






-
磷(phosphorus,P)是植物中脂质、核酸、二磷酸腺苷(ADP)、三磷酸腺苷(ATP)和糖类的重要组成元素,是含量仅次于氮的第二大限制性养分[1],在植物生长发育过程中发挥着不可替代的作用。植物吸收磷素主要以可溶性的磷酸根离子(
${\rm{PO}}_4^{3 - } $ 、${\rm{HPO}}_4^{2 - } $ 、${{\rm{H}}_2}{\rm{PO}}_4^ - $ )形式为主[2],虽然土壤中磷素含量很高,但是可供利用的无机磷素含量较少,是导致植物缺磷的主要原因之一[3]。植物磷转运蛋白(phosphate transporter,PHT)是植物从土壤中吸收无机磷酸盐并在体内进行再分配利用的载体。磷吸收动力学研究表明:植物在进化过程中形成了2类不同的磷吸收转运系统[4],分别为高亲和力磷吸收转运系统(high-affinity P uptake system)和低亲和力磷吸收转运系统(low-affinity P uptake system)[5]。研究者最早在拟南芥Arabidopsis thaliana中克隆出了磷转运蛋白基因AtPHT1.1,随后通过基因组测序、同源序列分析等方法陆续在大豆Glycine max[6]、玉米Zea mays[7]、大麦Triticum aestivum[8]和番茄Lycopersicon esculentum[9]等植物中克隆出了磷转运蛋白基因。植物磷酸盐转运蛋白属于MFS超级家族(major facilitator superfamily)[10],有12个疏水的跨膜区域,蛋白质结构高度相似,在氨基酸序列中存在保守特征序列GGDYPLSATIMSE,以及保守的磷酸化位点和糖基化位点。磷转运蛋白分为4个亚家族,分别为PHTⅠ、PHTⅡ、PHTⅢ、PHTⅣ[11]。拟南芥中,AtPHT1.1、AtPHT1.2和AtPHT1.3对拟南芥吸收磷的贡献非常大,而且AtPHT1.1在长距离运转磷的过程中具有重要作用[12]。研究表明:高磷环境中OsPHT1是水稻Oryza sativa吸收和转运磷素的关键PHTⅠ成员[13]。缺磷显著诱导OsPHT2、OsPHT4、OsPHT8、OsPHT9 和OsPHT10等的表达。OsPHT8几乎在水稻的各个器官中都强烈表达,是组成型磷转运子;OsPHT8下调会使新叶磷含量下降,老叶磷含量上升,处于灌浆期的胚乳和胚中的磷含量显著下降,说明水稻OsPHT8对磷素从源到库的再分配起到至关重要的作用[14]。大豆中,GmPHT7在根际成熟丛枝菌Arbuscular mycorrhiza (AM)根的根冠小柱细胞、皮层细胞和无菌根的侧根原基细胞中表达,在衰老叶片的维管束末端少数管胞中也有表达,主要负责向种子转运再活化的磷素[15]。此外,GmPHT10和GmPHT11也会受AM诱导表达[16],即磷酸盐转运蛋白基因广泛存在于各种植物中。毛竹Phyllostachys edulis属于禾本科Gramineae刚竹属Phyllostachys,具有经济价值高、用途广泛、栽培面积大等特点[17]。影响毛竹生长发育的因素有很多,如林地养分、水分不足[18],粗放的抚育管理和病虫害防治不及时等[19],其中,磷元素是限制毛竹林生长的重要营养元素之一。因此,研究毛竹磷素转运和吸收的相关基因的表达具有重要意义[20]。由于土壤对磷素的化学固定作用,磷素利用率普遍较低[21]。磷酸盐转运蛋白是植物吸收和转运磷素的重要参与者。本研究利用生物信息学方法鉴定毛竹PHTⅠ家族成员,分析其基因启动子、蛋白质理化性质、基因结构、氨基酸保守基序、基因在染色体的位置以及基因的组织表达特异性等,以期为深入研究毛竹PHT基因功能,探索毛竹磷素利用机制提供参考。
-
拟南芥和水稻PHT基因的CDS序列、基因序列以及氨基酸序列分别下载于Tair 和Rice Genome Annotation Project 。在毛竹基因组数据库Bamboo GDB 进行BlastP、BlastN (e-value=1e−10) 比对,获取毛竹PHTⅠ同源基因的候选序列。通过SMART数据库(
http://smart.embl-heidelberg.de )和PFAM数据库(http://pfam.xfam.org )确定获取的候选序列的保守结构域的准确性和完整性,保留具有编码完整保守结构域的候选序列,并进行基因命名(PePHTs)。 -
利用ProtParam 获取毛竹PHTⅠ基因编码蛋白质的基本理化特性,使用Gene Structure Display Server 2.0 在线工具分析PePHTs的基因结构,通过MEME Version 5.3.2 (
http://meme-suite.org/tools/meme )获取毛竹PHTⅠ的保守基序,利用PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )在线分析平台对PePHTs所含作用元件进行分析,在Excel软件中整理Tab结果文件, 用TBtools软件的Basic BioSequence View工具展示顺式作用元件的分布[22],使用Plant-mPLoc算法(http://www.csbio.sjtu.edu.cn )预测毛竹PHTⅠ的亚细胞位置。 -
使用Clustal X 1.8软件对毛竹、拟南芥、水稻等PHT基因的CDS序列进行多重比对,比对结果在MEGA 7.0软件中使用邻接法(neighbor-joining)构建系统进化树,bootstrap试验重复1000次[23],其他参数设置为默认值。利用在线工具Evolview 进行树图编辑[24]。
-
利用在线软件MEME Version 5.3.2 (
http://meme-suite.org/tools/meme )对毛竹PHTⅠ的motif进行预测和分析,motif数量设置为10。使用TBtools软件的Gene Location Visualize from GTF/GFF工具展示基因在染色体上的位置。使用KaKs_Calculator软件计算毛竹PHTⅠ基因的CDS序列的选择压力值(ω)[25]。 -
根据毛竹和竹笋不同组织的转录组数据[26],用PePHTs的RPKM (reads per kilo-bases per million reads)值表示基因的表达丰度,利用TBtools制作热图展示基因的表达丰度。
-
通过比对分析毛竹基因组,鉴定并确定编码完整MFs_1的候选基因共20个。植物PHT一般都含有MFs_1保守结构域,候选基因编码的蛋白质都含有保守的跨膜结构域,与MFS超家族的PHT家族特征相同。根据PHT候选基因在毛竹Scaffold中的位置以及同源基因的名称依次命名为:PePHT1~PePHT20。
对20个PHTⅠ家族基因进行生物信息学分析,其理化性质结果(表1)显示:PePHTs编码的氨基酸序列长度,最长为696 个氨基酸 (PePHT1),最短为432 个氨基酸(PePHT17),理论等电点为6.84~9.30,除PePHT15外,其他均为碱性蛋白质,分子量为48.61~76.37 kDa。疏水性测试显示:所有蛋白质疏水性值(grand average hydropathicity,GRAVY)>0,说明该家族蛋白质均为疏水性蛋白质。脂肪族氨基酸指数显示:PHTⅠ家族的蛋白质热稳定性为 83.79~105.07,热稳定性差异较大。亚细胞定位结果显示:PePHTs均定位于细胞膜中。
表 1 毛竹PHTⅠ家族基因编码蛋白序列的理化性质
Table 1. Physicochemical properties of proteins encoded by PHTⅠ gene family in Ph. edulis
基因名称 基因登录号 氨基酸/个 理论等电点 分子量/kDa 疏水性值 脂肪族氨基酸指数 亚细胞定位 PePHT1 PH02Gene03602 696 7.63 76.37 0.269 92.56 细胞膜 PePHT2 PH02Gene11006 531 9.10 57.69 0.538 98.91 细胞膜 PePHT3 PH02Gene14509 541 9.15 59.64 0.322 95.10 细胞膜 PePHT4 PH02Gene21291 532 8.45 58.05 0.371 89.40 细胞膜 PePHT5 PH02Gene24702 536 8.71 59.02 0.321 90.80 细胞膜 PePHT6 PH02Gene37931 547 8.31 58.97 0.412 92.32 细胞膜 PePHT7 PH02Gene39948 558 8.85 60.96 0.313 93.58 细胞膜 PePHT8 PH02Gene44007 536 8.03 58.49 0.408 91.27 细胞膜 PePHT9 PH02Gene44009 574 8.60 62.35 0.449 94.79 细胞膜 PePHT10 PH02Gene48053 507 8.01 55.77 0.259 87.59 细胞膜 PePHT11 PH02Gene48969 598 9.21 59.84 0.204 83.79 细胞膜 PePHT12 PH02Gene49859 567 8.99 61.45 0.376 92.31 细胞膜 PePHT13 PH02Gene50239 554 8.95 60.55 0.329 93.92 细胞膜 PePHT14 PH02Gene21248 518 8.02 57.42 0.494 105.04 细胞膜 PePHT15 PH02Gene21249 655 6.84 71.89 0.226 90.87 细胞膜 PePHT16 PH02Gene21250 505 9.30 55.46 0.473 105.07 细胞膜 PePHT17 PH02Gene21252 432 9.24 48.61 0.348 101.83 细胞膜 PePHT18 PH02Gene47590 520 8.67 58.31 0.324 90.25 细胞膜 PePHT19 PH02Gene47591 563 8.92 62.63 0.314 94.81 细胞膜 PePHT20 PH02Gene49564 527 8.04 58.50 0.391 97.53 细胞膜 -
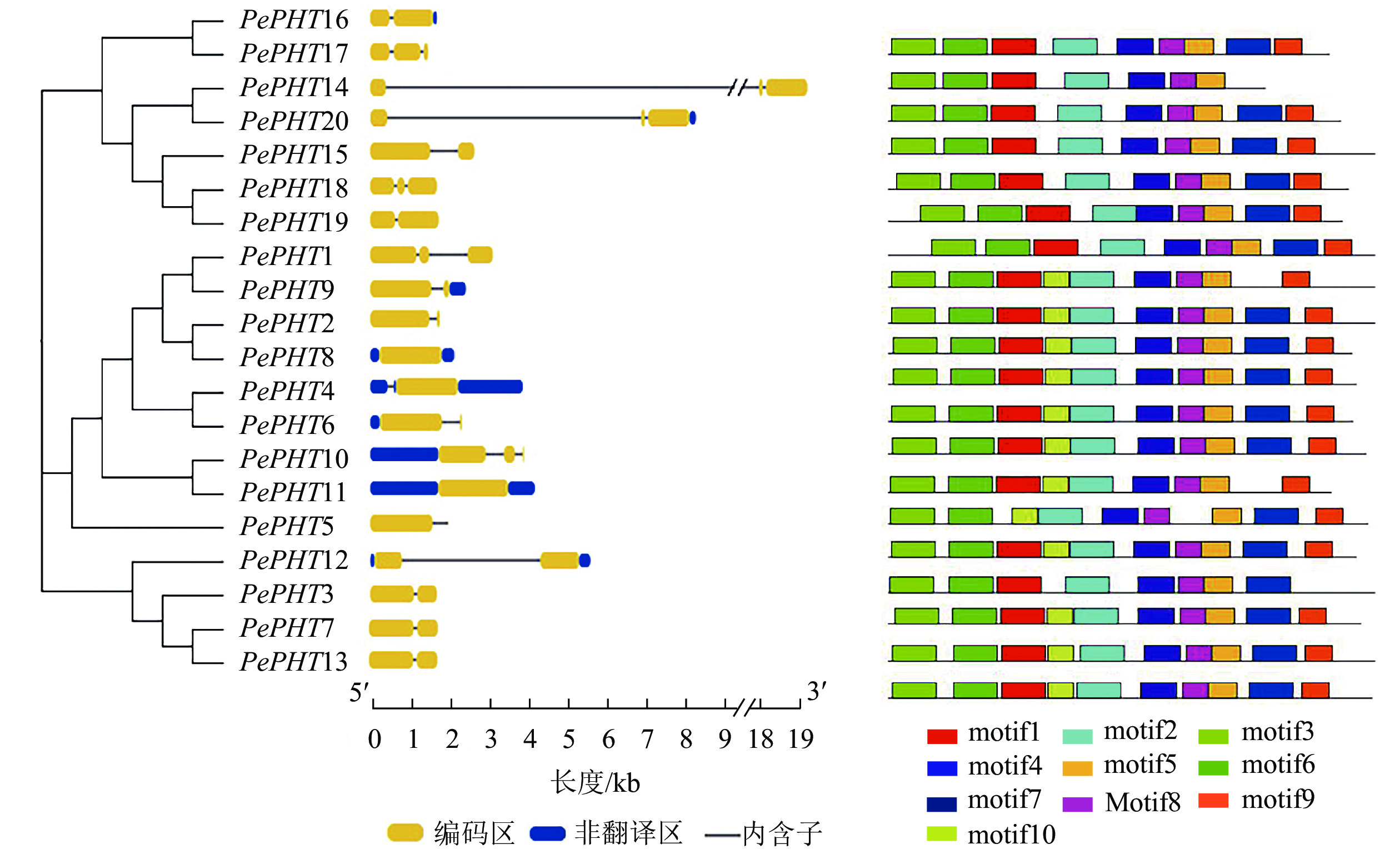
由图1可知:毛竹PHTⅠ家族基因多数含1~2个内含子(intron),在所有PePHTs中,PePHT14内含子区域最长,PePHT8和PePHT11内含子区域最短。保守基序分析显示:PePHTs含有7~10个保守基序,分别命名为motif1~motif10。其中有6个基序高度保守,分别是motif2、motif3、motif4、motif5、motif6、motif8,其他基序在部分序列中缺失。9个PePHTs中含有10个motif,其他11个PePHTs缺失1~3个motif,PePHT1、PePHT10都缺少motif7,PePHT11缺少motif1,PePHT12缺少motif9、motif10,PePHT14、PePHT15、PePHT16、PePHT18、PePHT19、PePHT20都缺少motif10,PePHT17则缺少motif7、motif9、motif10。PePHTs高度保守基序的氨基酸数目也不尽相同,最长的motif由50个氨基酸组成,最短的motif由29个氨基酸组成。
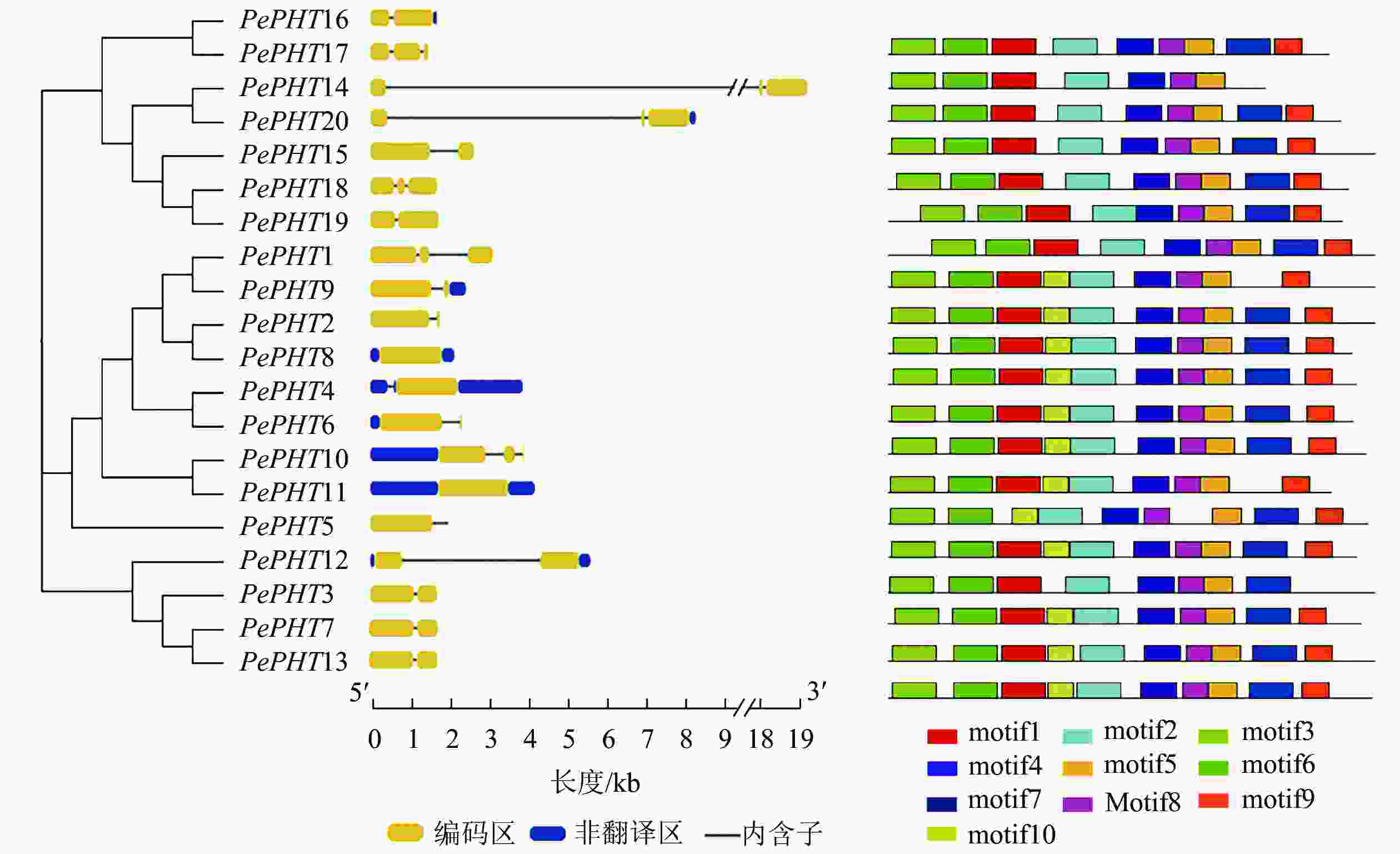
图 1 毛竹PHTⅠ家族基因的结构
Figure 1. Structures analysis of PHTⅠ gene family in Ph. edulis
为探究PePHTs受内在调控因子调节的情况和对外界环境的响应,选取PePHTs距离起始密码子上游2 000 bp的序列,对其所含顺式作用元件和应答元件进行分析。如图2所示:所有启动子的顺式作用元件种类比较相似,包括MYB转录因子结合的顺式作用元件,生长素、赤霉素、水杨酸、脱落酸等激素响应元件以及低温、干旱、缺氧、光等非生物胁迫响应元件。由此表明:PePHTs的转录表达可能会受到非生物胁迫和激素的影响。
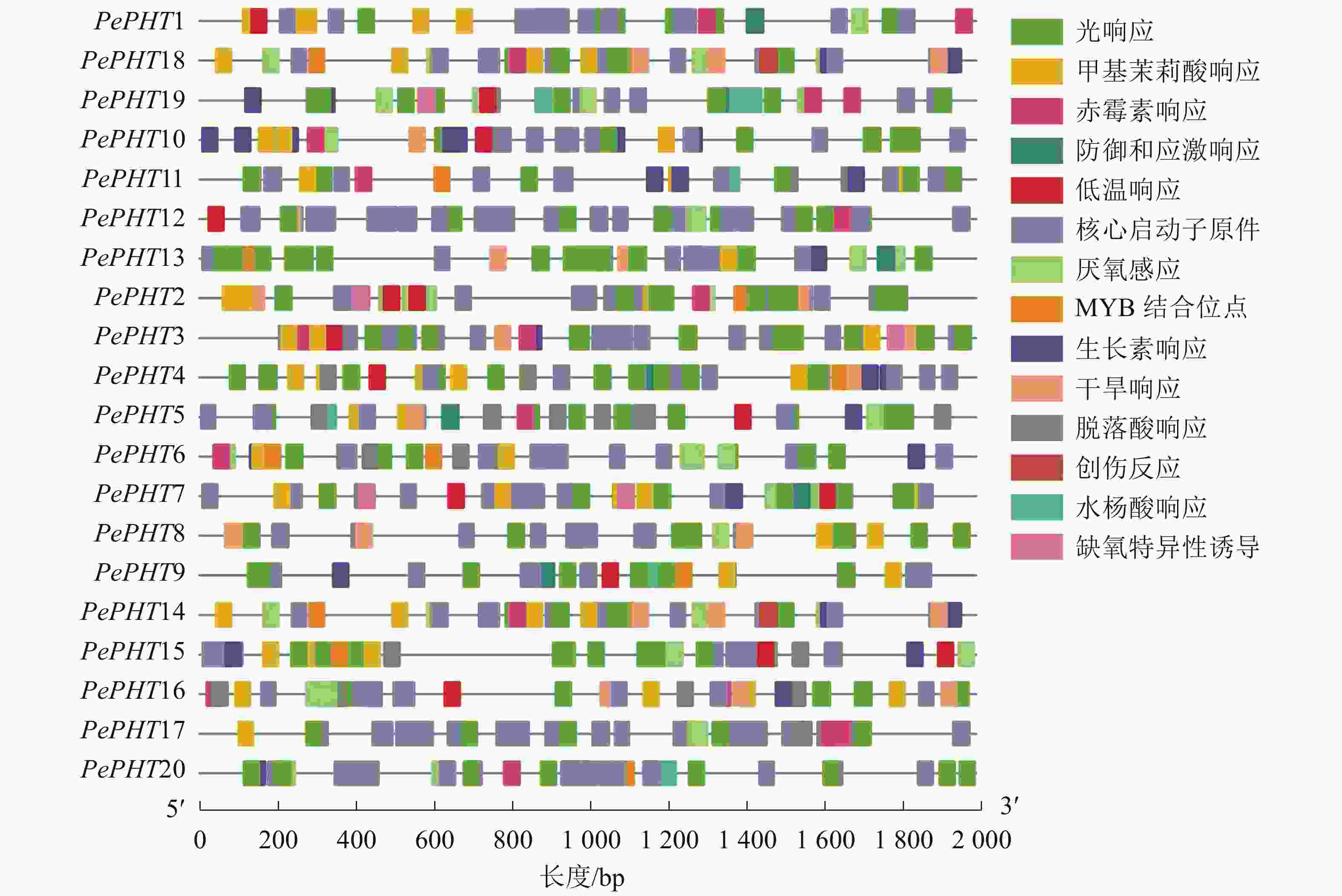
图 2 毛竹PHTⅠ家族基因启动子区顺式作用元件位置信息
Figure 2. Location information of cis-acting regulatory elements identified in the promoter region of PHTⅠ gene family in Ph. edulis
-
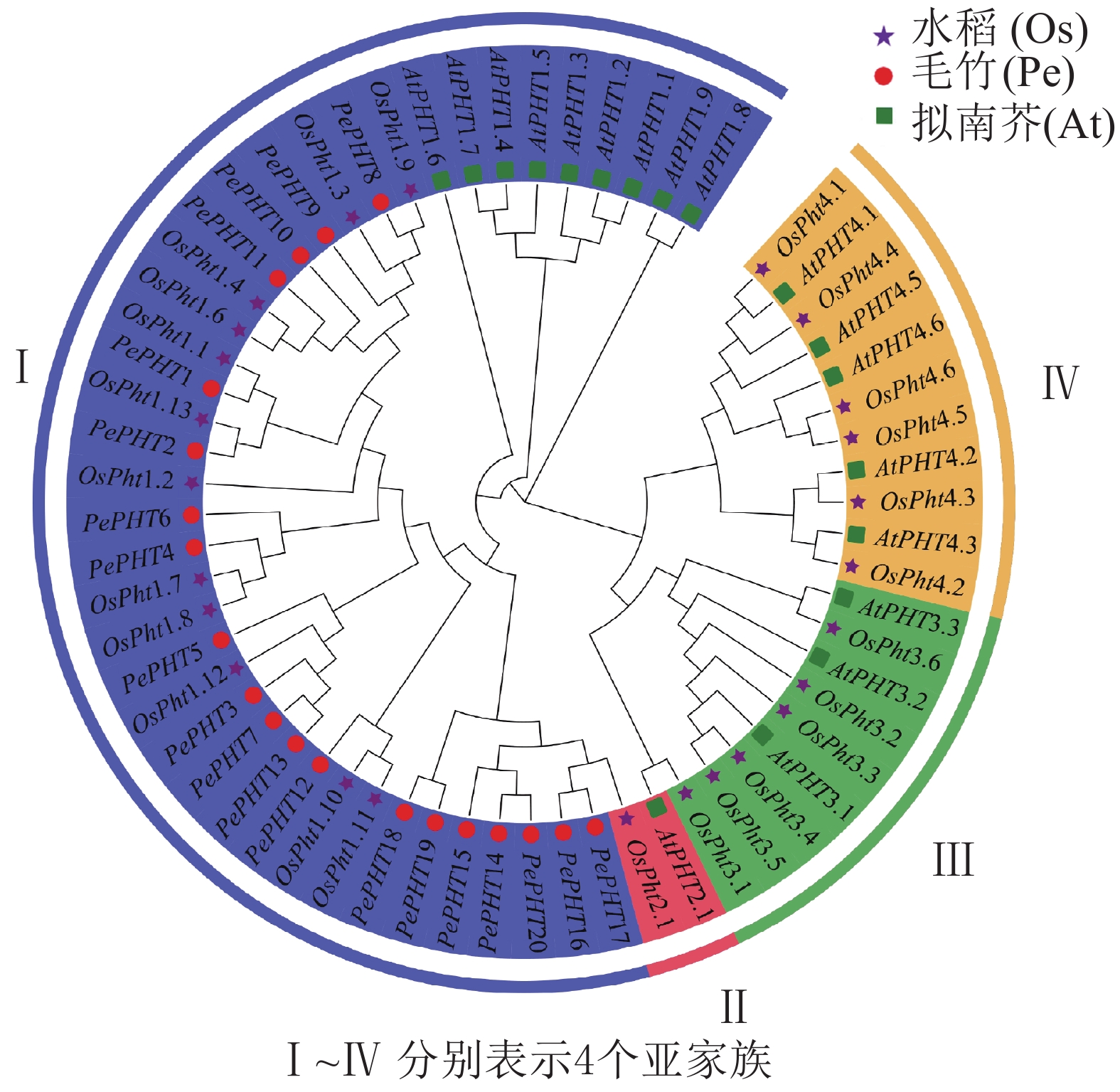
为了解PePHTs的进化关系,预测基因潜在功能,本研究提取20个毛竹、18个拟南芥和25个水稻的PHT基因的CDS序列进行多重序列比对,并利用MEGA 7.0软件根据邻接法构建系统进化树。由图3显示:毛竹、拟南芥和水稻的PHT基因被聚类到4个亚家族中,来自毛竹的20个PePHTs均分布在第Ⅰ亚家族的5个分支上,1个分支中只有PePHTs基因,另4个分支和水稻聚类在一起。此外,第Ⅰ亚家族中还包含9个拟南芥和12个水稻的PHT基因。PHT亚家族中的基因在功能上存在一定差异,如大多数第Ⅰ亚家族成员主要在直接接触根际环境的根毛和表皮细胞中表达,参与根系对环境中磷元素的吸收过程[27-28],定位在细胞膜上。相比拟南芥,毛竹PHT基因均优先与水稻PHT基因聚类,推测PePHTs在功能上可能与水稻同源基因更为相似。
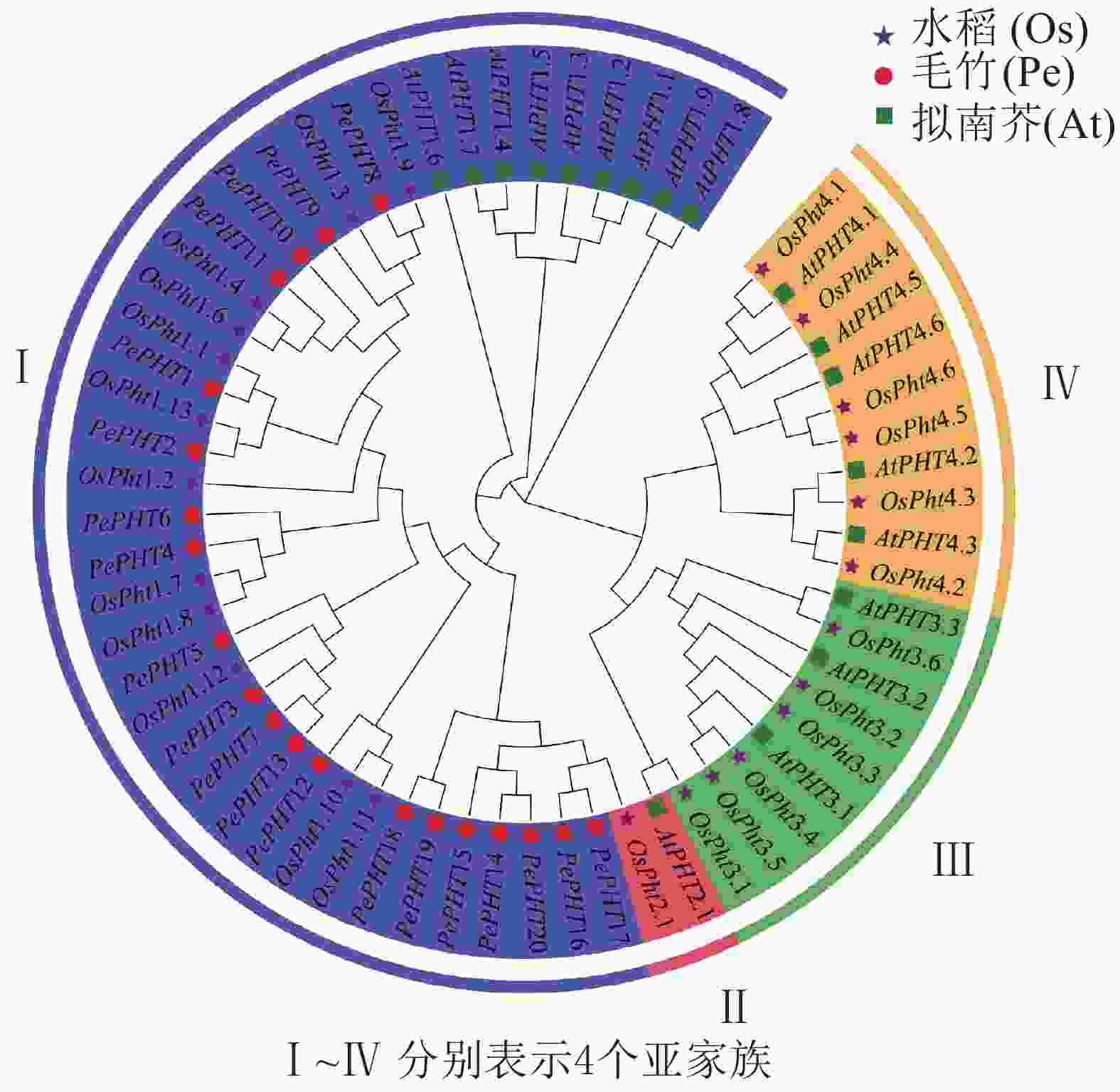
图 3 毛竹、拟南芥和水稻PHT家族基因系统进化树
Figure 3. Phylogentic tree of PHT gene family from Ph. edulis, A. thaliana and O. sativa
-
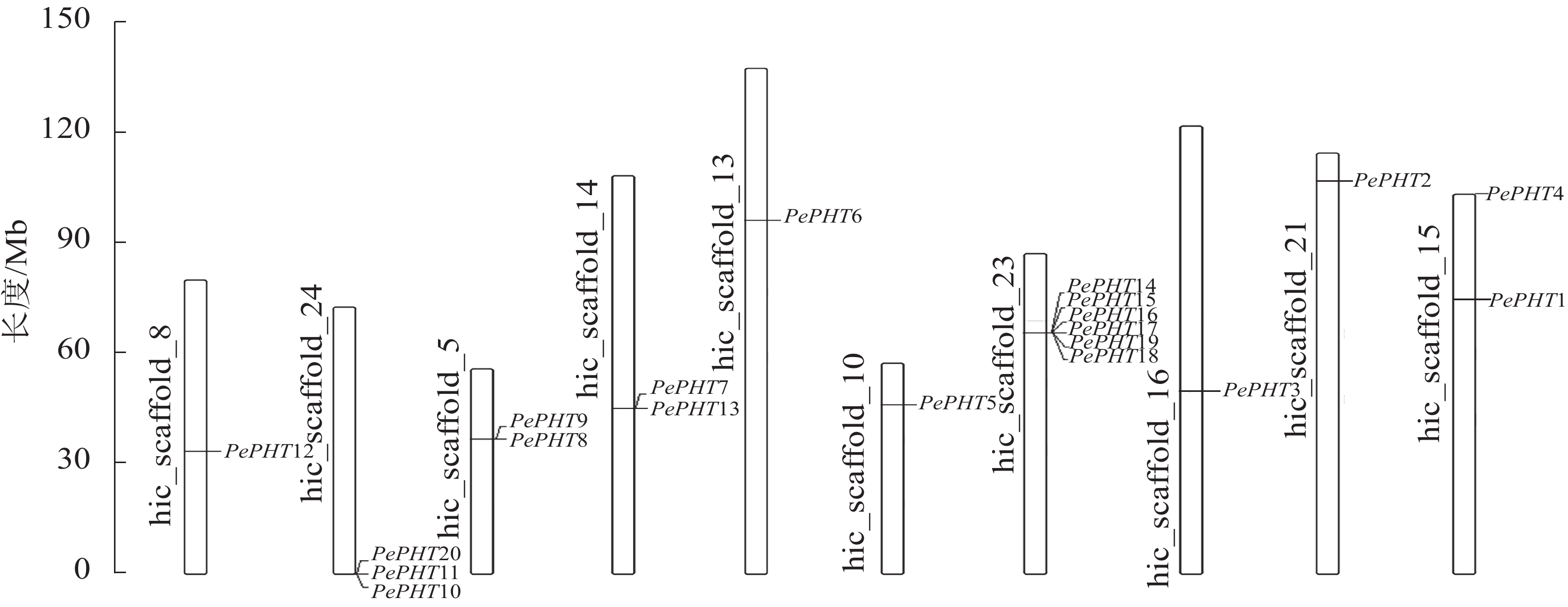
染色体定位显示:20个毛竹PHT基因位于10条染色体上,其中23号染色体上最多,有6个,分别是PePHT14、PePHT15、PePHT16、PePHT17、PePHT18、PePHT19;其次是24号染色体,有3个,分别是PePHT10、PePHT11、PePHT20,推测这2个基因簇中的基因可能分别编码催化2种新陈代谢途径中不同步骤的磷转运酶[29]。5号、14号以及15号染色体上各有2个基因,其余染色体各有1个(图4)。除数量分布不均匀外,各基因在染色体上的分布位置也不均匀,大多数基因位于染色体的中部,少量则位于顶部和底部。适应性分析结果表明:PePHT7的ω>0,其他基因的ω<0,表明PePHT7受到正选择压力,其他基因受到负选择压力[30]。
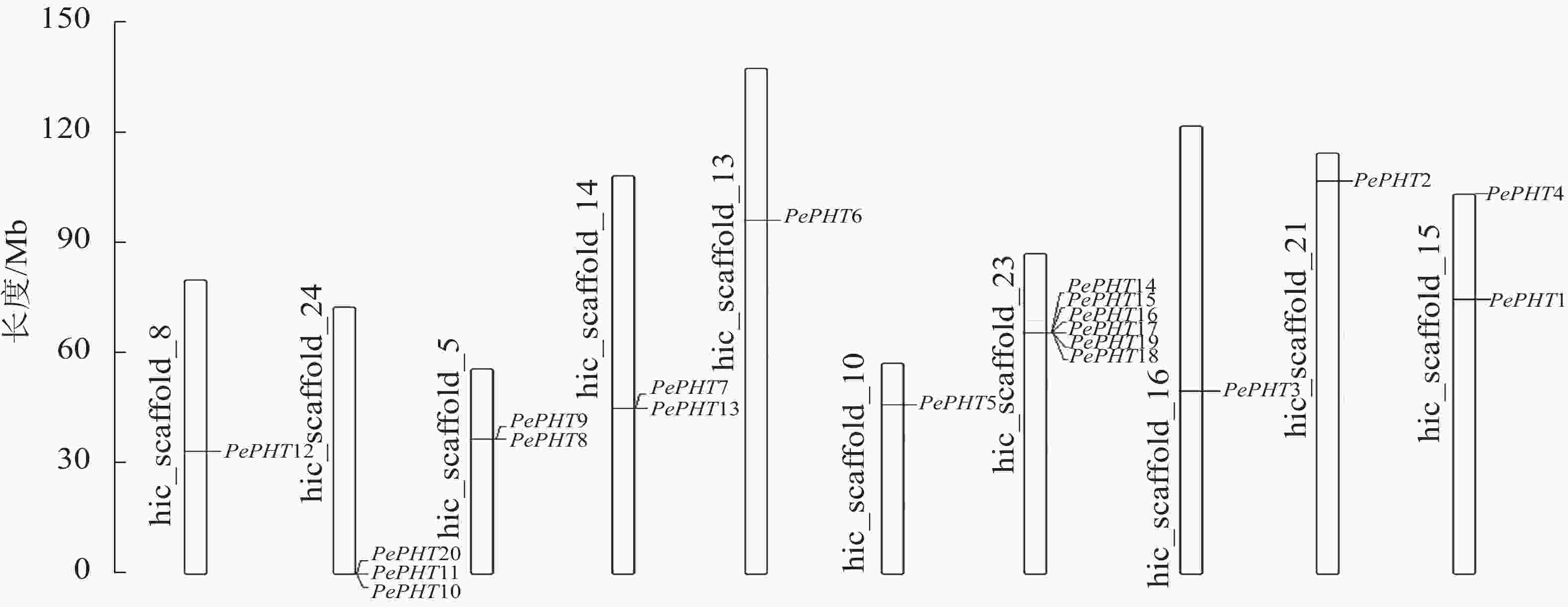
图 4 毛竹PHTⅠ基因在染色体上的位置
Figure 4. Chromosomal location of PHTⅠ genes from Ph. edulis
-
根据毛竹6个不同部位的转录组表达谱数据[26],对毛竹PHTⅠ家族基因进行组织特异性表达分析。由图5可知:毛竹不同组织中的PePHTs表达丰度差异较大,其中PePHT5、PePHT7、PePHT8、PePHT12、PePHT14、PePHT19在箨鞘中大量表达;PePHT2、PePHT3、PePHT6、PePHT13、PePHT15在根中表达丰度较高;PePHT18在叶中大量表达,PePHT10、PePHT20在叶鞘中大量表达,PePHT16、PePHT17在叶和叶鞘中也有少量表达。
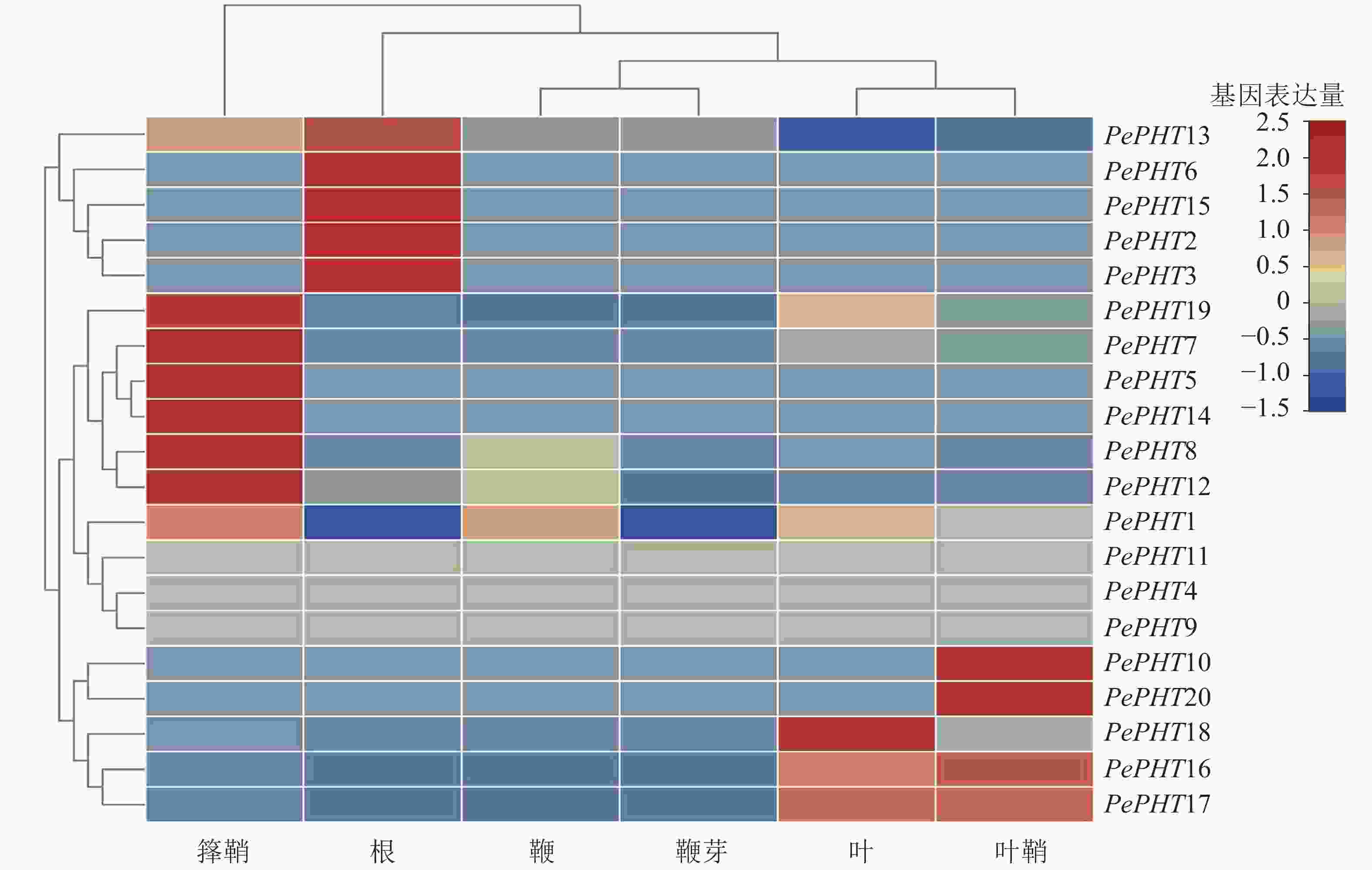
图 5 PePHTs在毛竹6个部位的表达
Figure 5. Expression analysis of PePHTs in 6 parts of Ph. edulis
-
植物从环境中吸收磷元素的过程中,磷酸盐转运蛋白起着至关重要的载体作用。植物PHTⅠ磷酸盐转运蛋白为膜蛋白,大多数属于高亲和力转运系统,并且具有相似的蛋白质序列和化学结构。在双子叶植物拟南芥和单子叶植物水稻中分别发现了9个[31-32]和12个PHTⅠ家族成员[6]。VERSAW等[33]证实拟南芥AtPHT2.1除了参与植物对磷元素的吸收与转运,还可能参与茎部对磷的转运,其蛋白质定位于叶绿体内膜上。PHTⅡ磷酸盐转运蛋白也在拟南芥[34]、马铃薯Solanum tuberosum [35]、茄Solanum melongena [36]、菠菜Spinacia oleracea [37]、烟草Nicotiana tabacum [38]和小麦Triticum aestivum [39]等多种植物中被发现。PHTⅢ磷酸盐转运蛋白最初在拟南芥中被克隆得到的。研究表明:PHTⅢ 磷酸盐转运家族与 PHTⅠ家族一样,通过 P/H+同向转运和P/OH−反向转运方式参与细胞质间磷的交换[39-40]。随后水稻、玉米和大豆等其他植物也克隆得到了该蛋白[41]。但是PHTⅣ磷酸盐转运蛋白只在少数几种植物中发现[42],研究报道很少。
植物磷转运蛋白是众多转运蛋白中的一类重要蛋白家族,它们在植物的根、茎、叶、花等器官都有分布,是磷元素吸收和运转的主要载体[1]。物种中均存在PHT基因,说明该基因具有重复性和多样性,这也是基因组重新排列和扩展的结果。本研究从毛竹基因组中鉴定出20个PHTⅠ家族基因,每个成员都含有Sugar_tr和MFs_1保守结构域,这是它们具有相似功能的基础。理化性质分析显示:PHT长度、理论等电点以及分子量区间跨度较大,这有可能是因为基因进行多次复制转录后进化的结果。本研究预测了毛竹PHTⅠ基因在细胞中的位置,发现毛竹PHTⅠ基因都分布在细胞膜上,与已有研究一致[3],说明鉴定出的毛竹PHTⅠ亚家族中的基因符合磷酸盐转运蛋白基因的特性。在植物进化过程中,选择压力能很好地体现发挥重要功能的蛋白的变化。基因适应性进化分析显示:大多数PePHTs基因受到较强烈的负选择压力,说明毛竹磷转运蛋白相对趋于稳定,是其保持原有重要功能的原因;同时PePHT7基因受到正选择压力,提示该基因编码的蛋白可能会延伸出一些新的功能[30]。植物基因组织特异性表达与基因的功能关系密切,毛竹磷转运蛋白在箨鞘、根、叶和叶鞘都有1个及以上的基因大量表达,PePHT4、PePHT9、PePHT11等3个基因在任何组织中都没有检测到,还需深入研究;PePHT1在根和鞭芽中明显下调,说明基因在不同组织中表达丰度不一样且发挥着不同的作用。
系统进化分析发现:毛竹PHTⅠ基因聚类在第Ⅰ亚家族的5个分支上,同一支中的基因可能具有相似的功能。拟南芥的PHTⅠ亚家族中有9个成员,其中AtPHT1.6在花粉中表达,其余8个在根中表达[43];当拟南芥缺磷时,AtPHT1.1和AtPHT1.4会大量表达,这2个磷酸盐转运蛋白提供了70%的磷酸盐转运活性[44]。水稻PHTⅠ亚家族中有12个成员[6],其中OsPHT1主要功能是吸收磷素和体内磷酸盐的再分配[13],OsPHT2主要在地上部分表达,也是水稻PHTⅠ亚家族中唯一一个低亲和力磷转运蛋白[45]。PHTⅠ家族在水稻和拟南芥吸收和转运磷的过程中具有重要作用[32, 45],推测在毛竹对磷的吸收转运过程中也可能具有重要作用,但需要进一步的转录组数据进行验证。
Identification and expression pattern of phosphorus transporter Ⅰ family genes of Phyllostachys edulis
-
摘要:
目的 鉴定毛竹Phyllostachys edulis磷转运蛋白Ⅰ (phosphate transporter 1, PHTⅠ)家族基因,分析其表达模式。 方法 利用生物信息学方法,鉴定毛竹PHTⅠ家族成员,分析基因启动子调控元件、编码蛋白的理化性质、基因结构、氨基酸保守基序、基因在染色体的位置、组织表达特异性、基因适应性进化及系统进化等。 结果 毛竹中共鉴定出20个PHTⅠ家族基因 (PePHTs),分布在10条染色体上,均定位于细胞膜。每个基因都含有1~2个内含子,PePHTs启动子序列中包含干旱、低温等非生物胁迫以及赤霉素等激素类响应元件。毛竹PHTⅠ大部分为碱性蛋白质,分子量为48.61~76.37 kDa,理论等电点为6.84~9.30,疏水性值均大于0,都属于疏水蛋白。基因适应性进化分析显示:大多数PePHTs的选择压力值小于0,说明多数基因受到负选择压力。转录组表达图谱表明:PePHTs在不同组织中的表达存在差异性,说明该基因家族在毛竹生长发育过程中发挥着不同的作用。系统进化树表明:PePHTs都聚类在第Ⅰ亚家族,并且优先和水稻Oryza sativa聚类在同一支上。 结论 PHTⅠ家族在植物吸收和转运磷的过程中扮演了重要作用。本研究结果为深入研究毛竹PHTⅠ家族基因的功能奠定了理论基础。图5表1参45 Abstract:Objective This study is aimed to investigate the family members of phosphorus transporter Ⅰ (PHTⅠ) protein of Moso bamboo (Phyllostachys edulis) and analyze their expression patterns. Method With the employment of the bioinformatics, the PHTⅠ family members of Moso bamboo were identified before an analysis was conducted of the regulatory elements of gene promoters, the physicochemical properties of coding proteins, gene structure, conserved motifs of amino acids, gene positions on chromosomes, tissue expression specificity, gene adaptive evolution and phylogeny. Result PHTⅠ family of Moso bamboo consisted of 20 members distributed on ten chromosomes in the cell membrane and each gene contained 1 to 2 introns while the PePHTs promoter sequence contained elements of abiotic stress such as drought and low temperature and hormone response elements such as Gibberellic acid. Most PHTⅠ from Ph.edulis were basic proteins with the range of molecular weight of PePHTs between 48.61 and 76.37 kDa and that of the theoretical isoelectric point between 6.84 and 9.30. The hydrophobicity value of all PePHTs was greater than zero, indicating that all the proteins in this family were hydrophobic. In terms of gene adaptive evolution, the ω value of most PePHTs genes were lower than zero, indicating that most genes were under negative selection pressure. PePHTs had different expressions in different tissues, which indicated that PePHTs played different roles in the growth and development of Moso bamboo. PePHTs were clustered in the first Ⅰ subfamily, preferentially in the same branch as rice (Oryza sativa). Conclusion PHTⅠ family plays an important role in the absorption and transport of phosphorus in plants and the results of this study have provided a favorable theoretical foundation for a further analysis of the functions of the PHTⅠ family genes in Moso bamboo. [Ch, 5 fig. 1 tab. 45 ref. ] -
图 2 毛竹PHTⅠ家族基因启动子区顺式作用元件位置信息
Figure 2 Location information of cis-acting regulatory elements identified in the promoter region of PHTⅠ gene family in Ph. edulis
图 3 毛竹、拟南芥和水稻PHT家族基因系统进化树
Figure 3 Phylogentic tree of PHT gene family from Ph. edulis, A. thaliana and O. sativa
表 1 毛竹PHTⅠ家族基因编码蛋白序列的理化性质
Table 1. Physicochemical properties of proteins encoded by PHTⅠ gene family in Ph. edulis
基因名称 基因登录号 氨基酸/个 理论等电点 分子量/kDa 疏水性值 脂肪族氨基酸指数 亚细胞定位 PePHT1 PH02Gene03602 696 7.63 76.37 0.269 92.56 细胞膜 PePHT2 PH02Gene11006 531 9.10 57.69 0.538 98.91 细胞膜 PePHT3 PH02Gene14509 541 9.15 59.64 0.322 95.10 细胞膜 PePHT4 PH02Gene21291 532 8.45 58.05 0.371 89.40 细胞膜 PePHT5 PH02Gene24702 536 8.71 59.02 0.321 90.80 细胞膜 PePHT6 PH02Gene37931 547 8.31 58.97 0.412 92.32 细胞膜 PePHT7 PH02Gene39948 558 8.85 60.96 0.313 93.58 细胞膜 PePHT8 PH02Gene44007 536 8.03 58.49 0.408 91.27 细胞膜 PePHT9 PH02Gene44009 574 8.60 62.35 0.449 94.79 细胞膜 PePHT10 PH02Gene48053 507 8.01 55.77 0.259 87.59 细胞膜 PePHT11 PH02Gene48969 598 9.21 59.84 0.204 83.79 细胞膜 PePHT12 PH02Gene49859 567 8.99 61.45 0.376 92.31 细胞膜 PePHT13 PH02Gene50239 554 8.95 60.55 0.329 93.92 细胞膜 PePHT14 PH02Gene21248 518 8.02 57.42 0.494 105.04 细胞膜 PePHT15 PH02Gene21249 655 6.84 71.89 0.226 90.87 细胞膜 PePHT16 PH02Gene21250 505 9.30 55.46 0.473 105.07 细胞膜 PePHT17 PH02Gene21252 432 9.24 48.61 0.348 101.83 细胞膜 PePHT18 PH02Gene47590 520 8.67 58.31 0.324 90.25 细胞膜 PePHT19 PH02Gene47591 563 8.92 62.63 0.314 94.81 细胞膜 PePHT20 PH02Gene49564 527 8.04 58.50 0.391 97.53 细胞膜  下载: 导出CSV
下载: 导出CSV
-
[1] ABEL S, TICCONI C A, DELATORRE C A. Phosphate sensing in higher plants [J]. Physiol Plant, 2002, 115(1): 1 − 8. [2] VANCE C P, UHDE-STONE C, ALLAN D L. Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource [J]. New Phytol, 2003, 157(3): 423 − 447. [3] SCHACHTMAN D P, REID R J, AYLING S M. Phosphorus uptake by plants: from soil to cell [J]. Plant Physiol, 1998, 116(2): 447 − 453. [4] POIRIER Y, BUCHER M. Phosphate transport and homeostasis in Arabidopsis [J/OL]. Am Soc Plant Biol, 2002: e0024 [2021-06-05]. doi: 10.1199/tab.0024. [5] LÓPEZ-ARREDONDO D L, LEYVA-GONZÁLEZ M A, GONZÁLEZ-MORALES S I, et al. Phosphate nutrition: improving low-phosphate tolerance in crops [J]. Annu Rev Plant Biol, 2014, 65: 95 − 123. [6] PASZKOWSKI U, KROKEN S, ROUX C, et al. Rice phosphate transporters include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal symbiosis [J]. Proc Nat Acad Sci, 2002, 99(20): 13324 − 13329. [7] NAGY R, VASCONCELOS M J V, ZHAO S, et al. Differential regulation of five Pht1 phosphate transporters from maize (Zea mays L. ) [J]. Plant Biol, 2006, 8(2): 186 − 197. [8] RAE A L, CYBINSKI D H, JARMEY J M, et al. Characterization of two phosphate transporters from barley; evidence for diverse function and kinetic properties among members of the Pht1 family [J]. Plant Mol Biol, 2003, 53(1/2): 27 − 36. [9] NAGY R, KARANDASHOV V, CHAGUE V, et al. The characterization of novel mycorrhiza-specific phosphate transporters from Lycopersicon esculentum and Solanum tuberosum uncovers functional redundancy in symbiotic phosphate transport in solanaceous species [J]. Plant J, 2005, 42(2): 236 − 250. [10] PAO S S, PAULSEN I T, SAIER M H. Major facilitator superfamily [J]. Microbiol Mol Biol Rev, 1998, 62(1): 1 − 34. [11] WANG Duoli, LÜ Sulian, JIANG Ping, et al. Roles, regulation, and agricultural application of plant phosphate transporters [J/OL]. Front Plant Sci, 2017, 8: 817[2021-06-05]. doi: 10.3389/fpls.201700817. [12] AYADI A, DAVID P, ARRIGHI J F, et al. Reducing the genetic redundancy of Arabidopsis PHOSPHATE TRANSPORTER1 transporters to study phosphate uptake and signaling [J]. Plant Physiol, 2015, 167(4): 1511 − 1526. [13] SUN Shubin, GU Mian, CAO Yue, et al. A constitutive expressed phosphate transporter, OsPht1;1, modulates phosphate uptake and translocation in phosphate-replete rice [J]. Plant Physiol, 2012, 159(4): 1571 − 1581. [14] LI Yiting, ZHANG Jun, ZHANG Xiao, et al. Phosphate transporter OsPht1;8 in rice plays an important role in phosphorus redistribution from source to sink organs and allocation between embryo and endosperm of seeds [J]. Plant Sci, 2015, 230: 23 − 32. [15] INOUE Y, KOBAE Y, OMOTO E, et al. The soybean mycorrhiza-inducible phosphate transporter gene, GmPT7, also shows localized expression at the tips of vein endings of senescent leaves [J]. Plant Cell Physiol, 2014, 55(12): 2102 − 2111. [16] GORDON-WEEKS R, TONG Yiping, DAVIES T G E, et al. Restricted spatial expression of a high-affinity phosphate transporter in potato roots [J]. J Cell Sci, 2003, 116(15): 3135 − 3144. [17] SONG Xinzhang, PENG Changhui, CIAIS P, et al. Nitrogen addition increased CO2 uptake more than non-CO2 greenhouse gases emissions in a Moso bamboo forest [J/OL]. Sci Adv, 2020, 6(12): eaaw5790[2021-06-20]. doi: 10.1126/sciadv.aaw5790. [18] 郑德华. 不同土壤类型对毛竹林生长的影响研究[J]. 世界竹藤通讯, 2014, 12(5): 32 − 34. ZHENG Dehua. Influence of soil type on the growth of Phyllostachys heterocycla cv. pubescens forest [J]. World Bamboo Rattan, 2014, 12(5): 32 − 34. [19] 严圣钦. 低产毛竹林的成因和改造技术[J]. 世界竹藤通讯, 2009, 7(3): 34 − 36. YAN Shengqin. Causes for low-yield moso forest and the improvement technology [J]. World Bamboo Rattan, 2009, 7(3): 34 − 36. [20] DU Enzai, TERRER C, PELLEGRINI A F A, et al. Global patterns of terrestrial nitrogen and phosphorus limitation [J]. Nat Geosci, 2020, 13(3): 221 − 226. [21] 赵光强, 付循成, 曹慧. 高等植物的磷营养研究[J]. 安徽农业科学, 2007, 35(31): 9851 − 9854. ZHAO Guangqiang, FU Xuncheng, CAO Hui. Research on the phosphorus nutrition in higher plant [J]. J Anhui Agric Sci, 2007, 35(31): 9851 − 9854. [22] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Mol Plant, 2020, 13(8): 1194 − 1202. [23] KUMAR S, STECHER G, TAMURA K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets [J]. Mol Biol Evol, 2016, 33(7): 1870 − 1874. [24] HE Zilong, ZHANG Huangkai, GAO Shenghan, et al. Evolview V2: an online visualization and management tool for customized and annotated phylogenetic trees [J]. Nucleic Acids Res, 2016, 44(W1): W236 − W241. [25] IVANOVA Z, SABLOK G, DASKALOVA E, et al. Chloroplast genome analysis of resurrection tertiary relict haberlea rhodopensis highlights genes important for desiccation stress response [J/OL]. Front Plant Sci, 2017, 8: 204[2021-06-21]. doi: 10.3389/fpls.2017.00204. [26] ZHAO Hansheng, GAO Zimin, WANG Le, et al. Chromosome-level reference genome and alternative splicing atlas of Moso bamboo (Phyllostachys edulis) [J/OL]. GigaScience, 2018, 7(10): giy115[2021-06-23]. doi: 10.1093/gigascience/giy115. [27] BUN-YA M, NISHIMURA M, HARASHIMA S, et al. The PHO84 gene of saccharomyces cerevisiae encodes an inorganic phosphate transporter [J]. Mol Cell Biol, 1991, 11(6): 3229 − 3238. [28] AZIZ T, FINNEGAN P M, LAMBERS H, et al. Organ-specific phosphorus-allocation patterns and transcript profiles linked to phosphorus efficiency in two contrasting wheat genotypes [J]. Plant Cell Environ, 2014, 37(4): 943 − 960. [29] 林翩翩, 白有煌, 周明兵. 毛竹细胞色素P450的基因组学分析[J]. 植物生理学报, 2014, 50(9): 1387 − 1400. LIN Pianpian, BAI Youhuang, ZHOU Mingbing. Genomics analysis of cytochrome P450 monooxygenase genes in Phyllostachys heterocycla [J]. Plant Physiol J, 2014, 50(9): 1387 − 1400. [30] 王捷, 魏爱丽, 石瑛, 等. 念珠藻属植物hetR基因的适应性进化分析[J]. 植物科学学报, 2020, 38(1): 23 − 31. WANG Jie, WEI Aili, SHI Ying, et al. Adaptive evolutionary analysis of hetR gene in Nostoc [J]. Plant Sci J, 2020, 38(1): 23 − 31. [31] YE Ying, YUAN Jing, CHANG Xiaojian, et al. The phosphate transporter gene OsPht1;4 is involved in phosphate homeostasis in rice [J/OL]. PLoS One, 2015, 10(5): e0126186[2021-06-23]. doi: 10.1371/journal.pone.0126186. [32] MUDGE S R, RAE A L, DIATLOFF E, et al. Expression analysis suggests novel roles for members of the Pht1 family of phosphate transporters in Arabidopsis [J]. Plant J, 2002, 31(3): 341 − 353. [33] VERSAW W K, HARRISON M J. A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses [J]. Plant Cell, 2002, 14(8): 1751 − 1766. [34] KARTHIKEYAN A S, VARADARAJAN D K, JAIN A, et al. Phosphate starvation responses are mediated by sugar signaling in Arabidopsis [J]. Planta, 2007, 225(4): 907 − 918. [35] GOFF S A, RICKE D, LAN T H, et al. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica) [J]. Science, 2002, 296(5565): 92 − 100. [36] CHEN Aiqun, HU Jiang, SUN Shubin, et al. Conservation and divergence of both phosphate- and mycorrhiza-regulated physiological responses and expression patterns of phosphate transporters in solanaceous species [J]. New Phytol, 2007, 173(4): 817 − 831. [37] FERRO M, SALVI D, RIVIÈRE-ROLLAND H, et al. Integral membrane proteins of the chloroplast envelope: identification and subcellular localization of new transporters [J]. Proc Natl Acad Sci, 2002, 99(17): 11487 − 11492. [38] GUO Chengjin, ZHAO Xiaolei, LIU Xiaoman, et al. Function of wheat phosphate transporter gene TaPHT2;1 in Pi translocation and plant growth regulation under replete and limited Pi supply conditions [J]. Planta, 2013, 237(4): 1163 − 1178. [39] RAUSCH C, BUCHER M. Molecular mechanisms of phosphate transport in plants [J]. Planta, 2002, 216(1): 23 − 37. [40] SAIA S, RAPPA V, RUISI P, et al. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat [J/OL]. Front Plant Sci, 2015, 6: 815[2021-04-15]. doi: 10.3389/fpls.2015.00815. [41] WOHLRAB H. Identification of the N-ethylmaleimide reactive protein of the mitochondrial phosphate transporter [J]. Biochemistry, 1979, 18(10): 2098 − 2102. [42] GUO Biwei, IRIGOYEN S, FOWLER T B, et al. Differential expression and phylogenetic analysis suggest specialization of plastid-localized members of the PHT4 phosphate transporter family for photosynthetic and heterotrophic tissues [J]. Plant Signaling Behav, 2008, 3(10): 784 − 790. [43] WOHLRAB H, BRIGGS C. Yeast mitochondrial phosphate transport protein expressed in Escherichia coli. Site-directed mutations at threonine-43 and at a similar location in the second tandem repeat (isoleucine-141) [J]. Biochemistry, 1994, 33(32): 9371 − 9375. [44] MISSON J, RAGHOTHAMA K G, JAIN A, et al. A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation [J]. Proc Natl Acad Sci, 2005, 102(33): 11934 − 11939. [45] AI Penghui, SUN Suibin, ZHAO Jianning, et al. Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and kinetic properties in uptake and translocation [J]. Plant J, 2009, 57(5): 789 − 809. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210471
 点击查看大图
点击查看大图
计量
- 文章访问数: 2427
- HTML全文浏览量: 538
- PDF下载量: 70
- 被引次数: 0



