




-
随着全球气候变暖问题日益突出,通过森林增汇应对气候变化问题受到各国政府和学术界的普遍关注。竹林较其他森林具有更强的固碳能力[1]。20世纪80年代,零星出现单株竹子生长过程中生物量及碳储量变化模拟分析[2]的竹林碳汇研究;20世纪90年代,转向研究竹子种群生物量[3-4],竹子碳汇的研究开始由单株碳汇拓展至竹林碳汇;进入21世纪,学者们开始对竹林生态系统碳汇量展开探索[5-7],并就管理措施如何提升竹林碳汇能力展开讨论[8-10]。近年来,国家和地区碳交易体系逐渐完善,竹林碳汇项目开发及碳交易研究成为热点[11-12],研究开始从单纯的探索竹林碳汇变化转变为依托竹林碳汇获利。随着竹林比较收益下降和退化竹林增加,竹林应对气候变化和改善农户生计的双重效应探索[13-15]成为当前竹林碳汇研究的重点,竹林增汇及可持续经营建议陆续提出。目前竹林碳汇研究的综述以传统的定性描述为主[16-17],鲜有使用定量方法进行归纳和总结;自《京都议定书》签订以来,竹林碳汇在应对气候变化中作用受到极大重视,积累了相当丰富的成果。本研究基于信息计量学的思想,借助CiteSpace软件,对Web of Science核心合集数据库中1998−2019年以竹林碳汇为主题的文献数据进行可视化、系统的回顾和总结,以展示自《京都议定书》签订以来全球竹林碳汇研究领域的总体特征和演变趋势,为开展相关研究提供参考。
-
信息计量学由文献计量学演化而来,是研究文献作者分布、机构分布、关键词信息等文献信息的重要方法[18]。随着计算机科学的进步和文献数据库兼容性的不断改进,Web of Science、中文社会科学引文索引(CSSCI)、中国知网(CNKI)等数据库借助信息计量学方法对文献作者、发文机构、关键词等信息进行检索并分析。知识图谱是信息计量学里用于展现科学知识发展进程与结构关系最为直观的形式,是总结各学科的研究态势与研究进展较为科学的方式。本研究借助知识图谱,对竹林碳汇相关研究的文献信息、知识网络进行可视化展现。鉴于CiteSpace软件在信息计量学可视化分析方面的应用,及在文献信息中潜在的知识结构、变化规律挖掘方面的特色[19],本研究选用2020年初更新的CiteSpace 5.6版本作为分析工具。
-
核心作者群指发文数量较多,影响力较大的作者群体,文献计量学中常用洛特卡定律(Lotka’s law)和普赖斯定律(Price’s law)分析核心作者群。洛特卡定律通过分析作者数量和发文数量的相关关系,揭示作者科研生产能力的频率分布特征,以反映该领域作者的科研生产能力,即发表n篇论文的作者数约为发表1篇论文的作者数的1/π2,发表1篇论文的作者数约为所有作者的60%[20],据此判断该领域核心作者群的发文水平。普赖斯定律以洛特卡定律为基础,利用大量文献统计进一步推导得出高产作者的数量[18],提供某领域核心作者的识别方法;即通过该领域最高产作者的发文数量推测核心作者的最低发文数量,以确定某作者是否为该领域的核心作者,核心作者的最低发文数量k=0.749
$ \sqrt {{n _{\max }}} $ ,其中nmax为该领域最高产的作者的发文数量,推导出某领域大约50%的论文是由核心作者所发表[21]。 -
Web of Science核心合集数据库是全球最大、覆盖学科最多的综合性学术信息资源,收录了全球最具影响力的近1万种核心学术期刊,可信度高,可以较为全面、准确的透视全球范围竹林碳汇相关研究进展情况。设置检索主题为竹林碳汇(包括bamboo forest carbon sink,bamboo forest carbon sequestration,bamboo forest carbon stock,bamboo forest carbon storage,bamboo forest carbon,bamboo carbon stock,bamboo carbon storage,bamboo carbon sink,bamboo carbon sequestration)。考虑到1997年《京都议定书》签订并提出“增加森林碳汇抵消减排”,竹林碳汇相关研究开始得以高度重视,而许多期刊2020年度文献不全面,设置搜索年限为1998−2019年。分析前逐条核对检索结果,对于信息不完整(无作者的文献、会议记录、访谈、总结、讨论、综述等)的内容予以剔除,最终得到文献1582篇,作为研究对象。
-
从图1可知:1998−2019年间,竹林碳汇研究领域发文量约75篇·a−1,2001年后发文量大幅度增加,2013年后超过100篇·a−1,且具有明显的逐年增长态势。说明《京都议定书》签订后,通过竹林增汇应对气候变化问题引起了学术界的普遍关注;相关主题经过3~5 a的酝酿,以文献的形式快速积累并逐年增多;随着进入该领域的机构和作者增多,竹林碳汇研究进一步蓬勃发展。1998−2019年间年度发文数量有所波动,但总体趋势逐年上升并有加速增长态势,目前处于快速发展期。表明竹林碳汇的研究正受到越来越多学者的广泛重视。
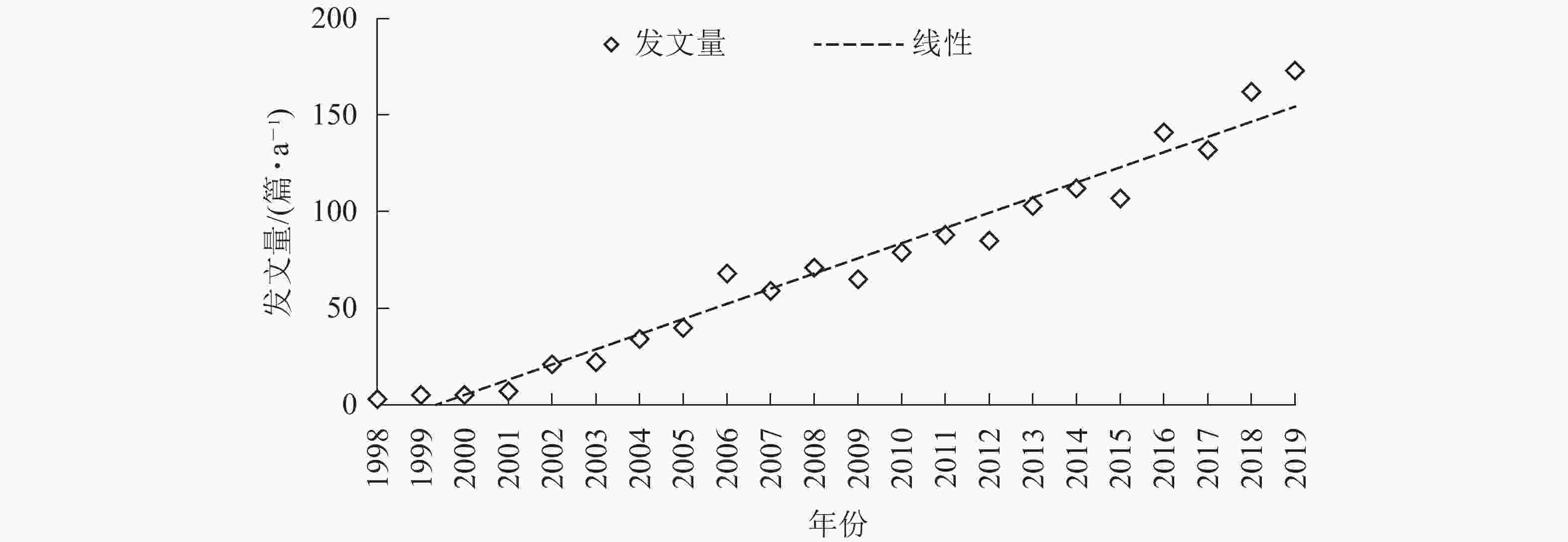
图 1 历年竹林碳汇文献数量变化
Figure 1. Changes in the number of studies on bamboo forest carbon sinks over the years
-
对发文作者的分析可知,“Guomo Zhou”(周国模)“Peikun Jiang”(姜培坤)“Huaqiang Du”(杜华强)在该领域的中心性地位较高;发文数量分别为56、26和23篇,远高于其他大多数学者。同时作者之间合作紧密,已形成了具有一定规模的学术团体。进一步调查发现:中心性地位较高的3位作者均来自浙江农林大学同一科研团队,全球首个竹林碳汇项目、全球竹林碳汇项目造林和经营的方法学[22]均由该团队开发和制定;标志着中国在竹林碳汇理论研究和实践方面均处于世界领先水平,以周国模为代表的一批中国学者正引领着全球竹林碳汇不断发展。
-
对竹林碳汇研究领域各个发文数量区段及对应作者数量的分析可知(表1):仅发表过1篇论文的作者数占比为54.53%,低于洛特卡定律的60%;说明在竹林碳汇领域,发文数量超过1篇的作者较其他学科的普遍水平更高;另外,发表2~6篇和26~56篇论文的作者数量与发表1篇论文的作者数量的比值,也高于洛特卡定律,而发表8~23篇的作者占比则低于洛特卡定律。表明竹林碳汇领域的作者研究较为专注且科研生产率高,尤其发文为8~23篇的作者群对该领域的贡献相比于其它学科普遍水平更高,发文潜力也更大。对该领域的杰出和高产作者作进一步分析,计算可知核心作者最低发文量k≈5.61,即判断是否为竹林碳汇领域核心作者的标准是该作者至少应当发表相关论文6篇。根据该标准,结合表1可知:目前竹林碳汇领域约有72名核心作者,共发文650篇,发文数量占比低于普莱斯定律的50%。以作者发文数量次多的26篇计算k值时,则发文数量占比接近于50%;表明竹林碳汇研究的核心作者具有较高产出率,核心作者群已基本形成。对搜索得到的排位前20的核心作者调查(表2)可知:核心作者主要为中国学者,成果主要刊登在林学、生态学等学科的国际主流期刊上,如Agricultural and Forest Meteorology、Journal of Environmental Management等。
表 1 发文数量和作者数量的对应分布
Table 1. Distribution of the number of literatures and the number of authors
发文数
量/篇作者数
量/人占全部作者
的比例/%占发文数量为1的
作者的比例/%发文数
量/篇作者数
量/人占全部作者的
比例/%占发文数量为1的
作者的比例/%1 686 54.53 11 2 0.16 0.29 2 245 19.48 35.71 12 1 0.08 0.15 3 132 10.49 19.24 13 2 0.16 0.29 4 81 6.44 11.81 14 1 0.08 0.15 5 42 3.34 6.12 16 1 0.08 0.15 6 28 2.23 4.08 19 1 0.08 0.15 7 19 1.51 2.77 20 1 0.08 0.15 8 6 0.48 0.87 23 1 0.08 0.15 9 3 0.24 0.44 26 1 0.08 0.15 10 4 0.32 0.58 56 1 0.08 0.15 合计 1 258 99.99 表 2 核心作者及论文分布
Table 2. Core authors and distribution of their studies
序号 作者姓名 国家 所属单位 主要成果分布的期刊 1 周国模 中国 浙江农林大学 Agricultural and Forest Meteorology、Journal of Environmental Management 2 姜培坤 中国 浙江农林大学 Plant and Soil、Forest Ecology and Management 3 杜华强 中国 浙江农林大学 ISPRS Journal of Photogrammetry and Remote Sensing、IEEE Transactions on
Geoscience and Remote Sensing4 李永夫 中国 浙江农林大学 Agricultural and Forest Meteorology、Plant and Soil 5 王兵 中国 中国林业科学研究院 Scandinavian Journal of Forest Research、BioResources 6 宋新章 中国 浙江农林大学 Agricultural and Forest Meteorology、Soil Biology & Biochemistry 7 宋照亮 中国 天津大学 Global Change Biology、Science of The Total Environment 8 王海龙 中国 浙江农林大学 Environmental Science and Pollution Research、Journal of Soils and Sediments 9 庄舜尧 中国 中国科学院 Forestry、Journal of Soils and Sediments 10 张小川 加拿大 阿尔伯塔大学 Plant and Soil、Ecology 11 范少辉 中国 国际竹藤中心 Catena、Journal of Cleaner Production 12 施拥军 中国 浙江农林大学 Forest Ecology and Management、International Journal of Remote Sensing 13 吴家胜 中国 浙江农林大学 Journal of Plant Nutrition and Soil Science、Soil Research 14 江洪 中国 南京大学 Forest Ecology and Management、Science of The Total Environment 15 李永春 中国 浙江农林大学 Soil Biology & Biochemistry、Plant and Soil 16 徐小军 中国 浙江农林大学 Agricultural and Forest Meteorology、Annals of Forest Science 17 官凤英 中国 国际竹藤中心 Catena、Scientific Reports 18 NATH Arun Jyoti 美国 俄亥俄州立大学 Global Ecology and Conservation 19 KOMATSU Hikaru 日本 九州大学 Agricultural and Forest Meteorology 20 HUFF Matthew D 美国 欧道明大学 Journal of Environmental Management -
对发文机构的分析可知:“Zhejiang A&F University”(浙江农林大学)在竹林碳汇领域处于最为突出的位置,“Nanjing Forestry University”(南京林业大学)“Beijing Forestry University”(北京林业大学)“International Centre for Bamboo and Rattan”(国际竹藤中心)“Chinese Academy of Forestry”(中国林业科学研究院)等机构在竹林碳汇领域研究方面也具有较大贡献。从机构发文数量来看,位列前5的机构总发文量均超过25篇,其中浙江农林大学发文量(148篇)远多于其他机构,对发文总量的贡献达9.36%,即在竹林碳汇研究的国际论文中,每发表10篇论文约有1篇来自浙江农林大学。其他机构如南京林业大学(39篇)、北京林业大学(37篇)、国际竹藤中心(36篇)、中国林业科学研究院(26篇)等也较为“高产”。可见高校和国际竹藤中心等专业研究机构是竹林碳汇研究领域的主要发文机构。
另外,几乎所有边缘机构都与浙江农林大学、南京林业大学、北京林业大学、国际竹藤中心、中国林业科学研究院等核心机构具有联结。这表明浙江农林大学、南京林业大学、北京林业大学、国际竹藤中心、中国林业科学研究院等机构的竹林碳汇研究对该领域产生了深远影响。排名前五的机构贡献了总发文量的18.09%,说明这些核心机构在竹林碳汇研究方面影响力巨大,主导了竹林碳汇领域主要研究方向。从发文机构的地域分布来看,中国、美国、日本等许多国家和地区均有机构展开该领域的研究,但是发文量排名前五的机构均在中国境内,空间集聚性明显。随着竹林碳汇研究的深入,发文机构的空间分布也在不断拓展,越来越多的国家和地区开始关注竹林碳汇相关问题,如日本东京大学、美国佛罗里达国际大学、韩国仁荷大学、澳大利亚科廷科技大学等机构在该领域的发文数量正逐渐增加;表明竹林碳汇研究领域的学术影响力在不断增强,随着气候变化问题日益严重,通过竹林碳汇应对气候变化问题正受到世界各国越来越多学者的关注。
-
对关键词的分析可知:“Bamboo”(竹子)、“Carbon storage”(碳储量)、“Biomass”(生物量)是中心性最高的关键词,分别出现了647、552和514 次;“Moso bamboo”(毛竹)(305次)、“Emission reduction”(减排)(235次)、“Land use change”(地类使用变化)(212次)、“Temperature”(温度)(186次)等词出现的频次也较高。综合来看,高频关键词涵盖了“竹子生长”“竹林碳汇”“减排效应”等基本问题,展示了竹林碳汇研究的主要内容。
在CiteSpace输出的结果中,按照对数似然率提取的10个标签词分别为竹子、碳汇、生物量,毛竹、生态系统、温度、地类使用变化、二氧化碳(CO2)、影响因素、额外性;表达了自《京都议定书》签订以来学者们对竹林减排效应的关注程度。《京都议定书》签订之初仅规定了发达国家的减排责任,之后1998年召开的缔约方大会,提出了发展中国家可以通过清洁发展机制(CDM项目)减少排放以获取外汇收入,并将林业碳汇项目也纳入其中,随后林业碳汇相关研究开始受到广泛关注。竹子生长周期短,母竹种植到成林一般只需要6~8 a即可进行批量采伐并循环经营,因此其固碳效应引起了学者的重视。
最初的研究主要关注竹子自身生物量,并且大多以毛竹Phyllostachys edulis作为研究对象;之后的研究由单株拓展至竹林,林下层的生物量也受到关注,并细分为“地上生物量”“地下生物量”等。2005年《京都议定书》正式生效,欧盟成立了碳交易市场,竹林碳汇项目层面的研究也开始受到广泛关注。总体而言,竹林的碳汇量、竹子的碳减排效果、不同经营模式对碳减排的影响、不同地类竹林碳汇变化情况、竹林减排与其他森林减排的比较研究等是学界关注的重点。
近几年,相关研究被拓展至竹产品层面,竹产品的替代减排效应也开始受到关注;在当前竹林比较收益下降、退化加剧、弃营竹林增多等背景下,学者开始探讨竹林经营的影响因素、竹林增汇和增收的双重效应等。不同时期减排的国际形势及竹林面临的问题有所不同,相关研究热点也随之变化。
-
对文献的具体研究内容进行归纳和梳理,可以发现现有文献的主要观点包括5个方面。
①竹林有着较高的碳储量。竹林植被层中,毛竹林植被层碳储量最高,为137.9 t·hm−2[4],龙头竹Fargesia dracocephala为60.5 t·hm−2[7],桂竹P. bambusoides为49.8 t·hm−2[23];竹林生态系统中,慈竹Neosinocalamus affinis林生态系统碳储量最大,为135.95 t·hm−2,苦竹Pleioblastus amarus为135.81 t·hm−2、龙竹Dendrocalamus giganteus为10.46 t·hm−2[24]。全球竹林碳储量巨大,有研究表明[13]:全球竹林碳储量约为1.81 Pg,占全球森林碳储量的0.50%,其中,中国竹林碳储量在中国森林生态系统碳总量中占比超过10%。学者们预测,未来50 a,竹林碳储量还将持续增长[7,25]。可见,竹林及其生态系统在区域和国家碳平衡中发挥重要作用,并具有较大的提升潜力。
②竹林固碳能力明显高于其他树林。相比于其他树种,竹子生长更快速,固碳能力更强[26]。对竹子与其他树种固碳能力的比较发现[13]:若以60 a为生长周期,毛竹固碳能力分别是杉木Cunninghamia lanceolata和桉树Eucalyptus spp.的1.22倍和1.26倍,毛竹林碳汇量(8.13 Mg·hm−2·a−1)是杉木(3.35 Mg·hm−2·a−1)的2.43倍[23];测定毛竹、台湾红柏树Chamaecyparis formosensis和日本雪松Cryptomeria japonica的固碳能力可知,三者碳汇量分别为8.13、2.83和4.44 Mg·hm−2·a−1,毛竹固碳能力明显更强[27]。对竹林和其他树林生态系统碳储量的对比研究发现[28],毛竹林生态系统碳储量为104.83 t·hm−2,高于杉木(95.66 t·hm−2)和马尾松Pinus massoniana(96.49 t·hm−2),固碳量(9.64 Mg·hm−2·a−1)分别是杉木和马尾松的1.69和1.63倍。不难发现,竹子具有更强的固碳能力,是碳汇林经营的优选树种。
③经营方式对竹林固碳能力影响显著。已有研究表明[17]:对竹林进行集约经营有利于提高竹林固碳能力,集约经营的毛竹林植被层固碳量为12.75 Mg·hm−2·a−1,是粗放经营(8.14 Mg·hm−2·a−1)的1.56倍。但是长期的集约经营也可能带来土壤退化、养分流失等问题,加剧竹林退化,因此越来越多的学者开始倡导适度经营。有研究表明[28]:适度经营模式下竹林中土壤碳储量约为59.94 Mg·hm−2·a−1,分别为集约经营和粗放经营的2.36倍和1.86倍,适度经营明显提高了竹林土壤固碳水平。同时对竹林结构进行改善,适度间伐[9],优化竹林经营管理策略[29]等也对提高竹林碳汇量具有积极影响。因此,竹林的科学经营是提高其固碳水平的必要举措。
④竹制品具有较大的替代减排潜力。竹制品碳储水平高,使用竹制品不仅可以减缓竹子的碳排放,还可以对非竹类产品产生替代减排效应[30]。对竹板材和木板材碳储水平的研究发现[31]:竹板材碳储水平更高,从全生命周期的视角来看,1 kg竹板材和木板材所封存的碳分别为0.68 kg和0.19 kg,前者明显高于后者。同时竹材的使用生态成本(0.52欧元·m−3)明显低于木材(2.11欧元·m−3)和钢材(10.86欧元·m−3)[32]。就性能而言,建筑中使用竹材比同质量的木材和混凝土强度和刚性更大[33],认为竹材具有较强的可替代性。可见,竹制品替代减排价值巨大。需要指出的是,竹制品替代减排效应的研究偏少,且主要就技术视角展开相关讨论,实际应用时还需考虑竹制品成本收益和消费者偏好等因素,相关研究值得进一步深入和拓展。
⑤竹林具备增加森林碳汇和增加农户收益的双重功效。随着竹子价格的下降和劳动力成本的增加,退化竹林、弃营竹林越来越多,传统的竹林经营效率下降,竞争优势逐渐消减。与之相反,碳汇竹林生态效益凸显[34],提高竹林综合产出水平潜力较大;同时碳汇竹林经营时减少扰动的技术要求,对增加竹林生态系统的综合固碳水平有利[35],对节约劳动成本也具有积极影响。有研究对竹林经营中碳汇收益进行了模拟分析[14,36],结果表明:考虑竹林碳汇收益能有效提高竹林的收益水平,可以弥补传统竹林比较优势下降而引起的收益损失;即竹林在应对气候变化和提升农户收益方面均具有积极效应[13]。这些研究在探讨竹林增汇功效的同时,也为研究如何应对传统竹林低效率的升级改造提供了参考。值得一提的是,尽管该方面的研究无论对于应对全球气候变化还是解决当前竹产业持续发展问题均尤为重要,但相关研究主要为理论探讨,未来可以结合碳汇项目发展等课题展开深入探讨,为解决现实问题提供切实可行的方案。
对竹林碳汇计量和监测的研究则认为:竹林碳汇计量范围应该包括乔木层生物量、灌木层生物量和地下层生物量[37];计算竹林碳汇量时可运用竹类植物生长模型或生物量变化模型[38],也可借助遥感等现代化技术对竹林碳汇变化进行估计[39],还可以使用全生命周期方法(LCA)对竹子从种植-采伐-加工-竹制品使用寿命结束的全过程或部分过程的碳变化进行估计[31]。
-
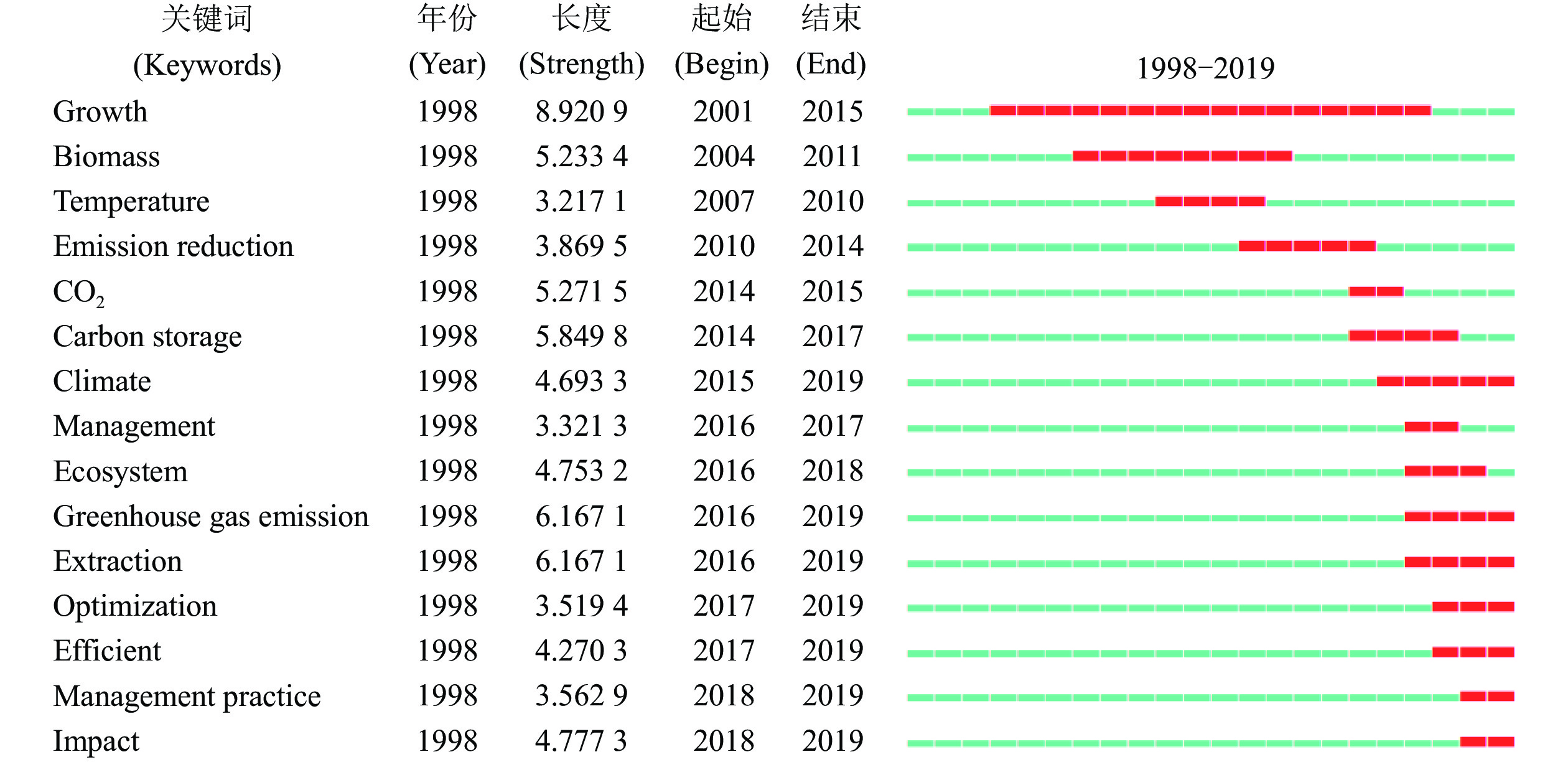
为探索不同时期竹林碳汇研究相关热点的变化情况,利用CiteSpace软件的突发检测功能对各期段的突发关键词进行归纳分析。由图2可知:自1998年以来,竹林碳汇领域突现了许多高频关键词,排名前15的突现高频关键词的突发变迁,与全球气候变化相关协议签订后,气候问题得到各国政府重视、森林碳汇在减排中的贡献凸显的时代背景息息相关,同时也体现了传统竹林经营比较收益下降背景下,学者们对通过竹林碳汇解决竹林持续经营问题的重视程度。
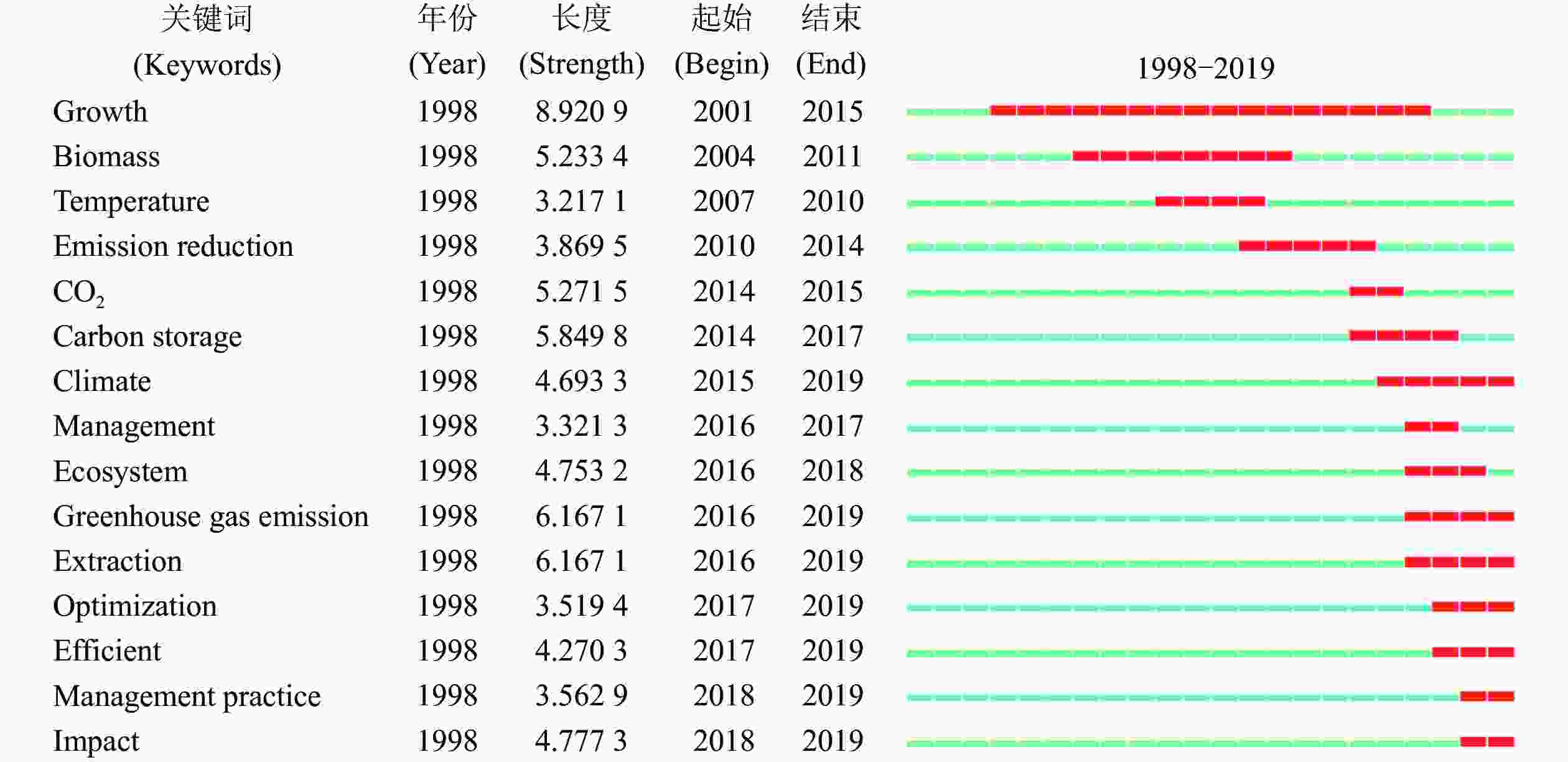
图 2 排名前15的突发关键词检测
Figure 2. Results of top 15 keywords burst detection
具体来看,随着时间推移,“生物量”“碳储量”“CO2”“气候”“温室气体排放”“优化”“管理实践”等突发关键词不断涌现。受日益加剧的气候变暖问题影响,各国政府开始积极探索减排策略及优化方案,关键词热点变化与国际控排形势变化出现趋同效应。关键词变迁一方面体现了在控排监测技术不断完善背景下,竹林碳汇研究的不断拓展和深化;另一方面也体现了对劳动力成本上升、竹林经营比较收益下降、竹林退化/弃营等问题的密切关注,并试图通过经营碳汇竹林为解决该类问题提出优化策略。研究热点最初集中于对该领域基本问题探索,之后逐渐转为现实思考与应用,通过竹林碳汇及减排解决竹林持续经营问题的理论思考,研究焦点也由论证竹林的固碳水平逐渐转为提出更优的竹林减排方案及竹林增收策略。
-
本研究以Web of Science核心合集数据库收录的竹林碳汇研究文献为对象,借助CiteSpace软件对该领域的文献信息进行综合分析,以展现该领域主要发展历程、知识体系及研究热点的演变特征。研究结果表明:①竹林碳汇发文量不断增多,年均文献量有明显的上升趋势。自《京都议定书》签订以来的20余a里,竹林碳汇发文量约75篇·a−1,2013年发文量超过100篇·a−1,发文数量有明显的加速增长趋势。②竹林碳汇的发文作者间合作紧密,核心发文机构具有地域集聚性。核心作者有较高的科学生产力和学术影响力,已形成核心作者群;核心发文机构的中心性地位较高,对该领域具有较大影响,主要位于中国境内,地域集聚特征明显。③现有文献集中于研究竹林碳储量、竹林固碳能力、经营方式、竹制品碳储量、竹林增汇和增收效应等方面。文献中的主要观点包括竹林有着较高的碳储量、竹林固碳能力明显高于其他树林、经营方式对竹林固碳能力具有显著影响、竹制品具有较大的替代减排潜力、竹林具备增加森林碳汇和增加农户收益的双重功效等。④竹林碳汇领域突发关键词的演变与国际气候政策变化、竹林经营中面临的困境联系较为紧密。许多高频关键词都体现或代表了国际社会气候政策变化的阶段性特征,随着竹林退化问题加剧、弃营竹林增加,通过竹林碳汇解决竹林可持续经营问题越来越受关注。
-
自《京都议定书》签订以来,竹林碳汇研究得到较高重视并迅速发展,一批以中国学者为主的科研人员建立并完善了竹林碳汇相关研究的学术体系,为全球竹林碳汇研究奠定了重要基础。总的来说,竹林碳汇经过20余a发展,相关研究已经得到不断拓展和深化,竹子固碳能力、减排效应、替代减排等领域取得了丰厚的研究成果。这些重要成果为往后相应领域的进一步研究提供了丰富资料。但是相关研究也存在一些不足,主要体现为:①核心作者群及核心发文机构均过于集中,研究的参与群体相对单薄。现有研究中主要的核心作者和核心发文机构均在中国,研究的主要参与群体为中国学者,空间集聚度较高,对长远发展不利,应吸引世界各地更多机构和学者积极参与。②内容过于集中于理论,与现实结合偏弱。现有研究大量是从理论视角、借助实验数据分析计算竹林碳储量和竹林固碳能力,如何将这些成果运用于应对气候变化以及解决现实中竹林退化、竹林增收等问题仍需要深化;对于竹制品的替代减排效应也大多停留于技术视角,缺乏现实应用方面的研究。③主要成果在促进减排和增收方面的实际贡献较低。目前对于减排和增收相结合的分析主要通过情景模拟分析等进行,其结论仅停留于理论参考层面,要实现实际应用还需考虑现实情景的多面性和复杂性,但相关研究较少。
鉴于上述不足,结合竹林碳汇研究发展趋势,提出以下建议:①加强学术合作与交流,扩大竹林碳汇领域的影响力。以核心作者群为基础,发挥团队效应,增加作者之间、机构之间、学科之间的合作和交流,尤其应该加强国际学术交流与合作,以吸引更多学者参与相关研究,拓宽竹林碳汇研究视野,丰富研究范式,进一步促进该领域的发展演进。②强化现实问题的对策类研究,提升竹林碳汇理论的现实应用价值。应使用竹林碳汇相关理论,研究制定解决现实问题的优化策略,例如对退化、弃营竹林进行碳汇林改造升级以促进竹林持续经营,展开竹制品消费偏好调查,从市场层面引导实现替代减排效应,以发挥竹林碳汇理论的实践价值。③着重挖掘竹林碳汇应对气候变化及提高林农收益方面的潜力,充分发挥竹林减排和增收的双重效应。在进一步探索竹林碳汇的多重减排效应,如竹林的增汇效应、竹产品的替代减排效应等基础上,基于国内外碳市场交易实践,积极开展竹林碳汇项目开发、交易获利等方面的深入探索,为传统低效率竹林的碳汇林转型改造升级提供从经营至创收整套的经营策略,达到森林增汇和农户增收的双重目标。
A review and prospect of global bamboo forest carbon sink research based on CiteSpace
-
摘要: 随着全球气候变暖问题日益严重,竹林碳汇研究已经逐渐演化为碳汇相关研究领域的重要分支之一。本研究借助CiteSpace软件,对自《京都议定书》签订以来,Web of Science核心合集数据库中研究竹林碳汇的文献进行了回顾和梳理,结果表明:①竹林碳汇研究处于快速发展期,该领域正受到越来越多学者的密切关注;②发文作者之间合作紧密,核心作者发文能力较强,并且已经基本形成核心作者群,核心发文机构地域集聚性明显,对该领域贡献巨大;③现有研究主要集中于探讨竹林碳储量、竹林固碳能力、经营模式、竹制品碳储量、竹林增汇和增收效应等方面,主要观点包括竹林有着较高的碳储量、竹林固碳能力明显高于其他树林、经营模式对竹林固碳能力具有显著影响、竹制品具有较大的替代减排潜力、竹林具备增加森林碳汇和增加农户收益的双重功效等;④关键词的变迁与通过竹林碳汇应对全球气候变化,以及促进竹林可持续经营密切关联。未来应该扩大竹林碳汇领域的影响力,加强现实问题的对策类研究,以挖掘竹林碳汇减排和增收的双重效应。图2表2参39Abstract: As climate warming further worsens, the studies of bamboo forest carbon sink have gradually evolved into one of the important branches of carbon sink research field. With the employment of CiteSpace software, a literature review was conducted of researches on bamboo forest carbon sink included in the Web of Science Core Collection for analysis and prospects into the future. The results showed that a) with close attention drawn from more and more researchers, bamboo forest carbon sink research is embracing a rapid development; b) with the close cooperation among authors, substantial progress has been achieved in this field with a collection of research papers published, forming a strong core author group while the core institutions have shown obvious geographical agglomeration with great contributions made to this field; c) the existing studies mainly focus on the discussion of bamboo forest carbon storage, bamboo forest carbon sequestration capacity, management methods, bamboo products carbon storage, and the effects of bamboo forests on the promotion of carbon storage and farmers’ income and it has been concluded that bamboo forests have a high level of carbon storage, the carbon sequestration capacity of bamboo forests is significantly higher than that of other forests, management methods have a significant impact on on the promotion of the carbon sequestration capacity of bamboo forest, bamboo products have great potential for substitution and emission reduction and bamboo forests have the double effect on the promotion of carbon sink and farmers’ income; d) the change of research keywords is a reflection of the efforts to cope with the global climate change and promote the sustainable management of bamboo forests by improving the carbon sequestration capacity. In the future, it is necessary to enhance the influence of bamboo forest carbon sinks and encourage countermeasure researches on practical issues to give full play to the double effect of bamboo forest carbon sinks on the reduction of emissions and the promotion of farmers’ income. [Ch, 2 fig. 2 tab. 39 ref.]
-
Key words:
- bamboo forest carbon sink /
- visual analysis /
- research focuses /
- CiteSpace /
- review
-
图 1 历年竹林碳汇文献数量变化
Figure 1 Changes in the number of studies on bamboo forest carbon sinks over the years
图 2 排名前15的突发关键词检测
红色区域代表高频关键词突发及持续的期段
Figure 2 Results of top 15 keywords burst detection
表 1 发文数量和作者数量的对应分布
Table 1. Distribution of the number of literatures and the number of authors
发文数
量/篇作者数
量/人占全部作者
的比例/%占发文数量为1的
作者的比例/%发文数
量/篇作者数
量/人占全部作者的
比例/%占发文数量为1的
作者的比例/%1 686 54.53 11 2 0.16 0.29 2 245 19.48 35.71 12 1 0.08 0.15 3 132 10.49 19.24 13 2 0.16 0.29 4 81 6.44 11.81 14 1 0.08 0.15 5 42 3.34 6.12 16 1 0.08 0.15 6 28 2.23 4.08 19 1 0.08 0.15 7 19 1.51 2.77 20 1 0.08 0.15 8 6 0.48 0.87 23 1 0.08 0.15 9 3 0.24 0.44 26 1 0.08 0.15 10 4 0.32 0.58 56 1 0.08 0.15 合计 1 258 99.99  下载: 导出CSV
下载: 导出CSV
表 2 核心作者及论文分布
Table 2. Core authors and distribution of their studies
序号 作者姓名 国家 所属单位 主要成果分布的期刊 1 周国模 中国 浙江农林大学 Agricultural and Forest Meteorology、Journal of Environmental Management 2 姜培坤 中国 浙江农林大学 Plant and Soil、Forest Ecology and Management 3 杜华强 中国 浙江农林大学 ISPRS Journal of Photogrammetry and Remote Sensing、IEEE Transactions on
Geoscience and Remote Sensing4 李永夫 中国 浙江农林大学 Agricultural and Forest Meteorology、Plant and Soil 5 王兵 中国 中国林业科学研究院 Scandinavian Journal of Forest Research、BioResources 6 宋新章 中国 浙江农林大学 Agricultural and Forest Meteorology、Soil Biology & Biochemistry 7 宋照亮 中国 天津大学 Global Change Biology、Science of The Total Environment 8 王海龙 中国 浙江农林大学 Environmental Science and Pollution Research、Journal of Soils and Sediments 9 庄舜尧 中国 中国科学院 Forestry、Journal of Soils and Sediments 10 张小川 加拿大 阿尔伯塔大学 Plant and Soil、Ecology 11 范少辉 中国 国际竹藤中心 Catena、Journal of Cleaner Production 12 施拥军 中国 浙江农林大学 Forest Ecology and Management、International Journal of Remote Sensing 13 吴家胜 中国 浙江农林大学 Journal of Plant Nutrition and Soil Science、Soil Research 14 江洪 中国 南京大学 Forest Ecology and Management、Science of The Total Environment 15 李永春 中国 浙江农林大学 Soil Biology & Biochemistry、Plant and Soil 16 徐小军 中国 浙江农林大学 Agricultural and Forest Meteorology、Annals of Forest Science 17 官凤英 中国 国际竹藤中心 Catena、Scientific Reports 18 NATH Arun Jyoti 美国 俄亥俄州立大学 Global Ecology and Conservation 19 KOMATSU Hikaru 日本 九州大学 Agricultural and Forest Meteorology 20 HUFF Matthew D 美国 欧道明大学 Journal of Environmental Management
下载: 导出CSV
-
[1] 周国模, 姜培坤. 毛竹林的碳密度和碳贮量及其空间分布[J]. 林业科学, 2004, 40(6): 20 − 24. ZHOU Guomo, JIANG Peikun. Density, storage and spatial distribution of in Phyllostachys pubescens forest [J]. Sci Sliv Sin, 2004, 40(6): 20 − 24. [2] 周芳纯. 毛竹竹冠结构的研究[J]. 南京林产工业学院学报, 1982(3): 46 − 73. ZHOU Fangchun. Study on the structure of bamboo crown of Phyllostachys pubescens [J]. J Nanjing Technol Coll For Prod, 1982(3): 46 − 73. [3] 聂道平. 毛竹林结构的动态特性[J]. 林业科学, 1994, 30(3): 201 − 208. NIE Daoping. Structural dynamics of bamboo forest stand [J]. Sci Sliv Sin, 1994, 30(3): 201 − 208. [4] ISAGI Y, KAWAHARA T, KAMO K, et al. Net production and carbon cycling in a bamboo Phyllostachys pubescens stand [J]. Plant Ecol, 1997, 130(1): 123. [5] 周国模. 毛竹林生态系统中碳储量、固定及其分配与分布的研究[D]. 杭州: 浙江大学, 2006. ZHOU Guomo. Carbon Storage, Fixation and Distribution in Mao Bamboo (Phyllostachys pubescens) Stands Ecosystem[D]. Hangzhou: Zhejiang University, 2006. [6] CHEN Xiangang, ZHANG Xiaoquan, ZHANG Yiping, et al. Changes of carbon stocks in bamboo stands in China during 100 years [J]. For Ecol Manage, 2009, 258(7): 1489 − 1496. [7] NATH A J, DAS G, DAS A K. Above ground standing biomass and carbon storage in village bamboos in North East India [J]. Biomass Bioenergy, 2009, 33(9): 1188 − 1196. [8] KUEHL Y, LI Y, HENLEY G. Impacts of selective harvest on the carbon sequestration potential in Moso bamboo (Phyllostachys pubescence) plantations [J]. For Trees Livelihoods, 2013, 22(1): 1 − 18. [9] MAO Fangjie, ZHOU Guomo, LI Pingheng, et al. Optimizing selective cutting strategies for maximum carbon stocks and yield of Moso bamboo forest using BIOME-BGC model [J]. J Environ Manag, 2017, 191: 126 − 135. [10] 敖贵艳, 吴伟光, 曹先磊, 等. 基于三阶段DEA模型的碳汇竹林生产效率分析: 来自浙江安吉的实证[J]. 农林经济管理学报, 2019, 18(5): 656 − 666. AO Guiyan, WU Weiguang, CAO Xianlei, et al. Analysis on production efficiency of carbon sink bamboo forest based on three-stage DEA model: evidence from Zhejiang Anji [J]. J Agro-For Econ Manage, 2019, 18(5): 656 − 666. [11] 张小全, 李怒云, 武曙红. 中国实施清洁发展机制造林和再造林项目的可行性和潜力[J]. 林业科学, 2005, 41(5): 139 − 143. ZHANG Xiaoquan, LI Nuyun, WU Shuhong. Analysis on feasibility and potentiality of afforestation and reforestation under the clean development mechanism of the Kyoto Protocol [J]. Sci Sliv Sin, 2005, 41(5): 139 − 143. [12] NATH A J, LAL R, DAS A K. Managing woody bamboos for carbon farming and carbon trading [J]. Global Ecol Conserv, 2015, 3: 654 − 663. [13] LOBOVIKOV M, SCHOENE D, LOU Yiping. Bamboo in climate change and rural livelihoods [J]. Mitigation Adapt Strategies Glob Change, 2012, 17(3): 261 − 276. [14] WU Weiguang, LIU Qiang, ZHU Zhen, et al. Managing bamboo for carbon sequestration, bamboo stem and bamboo shoots [J]. Small-scale For, 2015, 14(2): 233 − 243. [15] DWIVEDI A K, KUMAR A, BAREDAR P, et al. Bamboo as a complementary crop to address climate change and livelihoods -insights from India [J]. For Policy Econ, 2019, 102: 66 − 74. [16] 陈茂铨, 金晓春, 吴林森, 等. 竹林碳汇功能及其影响因子研究进展[J]. 竹子研究汇刊, 2010, 29(3): 5 − 9. CHEN Maoquan, JIN Xiaochun, WU Linsen, et al. Research progress of carbon sink and its influencing factors on bamboo forest [J]. J Bamboo Res, 2010, 29(3): 5 − 9. [17] ZHOU Guomo, MENG Cifu, JIANG Peikun. Review of carbon fixation in bamboo forests in China [J]. Bot Rev, 2011, 77(3): 262 − 270. [18] 邱均平, 刘敏. 1998−2007年我国竞争情报领域论文的计量规律研究[J]. 情报科学, 2009, 27(9): 1281 − 1285. QIU Junping, LIU Min. Research on the quantitive law of the article on competitive intelligence field from 1998 to 2007 in China [J]. Inf Sci, 2009, 27(9): 1281 − 1285. [19] 陈悦, 陈超美, 刘则渊, 等. CiteSpace知识图谱的方法论功能[J]. 科学学研究, 2015, 33(2): 242 − 253. CHEN Yue, CHEN Chaomei, LIU Zeyuan, et al. The methodolog function of CiteSpace mapping knowledge domains [J]. Stud Sci Sci, 2015, 33(2): 242 − 253. [20] 侯瑞芳, 李玲, 陈嘉勇, 等. 基于文献数据规律的机构知识库数据转换模型研究[J]. 图书情报工作, 2014, 58(17): 131 − 135. HOU Ruifang, LI Ling, CHEN Jiayong, et al. Exploring the regularities in bibliographic data: research on the data conversion model of institutional repository [J]. Libr Inf Serv, 2014, 58(17): 131 − 135. [21] 孙建军. 文献情报计量理论和方法[M]. 南京: 南京大学出版社, 1994. [22] 国家林业和草原局. 我国首个竹子碳汇造林方法学正式颁布实施[DB/OL]. (2012-12-03)[2020-11-25]. http://www.forestry.gov.cn/portal/main/s/72/content-573717.html. [23] YEN Tianming, LEE J S. Comparing aboveground carbon sequestration between moso bamboo (Phyllostschys heterocycla) and China fir (Cunninghamia lanceolata) forests based on the allometric model [J]. For Ecol Manage, 2011, 261(6): 995 − 1002. [24] ZHOU Yufeng, ZHOU Guomo, DU Huaqiang, et al. Biotic and abiotic influences on monthly variation in carbon fluxes in on-year and off-year Moso bamboo forest [J]. Trees, 2019, 33(1): 153 − 169. [25] LI Pingheng, ZHOU Guomo, DU Huaqiang, et al. Current and potential carbon stocks in Moso bamboo forests in China [J]. J Environ Manage, 2015, 156: 89 − 96. [26] SCURLOCK J M O, DAYTON D C, HAMES B. Bamboo: an overlooked biomass resource? [J]. Biomass Bioenerg, 2000, 19: 229 − 244. [27] JIANG Peikun, MENG Cifu, ZHOU Guomo, et al. Comparative study of carbon storage in different forest stands in subtropical China [J]. Bot Rev, 2011, 77(3): 242 − 251. [28] LI Chong, SHI Yongjun, ZHOU Guomo, et al. Effects of different management approaches on soil carbon dynamics in Moso bamboo forest ecosystems [J]. Catena, 2018, 169: 59 − 68. [29] XU Lin, SHI Yongjun, ZHOU Guomo, et al. Structural development and carbon dynamics of Moso bamboo forests in Zhejiang Province, China [J]. For Ecol Manage, 2018, 409: 479 − 488. [30] GU Lei, ZHOU Yufeng, MEI Tingting, et al. Carbon footprint analysis of bamboo scrimber flooring: implications for carbon sequestration of bamboo forests and its products[J]. Forests, 2019, 10: 51. doi: 10.3390/f10010051 [31] VOGTLÄNDER J G, van der VELDEN N M, van der LUGT P. Carbon sequestration in LCA, a proposal for a new approach based on the global carbon cycle; cases on wood and on bamboo [J]. Int J Life Cycle Assess, 2014, 19: 13 − 23. [32] VOGTLÄNDER J, van der LUGT P, BREZET H. The sustainability of bamboo products for local and Western European applications LCAs and land-use [J]. J Cleaner Prod, 2010, 18(13): 1260 − 1269. [33] JANSSEN J J A. Designing and building with bamboo INBAR technical report 20[R]. Beijing: International Network for Bamboo and Rattan (INBAR), 2000. [34] SONG Xinzhang, ZHOU Guomo, JIANG Hong, et al. Carbon sequestration by Chinese bamboo forests and their ecological benefits: assessment of potential, problems, and future challenges [J]. Environ Rev, 2011, 19: 418 − 428. [35] YIN Jiayang, GE Zhipeng, DENG Xu, et al. Abandonment lead to structural degradation and changes in carbon allocation patterns in Moso bamboo forests[J]. For Ecol Manage, 2019, 449: 117449. doi: 10.1016/j.foreco.2019.117449. [36] GU Lei, WU Weiguang, JI Wei, et al. Evaluating the performance of bamboo forests managed for carbon sequestration and other co-benefits in Suichang and Anji, China[J]. For Policy Econ, 2019, 106: 101947. doi: 10.1016/j.forpol.2019.101947. [37] EGGLESTON H S, BUENDIA L, MIWA K, et al. IPCC guidelines for national greenhouse gas inventories[R]. Geneva: IPCC, 2006 [38] LUYSSAERT S, SCHULZE E D, BÖRNER A, et al. Old-growth forests as global carbon sinks [J]. Nature, 2008, 455: 213 − 215. [39] DU Huaqiang, ZHOU Guomo, GE Hongli, et al. Satellite-based carbon stock estimation for bamboo forest with a non-linear partial least square regression technique [J]. Int J Remote Sensing, 2012, 33(6): 1917 − 1933. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200501
 点击查看大图
点击查看大图

计量
- 文章访问数: 3217
- HTML全文浏览量: 830
- PDF下载量: 110
- 被引次数: 0

