




-
海岛是保护海洋环境、维护生态平衡的重要平台,受大陆性和海洋性气候的共同影响,形成了海岛特殊独立的生境。这种生境兼具海陆双重特征[1],是生物多样性保护的关键区域。群落结构是认识群落组成、变化及发展趋势的基础,体现群落对环境的适应性、动态特征及生态功能,并能影响生态系统的功能特征[2]。目前,针对海岛植物群落结构已有部分研究,但由于独特的地理位置、有限的面积和清晰的空间边界,不同海岛的植物群落结构可能存在一定差异[3]。郑丽婷等[4]研究了山东省庙岛群岛典型植物多样性及其对环境因子的响应,庙岛群岛有麻栎Quercus acutissima、刺槐Robinia pseudoacacia、黑松Pinus thunbergii、荆条Vitex negundo等4种典型植物群落。杨佰润等[5]对浙江省大陈岛植物群落结构、物种组成及其与土壤因子的关系进行研究,发现大陈岛植物群落主要以木麻黄Casuarina equisetifolia、柃木Eurya japonica、天仙果Ficus erecta为优势物种。周厚诚等[6]分析了广东省汕头市南澳岛次生林的群落结构,包括物种结构、水平结构、垂直结构和组织水平结构。YANG等[7]调查研究了南海20个热带岛屿的木本植物,并收集了土壤样本,分析岛屿面积、隔离、气候和土壤因素如何影响不同生活型的植物群落。浙江省拥有中国最丰富的海岛资源,但对海岛植被缺乏系统性研究,对海岛群落的组成、结构、植物区系和多样性特征尚未形成系统性的科学认知。在岱山岛开展典型森林群落树种组成与结构研究,对于丰富和完善浙江省海岛森林群落的认知具有重要的理论与应用价值。
岱山岛位于浙江省舟山市岱山县境内,处于中亚热带常绿阔叶林地带,属浙闽山区甜槠Castanopsis eyrei、木荷Schima superba林区以及天台山、括苍山地、岛屿植被片。由于长期人为干扰,原生植被破坏殆尽,近年来,在造林、抚育等生态修复措施大力推动下,岱山岛森林群落得到较好恢复。目前,未见关于岱山岛森林群落树种组成结构的研究报道。本研究以浙江省岱山岛1 hm2样地内木本植物为研究对象,开展森林群落树种组成和结构研究,探讨岱山岛森林群落特征,包括物种的组成、区系成分、径级结构、物种多样性、树种组合等,有助于深入了解岱山岛森林群落,旨在为岱山岛森林群落恢复、群落多样性维护、群落稳定性提高和群落树种配置优化提供指导。
-
岱山岛位于浙江省北部沿海,隶属于浙江省舟山市岱山县,为岱山县境内最大岛,面积为104.97 km2,为舟山群岛第二大岛。地理位置为30°07′~30°38′N,121°31′E~123°17′E。岱山岛属北亚热带南缘季风海洋性气候区,年平均气温为18.0 ℃,最高气温为35.0 ℃,最低气温为−4.0 ℃,年平均降水量为1 038.4 mm,年平均风速为6.1 m·s−1。岛上森林类型主要为常绿落叶阔叶混交林,乔木层主要树种有麻栎、樟树Cinnamomum camphora等。
-
为深入剖析岱山岛森林群落结构,在查阅相关资料的基础上,初步了解该区域植被变化过程,并通过现场踏查比较,在岱山岛森林分布集中连片、面积最大、保护最好的磨心山风景区的山体中部,选取具有代表性、相对均匀、近期受人类干扰少的区域设立样地。样地总面积为1 hm2,中心点坐标为30°15′20″N,122°11′13″E,海拔为80 m,东南坡。在该样地中共设置25个20 m×20 m的连续调查样方(图1)。以样地西南角为原点,从原点出发,横轴(x轴)向正东方向,纵轴(y轴)向正北方向,对样地内所有胸径(DBH)≥1 cm的木本植物个体进行调查,记录个体所处样方号、坐标、物种名称、胸径、树高、生长状况等信息。
图 1 岱山岛样方编号示意图
Figure 1. Schematic diagram of plot numbers on Daishan Island
-
森林群落研究中常采用重要值来衡量某一树种在群落中的重要性,通过重要值分析树种在森林群落中的地位和优势程度。物种重要值(VI)=(相对优势度+相对多度+相对频度)/3[8]。其中:相对优势度=(样方内某一树种的个体胸高断面积之和/所有树种个体胸高断面积总和)×100%;相对多度=(样方内某一树种的密度/所有树种的总密度)×100%;相对频度=(样方内某一树种的频度/所有树种的频度总和)×100%。
-
森林的垂直结构是森林群落的基本特征之一,林层的变化会影响群落内的植物生长和物种多样性等。在本研究中,运用K-means聚类方法将样地内DBH≥1 cm的全部个体的树高进行分类,用轮廓系数法对分类结果进行检验[9]。
-
群落的年龄结构以径级结构代替,以1 cm为起测径级。根据岱山岛木本植物的基本情况,以5 cm为递增单元划分成9个区间,最大区间为40 cm及以上,并以5 cm为作图单元,最大作图区间为40~50 cm。
-
本研究采用群落尺度上常用的Shannon-Wiener多样性指数、Pielou均匀度指数、Simpson多样性指数和Margalef丰富度指数表示植物群落的物种多样性。计算公式参照文献[10]。
-
以25个样方中各物种的重要值为基础数据,建立物种-样方矩阵,通过Ward离差平方和法进行层次聚类,距离测度选用经Hellinger转化的欧氏距离以适配重要值数据特性,初步识别样方分组结构,采用TWINSPAN (Two-way Indicator Species Analysis)双向指示种分析法优化分类[11]。TWINSPAN分析由PC-ORD Version 5完成。
为验证分类结果的生态意义,采用相似性分析 (Analysis of Similarities,ANOSIM)进行组间差异检验,基于Bray-Curtis距离值的秩次进行组间差异显著性检验,通过非参数置换检验(999次置换)计算统计量R。通过R检验组间差异是否显著大于组内差异,从而判断分组是否有意义。R取值范围为[−1,1],0<R≤1表示组间差异大于组内差异,−1≤R<0则表示组内差异大于组间差异,R=0表示组间和组内没有明显差异。通过P值检验组间差异是否有显著性,P<0.01表示组间差异极显著[12]。
-
研究样地共记录木本植物(DBH≥1 cm) 8 767株,隶属18科28属29种。群落平均树高为(4.11±1.95) m,平均胸径为(4.35±4.57) cm。常绿树种有12种1 853株,占总种数的41.38%,占总株数的21.14%,重要值为34.66%;落叶树种有17种
6914 株,占总种数的58.62%,占总株数的78.86%,重要值为65.34%。群落中优势树种基本特征详见表1。表 1 群落重要值排名前10树种统计
Table 1. Statistics of top 10 tree species with important value of community
树种 多度/
(株·hm−2)平均胸径/cm 平均树高/m 重要值/% 树种 多度/
(株·hm−2)平均胸径/cm 平均树高/m 重要值/% 天仙果 6 103 2.87±1.72 3.55±1.19 32.53 枸骨 221 2.32±1.16 3.23±0.92 3.69 麻栎 244 17.01±10.56 8.89±3.62 12.94 黄檀 220 5.03±2.85 5.22±1.42 3.23 杉木 618 9.48±3.93 5.43±1.85 11.37 算盘子 74 8.27±3.28 5.73±1.35 2.87 细齿叶柃 764 4.38±2.63 4.22±1.31 7.62 水杉 44 18.93±7.60 12.13±3.21 2.70 樟树 146 14.45±8.77 7.17±3.01 7.12 女贞 71 8.51±5.86 6.69±2.35 2.68 说明:天仙果Ficus erecta,麻栎Quercus acutissima,杉木Cunninghamia lanceolata,细齿叶柃Eurya nitida,樟树Cinnamomum camphora,枸骨Ilex cornuta,黄檀Dalbergia hupeana,算盘子Glochidion puberum,水杉Metasequoia glyptostroboides,女贞Ligustrum lucidum。 参考《中国植被》[13],将群落中物种个体数≤1株·hm−2的种定义为稀有种,个体数介于1~100株·hm−2的种定义为偶见种,个体数>100株·hm−2的种定义为常见种。结果显示:群落共有常见种7种,偶见种19种,稀有种3种。常见种包括天仙果(6 103株)、细齿叶柃Eurya nitida (764株)、杉木Cunninghamia lanceolata (618株)、麻栎(244株)、枸骨Ilex cornuta (221株)、黄檀Dalbergia hupeana (220株)、樟树(146株),常见种个体数之和占总个体数的94.86%。稀有种包括板栗Castanea mollissima、冬青Ilex chinensis、乌桕Sapium sebiferum,说明群落是少数物种占据主导地位。
-
将调查样地内DBH≥1 cm全部个体的树高进行K-means非监督聚类,当k=2时平均轮廓系数达最大值(0.68)。将群落分为上下2层(表2),群落上层共有783株,占总个体数8.93%,平均树高为(8.64±2.46) m,平均胸径为(14.8±7.71) cm,主要树种为麻栎、杉木、樟树、天仙果和水杉;群落下层共有7 984株,占总个体数91.07%,平均树高为(3.67±1.17) m,平均胸径为(3.33±2.32) cm,主要树种为天仙果、杉木、细齿叶柃、枸骨和樟树。
表 2 岱山岛森林群落垂直结构
Table 2. Vertical stratification of the forest community on Daishan Island
林层 主要树种 最低树高/m 最高树高/m 平均树高/m 平均胸径/cm 多度/(株·hm−2) 重要值/% 群落上层 麻栎 6.50 17.00 10.55±2.77 21.46±8.87 176 25.42 杉木 6.50 11.00 7.83±1.08 13.13±3.29 174 15.72 樟树 6.50 15.50 9.07±2.69 19.10±8.17 81 12.22 天仙果 6.50 11.00 7.02±0.71 8.73±3.00 122 10.22 水杉 6.50 18.00 12.13±3.21 18.93±7.60 44 7.24 群落下层 天仙果 1.00 6.00 3.48±1.09 2.75±1.46 5981 42.95 杉木 1.60 6.00 4.48±1.08 8.04±3.16 444 13.57 细齿叶柃 1.50 6.00 4.11±1.23 4.19±2.46 735 10.78 枸骨 1.80 5.50 3.22±0.89 2.31±1.16 220 4.47 樟树 2.00 6.00 4.81±1.17 8.65±5.42 65 4.36 -
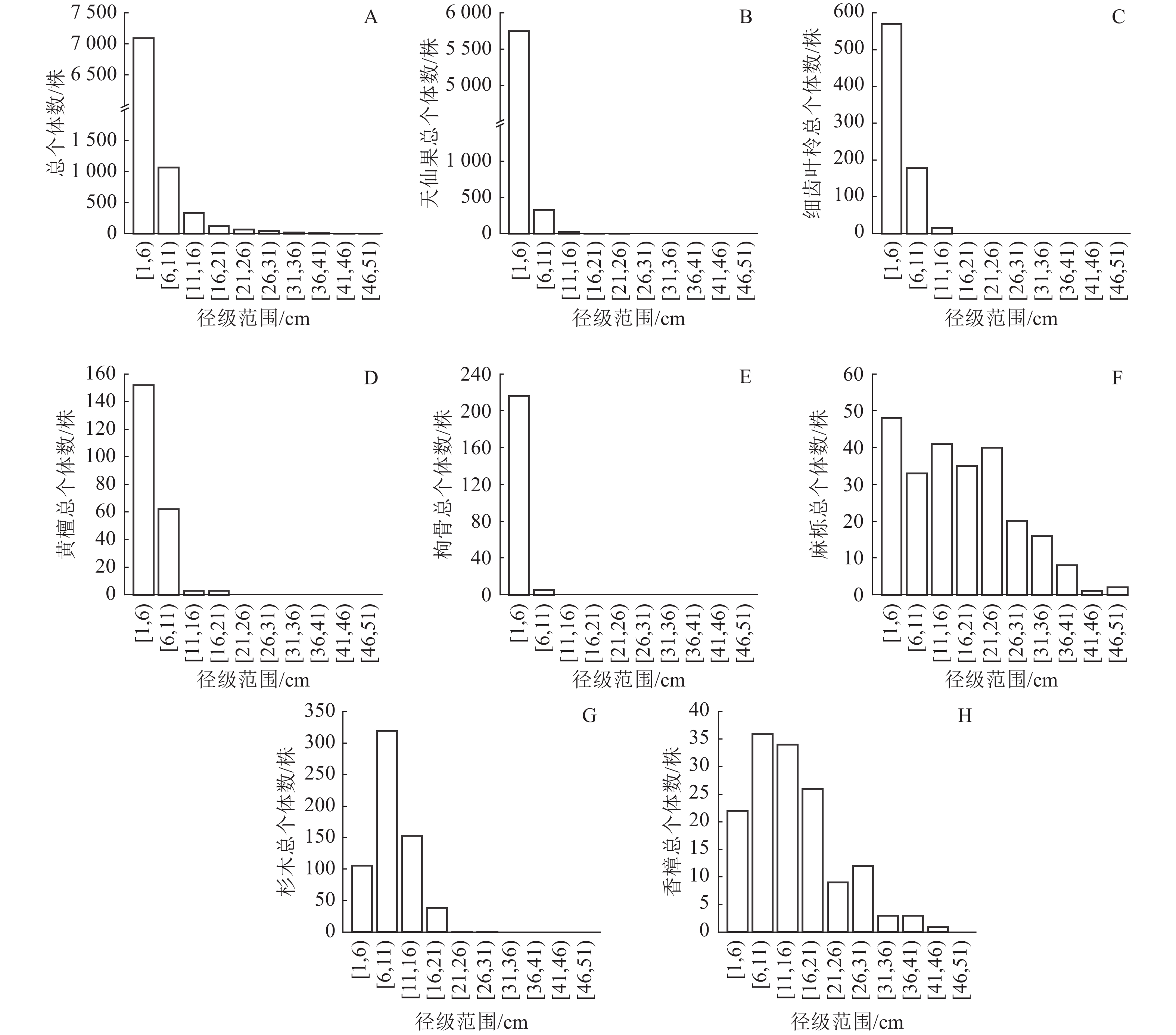
从图2A可以看出:群落中全部树种径级结构呈倒“J”型,1 cm≤DBH<6 cm的个体有7 090株(占总个体数的80.87%),6 cm≤DBH<11 cm的个体有1 070株(12.20%),11 cm≤DBH<16 cm的个体有333株(3.80%),16 cm≤DBH<21 cm的个体有127株(1.45%),DBH≥21 cm的个体有147株(1.68%)。群落平均胸径为4.35 cm,最大胸径的个体为麻栎,胸径达50.00 cm。说明整个群落以小径级和中小径级个体占绝对优势,占总个体数的93.07%,群落正处于恢复过程初期。
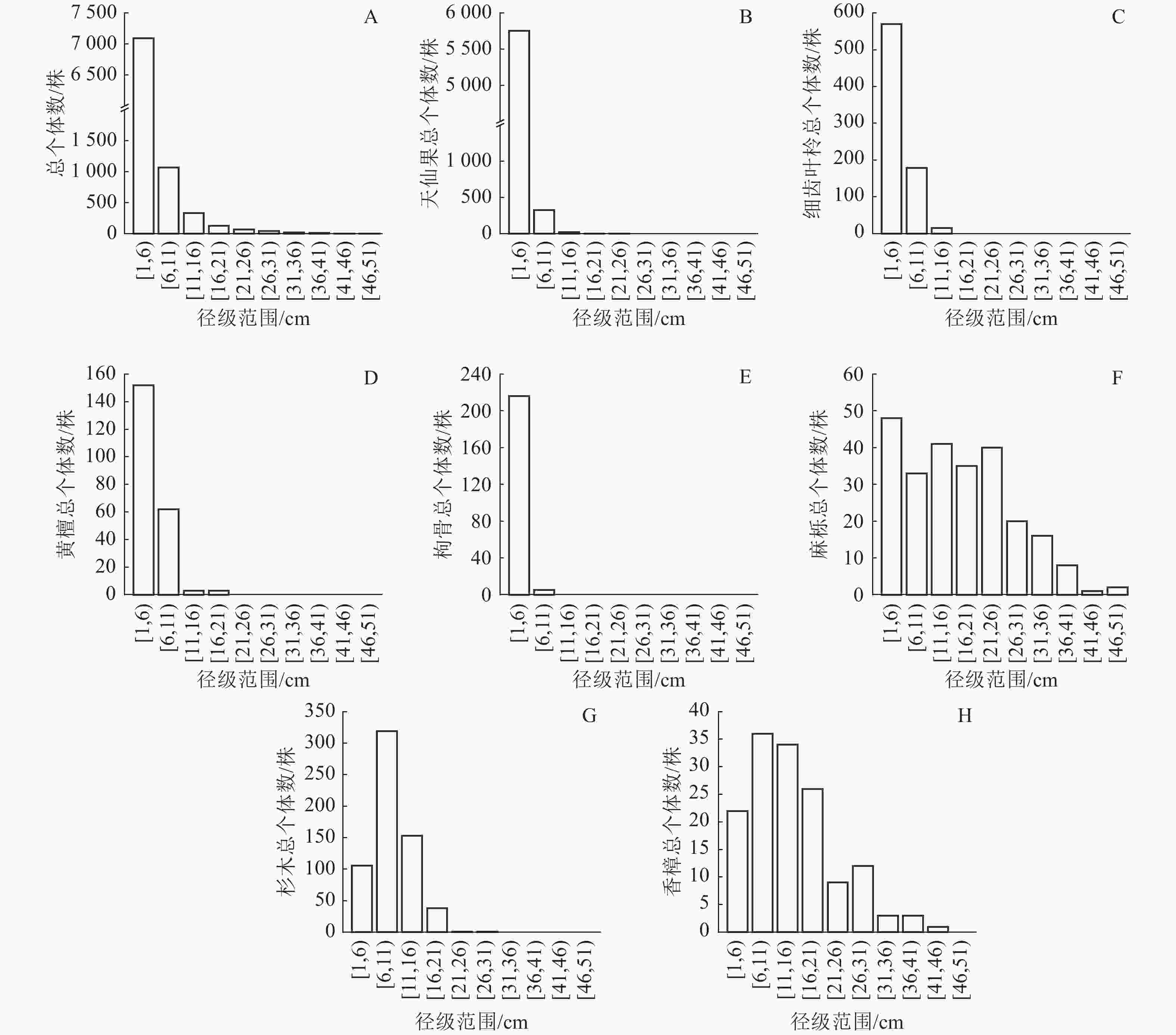
图 2 岱山岛群落常见种径级分布
Figure 2. Diameter class distribution of common species in Daishan Island community
从图2B可以看出:天仙果作为林下树种,个体数量多,胸径集中分布在1 cm≤DBH<6 cm,在群落中具有较高的更新能力和竞争力。由于天仙果有很强的萌生能力,容易产生大量幼苗和小径级个体,占据大量的林下空间,应当在森林管理中加以控制,为其他树种生长提供空间。从图2C和图2D可以看出:细齿叶柃和黄檀个体的胸径集中分布在1 cm≤DBH<11 cm。细齿叶柃是林下常绿树种,黄檀是珍贵树种,在群落中也具有一定的更新能力,应当促进其发展。从图2F可以看出:麻栎是落叶大乔木,在各个径级上分布较均匀,在群落中表现出较好的稳定性和竞争能力,能够在各个径级维持相对稳定的个体数量。从图2G可以看出:杉木以中小径级个体为主,缺少大径级个体,说明虽然其萌生能力较强,但在群落中竞争能力一般,难以形成大径级个体。樟树是亚热带常绿阔叶林主要树种,在更新能力、竞争能力和稳定性之间取得了较好的平衡,能够在林相改善中持续发挥重要作用,促进群落向常绿阔叶林演变(图2H)。
-
参照吴征镒[14−15]对科、属分布类型的划分方法,将分布区类型划分为4个大类,包括世界分布、热带成分(2~7)、温带成分(8~14)和中国特有。从表3可以看出:在科水平上,以热带成分为主,其中世界分布有5科,占27.78%,热带成分有9科,占50.00%,温带成分有4科,占22.22%。在属水平上,以热带成分为主的情况更加明显,其中热带成分有18属,占64.29%,温带成分有8属,占28.57%,中国特有属有2属,占7.14%。在种水平上,仍以热带成分为主,其中热带成分有19种,占65.52%,温带成分有8种,占27.58%,中国特有种有2种,占6.90%。
表 3 岱山岛群落木本植物区系类型
Table 3. Flora types of woody plants in the forest communities on Daishan Island
分布区类型 分布区类型 科数 属数 种数 世界分布 1 广布 5 0 0 热带成分
(2~7)2 泛热带 6 10 11 3 热带亚洲和热带美洲间断分布 2 1 1 4 旧世界热带分布 0 3 3 5 热带亚洲和热带大洋洲分布 0 2 2 6 热带亚洲至热带非洲分布 0 0 0 7 热带亚洲(印度、马来西亚)分布 1 2 2 温带成分
(8~14)8 北温带分布 4 4 4 9 东亚和北美间断分布 0 0 0 10 旧世界温带分布 0 1 1 11 温带亚洲分布 0 0 0 12 地中海、西至中亚分布 0 0 0 13 中亚分布 0 0 0 14 东亚分布(东喜马拉雅—日本) 0 3 3 中国特有 15 中国特有分布 0 2 2 -
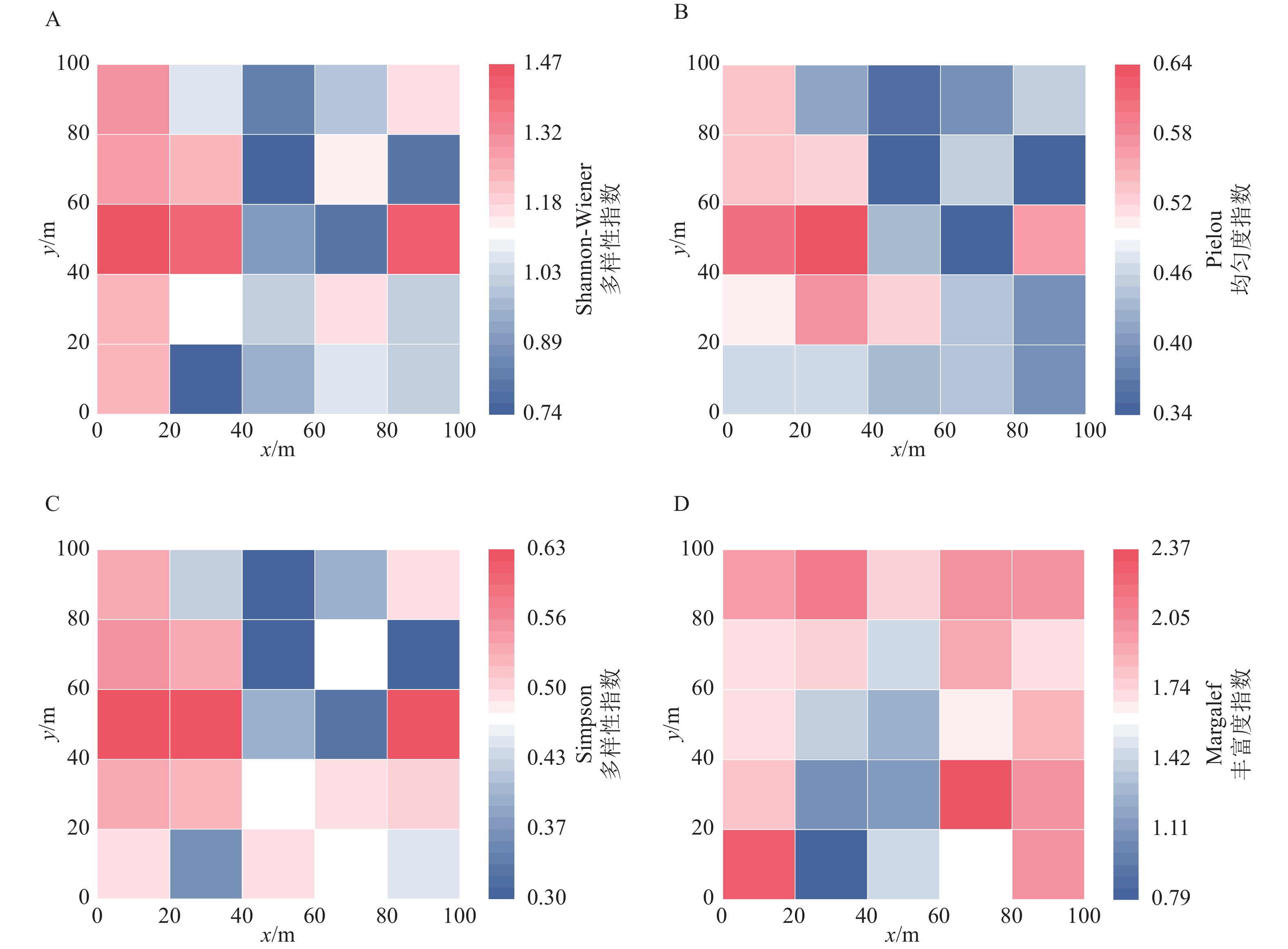
Shannon-Wiener多样性指数反映群落的物种丰富度和均匀度,数值越高多样性越高。1 hm2样地群落Shannon-Wiener多样性指数为1.28。从图3A可以看出:在20 m×20 m尺度上25个样方Shannon-Wiener多样性指数平均值为1.09;样地西侧相对较高,最高的1.47与最低0.74相比,相差98.65%。表明在样地空间上Shannon-Wiener多样性指数异质性很大,即使是相邻的2个样方都可能有很大差异。
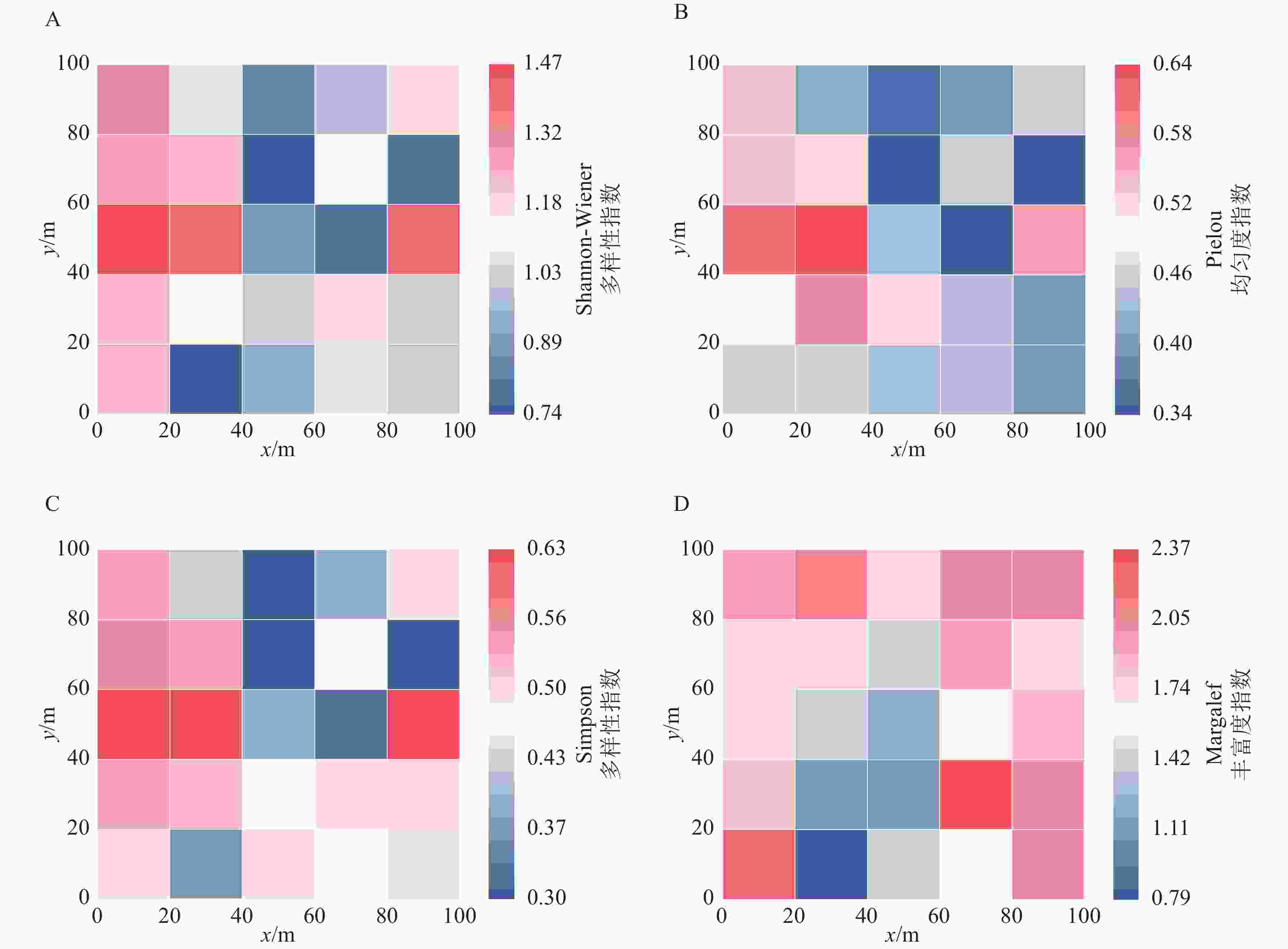
图 3 岱山岛群落物种多样性指数
Figure 3. Species diversity index of the Daishan Island community
Pielou均匀度指数衡量群落中物种个体分布的均匀程度,数值越接近1,物种分布越均匀。1 hm2样地群落Pielou均匀度指数为0.38,表明物种分布不均匀,少数物种占据主导地位。从图3B可以看出:在20 m×20 m尺度上25个样方Pielou均匀度指数平均值为0.46,最低为0.34,最高为0.64,同样说明样方之间物种分布的均匀性差异较大。
Simpson多样性指数反映优势种对群落的主导性,侧重常见物种的贡献。1 hm2样地群落Simpson多样性指数为0.50。图3C表明:在20 m×20 m尺度上25个样方Simpson多样性指数平均值为0.47,最低0.30,最高0.63,说明不同小尺度样方之间的多样性差异较大,但整体上多样性水平不高。从图3B和图3C可以看出:Simpson多样性指数与Pielou均匀度指数在整体上具有一致性。Margalef指数表征群落的物种丰富度,数值越大,表明单位个体数对应的物种数越多。1 hm2样地群落Margalef指数为3.08,在20 m×20 m尺度上25个样方Margalef指数平均值为1.72,最低0.79,最高2.37,在1204号样方最高(图3D)。
-
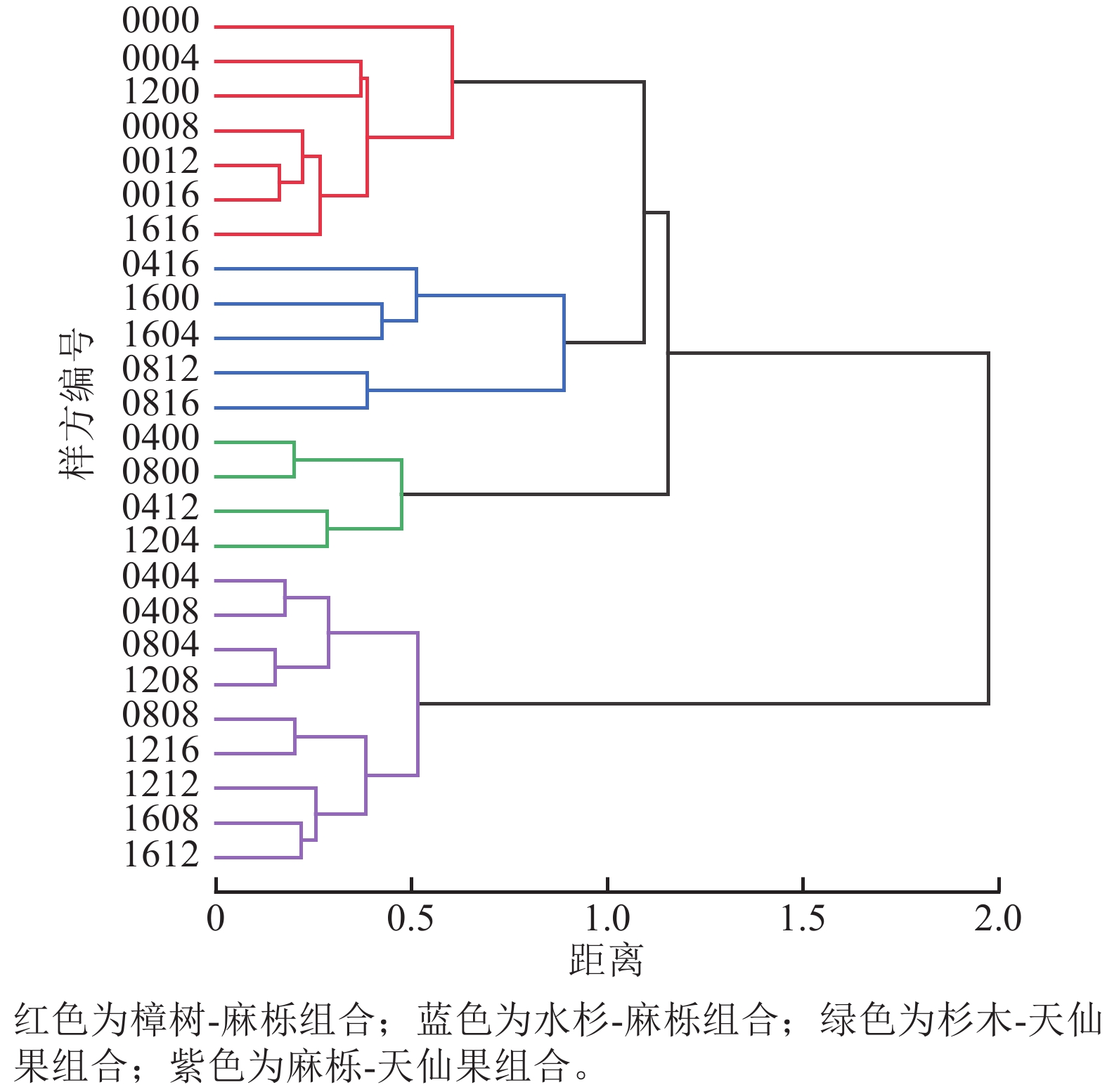
样地群落Pielou均匀度指数为0.38,表明物种分布不均匀。为了进一步了解群落树种组合类型,运用TWINSPAN分析,对25个样方群落上层树种组合进行分类,划分出4个树种组合(图4)。第1类为樟树-麻栎组合,共7个样方,主要组成树种为樟树、麻栎和杉木,集中分布在样地西侧。第2类为水杉-麻栎组合,共5个样方,主要组成树种为水杉、麻栎、算盘子,集中分布在样地北侧。第3类为杉木-天仙果组合,共4个样方,主要组成树种为杉木、天仙果、麻栎,集中分布在样地南侧。第4类为麻栎-天仙果组合,共9个样方,主要组成树种为麻栎、天仙果、杉木,集中分布在样地东侧。对4种组合进行ANOSIM分析,结果显示R=0.526>0,说明4种组合间差异大于组内差异;P=0.001<0.01,表明组合间差异极显著。
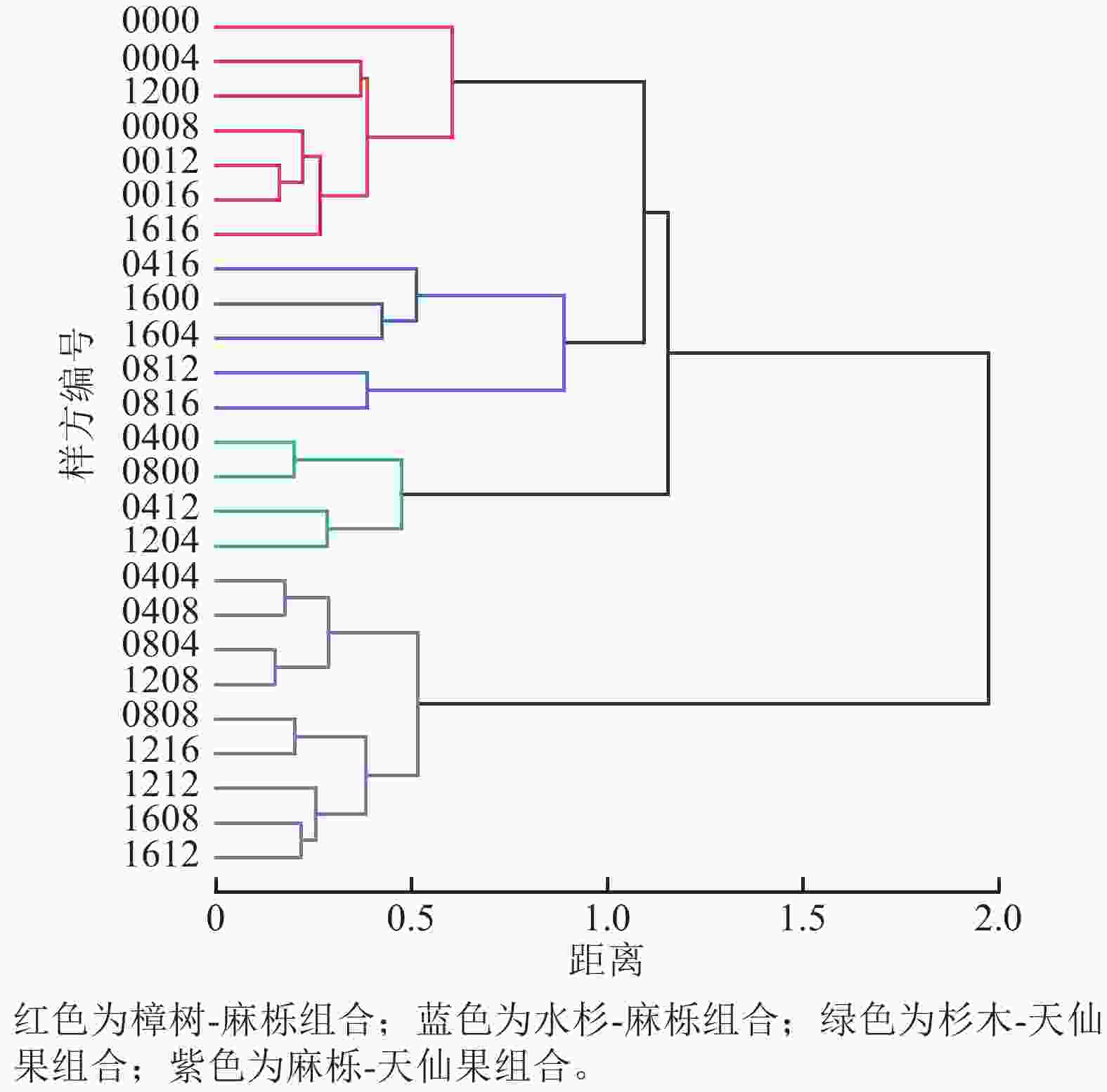
图 4 群落上层树种组合
Figure 4. Combination types of upper tree species in the community
4种树种组合林分指标和多样性指数(表4)表明:麻栎-天仙果组合的个体数量最多,但树木多度从大到小依次为杉木-天仙果组合、麻栎-天仙果组合、樟树-麻栎组合、水杉-麻栎组合。麻栎-天仙果组合的平均胸径、平均树高在4个组合中最高,其次是水杉-麻栎组合。物种多样性指数均以水杉-麻栎组合最高,说明水杉-麻栎组合中存在许多不同的物种,且各个物种的个体数量相对均衡分布。
表 4 岱山岛群落上层树种组合林分指标和多样性指数
Table 4. Stand indicators and diversity indices of dominant tree species combinations in Daishan Island forest community
树种组合 个体数/株 多度/(株·hm−2) 平均胸径/cm 平均树高/m Shannon-Wiener
多样性指数Pielou均匀
度指数Simpson多样
性指数Margalef
丰富度指数整体 783 783 14.80±7.71 8.64±2.46 2.17 0.78 0.85 2.25 樟树-麻栎 196 700 14.37±5.88 7.92±1.60 2.11 0.80 0.84 2.46 水杉-麻栎 90 450 14.93±8.30 8.86±2.75 2.23 0.87 0.87 2.67 杉木-天仙果 189 1 181 13.76±6.94 8.54±2.13 1.66 0.65 0.69 2.29 麻栎-天仙果 308 856 15.68±8.87 9.09±2.87 1.95 0.74 0.79 2.27 -
调查结果显示:麻栎、樟树和天仙果等是岱山岛森林群落的主要优势树种。麻栎、樟树是常见的人工造林树种,在浙江海岛适地适树技术研究[16]中被推荐。根据历史资料记载,岱山自20世纪80年代在全民义务植树活动中开始引种麻栎、樟树。麻栎根系发达、树冠疏透,可有效抵御海风,是一种适应性极强的树种,能够在多种环境中生长[17]。同时,麻栎与松类混交时,可通过根系分泌物质改善土壤结构,促进共生真菌活动,增强群落稳定性[18]。在杨佰润等[5]对大陈岛植物群落的研究中,天仙果也被记录为该岛的主要优势物种,与柃木等矮小乔木组成的群落,具有绝对的竞争力且物种数量明显较多。因此,岱山岛森林群落为通过人工种植促进恢复的次生林。
岱山岛森林群落树种径级结构呈现倒“J”型分布,说明群落树种更新能力较强,也反映了树种对光照、土壤、水分等资源的竞争强度相对较高。种群竞争导致少数个体存活并发育成大径级个体,大多数植株在竞争过程中被排斥。与杨佰润等[5]对大陈岛植物群落的研究结果相似。从常见种的径级结构比较来看,由于枸骨、天仙果属于小乔木,往往受到大乔木的强烈竞争,限制了其生长和径级结构的发育,个体的胸径集中于1 cm≤DBH<5 cm,因此大径级个体数相对较少。
岱山岛位于30°N,在科水平上,世界分布占27.78%,热带成分占50.00%,温带成分占22.22%。这与黄继红等[19]的研究结果相似:在30°N左右的海岛植被区系中,热带成分通常占主导地位,世界分布科的比例较高,而温带成分较少。
海岛植物多样性受到多种因素的综合影响,其中面积、土壤养分和气候条件是最主要的因素[20],除自然因素外,人类活动也会对海岛植物多样性产生影响[21]。研究显示:岱山岛作为相对较小且具有一定隔离性的海岛,物种迁入和扩散能力有限,导致物种分布不均匀[22],不同小尺度样方间的多样性差异较大,样地内部的生态环境存在较大异质性,类似结果在福建海岛[23]和庙岛群岛[24]研究中均有发现。
岱山岛森林群落上层以樟树、麻栎、水杉、女贞为优势树种的组合更适宜作为海岛群落修复的造林树种。樟树拥有由强健主根、多级侧根及密集须根组成的根系网络系统,为主干提供稳固的支撑,使其能够抵抗较强的风力和不良环境条件。樟树能够在不同的环境条件下利用多种资源,减少与其他树种的生态位重叠,缓解了竞争压力,增加了自身在群落中的稳定性和优势地位。麻栎作为一种适应性强的树种,具有较深的根系,且侧根发达,能有效锚固沙质土壤,抵御台风侵袭[25]。此外,麻栎对土壤要求低,能够适应海岛季节性的干旱条件,其落叶还能增加土壤有机质,改善贫瘠土壤。同时,麻栎的果实为小型哺乳动物和鸟类提供食物,有利于食物链的稳定[26]。在嵊泗县麻栎被列为海岛森林抚育补植的推荐树种之一[27]。水杉为速生落叶乔木,兼具强耐寒性、耐水淹及抗旱性,树形挺拔,叶色季相变化显著,具有较高的观赏价值。常被选作植被恢复树种,种植于海岛潮间带、河口湿地等区域,可以有效防止水土流失,同时为其他动物提供栖息地[28]。杉木作为该群落的常见种之一,拥有较多的个体数,但个体多集中在小径级,且存在显著的株高生长抑制、冠幅发育不良,因此不适宜作为岱山岛生态修复树种。
岱山岛森林群落下层以天仙果、细齿叶柃、算盘子、枸骨为优势树种的组合更适宜作为海岛群落修复的造林树种。天仙果作为桑科Moraceae 榕属Ficus植物,具有较为发达的根系,能深入土壤,更好地吸收水分和养分[29],增强对土壤的固定作用,使其在海岛多风、土壤条件复杂的环境中不易被吹倒,能稳定生长。同时天仙果在生态系统中占据着较为广泛的生态位,能与多种植物和动物相互作用。细齿叶柃凭借广域生态幅和低光补偿点的耐荫机制[30],可以在群落下层实现种群持续,稳定占据空间生态位。细齿叶柃细锯齿形态的叶片,通过调控叶脉网络拓扑结构,降低叶片边界层厚度,促进水气交换,提高水分运输效率。此外,通过增强叶肉导度,在低温胁迫期提高光合、蒸腾速率,促进碳吸收,弥补低温对光合作用的影响,使细齿叶柃在不同气候条件下均能发挥良好的生态功能。反季节花物候有利于促进生态系统的物质循环和能量流动,维持生态平衡。算盘子是一种耐盐碱灌木,其革质化叶片可通过减少气孔密度和表皮渗透性,有效降低盐雾环境下盐分的吸收与蒸腾损伤,从而适应海岛边缘高盐、干旱的胁迫条件。这种植物的匍匐生长特性可以快速覆盖地表,有效减少水土流失,尤其在陡坡或侵蚀严重区域表现出显著的效果。此外,算盘子的果实不仅可以入药,还可以作为染料的原料,在实现生态修复的同时还可为当地居民带来经济价值。
本研究主要对岱山岛森林群落树种的组成和结构进行了系统研究,受制于野外调查时间窗口的局限性,加之由于木本植物密度很高,林下草本植物稀少,未对草本层和层间植物进行专项调查。未来应选择合适季节对草本层和层间植物进行补充调查,掌握草本层和层间植物的分布特征、物种组成,更加全面准确地反映岱山岛森林群落组成与结构。
Structure and composition of typical forest communities on Daishan Island in Zhejiang Province
-
摘要:
目的 探索浙江省岱山岛森林群落树种的组成和结构,为海岛森林群落的修复提供树种组合模式参考。 方法 利用样方调查法,调查样地(100 m×100 m)内胸径≥1 cm的木本植物,分析其树种组成、区系成分、径级结构、物种多样性、树种组合等特征。 结果 岱山岛样地内共有木本植物8 767株,隶属18科28属29种,平均树高为4.11 m,平均胸径为4.35 cm。麻栎Quercus acutissima、樟树Cinnamomum camphora和天仙果Ficus erecta等为该群落的主要优势树种。在科水平上,群落树种的世界分布占27.78%,热带成分占50.00%,温带成分占22.22%;在属水平上,热带成分占64.29%,温带成分占28.57%,中国特有占7.14%。样地树种径级结构呈倒“J”型分布,小径级和较小径级的植物数量占比达91.80%。群落Shannon-Wiener多样性指数、Pielou均匀度指数、Simpson多样性指数和Margalef丰富度指数分别为1.28、0.38、0.50、3.08。群落有樟树-麻栎、水杉Metasequoia glyptostroboides-麻栎、麻栎-天仙果等具有较好恢复力的树种组合。 结论 岱山岛森林群落以麻栎、樟树、天仙果、细齿叶柃Eurya nitida、枸骨Ilex cornuta等为优势树种,径级结构呈现倒“J”型分布。该群落植物区系以热带成分占主导,群落整体上树种分布不均匀,形成了不同的树种组合。图4表4参30 Abstract:Objective This study aims to investigate the species composition and community structure of forest stands on Daishan Island, Zhejiang Province, and provide recommendations for tree species combination models for the restoration of forest communities on islands. Method A quadrat survey was conducted to investigate woody plants with a diameter at breast height (DBH) ≥ 1 cm within the sample plots (100 m×100 m). Species composition, floristic elements, DBH class distribution, species diversity, and tree species combination were analyzed. Result A total of 8 767 woody plant individuals were recorded, belonging to 29 species, 28 genera, and 18 families, with an average tree height of 4.11 m and an average DBH of 4.35 cm. Quercus acutissima, Cinnamomum camphora, and Ficus erecta were the dominant species in the community. At the family level, the flora consisted of 27.78% cosmopolitan, 50.00% tropical, and 22.22% temperate components. At the genus level, 64.29% were tropical, 28.57% temperate, and 7.14% unique to China. The DBH of tree species in the community showed an inverted “J” shape distribution, with 91.80% of plants in the small and relatively small diameter classes. The Shannon-Wiener diversity index, Pielou evenness index, Simpson diversity index, and Margalef richness index were 1.28, 0.38, 0.50, and 3.08, respectively. The community included tree species combinations with good resilience, such as C. camphora-Q. acutissima, Metasequoia glyptostroboides-Q. acutissima, and Q. acutissima-F. erecta. Conclusion The forest community on Daishan Island is dominated by Q. acutissima, C. camphora, F. erecta, Eurya nitida, and Ilex cornuta. The DBH structure displays an inverted “J” shape pattern, and the plant community in this area is dominated by tropical species. The overall distribution of tree species in the community is uneven, resulting in different combinations of tree species. [Ch, 4 fig. 4 tab. 30 ref.] -
Key words:
- species composition /
- community structure /
- tree species combination /
- Daishan Island
-
图 2 岱山岛群落常见种径级分布
Figure 2 Diameter class distribution of common species in Daishan Island community
表 1 群落重要值排名前10树种统计
Table 1. Statistics of top 10 tree species with important value of community
树种 多度/
(株·hm−2)平均胸径/cm 平均树高/m 重要值/% 树种 多度/
(株·hm−2)平均胸径/cm 平均树高/m 重要值/% 天仙果 6 103 2.87±1.72 3.55±1.19 32.53 枸骨 221 2.32±1.16 3.23±0.92 3.69 麻栎 244 17.01±10.56 8.89±3.62 12.94 黄檀 220 5.03±2.85 5.22±1.42 3.23 杉木 618 9.48±3.93 5.43±1.85 11.37 算盘子 74 8.27±3.28 5.73±1.35 2.87 细齿叶柃 764 4.38±2.63 4.22±1.31 7.62 水杉 44 18.93±7.60 12.13±3.21 2.70 樟树 146 14.45±8.77 7.17±3.01 7.12 女贞 71 8.51±5.86 6.69±2.35 2.68 说明:天仙果Ficus erecta,麻栎Quercus acutissima,杉木Cunninghamia lanceolata,细齿叶柃Eurya nitida,樟树Cinnamomum camphora,枸骨Ilex cornuta,黄檀Dalbergia hupeana,算盘子Glochidion puberum,水杉Metasequoia glyptostroboides,女贞Ligustrum lucidum。  下载: 导出CSV
下载: 导出CSV
表 2 岱山岛森林群落垂直结构
Table 2. Vertical stratification of the forest community on Daishan Island
林层 主要树种 最低树高/m 最高树高/m 平均树高/m 平均胸径/cm 多度/(株·hm−2) 重要值/% 群落上层 麻栎 6.50 17.00 10.55±2.77 21.46±8.87 176 25.42 杉木 6.50 11.00 7.83±1.08 13.13±3.29 174 15.72 樟树 6.50 15.50 9.07±2.69 19.10±8.17 81 12.22 天仙果 6.50 11.00 7.02±0.71 8.73±3.00 122 10.22 水杉 6.50 18.00 12.13±3.21 18.93±7.60 44 7.24 群落下层 天仙果 1.00 6.00 3.48±1.09 2.75±1.46 5981 42.95 杉木 1.60 6.00 4.48±1.08 8.04±3.16 444 13.57 细齿叶柃 1.50 6.00 4.11±1.23 4.19±2.46 735 10.78 枸骨 1.80 5.50 3.22±0.89 2.31±1.16 220 4.47 樟树 2.00 6.00 4.81±1.17 8.65±5.42 65 4.36
下载: 导出CSV
表 3 岱山岛群落木本植物区系类型
Table 3. Flora types of woody plants in the forest communities on Daishan Island
分布区类型 分布区类型 科数 属数 种数 世界分布 1 广布 5 0 0 热带成分
(2~7)2 泛热带 6 10 11 3 热带亚洲和热带美洲间断分布 2 1 1 4 旧世界热带分布 0 3 3 5 热带亚洲和热带大洋洲分布 0 2 2 6 热带亚洲至热带非洲分布 0 0 0 7 热带亚洲(印度、马来西亚)分布 1 2 2 温带成分
(8~14)8 北温带分布 4 4 4 9 东亚和北美间断分布 0 0 0 10 旧世界温带分布 0 1 1 11 温带亚洲分布 0 0 0 12 地中海、西至中亚分布 0 0 0 13 中亚分布 0 0 0 14 东亚分布(东喜马拉雅—日本) 0 3 3 中国特有 15 中国特有分布 0 2 2
下载: 导出CSV
表 4 岱山岛群落上层树种组合林分指标和多样性指数
Table 4. Stand indicators and diversity indices of dominant tree species combinations in Daishan Island forest community
树种组合 个体数/株 多度/(株·hm−2) 平均胸径/cm 平均树高/m Shannon-Wiener
多样性指数Pielou均匀
度指数Simpson多样
性指数Margalef
丰富度指数整体 783 783 14.80±7.71 8.64±2.46 2.17 0.78 0.85 2.25 樟树-麻栎 196 700 14.37±5.88 7.92±1.60 2.11 0.80 0.84 2.46 水杉-麻栎 90 450 14.93±8.30 8.86±2.75 2.23 0.87 0.87 2.67 杉木-天仙果 189 1 181 13.76±6.94 8.54±2.13 1.66 0.65 0.69 2.29 麻栎-天仙果 308 856 15.68±8.87 9.09±2.87 1.95 0.74 0.79 2.27
下载: 导出CSV
-
[1] NOGUÉ S, de NASCIMENTO L, FERNÁNDEZ-PALACIOS J M, et al. The ancient forests of La Gomera, Canary Islands, and their sensitivity to environmental change[J]. Journal of Ecology, 2013, 101(2): 368−377. DOI: 10.1111/1365-2745.12051. [2] 姚兰, 艾训儒, 易咏梅, 等. 鄂西南亚热带常绿落叶阔叶混交林优势种群的结构与动态特征[J]. 林业科学, 2017, 53(2): 10−18. YAO Lan, AI Xunru, YI Yongmei, et al. Structure and dynamics of dominant populations in the mixed forest of subtropical evergreen and deciduous broad-leaved tree species in the southwest of Hubei Province[J]. Scientia Silvae Sinicae, 2017, 53(2): 10−18. DOI: 10.11707/j.1001-7488.20170202. [3] 池源, 石洪华, 郭振, 等. 海岛生态脆弱性的内涵、特征及成因探析[J]. 海洋学报, 2015, 37(12): 93−105. CHI Yuan, SHI Honghua, GUO Zhen, et al. Connotation, features and causes of island ecological vulnerability[J]. Haiyang Xuebao, 2015, 37(12): 93−105. DOI: 10.3969/j.issn.0253-4193.2015.12.010. [4] 郑丽婷, 苏田, 刘翔宇, 等. 庙岛群岛典型植物群落物种、功能、结构多样性及其对环境因子的响应[J]. 应用生态学报, 2018, 29(2): 343−351. ZHENG Liting, SU Tian, LIU Xiangyu, et al. Species, functional, structural diversity of typical plant communities and their responses to environmental factors in Miao Archipelago, China[J]. Chinese Journal of Applied Ecology, 2018, 29(2): 343−351. DOI: 10.13287/j.1001-9332.201802.037. [5] 杨佰润, 吴江, 官凯程, 等. 大陈岛植物群落结构及物种组成[J]. 浙江农林大学学报, 2025, 42(2): 321−328. YANG Bairun, WU Jiang, GUAN Kaicheng, et al. Community structure and species composition in Dachen Island[J]. Journal of Zhejiang A&F University, 2025, 42(2): 321−328. DOI: 10.11833/j.issn.2095-0756.20240423. [6] 周厚诚, 任海, 彭少麟. 广东南澳岛次生林的群落结构分析[J]. 广西植物, 2001, 21(3): 209−214, 251. ZHOU Houcheng, REN Hai, PENG Shaolin. Community structure of three secondary forests in Nan’ao Island, Guangdong[J]. Guihaia, 2001, 21(3): 209−214, 251. DOI: 10.3969/j.issn.1000-3142.2001.03.005. [7] YANG Chengfeng, ZHAO Jingyan, HUANG Shengzhuo, et al. Distribution of different plant life forms on tropical islands: patterns and underlying mechanisms[J]. Frontiers in Plant Science, 2025, 16: 1566156. DOI: 10.3389/fpls.2025.1566156. [8] 郑枭, 王通, 庞春梅, 等. 气候变化背景下天目山落叶阔叶林乔木层1996—2017年动态研究[J]. 浙江农林大学学报, 2023, 40(6): 1250−1260. ZHENG Xiao, WANG Tong, PANG Chunmei, et al. Dynamic study on tree layer of deciduous broad-leaved forest in Mount Tianmu from 1996 to 2017 under climate change[J]. Journal of Zhejiang A&F University, 2023, 40(6): 1250−1260. DOI: 10.11833/j.issn.2095-0756.20230185. [9] 楼一恺, 范忆, 戴其林, 等. 天目山常绿落叶阔叶林群落垂直结构与群落整体物种多样性的关系[J]. 生态学报, 2021, 41(21): 8568−8577. LOU Yikai, FAN Yi, DAI Qilin, et al. Relationship between vertical structure and overall species diversity in an evergreen deciduous broad-leaved forest community of Tianmu Mountain Natural Reserve[J]. Acta Ecologica Sinica, 2021, 41(21): 8568−8577. DOI: 10.5846/stxb202007301989. [10] 许辉林, 付煜阳, 余树全, 等. 天目山常绿落叶阔叶林生产力及其影响因素[J]. 东北林业大学学报, 2024, 52(10): 40−46, 65. XU Huilin, FU Yuyang, YU Shuquan, et al. Productivity and influencing factors of evergreen and deciduous broad-leaved forests in Tianmu Mountain[J]. Journal of Northeast Forestry University, 2024, 52(10): 40−46, 65. DOI: 10.3969/j.issn.1000-5382.2024.10.006. [11] 刘璐, 刘志成, 杨瑞莹, 等. 永定河干涸段(卢梁段)河流廊道植物群落数量分类及多样性特征[J]. 生态学报, 2020, 40(24): 9129−9137. LIU Lu, LIU Zhicheng, YANG Ruiying, et al. The quantitative classification and diversity pattern of plant communities in the river corridor of the dry section of Yongding River (Lu-Liang section)[J]. Acta Ecologica Sinica, 2020, 40(24): 9129−9137. DOI: 10.5846/stxb202003050419. [12] CHAPMAN M G, UNDERWOOD A J. Ecological patterns in multivariate assemblages: information and interpretation of negative values in ANOSIM tests[J]. Marine Ecology Progress Series, 1999, 180: 257−265. DOI: 10.3354/meps180257. [13] 吴征镒. 中国植被[M]. 北京: 科学出版社, 1980. WU Zhengyi. Vegetation in China[M]. Beijing: Science Press, 1980. [14] 吴征镒. 《世界种子植物科的分布区类型系统》的修订[J]. 云南植物研究, 2003, 25(5): 535−538. WU Zhengyi. Revision of the distribution type system of the world seed plants family[J]. Acta Botanica Yunnanica, 2003, 25(5): 535−538. [15] 吴征镒, 孙航, 周浙昆, 等. 中国种子植物区系地理[M]. 北京: 科学出版社, 2011. WU Zhengyi, SUN Hang, ZHOU Zhekun, et al. Floristics of Seed Plants from China[M]. Beijing: Science Press, 2011. [16] 陶吉兴. 浙江海岛适地适树技术研究[J]. 浙江林学院学报, 2003, 20(4): 346−352. TAO Jixing. A study of matching species with the site in island areas of Zhejiang[J]. Journal of Zhejiang Forestry College, 2003, 20(4): 346−352. [17] 朱文成, 李宗泰, 舒秀阁, 等. 不同坡向和播种季节对麻栎种子发芽率的影响[J]. 山东林业科技, 2019, 49(4): 54−55. ZHU Wencheng, LI Zongtai, SHU Xiuge, et al. Effects of different slope aspect and sowing season on seed germination rate of Quercus acutissima[J]. Journal of Shandong Forestry Science and Technology, 2019, 49(4): 54−55. DOI: 10.3969/j.issn.1002-2724.2019.04.014. [18] 张强. 油松麻栎混交林土壤物理性质研究[J]. 山东林业科技, 2019, 49(6): 61−62, 67. ZHANG Qiang. Study on soil physical properties of mixed pine oak forest[J]. Journal of Shandong Forestry Science and Technology, 2019, 49(6): 61−62, 67. DOI: 10.3969/j.issn.1002-2724.2019.06.017. [19] 黄继红, 马克平, 应俊生. 中国特有植物地理成分及其与气候因素关系[C]//马克平. 第九届全国生物多样性保护与持续利用研讨会论文集. 北京: 气象出版社, 2010: 97−108. HUANG Jihong, MA Keping, YING Junsheng. Phytogeographical patterns of genera of endemic seed plants and their relationship with climatic gradients in China[C]//MA Keping. Proceedings of the 9th National Conference on the Conservation and Sustainable Use of Biodiversity in China. Beijing: China Meteorological Press, 2010: 97−108. [20] LI Shengchun, TU Tieyao, LI Shaopeng, et al. Different mechanisms underlie similar species-area relationships in two tropical archipelagoes[J]. Plant Diversity, 2024, 46(2): 238−246. DOI: 10.1016/j.pld.2023.08.006. [21] 王鸣雷, 邱思妍, 史文娇. 珠三角地区至2050年土地利用变化对生物多样性的影响[J]. 生态学报, 2024, 44(16): 7183−7197. WANG Minglei, QIU Siyan, SHI Wenjiao. Impacts of future land use change on biodiversity in the Pearl River Delta[J]. Acta Ecologica Sinica, 2024, 44(16): 7183−7197. DOI: 10.20103/j.stxb.202312182744. [22] 张新阳, 池源, 刘大海, 等. 基于“景观-植被-土壤” 框架的海岛生态系统遥感评价——以辽宁长海县为例[J]. 海洋科学进展, 2025, 43(1): 231−246. ZHANG Xinyang, CHI Yuan, LIU Dahai, et al. Remote sensing-based island ecological evaluation under the framework of “landscape-vegetation-soil”: Changhai (a county in Liaoning Province, China) case study[J]. Advances in Marine Science, 2025, 43(1): 231−246. DOI: 10.12362/j.issn.1671-6647.20231022001. [23] 谢艳秋, 黄晖, 王春晓, 等. 福建海岛滨海特有植物种–面积关系及物种丰富度决定因素[J]. 生物多样性, 2023, 31(5): 5−14. XIE Yanqiu, HUANG Hui, WANG Chunxiao, et al. Determinants of species–area relationship and species richness of coastal endemic plants in the Fujian islands[J]. Biodiversity Science, 2023, 31(5): 5−14. DOI: 10.17520/biods.2022345. [24] 刘聪, 骆杨青, 闫昱晶, 等. 岛屿生物多样性的形成、维持与丧失[J]. 遗传, 2025, 47(1): 101−132. LIU Cong, LUO Yangqing, YAN Yujing, et al. The formation, maintenance, and loss of island biodiversity[J]. Hereditas (Beijing), 2025, 47(1): 101−132. DOI: 10.16288/j.yczz.24-223. [25] 王霞, 胡海波, 张世豪, 等. 不同林龄麻栎林地下部分生物量与碳储量研究[J]. 生态学报, 2019, 39(22): 8556−8564. WANG Xia, HU Haibo, ZHANG Shihao, et al. Underground biomass and carbon storage for Quercus acutissima forests of different ages[J]. Acta Ecologica Sinica, 2019, 39(22): 8556−8564. DOI: 10.5846/stxb201809141994. [26] 方炎杰, 金祖达, 周天焕, 等. 以嵊泗县为例谈海岛森林抚育补植技术和树种选择[J]. 防护林科技, 2014(11): 71−73, 93. FANG Yanjie, JIN Zuda, ZHOU Tianhuan, et al. Taking Shengsi County as an example to talk about island forest tending and replanting technology and tree species selection[J]. Protection Forest Science and Technology, 2014(11): 71−73, 93. DOI: 10.13601/j.issn.1005-5215.2014.11.031. [27] 袁学勤. 宣州区南漪湖入湖河口湿地生态修复对策研究[J]. 乡村科技, 2022, 13(5): 147−150. YUAN Xueqin. Study on ecological restoration countermeasures of Nanyi Lake estuary wetland in Xuanzhou District[J]. Rural Science and Technoloty, 2022, 13(5): 147−150. DOI: 10.3969/j.issn.1674-7909.2022.05.041. [28] ZHANG Zhengren, YANG Xue, LI Weiying, et al. Comparative chloroplast genome analysis of Ficus (Moraceae): insight into adaptive evolution and mutational hotspot regions[J]. Frontiers in Plant Science, 2022, 13: 965335. DOI: 10.3389/fpls.2022.965335. [29] 王燚, 包志毅. 我国柃属植物种质资源及其园林应用前景[J]. 林业科学, 2007, 43(8): 118−122. WANG Yi, BAO Zhiyi. Wild plant resources and landscape application prospects of Eurya in China[J]. Scientia Silvae Sinicae, 2007, 43(8): 118−122. DOI: 10.3321/j.issn:1001-7488.2007.08.020. [30] 童冉, 周本智, 姜丽娜, 等. 我国杉木人工林可持续经营面临的问题及发展策略——基于全国分布区的调查[J]. 世界林业研究, 2019, 32(2): 90−96. TONG Ran, ZHOU Benzhi, JIANG Li’na, et al. Problems and development strategy for sustainable management of Chinese fir plantation: based on the investigation of its national distribution[J]. World Forestry Research, 2019, 32(2): 90−96. DOI: 10.13348/j.cnki.sjlyyj.2018.0106.y. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250196
 点击查看大图
点击查看大图
计量
- 文章访问数: 803
- HTML全文浏览量: 282
- PDF下载量: 157
- 被引次数: 0
