






-
光照是影响植物生长的重要因素,光合作用通过影响植物的新陈代谢和次生代谢物的合成,调控植物的产量和活性成分积累[1−2]。通常,过强的光照会使叶绿体的光合反应中心失活或受损,引起光抑制,并可能抑制植物的生长。但长期的弱光环境可能会减少光能的吸收,并通过影响净光合速率来抑制植物的生长发育和代谢产物积累[3]。由此,适度光强可以促进光合作用,激活次生代谢产物的合成和积累过程,保障药用植物的正常生长和有效成分积累[4]。植物对光强的偏好性因物种而异,“光照-寒热药性”假说认为:延胡索Corydalis yanhusuo等热性中药偏好弱光环境[5]。郁闭度是影响林地光照强度的决定性因子,研究药用植物生长和有效成分含量对林冠郁闭度的响应特征,已成为林下药用植物种植管理和提质增效的重要问题。
凋落物分解是联系植物-土壤系统物质循环和能量流动的纽带[6],为林下药用植物的生长发育提供所需养分。研究表明:厚度过高的凋落物层由于遮荫、物理阻碍等可能对药用植物生长有负面效应,而适量的凋落物覆盖则有助于药用植物的生长[7]。目前,凋落物对药用植物生长的作用难以界定,它随着森林类型、凋落物量和药用植物的种类以及微生境的特征不同而存在差异[8]。延胡索名列“浙八味”道地药材之一,其块茎中含有的延胡索乙素在止痛、镇静和催眠等方面应用广泛。延胡索在浙江省栽培历史悠久,大盘山保护区内的杉木Cunninghamia lanceolata-延胡索群落是国内仅有的野生延胡索成片分布区域[9]。杉木是中国亚热带人工林的主要造林树种之一[10],其凋落物不易分解,纯林连栽后土壤中有效养分呈下降趋势,但亦有研究证实去除杉木凋落物会降低土壤养分含量[11]。然而,杉木林中凋落物输入如何影响林下延胡索的产量和有效成分却鲜有报道。
延胡索只在母球茎下生根,不发达的根系吸收水分较少,难以应对过强光照产生的蒸腾作用[12],过弱光照又不能满足生长所需要的能量,从而决定了延胡索对生长环境的要求较高[13]。但近年来,由于植被的过度保护,造成自然保护区内杉木林郁闭度较高,引起林内光照不足,以及林下凋落物层积累较厚等问题,这已严重阻碍了林下延胡索植株的萌发生长和开花结实。有研究表明:适度的遮荫和凋落物输入可满足野生药用植物的光照和养分需求[14−15]。然而,如何通过调节郁闭度和凋落物等改善林间光照和养分资源供应,促进林下药用植物产量和有效成分含量提升尚无研究报道。因此,本研究以杉木林林下种植的延胡索为研究对象,通过调控林冠郁闭度和移除凋落物等方式,研究营林措施对林下植株生物量分配、根区土壤养分特征以及延胡索块茎产量和延胡索乙素质量分数的影响,以期为块茎类药用植物的栽培提供理论支撑。
-
研究样地位于金华市磐安县大盘山国家级自然保护区(28°57′N,120°28′E),该区属亚热带季风气候,高温和降水主要分布于夏季,年均气温为17.3 ℃,年均降水量为1 116.0 mm,年日照时数为1 822.8 h,海拔为580.6~712.5 m。保护区乔木层主要为人工栽培杉木,株行距为2 m × 1 m,林龄均为30~40 a。保护区内有许多珍贵的植物和道地药材,由于保护区禁止砍伐,加上杉木林纯化和多代连栽后,造成林冠郁闭度较高、透光性弱,同时较厚的凋落物层致使林下药用植物数量和长势逐年递减。
-
2020年10月,依据坡向、坡度和海拔相近,母岩相同的原则,在大盘山国家级自然保护区内选取3个试验位点(白雨伞、大岩后和毛坞孔)。研究区3个试验位点初始郁闭度与土壤理化性质分别为:郁闭度0.9、0.8和0.9;pH 5.25、4.93和5.09;有机碳31.20、47.10和33.10 g·kg−1;全氮1.92、3.32和2.55 g·kg−1;可溶性有机碳23.90、23.90和24.70 mg·kg−1;可溶性有机氮18.80、23.20和20.70 mg·kg−1。每个位点分别选取3个20 m×20 m的标准样地,各样地间隔大于10 m,每个样地内设置6个小区,共54个试验小区。标准样地通过疏伐林冠来建立3个郁闭度的固定样地。
本研究为双因素裂区试验,郁闭度处理为主因素,凋落物处理为副因素。每个处理组合设置3个1 m×4 m的条带状小区,在小区内进行种植。每年播种前进行林冠疏伐以确保达到以下试验要求:高郁闭度0.7 (光强≤65 W·m−2)、中郁闭度0.5 (光强为65~190 W·m−2)、低郁闭度0.3 (光强≥190 W·m−2)。郁闭度采用抬头法和鱼眼相机[16]相结合的方法确定。本研究共设置3个位点×3个郁闭度×2种凋落物处理×3个重复。
2021年11月,选择长势相似、大小均匀的延胡索新鲜块茎进行播种,株行距为10 cm×25 cm。整个试验期间保持野生生境,不进行水肥管理。在开花期(2022年3月)进行净光合速率、植株生物量、根生物量等生长指标测定;同时,去除小区内地表覆盖的凋落物,在延胡索种植小区采用五点取样法用土钻采集0~20 cm土层的延胡索根区土壤,每个处理3个重复,带回实验室后去除杂质,并过2 mm筛,在实验室经自然风干后,一部分土壤用于理化性质测定,另一部分土壤于−80 ℃储藏,用于相关微生物丰度测定。收获时(2022年5月)在每个试验小区选取2个1 m×1 m区域,挖出全部块茎进行延胡索产量和延胡索乙素质量分数的测定。
-
土壤有机碳(SOC)采用重铬酸钾容量法-稀释热法测定。全氮(TN)采用凯氏定氮仪-蒸馏法测定。全磷(TP)采用硫酸-高氯酸消煮-钼锑抗比色法测定。速效磷(AVP)采用碳酸氢钠法测定。可溶性碳(DOC)和可溶性氮(DON)使用总有机碳分析仪(TOC)测定。土壤理化性质测定方法参照《土壤农化分析》[17]。
-
每个处理选取生长均匀的延胡索植株3株,将其完整取出。摘除植株块茎,保留其余部分,用去离子水洗净,在烘箱中用105 ℃杀青30 min,75 ℃烘至恒量后称取质量,分别得到植株地上部分、根生物量。叶片净光合速率采用LI-6400XT便携式光合作用测量系统(LI-COR Inc., Lincoln)测定,选择晴天9:00—11:00测定,每株选取叶位一致、完全展开的3片功能叶片,每个处理测定3株植株,待每片叶片的净光合速率增减幅度小于0.5 μmol·m−2·s−1时,记录数值并取平均值,即为植株净光合速率。
-
产量测定用称量法,样品采收于2022年5月。对每个小区收获的延胡索块茎单独分装,用自来水冲洗3遍,再用去离子水清洗2遍,将洗净的块茎置于通风处自然阴干,至表面无明显水分,用电子天平称其鲜质量,记为每个小区产量,然后计算每个处理的产量。
延胡索块茎产量测定后,采用张静等[18]的高效液相色谱法(HPLC)进行有效成分延胡索乙素质量分数的测定。每个处理重复3次。
-
称取土壤样品0.5 g,按照PowerSoilTM总DNA提取试剂盒(Mobio公司)说明书提取土壤微生物总DNA;经质量体积比为1%的琼脂糖凝胶电泳检测,保存于−20 ℃冰箱备用[19]。采用实时荧光定量PCR (RT-qPCR)测定土壤细菌和真菌群落丰度,选择引物338F (ACTCCTACGGGAGGCAGCAG)和518R (ATTAC-CGCGGCTGCTGG)扩增16S rDNA,选择引物NS1 (GTAGTCATATGCTTGTCC)和Fung (CATTC-CCCGTTACCCGTTG)扩增18S rDNA。每个样本重复3次[20]。
-
将植株生物量、根生物量、净光合速率、块茎产量和延胡索乙素质量分数作为评价指标,根据式(1)计算雷达图的面积(Si),根据式(2)计算雷达图的周长(Li):
$$ {S}_{i}=\sum _{j=1}^{m}\left[\frac{1}{2}{N}_{ij}{N}_{i\left(j+1\right)}\mathrm{s}\mathrm{i}\mathrm{n}\mathrm{\alpha }\right]\text{;}$$ (1) $${L}_{i}=\sum _{j=1}^{m}\sqrt{{N}_{ij}^{2}+{N}_{i\left(j+1\right)}^{2}-{2N}_{ij}^{2}{N}_{i\left(j+1\right)}^{2}\mathrm{cos}\mathrm{\alpha }}。$$ (2) 式(1)~(2)中:m表示评价指标总数;Nij是第i个对象第j项评价指标;α为相邻射线轴之间的夹角。
根据式(3)计算评价指标的综合评价值(Y):
$$Y=\sqrt{\frac{{S}_{i}}{{S}_{\mathrm{m}\mathrm{a}\mathrm{x}}}\times \frac{{S}_{i}}{{{\text{π}} \left(\dfrac{{L}_{i}}{2{\text{π}} }\right)}^{2}}}。 $$ (3) 式(3)中:Smax表示雷达图的最大面积。
-
采用SPSS 19.0进行数据分析。采用双因素方差分析,解析郁闭度、凋落物及两者交互作用对植株和土壤指标的效应。利用单因素方差分析对同一郁闭度条件下不同凋落物处理的影响进行Duncan多重比较,显著性水平为0.05。采用线性回归与逐步回归分析方法解析影响延胡索块茎产量和延胡索乙素质量分数的主要因素。用GraphPad PRISM®软件进行绘图。数据结果采用平均值±标准差表示。
-
无论覆盖或去除凋落物,降低郁闭度均提高了延胡索生物量(图1)。在覆盖凋落物时,3个试验位点的植株生物量在低郁闭度处理中分别较中郁闭度和高郁闭度处理平均提高了67.3%和109.0%;在去除凋落物时,低郁闭度处理的植株生物量分别较中郁闭度和高郁闭度处理平均提高了59.0%和137.0%。此外,根生物量和净光合速率也表现出相似的趋势。

图 1 不同郁闭度和凋落物处理对延胡索植株生长的影响
Figure 1. Effects of different canopy density and litter treatment on plant growth of C. yanhusuo
郁闭度、凋落物显著影响了延胡索的块茎产量和延胡索乙素质量分数(表1,P<0.05)。在3个试验位点,去除凋落物时(图2),低郁闭度处理下延胡索块茎产量分别比中郁闭度、高郁闭度处理平均增加了279.0%、320.0%;在覆盖凋落物时,白雨伞和大岩后2个位点延胡索产量随着郁闭度的降低而增加。去除凋落物时,延胡索乙素质量分数随着郁闭度的降低而降低。且在高郁闭度处理中,3个试验位点去除凋落物后延胡索乙素质量分数均显著(P<0.05)高于覆盖凋落物。
表 1 郁闭度及凋落物处理对延胡索产量与延胡索乙素质量分数影响的方差分析
Table 1. Analysis of variance of canopy density and litter treatment on yield and tetrahydropyridine content of C. yanhusuo
因素 块茎产量 延胡索乙素质量分数 F P F P 郁闭度 195.06 <0.001 44.44 <0.001 凋落物 5.69 <0.001 89.89 <0.001 位点 102.45 <0.001 0.83 >0.05 郁闭度×凋落物 10.81 <0.001 8.01 <0.05 郁闭度×位点 6.15 <0.05 1.11 >0.05 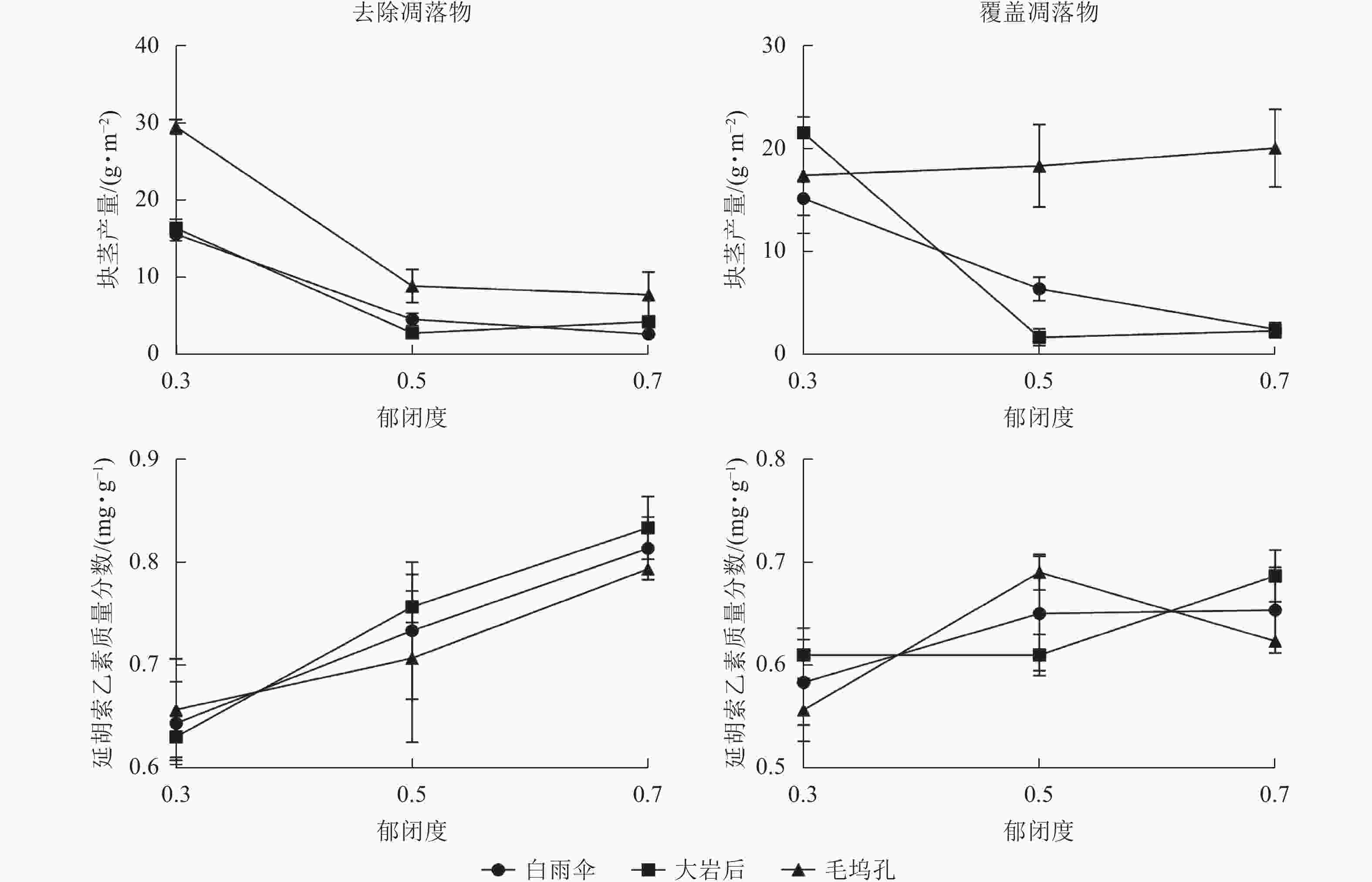
图 2 延胡索产量和延胡索乙素质量分数对不同郁闭度处理的响应
Figure 2. Response of C. yanhusuo yield and tetrahydropyridine content to different canopy densities
-
利用雷达图综合评估不同处理对延胡索生长、产量和品质影响(图3)。结果表明:3个试验位点平均综合评价值得分最高的是低郁闭度+去除凋落物处理,其次为低郁闭度+覆盖凋落物处理。
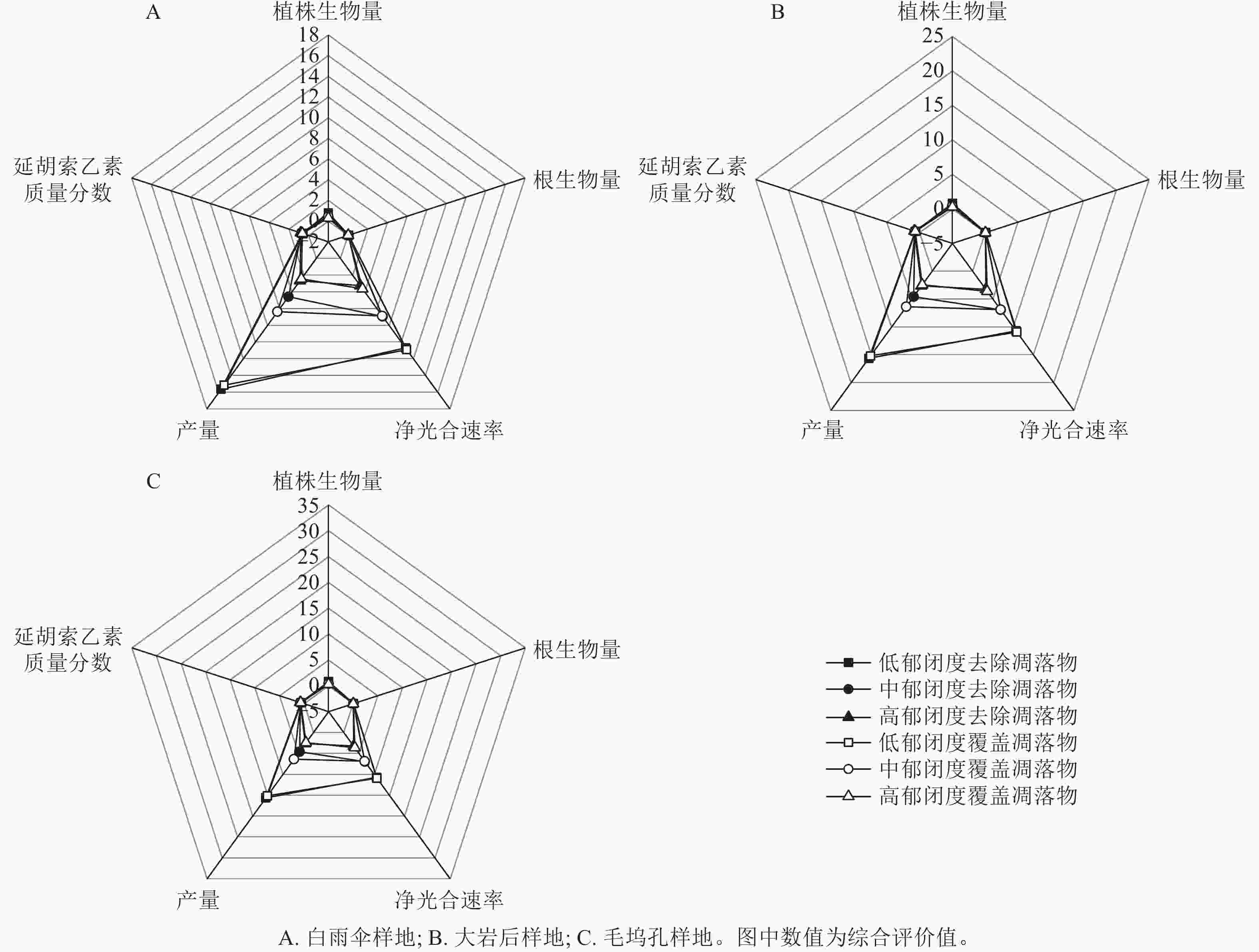
图 3 郁闭度和凋落物影响下延胡索生长、产量和延胡索乙素质量分数的综合评估
Figure 3. Comprehensive evaluation of the effects of canopy density and litter on growth, yield and tetrahydropyridine content of C. yanhusuo
-
去除凋落物时,土壤有机碳、全氮质量分数随着郁闭度的降低而升高(表2);保留凋落物时,白雨伞和大岩后试验位点可溶性氮质量分数随着郁闭度的降低而降低。在低郁闭度处理下,去除凋落物时的可溶性氮质量分数均低于覆盖凋落物处理。在去除凋落物时,白雨伞和毛坞孔试验位点可溶性碳质量分数随着郁闭度的降低而升高。保留凋落物时,3个试验位点低郁闭度处理下土壤真菌群落丰度比高郁闭度处理平均降低41.3%。覆盖凋落物时,大岩后和毛坞孔试验位点的细菌群落丰度在低郁闭度处理下比高郁闭度处理平均降低65.8%。
表 2 郁闭度及凋落物处理对根区土壤养分含量、细菌和真菌丰度的影响
Table 2. Effects of different canopy densities and litter treatment on soil nutrients and the abundance of bacteria and fungi in rhizosphere soil
试验位点 处理 SOC/
(g·kg−1)TN/
( g·kg−1)TP/
( g·kg−1)DOC/
(mg·kg−1)DON/
(mg·kg−1)AVP/
(mg·kg−1)真菌数量/
(×109拷贝·g−1)细菌数量/
(×1010拷贝·g−1)白雨伞 -L 38.36±0.47 aA 3.13±0.08 aA 0.07±0.02 aA 57.31±3.70 aA 98.29±3.08 aB 1.63±0.52 bA 8.89±0.57 aA 4.96±1.08 aA -M 35.41±2.13 aA 3.44±0.29 aA 0.19±0.04 aA 39.54±2.85 bA 58.63±4.84 bA 3.76±0.70 aA 4.72±1.16 bA 4.77±1.02 aA -H 22.48±0.82 bB 2.36±0.13 bA 0.06±0.01 aB 30.33±4.26 cA 87.68±5.94 aA 1.68±0.10 bB 8.08±1.52 aA 3.20±0.16 aA +L 30.45±0.92 aB 3.33±0.20 aA 0.12±0.04 aA 39.69±7.19 aB 129.08±11.94 aA 2.64±0.76 aA 9.33±1.53 bA 5.60±0.88 aA +M 34.03±1.02 aA 2.73±0.10 aA 0.18±0.04 aA 42.56±4.83 aA 73.16±3.41 bA 2.52±0.44 aA 12.83±0.75 aA 5.73±0.99 aA +H 31.78±0.52 aA 2.56±0.11 aA 0.22±0.05 aA 37.57±8.07 aA 36.05±3.71 cA 2.25±0.62 aA 12.68±0.35 aA 4.47±0.54 aA 大岩后 -L 38.59±0.13 aA 3.28±0.21 aA 0.30±0.07 aB 16.74±4.78 bA 75.09±16.71 bB 2.25±0.86 bA 8.64±0.18 aA 3.54±0.50 bA -M 31.11±0.48 bA 2.75±0.13 bA 0.30±0.04 aA 28.32±3.20 aA 124.62±9.71 aA 8.31±0.07 aA 6.35±0.20 bB 1.46±0.10 aA -H 39.35±0.39 aA 2.19±0.16 cA 0.28±0.02 aB 21.47±3.18 abA 64.71±10.69 bA 7.75±0.61 aA 8.09±1.21 abA 2.74±0.32 aA +L 39.95±0.15 aA 2.95±0.10 aB 0.43±0.01 bA 27.08±1.15 aA 132.53±17.23 aA 1.34±0.31 bA 8.68±0.24 cA 3.76±0.39 bA +M 35.00±0.25 bA 3.13±0.06 aA 0.30±0.02 cA 27.39±1.05 aA 92.23±5.40 bA 4.98±1.01 aB 12.21±1.66 bA 4.68±1.01 bA +H 29.20±0.51 cA 2.49±0.23 bA 0.53±0.01 aA 31.27±3.44 aA 86.15±1.22 bA 5.00±0.74 aB 19.99±0.77 aA 13.69±0.75 aA 毛坞孔 -L 38.81±0.79 aA 2.89±0.06 aA 0.64±0.04 aA 36.59±0.76 aA 98.78±1.28 aA 5.28±0.63 aA 8.69±0.43 aA 4.29±0.11 bA -M 37.45±1.14 aA 2.87±0.07 aA 0.76±0.08 aA 30.02±5.20 aA 86.87±3.97 aB 3.11±0.43 bA 5.81±0.47 bA 3.51±0.91 bA -H 33.32±1.14 bA 2.41±0.10 bA 0.69±0.06 aA 29.37±0.93 aA 72.78±8.84 bB 2.88±0.33 bB 6.57±0.39 bA 12.31±1.84 aA +L 41.60±1.43 aA 3.23±0.23 aA 0.62±0.05 bA 18.39±2.90 aB 105.46±6.99 bA 2.23±0.38 cB 8.71±0.44 cA 6.49±1.00 bA +M 40.00±0.73 bA 3.01±0.09 abA 0.77±0.05 aA 33.55±4.22 aA 84.36±6.42 bA 4.21±0.46 bA 11.00±0.59 bA 7.04±1.44 bA +H 35.16±0.56 cA 2.65±0.08 bA 0.66±0.03 bA 23.93±3.14 bA 129.08±11.94 aA 6.84±0.30 aA 12.87±0.38 aA 16.21±2.09 aA 说明:H. 高郁闭度;M. 中郁闭度;L. 低郁闭度;-表示去除凋落物;+表示覆盖凋落物。SOC. 土壤有机碳;TN. 土壤全氮;TP. 土壤全磷;DOC. 土壤可溶性碳;DON. 土壤可溶性氮;AVP. 土壤速效磷。不同小写字母表示相同凋落物不同郁闭度之间差异显著(P<0.05),不同大写字母表示相同郁闭度不同凋落物之间差异显著(P<0.05)。 -
从表3可见:延胡索块茎产量与土壤有机碳、土壤可溶性氮、植株生物量和植株净光合速率具有显著的回归关系(P<0.01);而延胡索乙素质量分数与土壤全氮、植株净光合速率具有显著的回归关系(P<0.01)。
表 3 延胡索产量和延胡索乙素质量分数与各因素间的回归分析
Table 3. Regression analysis between the C. yanhusuo yield, tetrahydropalmatine content and various factors
因变量 延胡索产量 延胡索乙素质量分数 a b R2 P a b R2 P 土壤有机碳 0.970 −23.19 0.31 <0.01 −0.006 0.89 0.110 <0.05 土壤全氮 6.980 −8.97 0.10 <0.05 −0.120 1.02 0.310 <0.01 土壤全磷 14.370 5.29 0.17 <0.05 −0.020 0.69 0.003 >0.05 土壤可溶性碳 −0.004 11.09 −0.02 >0.05 0.000 0.69 0.002 >0.05 土壤可溶性氮 0.140 −1.89 0.21 <0.01 −0.001 0.77 0.110 <0.05 土壤速效磷 −0.530 12.98 0.02 >0.05 0.010 0.64 0.090 <0.05 植株生物量 16.160 0.46 0.41 <0.01 −0.110 0.75 0.180 <0.05 根生物量 73.960 5.85 0.12 <0.05 −0.910 0.74 0.180 <0.05 净光合速率 1.580 −0.14 0.36 <0.01 −0.010 0.78 0.310 <0.01 真菌数量 0.020 10.80 0.00 >0.05 −0.010 0.77 0.130 <0.05 细菌数量 0.290 9.40 0.02 >0.05 −0.010 0.72 0.090 >0.05 说明:R2表示拟合度;a表示系数;b表示截距。 为了进一步揭示主导延胡索块茎产量和延胡索乙素质量分数的影响因子,选取与两者显著(P<0.05)相关的土壤和植物因子与延胡索块茎产量和延胡索乙素质量分数构建多元回归方程(表4)。去除凋落物时,影响延胡索块茎产量的最主要因子是植株生物量,影响延胡索乙素质量分数的最主要因子是净光合速率。覆盖凋落物时,影响延胡索块茎产量的主要因子是根生物量、土壤有机碳和土壤可溶性氮质量分数;影响延胡索乙素质量分数的主要因子是净光合速率、土壤速效磷和土壤有机碳质量分数。
表 4 延胡索产量和延胡索乙素质量分数与各因素间的多元线性回归模型
Table 4. Multiple linear regression model between the C. yanhusuo yield, tetrahydropalmatine content and various factors
因变量 处理 回归方程 R2 重要性排序 延胡索产量(Y) 去除凋落物 Y=27.83X植株−7.41
(X植株=0.79)0.63 植株生物量(X植株) 覆盖凋落物 Y=0.14XDON+1.47XSOC−69.49X根−33.22
(XDON=0.55,XSOC=0.77,X根=−0.41)0.71 从大到小依次为土壤有机碳(XSOC)、土
壤可溶性氮(XDON)、根生物量(X根)延胡索乙素质量分数(Y) 去除凋落物 Y=−0.021XPn+0.880
(XPn=−0.640)0.70 净光合速率($ X_{\mathrm{Pn}} $) 覆盖凋落物 Y=−0.018XPn +0.011XAVP−0.003XSOC+0.930
(XPn =−0.702,XAVP=0.320,XSOC=−0.220)0.79 从大到小依次为净光合速率($ X_{\mathrm{P}{\mathrm{n}}} $)、速
效磷(XAVP)、土壤有机碳(XSOC) -
光照是植物生长过程中必不可少的环境因子,对植物的生长发育、形态建成以及生理生化等均有重要影响[21]。在本研究中,低郁闭度处理下植株净光合速率和生物量显著高于高郁闭度处理,可见降低郁闭度有利于促进延胡索光合作用和植株生物量积累。这与盛海燕等[22]发现遮荫后延胡索最大净光合速率均显著降低的结果一致,随着最大净光合速率的降低,植物的生长和生物量的积累将会受到抑制[23]。回归分析结果表明:延胡索生物量是促进延胡索块茎产量增加的关键因素,说明降低郁闭度是促进延胡索光合速率和生物量积累的关键。
适宜的土壤生境,包括温度和养分供给,也是影响作物产量的主要因素[24]。本研究的3个试验位点,降低郁闭度均显著增加了土壤有机碳和全氮。回归分析也表明:土壤有机碳和土壤可溶性氮是增加产量的重要因素,这与前人[25−26]的研究结果一致,即增加土壤有机碳和养分含量对提高作物产量至关重要。降低郁闭度可以改善土壤养分供应和增加延胡索产量,主要因为:①随着郁闭度的降低,更多的雨水能接触地表凋落物层,而降水的增加能使凋落物中进入土壤的碳和氮显著提高[27];②随着郁闭度的降低,光照增强,地表温度升高,土壤中微生物更加活跃,促进了土壤有机质的矿化[28]。因此通过降低郁闭度可以提高土壤有机碳和可溶性氮质量分数,进而增加延胡索产量。这与上述通过增加养分供应提高作物产量的研究相符。综上所述,降低郁闭度有利于增加土壤养分,从而促进植株生物量的积累,提高延胡索块茎的产量。
-
药用植物适宜的生长环境能促进植株代谢,从而有效促进药用成分积累,最终改善药材品质[29]。本研究发现:降低郁闭度可显著降低块茎中延胡索乙素质量分数。这与前人[30]的研究结果一致,即低光强被认为是刺激某些植物产生次生代谢物的适宜环境,光照强度的减弱诱导植物组织中生物碱的增加。本研究3个试验位点去除凋落物处理的延胡索乙素质量分数均高于覆盖凋落物处理,特别在高郁闭度处理。原因可能是,延胡索乙素质量分数随着延胡索块茎直径的增大呈降低趋势,而覆盖凋落物促进了块茎产量的增加,使得块茎单位质量中的延胡索乙素质量分数降低。因此,适当提高郁闭度和减少凋落物覆盖可以提高延胡索乙素质量分数,从而提升延胡索品质。
-
本研究发现:延胡索在低郁闭度下有利于获得植株生长优势,如提高延胡索植株生物量等,从而获得较高的产量,但是却不利于次生代谢产物延胡索乙素的积累。这与前人[31]的研究具有相似性,如浙贝母Phyllostachys edulis的生物碱质量分数随着郁闭度的变化,其趋势与块茎产量相反[32]。其原因主要是:①根据生长/分化平衡假说,光合产物分配给细胞生长的投入增加,就会减少分化的投入,细胞生长与分化为负相关[33];②延胡索块茎周皮是延胡索有效成分积累的核心区,增产必然造成块茎膨大,进而降低有效成分。在作物育种中,产量和矿物营养元素以及其他活性物质之间存在“稀释效应”[34−35],也可以在一定程度上解释延胡索产量和延胡索乙素质量分数变化趋势呈相反的现象。
药用植物产量与品质通常呈负相关关系,这使得延胡索产量和品质的同时提升成为一项艰巨的任务。本研究去除凋落物处理有利于延胡索乙素质量分数的积累,但是不利于延胡索块茎产量的提高。雷达图分析表明:低郁闭度去除凋落物处理增加产量的同时,可以维持相对较高的延胡索乙素质量分数。回归分析表明:土壤速效磷是增加延胡索乙素质量分数的主要驱动因素,以往的研究[9]也证实本研究试验地存在磷限制,因此提高土壤速效磷质量分数是提升延胡索块茎有效成分的重要措施。后续研究应进一步筛选最适凋落物覆盖量和郁闭度,同时配合林下配方施肥,并实时调整种植方案,在稳定产量的前提下,提升药用植物的品质。
-
延胡索的产量随着郁闭度的降低而增加,而延胡索乙素质量分数随着郁闭度的降低而降低。通过降低郁闭度,提高延胡索植株生物量、土壤有机碳和土壤可溶性氮质量分数来增加延胡索块茎的产量;通过保持较高郁闭度,抑制净光合速率来刺激延胡索乙素质量分数的积累。
Response of yield and tetrahydropalmatine content of Corydalis yanhusuo to canopy density and litter treatment of Chinese fir forest
-
摘要:
目的 探究林下药用植物产量和有效成分积累对林冠郁闭度调节和凋落物处理的响应,为药用植物栽培中调控林下光照强度和养分提供理论依据。 方法 以杉木Cunninghamia lanceolata人工林林下3个试验位点(白雨伞、大岩后和毛坞孔)的药用植物延胡索Corydalis yanhusuo为研究对象,采用双因素裂区试验,探究郁闭度调节(高、中、低)和凋落物处理(覆盖、去除)对延胡索产量和品质指标——延胡索乙素质量分数的影响及其驱动因素。 结果 延胡索块茎产量和延胡索乙素质量分数受到郁闭度、凋落物和两者交互作用的影响显著(P<0.05)。去除凋落物时,随着郁闭度的降低,延胡索块茎产量增加,延胡索乙素质量分数则呈降低趋势;覆盖凋落物时,3个试验位点的延胡索块茎产量和延胡索乙素质量分数表现规律不一致。植株生物量、土壤有机碳和可溶性氮质量分数是影响延胡索块茎产量的重要因素,而延胡索净光合速率、土壤速效磷和土壤有机碳质量分数是影响延胡索乙素质量分数的重要因素。 结论 低郁闭度去除凋落物处理在显著增加延胡索块茎产量的同时,还能维持延胡索乙素的质量分数。图3表4参35 Abstract:Objective This study, with an investigation into the response of yield and active component accumulation of medicinal plants to the regulation of forest canopy density and litter treatment, is aimed to provide theoretical basis for the regulation of light intensity and nutrient content of medicinal plants in simulated cultivation. Method Corydalis yanhusuo, a medicinal plant under Chinese fir (Cunninghamia lanceolata) forest plantation, was first selected as the research object before two-factorial split plot design was employed to explore the impact of three canopy density treatments (high, medium and low) and two litter treatments (removal and mulching) on the yield and active accumulation component of Corydalis yanhusuo. Result The yield of tuber and tetrahydropalmatine content of C. yanhusuo were significantly affected by canopy density, litter and their interaction with the yield decreasing when the litter was removed and the tetrahydropalmatine content increasing with the increase of canopy density. The yield and tetrahydropalmatine content of the three treatments did not show a consistent pattern with the increase of canopy density when litter was covered. According to the radar map, the best planting scheme was to remove litter at low canopy density. Plant biomass, soil organic carbon and soluble nitrogen contents were the important factors affecting tuber yield, while net photosynthetic rate, soil available phosphorus and organic carbon contents were the important factors affecting tetrahydropalmatine content. Conclusion The litter removal treatment with low canopy density can significantly increase the tuber yield of C. yanhusuo and maintain the tetrahydropalmatine content. [Ch, 3 fig. 4 tab. 35 ref.] -
Key words:
- canopy density /
- litter /
- Corydalis yanhusuo /
- tuber yield /
- tetrahydropalmatine
-
图 1 不同郁闭度和凋落物处理对延胡索植株生长的影响
Figure 1 Effects of different canopy density and litter treatment on plant growth of C. yanhusuo
图 2 延胡索产量和延胡索乙素质量分数对不同郁闭度处理的响应
Figure 2 Response of C. yanhusuo yield and tetrahydropyridine content to different canopy densities
图 3 郁闭度和凋落物影响下延胡索生长、产量和延胡索乙素质量分数的综合评估
Figure 3 Comprehensive evaluation of the effects of canopy density and litter on growth, yield and tetrahydropyridine content of C. yanhusuo
表 1 郁闭度及凋落物处理对延胡索产量与延胡索乙素质量分数影响的方差分析
Table 1. Analysis of variance of canopy density and litter treatment on yield and tetrahydropyridine content of C. yanhusuo
因素 块茎产量 延胡索乙素质量分数 F P F P 郁闭度 195.06 <0.001 44.44 <0.001 凋落物 5.69 <0.001 89.89 <0.001 位点 102.45 <0.001 0.83 >0.05 郁闭度×凋落物 10.81 <0.001 8.01 <0.05 郁闭度×位点 6.15 <0.05 1.11 >0.05  下载: 导出CSV
下载: 导出CSV
表 2 郁闭度及凋落物处理对根区土壤养分含量、细菌和真菌丰度的影响
Table 2. Effects of different canopy densities and litter treatment on soil nutrients and the abundance of bacteria and fungi in rhizosphere soil
试验位点 处理 SOC/
(g·kg−1)TN/
( g·kg−1)TP/
( g·kg−1)DOC/
(mg·kg−1)DON/
(mg·kg−1)AVP/
(mg·kg−1)真菌数量/
(×109拷贝·g−1)细菌数量/
(×1010拷贝·g−1)白雨伞 -L 38.36±0.47 aA 3.13±0.08 aA 0.07±0.02 aA 57.31±3.70 aA 98.29±3.08 aB 1.63±0.52 bA 8.89±0.57 aA 4.96±1.08 aA -M 35.41±2.13 aA 3.44±0.29 aA 0.19±0.04 aA 39.54±2.85 bA 58.63±4.84 bA 3.76±0.70 aA 4.72±1.16 bA 4.77±1.02 aA -H 22.48±0.82 bB 2.36±0.13 bA 0.06±0.01 aB 30.33±4.26 cA 87.68±5.94 aA 1.68±0.10 bB 8.08±1.52 aA 3.20±0.16 aA +L 30.45±0.92 aB 3.33±0.20 aA 0.12±0.04 aA 39.69±7.19 aB 129.08±11.94 aA 2.64±0.76 aA 9.33±1.53 bA 5.60±0.88 aA +M 34.03±1.02 aA 2.73±0.10 aA 0.18±0.04 aA 42.56±4.83 aA 73.16±3.41 bA 2.52±0.44 aA 12.83±0.75 aA 5.73±0.99 aA +H 31.78±0.52 aA 2.56±0.11 aA 0.22±0.05 aA 37.57±8.07 aA 36.05±3.71 cA 2.25±0.62 aA 12.68±0.35 aA 4.47±0.54 aA 大岩后 -L 38.59±0.13 aA 3.28±0.21 aA 0.30±0.07 aB 16.74±4.78 bA 75.09±16.71 bB 2.25±0.86 bA 8.64±0.18 aA 3.54±0.50 bA -M 31.11±0.48 bA 2.75±0.13 bA 0.30±0.04 aA 28.32±3.20 aA 124.62±9.71 aA 8.31±0.07 aA 6.35±0.20 bB 1.46±0.10 aA -H 39.35±0.39 aA 2.19±0.16 cA 0.28±0.02 aB 21.47±3.18 abA 64.71±10.69 bA 7.75±0.61 aA 8.09±1.21 abA 2.74±0.32 aA +L 39.95±0.15 aA 2.95±0.10 aB 0.43±0.01 bA 27.08±1.15 aA 132.53±17.23 aA 1.34±0.31 bA 8.68±0.24 cA 3.76±0.39 bA +M 35.00±0.25 bA 3.13±0.06 aA 0.30±0.02 cA 27.39±1.05 aA 92.23±5.40 bA 4.98±1.01 aB 12.21±1.66 bA 4.68±1.01 bA +H 29.20±0.51 cA 2.49±0.23 bA 0.53±0.01 aA 31.27±3.44 aA 86.15±1.22 bA 5.00±0.74 aB 19.99±0.77 aA 13.69±0.75 aA 毛坞孔 -L 38.81±0.79 aA 2.89±0.06 aA 0.64±0.04 aA 36.59±0.76 aA 98.78±1.28 aA 5.28±0.63 aA 8.69±0.43 aA 4.29±0.11 bA -M 37.45±1.14 aA 2.87±0.07 aA 0.76±0.08 aA 30.02±5.20 aA 86.87±3.97 aB 3.11±0.43 bA 5.81±0.47 bA 3.51±0.91 bA -H 33.32±1.14 bA 2.41±0.10 bA 0.69±0.06 aA 29.37±0.93 aA 72.78±8.84 bB 2.88±0.33 bB 6.57±0.39 bA 12.31±1.84 aA +L 41.60±1.43 aA 3.23±0.23 aA 0.62±0.05 bA 18.39±2.90 aB 105.46±6.99 bA 2.23±0.38 cB 8.71±0.44 cA 6.49±1.00 bA +M 40.00±0.73 bA 3.01±0.09 abA 0.77±0.05 aA 33.55±4.22 aA 84.36±6.42 bA 4.21±0.46 bA 11.00±0.59 bA 7.04±1.44 bA +H 35.16±0.56 cA 2.65±0.08 bA 0.66±0.03 bA 23.93±3.14 bA 129.08±11.94 aA 6.84±0.30 aA 12.87±0.38 aA 16.21±2.09 aA 说明:H. 高郁闭度;M. 中郁闭度;L. 低郁闭度;-表示去除凋落物;+表示覆盖凋落物。SOC. 土壤有机碳;TN. 土壤全氮;TP. 土壤全磷;DOC. 土壤可溶性碳;DON. 土壤可溶性氮;AVP. 土壤速效磷。不同小写字母表示相同凋落物不同郁闭度之间差异显著(P<0.05),不同大写字母表示相同郁闭度不同凋落物之间差异显著(P<0.05)。
下载: 导出CSV
表 3 延胡索产量和延胡索乙素质量分数与各因素间的回归分析
Table 3. Regression analysis between the C. yanhusuo yield, tetrahydropalmatine content and various factors
因变量 延胡索产量 延胡索乙素质量分数 a b R2 P a b R2 P 土壤有机碳 0.970 −23.19 0.31 <0.01 −0.006 0.89 0.110 <0.05 土壤全氮 6.980 −8.97 0.10 <0.05 −0.120 1.02 0.310 <0.01 土壤全磷 14.370 5.29 0.17 <0.05 −0.020 0.69 0.003 >0.05 土壤可溶性碳 −0.004 11.09 −0.02 >0.05 0.000 0.69 0.002 >0.05 土壤可溶性氮 0.140 −1.89 0.21 <0.01 −0.001 0.77 0.110 <0.05 土壤速效磷 −0.530 12.98 0.02 >0.05 0.010 0.64 0.090 <0.05 植株生物量 16.160 0.46 0.41 <0.01 −0.110 0.75 0.180 <0.05 根生物量 73.960 5.85 0.12 <0.05 −0.910 0.74 0.180 <0.05 净光合速率 1.580 −0.14 0.36 <0.01 −0.010 0.78 0.310 <0.01 真菌数量 0.020 10.80 0.00 >0.05 −0.010 0.77 0.130 <0.05 细菌数量 0.290 9.40 0.02 >0.05 −0.010 0.72 0.090 >0.05 说明:R2表示拟合度;a表示系数;b表示截距。
下载: 导出CSV
表 4 延胡索产量和延胡索乙素质量分数与各因素间的多元线性回归模型
Table 4. Multiple linear regression model between the C. yanhusuo yield, tetrahydropalmatine content and various factors
因变量 处理 回归方程 R2 重要性排序 延胡索产量(Y) 去除凋落物 Y=27.83X植株−7.41
(X植株=0.79)0.63 植株生物量(X植株) 覆盖凋落物 Y=0.14XDON+1.47XSOC−69.49X根−33.22
(XDON=0.55,XSOC=0.77,X根=−0.41)0.71 从大到小依次为土壤有机碳(XSOC)、土
壤可溶性氮(XDON)、根生物量(X根)延胡索乙素质量分数(Y) 去除凋落物 Y=−0.021XPn+0.880
(XPn=−0.640)0.70 净光合速率($ X_{\mathrm{Pn}} $) 覆盖凋落物 Y=−0.018XPn +0.011XAVP−0.003XSOC+0.930
(XPn =−0.702,XAVP=0.320,XSOC=−0.220)0.79 从大到小依次为净光合速率($ X_{\mathrm{P}{\mathrm{n}}} $)、速
效磷(XAVP)、土壤有机碳(XSOC)
下载: 导出CSV
-
[1] SONG Jinnan, YANG Jingli, JEONG B R. Difference between day and night temperature (DIF) and light intensity affect growth and photosynthetic characteristics of Panax ginseng Meyer sprouts [J/OL]. Plants, 2023, 12(15): 2820[2024-07-01]. DOI: 10.3390/plants12152820. [2] 于耀泓, 林熙, 谭锦豪, 等. 岭南地区林药复合系统5种药用植物生理生态适应性研究[J]. 中药材, 2022, 45(12): 2798−2804. YU Yaohong, LIN Xi, TAN Jinhao, et al. Study on physiological and ecological adaptability of five medicinal plants in compound forest-medicinal plant system in lingnan region [J]. Journal of Chinese Medicinal Materials, 2022, 45(12): 2798−2804. [3] 李冬林, 金雅琴, 崔梦凡, 等. 夏季遮光对连香树幼苗形态、光合作用及叶肉细胞超微结构的影响[J]. 浙江农林大学学报, 2020, 37(3): 496−505. LI Donglin, JIN Yaqin, CUI Mengfan, et al. Growth, photosynthesis and ultrastructure of mesophyll cells for Cercidiphyllum japonicum seedlings with shading in summer [J]. Journal of Zhejiang A&F University, 2020, 37(3): 496−505. [4] ABIDI F, GIRAULT T, DOUILLET O, et al. Blue light effects on rose photosynthesis and photomorphogenesis [J]. Plant Biology, 2013, 15(1): 67−74. [5] 史红专, 严晓芦, 郭巧生, 等. 中药寒热药性与其基原植物生境光照条件相关性分析[J]. 中国中药杂志, 2018, 43(10): 2032−2037. SHI Hongzhuan, YAN Xiaolu, GUO Qiaosheng, et al. Correlation analysis between cold and heat property of traditional Chinese medicines and light conditions of their original plants in habitat [J]. China Journal of Chinese Materia Medica, 2018, 43(10): 2032−2037. [6] 张晓曦, 刘凯旋, 车文绒, 等. 针阔叶凋落物混合分解对酚类物质释放的非加和性影响[J]. 生态学杂志, 2023, 42(12): 2885−2894. ZHANG Xiaoxi, LIU Kaixuan, CHE Wenrong, et al. Non-additive effects of mixed decomposition of coniferous and broad-leaved species litter on the release of phenolics [J]. Chinese Journal of Ecology, 2023, 42(12): 2885−2894. [7] 刘桂霞, 姚翠芳, 王静, 等. 羊草草地凋落物对药用植物柴胡和防风的化感作用[J]. 种子, 2012, 31(11): 1−3, 8. LIU Guixia, YAO Cuifang, WANG Jing, et al. Allelopathic effects of Leymus chinensis grassland litter on medicinal plants of Saposhnikovia divaricata (Turcz) schischk and Bupleurum chinense [J]. Seed, 2012, 31(11): 1−3, 8. [8] 张文卓, 张前前, 俞叶飞, 等. 森林郁闭度和凋落物对延胡索生长和土壤酶化学计量的影响[J]. 农业生物技术学报, 2024, 32(2): 322−333. ZHANG Wenzhuo, ZHANG Qianqian, YU Yefei, et al. Effects of forest canopy density and litter manipulation on Corydalis yanhusuo growth and soil enzyme stoichiometry [J]. Journal of Agricultural Biotechnology, 2024, 32(2): 322−333. [9] 曹瑶, 李凯, 原军宁, 等. 延胡索不同生长时期和不同部位中8种生物碱含量动态比较[J]. 中药材, 2021, 44(1): 41−46. CAO Yao, LI Kai, YUAN Junning, et al. Dynamic comparison of eight alkaloids contents in different growth stages and different parts of Corydalis yanhusuo [J]. Journal of Chinese Medicinal Materials, 2021, 44(1): 41−46. [10] 徐睿, 刘静, 王利艳, 等. 不同地理种源杉木根叶功能性状与碳氮磷化学计量分析[J]. 生态学报, 2022, 42(15): 6298−6310. XU Rui, LIU Jing, WANG Liyan, et al. Analysis of root and leaf functional traits and C, N, P stoichiometry of Cunninghamia lanceolata from different provenances [J]. Acta Ecologica Sinica, 2022, 42(15): 6298−6310. [11] 彭思瑞, 张慧玲, 孙兆林, 等. 长期凋落物去除对亚热带杉木林土壤有机碳及其组分的影响[J]. 植物生态学报, 2024, 48(8): 1708−1088. PENG Sirui, ZHANG Huiling, SUN Zhaolin, et al. Effects of long-term litter removal on soil organic carbon and multiple components in subtropical Cunninghamia lanceolata forest [J]. Chinese Journal of Plant Ecology, 2024, 48(8): 1708−1088. [12] 汪一敏, 陈茜茜, 俞冰, 等. 不同种植模式对延胡索产量及品质形成的影响[J]. 中国现代应用药学, 2019, 36(9): 1042−1049. WANG Yimin, CHEN Xixi, YU Bing, et al. Effects of different cropping patterns on the yield and quality formation of Corydalis rhizoma [J]. Chinese Journal of Modern Applied Pharmacy, 2019, 36(9): 1042−1049. [13] 陈彩霞, 李艾莲, 谢赛萍. 延胡索种茎分级与药材产量及质量关系的研究[J]. 中国农业大学学报, 2017, 22(3): 37−50. CHEN Caixia, LI Ailian, XIE Saiping, et al. Influence of seed-stems grading on plant growth, yield and quality of Corydalis yanhusuo [J]. Journal of China Agricultural University, 2017, 22(3): 37−50. [14] 袁秀云, 许申平, 周一冉, 等. 遮荫对白及形态及叶片结构的影响[J]. 植物研究, 2021, 41(6): 974−981. YUAN Xiuyun, XU Shenping, ZHOU Yiran, et al. Shading effect on morphology and leaf structure of Bletilla striata [J]. Bulletin of Botanical Research, 2021, 41(6): 974−981. [15] 吴海平, 叶根华, 李伟成. 毛竹根系、新鲜枝叶、凋落物及土壤浸提液对浙贝母的化感作用[J]. 浙江农林大学学报, 2023, 40(3): 648−657. WU Haiping, YE Genhua, LI Weicheng. Allelopathic effects of Phyllostachys edulis extracts on Fritillaria thunbergii [J]. Journal of Zhejiang A&F University, 2023, 40(3): 648−657. [16] 王鹏杰, 田昕, 陈树新, 等. 基于多源遥感数据及几何光学模型的森林郁闭度估测[J]. 遥感技术与应用, 2023, 38(2): 383−392. WANG Pengjie, TIAN Xin, CHEN Shuxin, et al. Estimation of forest canopy closure based on multi-source remote sensing data and geometric optical model [J]. Remote Sensing Technology and Application, 2023, 38(2): 383−392. [17] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000: 264−270. BAO Shidan. Soil Agrochemical Analysis [M]. 3rd ed. Beijing: China Agriculture Press, 2000: 264−270. [18] 张静, 周浓, 祁俊生, 等. HPLC同时测定不同产地延胡索中的6种生物碱[J]. 华西药学杂志, 2016, 31(4): 415−419. ZHANG Jing, ZHOU Nong, QI Junsheng, et al. Simultaneous determination of 6 alkaloids in Corydalis yanhusuo from different habitats by HPLC [J]. West China Journal of Pharmaceutical Sciences, 2016, 31(4): 415−419. [19] 王越, 罗雪媛, 李永春, 等. 毛竹与阔叶林凋落物添加对土壤细菌群落的影响[J]. 农业生物技术学报, 2020, 28(6): 951−962. WANG Yue, LUO Xueyuan, LI Yongchun, et al. Effects of litter addition of moso bamboo (Phyllostachys edulis) and broadleaf forest on soil bacterial community [J]. Journal of Agricultural Biotechnology, 2020, 28(6): 951−962. [20] 许雄钞, 张前前, 滕秋梅, 等. 毛竹和次生阔叶林优势树种根际效应对土壤有机碳矿化的影响[J]. 应用生态学报, 2023, 34(9): 2374−2382. XU Xiongchao, ZHANG Qianqian, TENG Qiumei, et al. Rhizosphere effects of moso bamboo and dominant tree species of secondary broadleaved forest on soil organic carbon mineralization [J]. Chinese Journal of Applied Ecology, 2023, 34(9): 2374−2382. [21] 黄财智, 张文辉, 邢忠利, 等. 间伐强度对黄桥林区辽东栎生殖构件的影响[J]. 应用生态学报, 2016, 27(12): 3838−3844. HUANG Caizhi, ZHANG Wenhui, XING Zhongli, et al. Effects of thinning intensities on reproductive modules of Quercus liaotungensis in Huanglong and Qiaoshan Mountains, Northwest China [J]. Chinese Journal of Applied Ecology, 2016, 27(12): 3838−3844. [22] 盛海燕, 李伟成. 延胡索光合与生长可塑性对光照的响应[J]. 生态科学, 2018, 37(6): 168−174 SHENG Haiyan, LI Weicheng. Responses of photosynthesis and growth plasticity of Corydalis yanhusuo to light [J]. Ecological Science, 2018, 37(6): 168−174. [23] 李鑫垚, 黄佳彬, 郭巧生, 等. 光强对菘蓝生长、化学成分及抗氧化活性的影响[J]. 浙江农林大学学报, 2023, 40(2): 356−364. LI Xinyao, HUANG Jiabin, GUO Qiaosheng, et al. Effects of light intensity on growth, chemical composition and antioxidant activity of Isatis indigotica [J]. Journal of Zhejiang A&F University, 2023, 40(2): 356−364. [24] 杨莹攀, 尹娟, 朱银浩, 等. 水氮调控对宁夏中部旱区马铃薯产量与土壤质量的影响[J]. 农业机械学报, 2024, 55(9): 358−370, 458. YANG Yingpan, YIN Juan, ZHU Yinhao, et al. Effect of water and nitrogen regulation on potato yield and soil quality evaluation [J]. Transactions of the Chinese Society for Agricultural Machinery, 2024, 55(9): 358−370, 458. [25] 陈群辉, 柯燕萍, 魏祖晨, 等. 不同基肥对浙贝母根际土壤养分及其入药品质的影响[J]. 南方农业学报, 2021, 52(12): 3358−3365. CHEN Qunhui, KE Yanping, WEI Zuchen, et al. Effects of soil nutrients content in rhizosphere and medicinal quality of Fritillaria thunbergii Miq. with different basal fertilizers [J]. Journal of Southern Agriculture, 2021, 52(12): 3358−3365. [26] QUAN Hao, WANG Bin, WU Lihong, et al. Impact of plastic mulching and residue return on maize yield and soil organic carbon storage in irrigated dryland areas under climate change [J/OL]. Agriculture, Ecosystems & Environment, 2024, 362(1): 108838[2024-07-01]. DOI:10.1016/j.agee.2023.108838. [27] 周祉蕴, 王奕钧, 杨艳丽, 等. 脉冲降水和凋落物对温性草原土壤碳矿化激发效应的影响[J]. 草地学报, 2024, 32(3): 879−888. ZHOU Zhiyun, WANG Yijun, YANG Yanli, et al. Influence of pulsed precipitation and litter on the priming effects of soil carbon mineralization in temperate steppe [J]. Acta Agrestia Sinica, 2024, 32(3): 879−888. [28] 李娜, 赵传燕, 郝虎, 等. 海拔和郁闭度对祁连山青海云杉林叶凋落物分解的影响[J]. 生态学报, 2021, 41(11): 4493−4502. LI Na, ZHAO Chuanyan, HAO Hu, et al. Decomposition and its nutrients dynamic of Qinghai spruce leaf litter with elevation gradient in Qilian Mountains [J]. Acta Ecologica Sinica, 2021, 41(11): 4493−4502. [29] 何冬梅, 王海, 陈金龙, 等. 中药微生态与中药道地性[J]. 中国中药杂志, 2020, 45(2): 290−302. HE Dongmei, WANG Hai, CHEN Jinlong, et al. Microecology and geoherbalism of traditional Chinese medicine [J]. China Journal of Chinese Materia Medica, 2020, 45(2): 290−302. [30] 全妙华, 佘朝文, 欧立军, 等. 遮荫对忽地笑鳞茎中石蒜碱和加兰他敏含量的影响[J]. 中国农业大学学报, 2012, 17(4): 58−61. QUAN Miaohua, SHE Chaowen, OU Lijun, et al. Effect of shading on contents of lycorine and galantamine in the bulb of Lycoris aurea [J]. Journal of China Agricultural University, 2012, 17(4): 58−61. [31] 曹伍林, 付晓莹, 孟祥才, 等. 秋季全光照及水杨酸对人参叶片生理光合特性及人参皂苷类成分含量的影响[J]. 中国实验方剂学杂志, 2016, 22(1): 72−77. CAO Wulin, FU Xiaoying, MENG Xiangcai, et al. Effects of salicylic acid on photosynthesis, physio-biochemistry and quality of ginseng radix et rhizoma under full illumination in autumn [J]. Chinese Journal of Experimental Traditional Medical Formulae, 2016, 22(1): 72−77. [32] 陈如兵, 胡杰, 范慧艳, 等. 不同遮光处理对浙贝母品质的影响[J]. 中国现代应用药学, 2019, 36(3): 312−316. CHEN Rubing, HU Jie, FAN Huiyan, et al. Influence of different shading treatments on Fritillaria thunbergii Miq. quality [J]. Chinese Journal of Modern Applied Pharmacy, 2019, 36(3): 312−316. [33] 苏文华, 张光飞, 李秀华, 等. 植物药材次生代谢产物的积累与环境的关系[J]. 中草药, 2005, 36(9): 139−142. SU Wenhua, ZHANG Guangfei, LI Xiuhua, et al. Relationship between accumulation of secondary metabolism in medicinal plant and environmental condition [J]. Chinese Traditional and Herbal Drugs, 2005, 36(9): 139−142. [34] 李晓靖. 氮肥和前茬作物对小麦籽粒产量和氮、锌等矿质营养元素累积的影响[D] . 济南: 山东师范大学, 2023. LI Xiaojing. Dual Isotopes Tracing Carbon and Nitrogen Dynamics during Leaf Litter Decomposition under Different Precipitation Regimes [D]. Ji’nan: Shandong Normal University, 2023. [35] 陈先敏, 梁效贵, 赵雪, 等. 历年国审玉米品种产量和品质性状变化趋势分析[J]. 中国农业科学, 2018, 51(21): 4020−4029. CHEN Xianmin, LIANG Xiaogui, ZHAO Xue, et al. Analysis on the trends of yield and quality related traits for maize hybrids released in China over the past years [J]. Scientia Agricultura Sinica, 2018, 51(21): 4020−4029. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240461
 点击查看大图
点击查看大图

计量
- 文章访问数: 1547
- HTML全文浏览量: 640
- PDF下载量: 74
- 被引次数: 0
