




-
毛竹Phyllostachys edulis为禾本科Gramineae刚竹属Phyllostachys单轴散生型竹类植物,具有生长更新快,自然成熟期短等优点,是中国分布最广、面积最大的经济竹种[1-2]。毛竹独特的生长习性决定了秆形结构因子及其在竹秆上的分布具有一定的规律性。毛竹的主体是竹秆,为提高毛竹林的经营利用水平,长期以来学者们十分重视毛竹秆形结构研究[3-5]。毛竹的秆形结构通常用竹节数、节长、节间周长(或直径)等因子描述。毛竹的竹节数在笋芽分化时期就已经确定,出土后不再增加新节[3]。毛竹的高生长主要依靠竹节之间的拉伸[4],节间伸长是从基部开始,靠居间分生组织细胞分裂和细胞伸长而逐节伸长的[6-8]。周芳纯[9]较早研究了节间长度、围度等在竹秆上的分布规律,依据毛竹竹秆上自近中央节位向基部和梢部两端推移,毛竹节间长度逐渐变小的特征,用正态分布近似描述节间长度的分布规律,表明节间长度通过正态分布难以反映节长的分布规律。谢芳[10]和甘代奎[11]则对标准竹竹节进行编号,赋值基部第1节竹节竹号为1,并自下而上依次编号,分析秆形结构因子在竹秆上分布规律,表明从基部向上,节长依节号的变化呈抛物线型,竹节周长与节号呈线性负相关。但是,不同毛竹的竹节数往往不同,以节号作为自变量,得出秆形结构因子与节号的关系方程不具有普遍意义。可见,现有描述毛竹秆形结构因子及其在竹秆上的分布规律仍是未解的科学问题。本研究以浙江省受人为干扰较少的毛竹林为研究对象,以竹节所处高度与竹高的比值(即相对高,取值范围0~1)代替实际高,分析秆形结构因子依赖于相对高的分布规律,旨在为毛竹林经营利用提供理论依据。
-
浙江省位于27°01′~31°10′N,118°01′~123°08′E,年均气温为16~19 ℃,四季分明,光照充足,雨水充沛,是典型的亚热带湿润季风气候。浙江省是中国毛竹的主产区之一,有“七山一水二分田”之说,山地和丘陵占74.63%,因而比较适合毛竹的生长,毛竹广泛分布于海拔400~800 m的丘陵、低山山麓地带。根据《2018年浙江省森林资源及其生态功能价值公告》,浙江省森林面积为607.82万 hm2,森林覆盖率为59.71%,竹林面积为92.70万 hm2,占森林面积的15.25%,其中毛竹林面积为81.67万 hm2,占竹林面积的88.10%。
-
2014−2018年,根据浙江省森林资源一类调查系统抽样样地中的毛竹林样地分布较多的区域,共选择10个县市(区)为调查区域,包括庆元县、泰顺县、常山县、武义县、诸暨市、宁海县、黄岩区、安吉县、临安区和余姚市(表1)。在调查区域内,选择近5 a没有采伐、垦复、施肥和钩梢等人为经营管理措施干扰的近自然毛竹林,每个区域随机设置10 m×10 m的样地至少4个,共52个样地。根据样地每木调查结果,按照胸径分布范围,确定大、中、小径级,并在大、中、小径级的样竹中,各随机选取1株梢头完整且无病虫害的样竹,共159株,总竹节数为9 431节。用锯子在竹秆基部齐地伐倒,用皮尺测量竹高。自竹秆基部向上,将竹节从小到大依次编号,称之为节号。用围尺测量每个竹节的竹节长和竹节中央直径。用游标卡尺测量竹秆基部,胸高处,及1/2高处东、南、西、北4个方位的壁厚,取4个方位壁厚的平均值作为对应位置的壁厚。
表 1 研究区概况和样地样竹调查
Table 1. Study area overview and sample setting
地名 纬度
(N)经度
(E)年均气
温/℃年均降
水量/mm样地数/
个样竹数/
株余姚 30°03′ 121°09′ 16.2 1 361 8 24 临安 30°23′ 118°51′ 16.4 1 628 4 12 诸暨 29°43′ 120°32′ 16.3 1 373 6 18 安吉 30°38′ 119°40′ 15.8 1 420 4 12 宁海 29°29′ 121°25′ 16.4 1 480 6 19 泰顺 27°30′ 119°42′ 17.9 1 670 4 12 黄岩 28°38′ 121°17′ 17.0 1 676 4 12 武义 28°54′ 119°48′ 17.9 1 546 6 20 常山 28°51′ 118°30′ 16.3 1 700 6 18 庆元 27°27′ 119°30′ 17.4 1 760 4 12 -
采用SPSS 20.0对毛竹秆形结构因子在区域间的差异性进行了方差分析,对秆形结构因子之间的关系进行了相关性分析,对秆形结构因子间的主导因子进行了因子分析。
-
由表2可知:10个地区毛竹的竹节数最小为39节,最大为76节,平均为59节。竹节数和1/2高节号的变异系数最小,均为0.12。胸高节号的变异系数最大,为0.20。最长竹节长与1/2高节长相近。10个研究区间,毛竹的竹节数、1/2高节长、最长竹节长、基部壁厚、1/2高壁厚、竹高和胸径间差异显著(P<0.05),其他秆形结构在10个地区间差异不显著。
表 2 毛竹秆形结构因子描述统计特征
Table 2. Description statistical characteristics of factor of culm form
指标 竹节数 1/2高节号 胸高节号 1/2高节长/cm 最长竹节长/cm 胸高竹节长/cm 极小值 39 18 6 17.30 18.30 11.70 极大值 76 39 16 46.00 46.00 30.60 均值 59 28 9 34.45 35.43 22.27 标准差 7.38 3.4 1.79 5.35 5.23 3.38 偏度 −0.30 −0.33 1.29 −0.22 −0.32 −0.10 峰度 −0.09 0.38 2.44 −0.20 −0.02 0.78 变异系数 0.12 0.12 0.20 0.16 0.15 0.15 地区显著性 P<0.05 P=0.08 P=0.32 P<0.05 P<0.05 P=0.06 指标 基部壁厚/mm 胸高壁厚/mm 1/2高壁厚/mm 竹高/m 胸径/cm 极小值 8.35 4.93 3.81 7.20 4.20 极大值 25.76 14.52 10.29 20.14 15.30 均值 16.61 10.32 6.53 14.56 10.08 标准差 3.21 2.01 1.10 2.64 2.34 偏度 0.26 −0.16 0.18 −0.23 −0.18 峰度 0.06 −0.40 0.63 −0.07 −0.44 变异系数 0.19 0.19 0.17 0.18 0.23 地区显著性 P<0.05 P=0.83 P<0.05 P<0.05 P<0.05 -
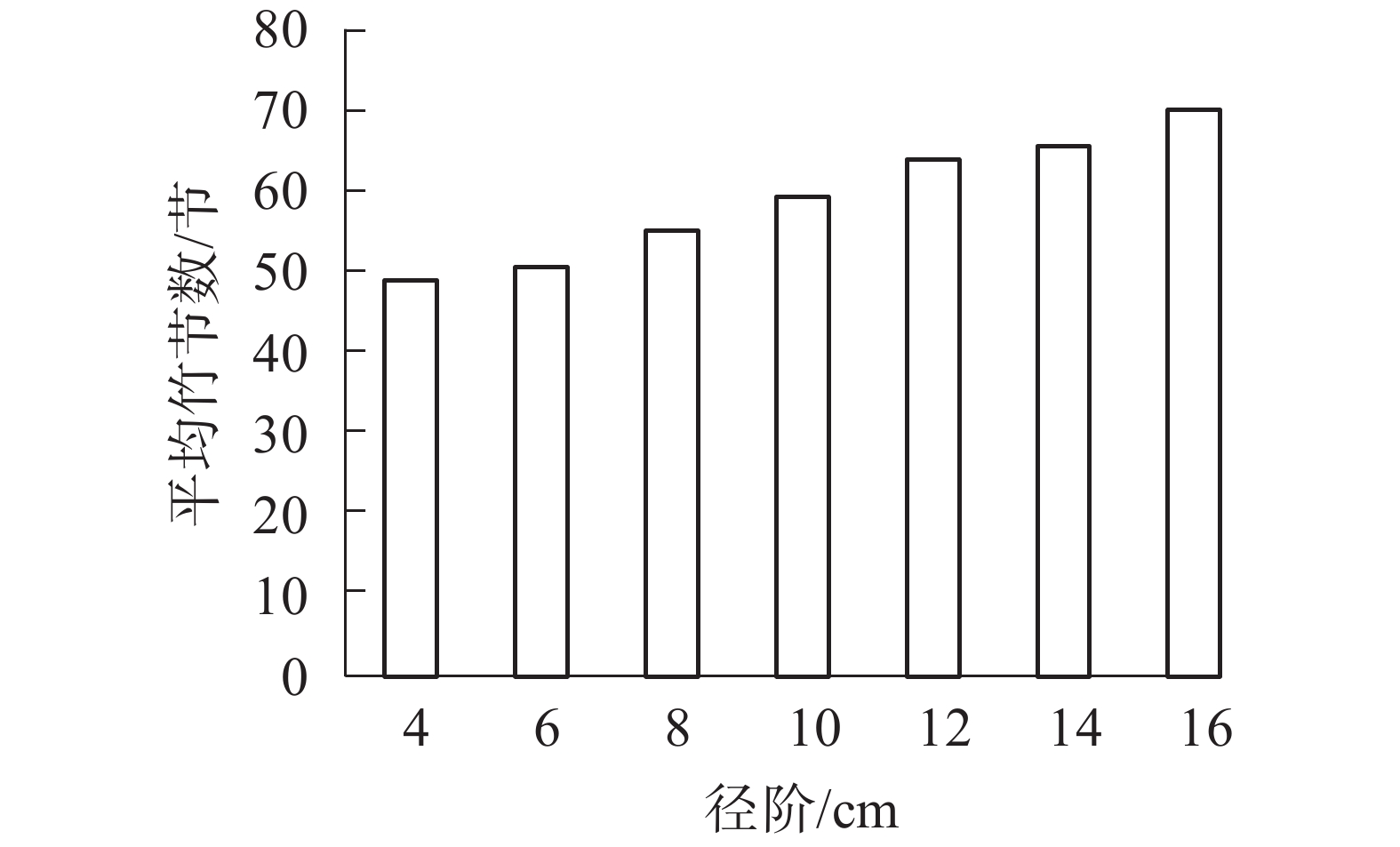
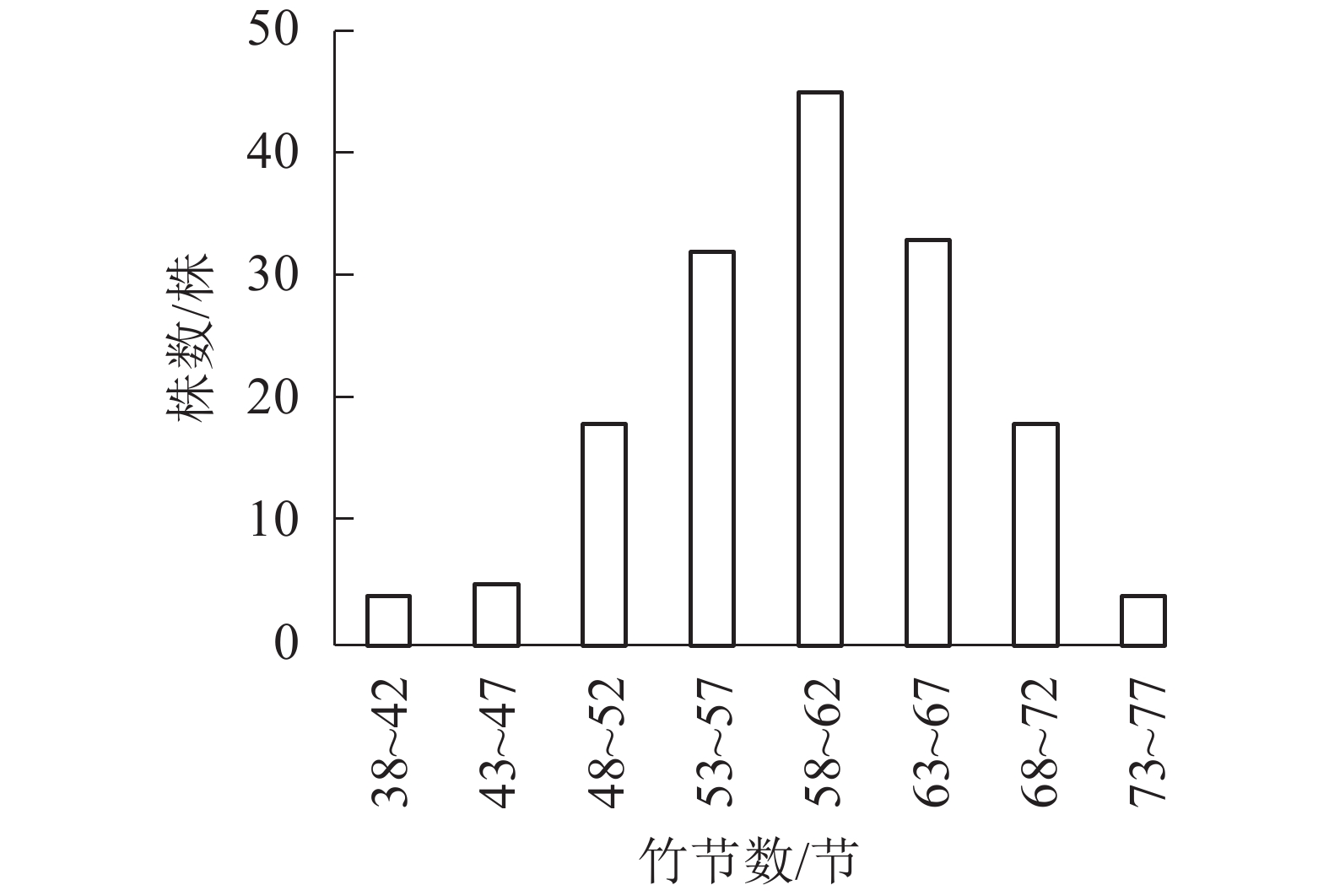
以5节竹节为竹节数级距,分析所采集样竹的竹节数频数分布特征。从图1可知:毛竹的竹节数主要集中在53~67节,且毛竹竹节数的频数分布符合正态分布(P<0.01)。从图2可见:随着径阶的增大,平均竹节数有增大的趋势,即粗壮竹子的竹节数较细小竹子多。
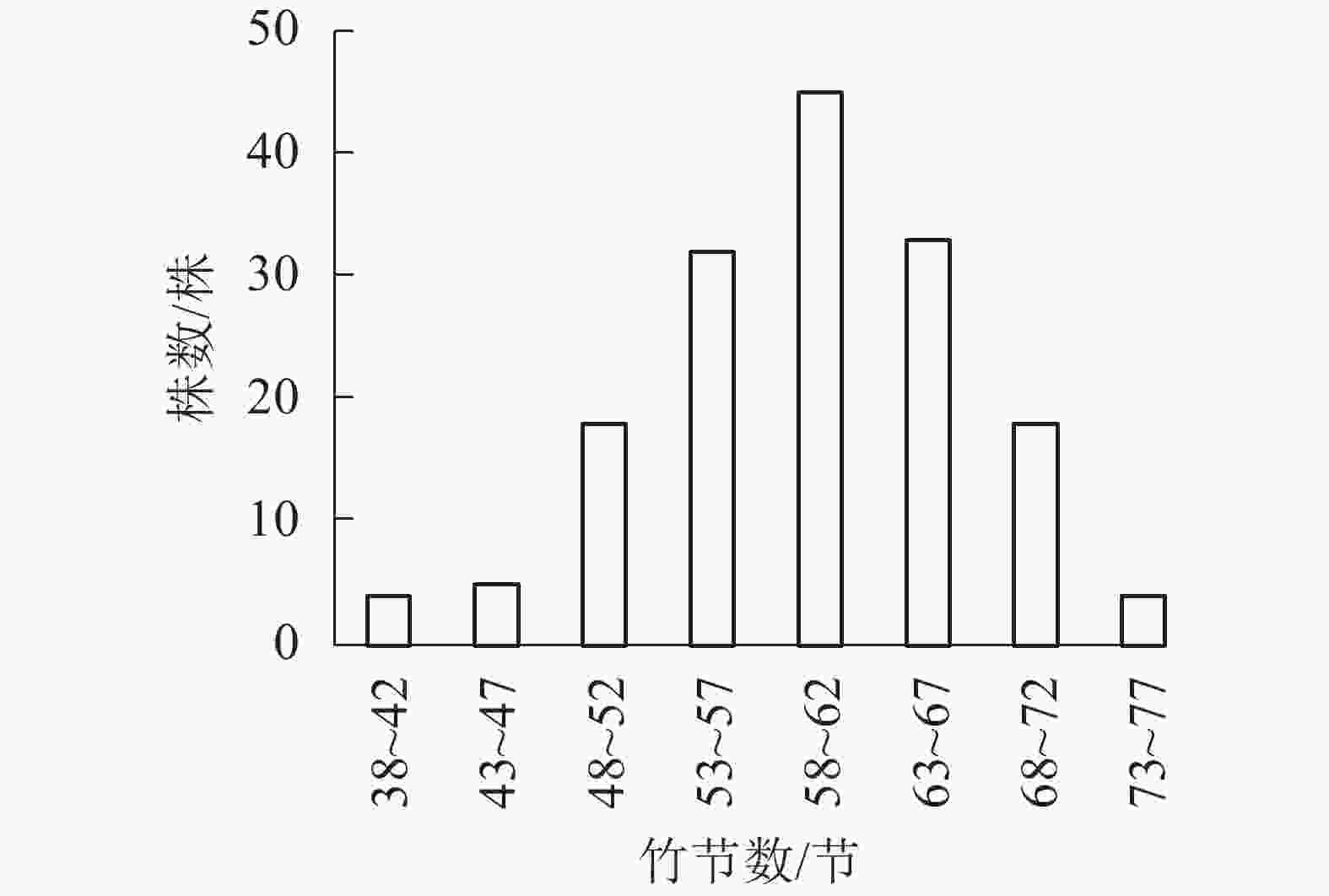
图 1 竹节数的分布频率
Figure 1. Frequency of number of bamboo node

图 2 不同径阶竹节数
Figure 2. Number of bamboo node in different diameter class
-
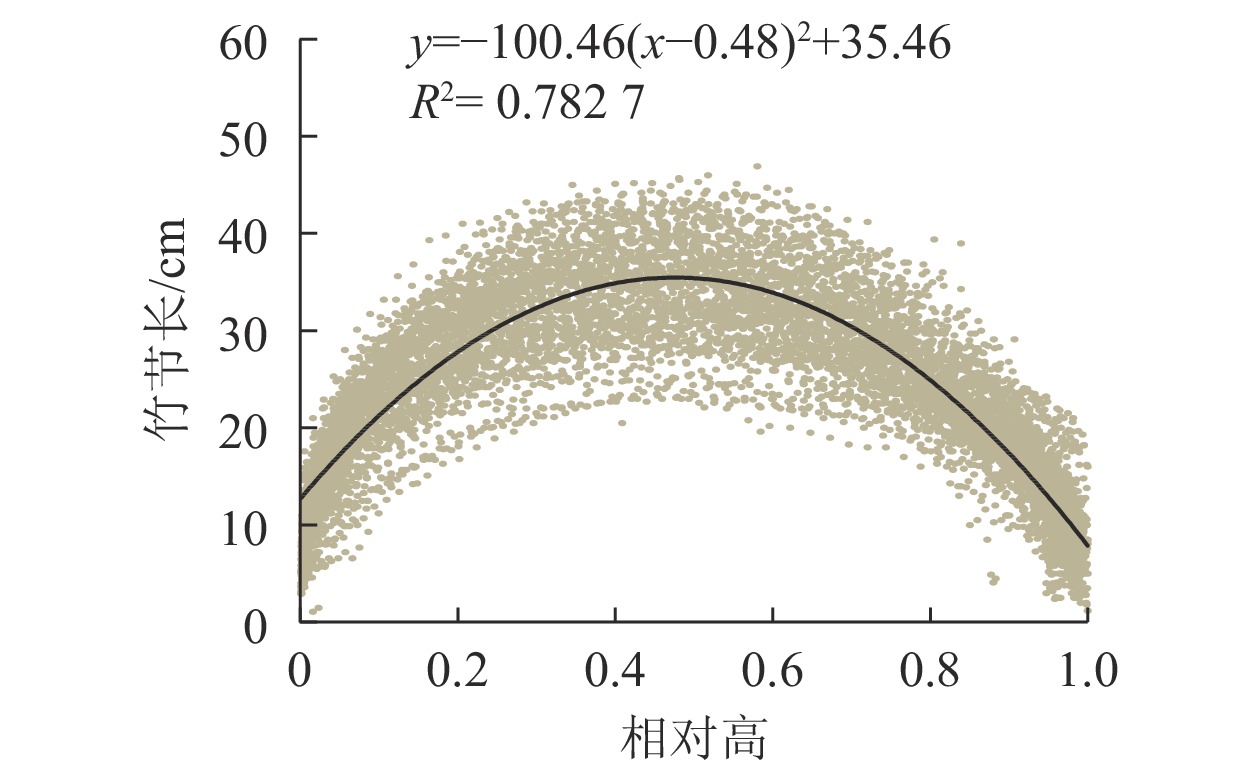
不同毛竹的竹节数、竹节长和竹高往往不同。为消除不同竹高的影响,以竹节所处高度与竹高的比值(即相对高,取值为0~1)代替实际高,分析竹节长在竹秆上的分布规律。从图3可见:随着竹节相对高的增加,竹节长自基部至梢头呈先增大后减小的趋势,并且竹节长在竹秆上的分布具有对称性。因此,可用抛物线描述竹节长在竹秆上的分布规律。根据函数拟合结果,在相对高为0.48时,即约1/2竹高处时,最长竹节为35.46 cm(图3)。
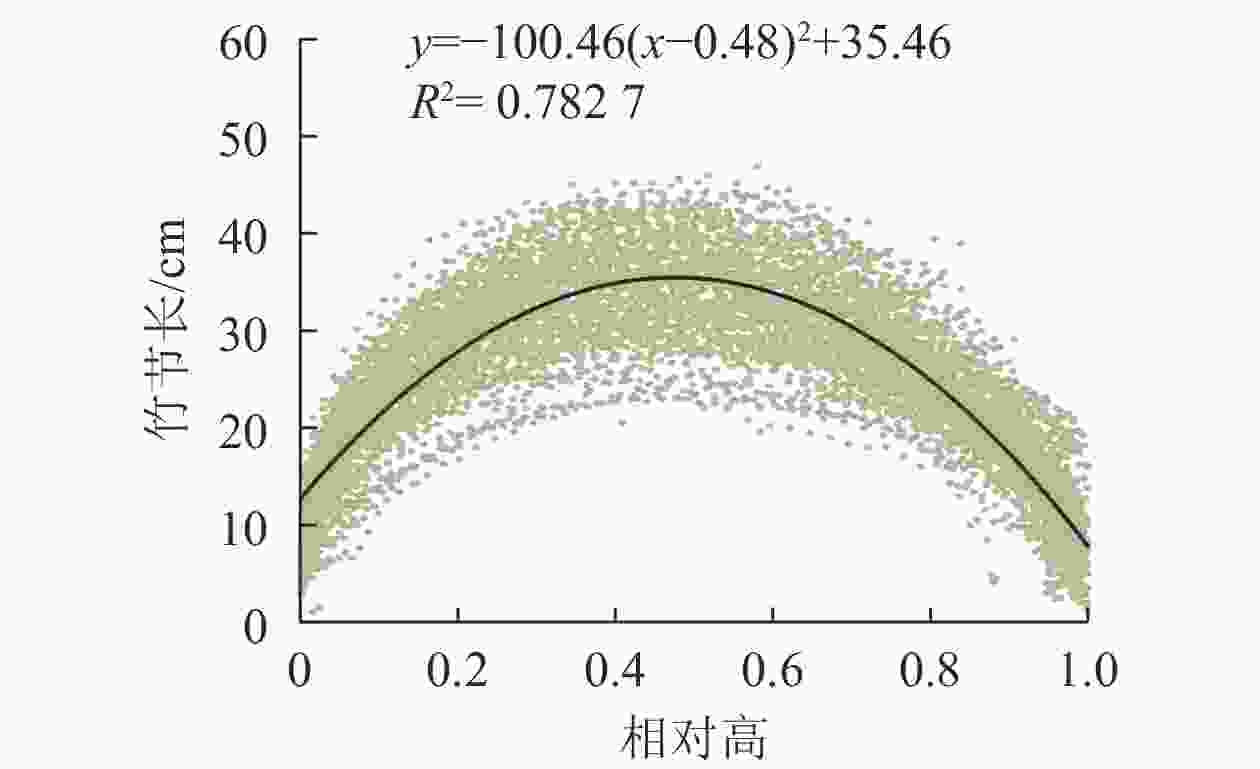
图 3 毛竹竹节长与相对高的关系
Figure 3. Relationship between relative height and length of node
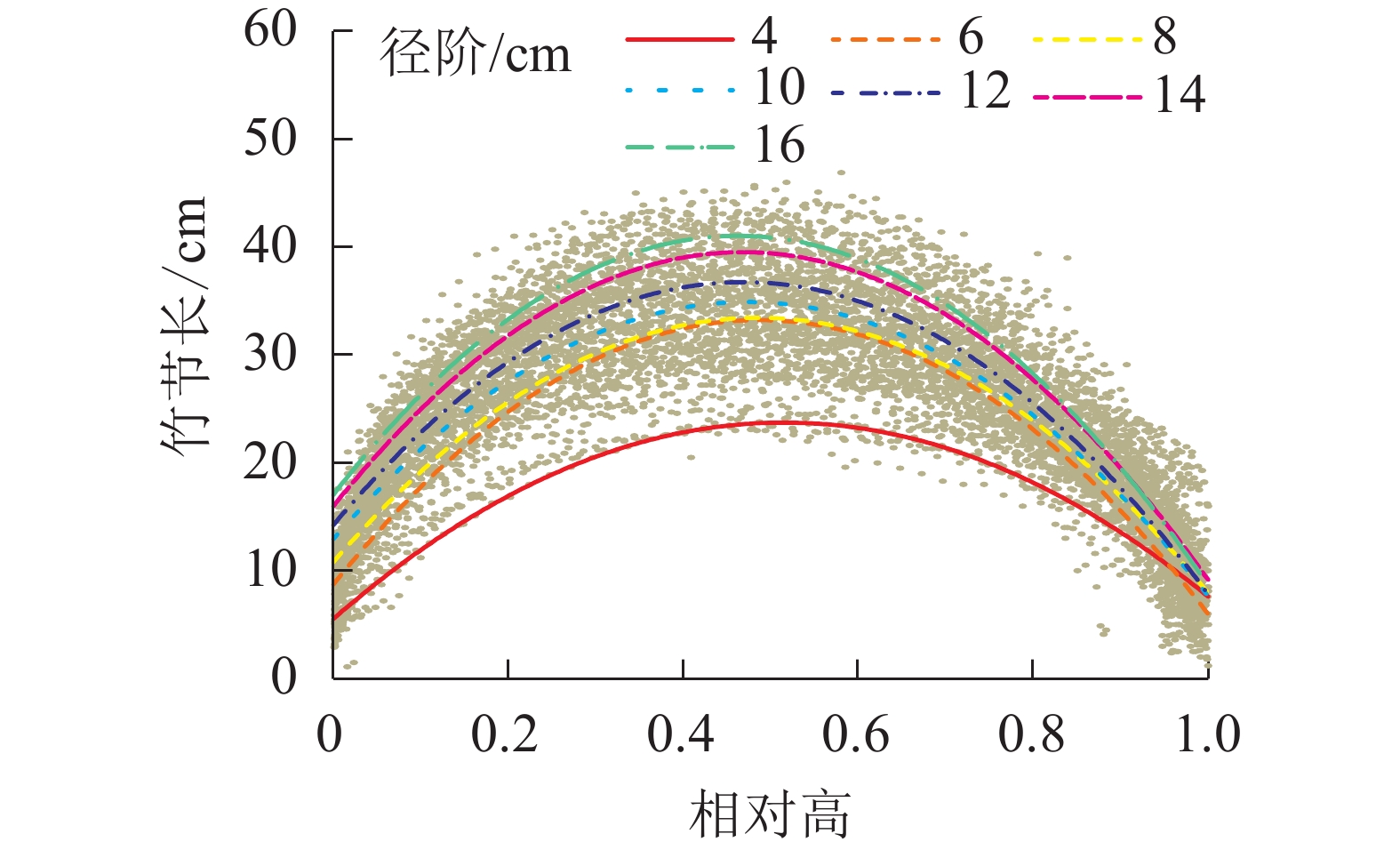
为比较不同径阶毛竹竹节长在竹秆上分布的差异性,绘制了不同径阶的关系图(图4),并拟合抛物线(表3)。经检验,拟合方程均达显著水平(P<0.01)。根据方程拟合结果和不同径阶最长竹节长对应相对高的实测值,可以解析不同径阶最长竹节长对应的相对高,以及某一相对高对应的理论节长的分布规律。从表3可知:不同径阶毛竹最长竹节对应的相对高差异不大,取值均为0.47~0.52,但最长竹节长随着径阶的增大有逐渐变大的趋势。
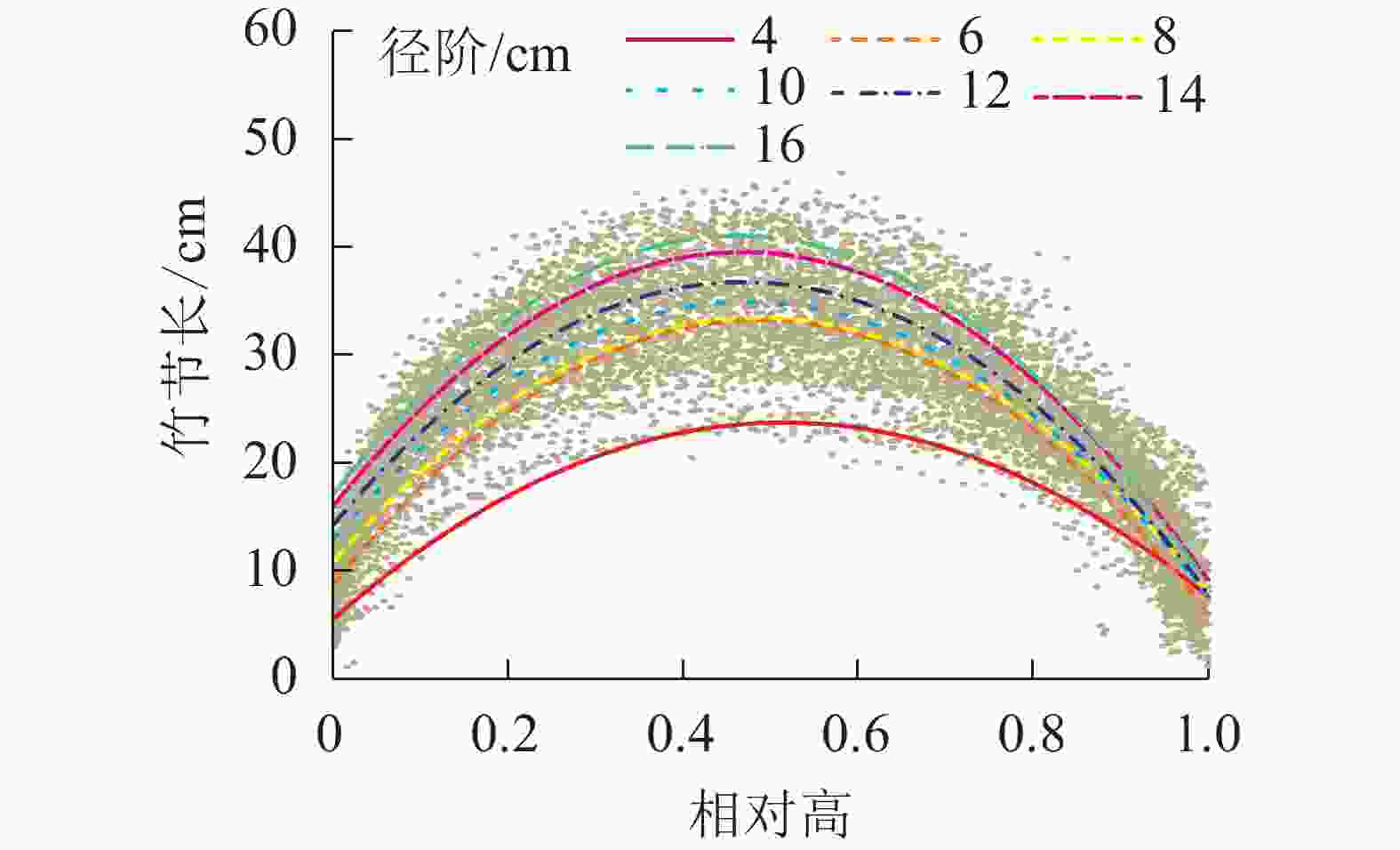
图 4 不同径阶毛竹竹节长在竹秆上的分布
Figure 4. Distribution of length of bamboo node on culm in different diameter class
表 3 不同径阶毛竹竹节长分布特征拟合结果
Table 3. Fitting results of distribution of length of node on culm in different diameter class
径阶/cm a b c R2 ML/cm MRH 4 −68.54 0.52 23.71 0.87 24.40 0.48 6 −103.39 0.49 33.20 0.79 33.45 0.49 8 −96.19 0.49 33.42 0.77 33.32 0.51 10 −97.87 0.47 34.82 0.80 34.76 0.50 12 −102.09 0.47 36.65 0.85 36.60 0.49 14 −107.00 0.47 39.41 0.88 39.16 0.51 16 −111.59 0.46 40.87 0.92 42.40 0.50 说明:拟合函数为y=−a(x−b)2+c,其中x为相对高,y为某相对 高所对应的竹节长度,a和b为函数所对应的参数。R2为 拟合决定系数;ML为实测平均最长竹节长;MRH为实 测最长竹节长对应的相对高 -
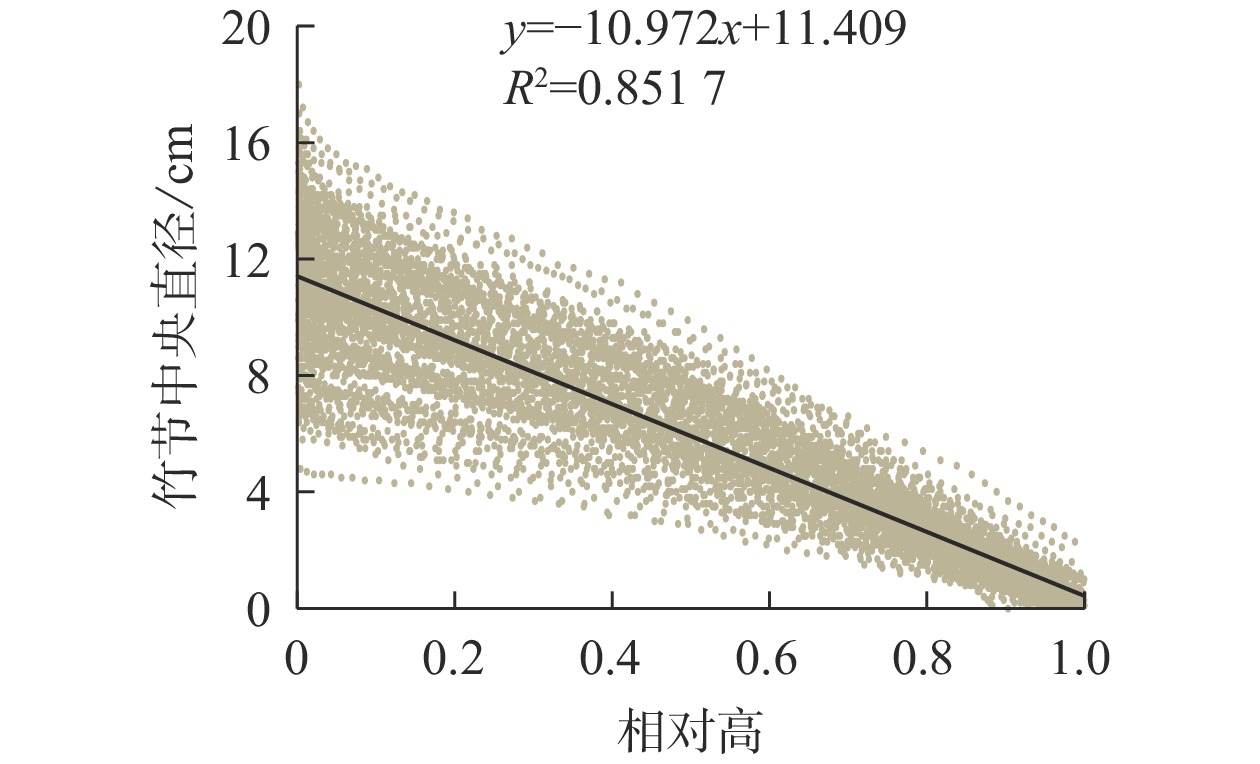
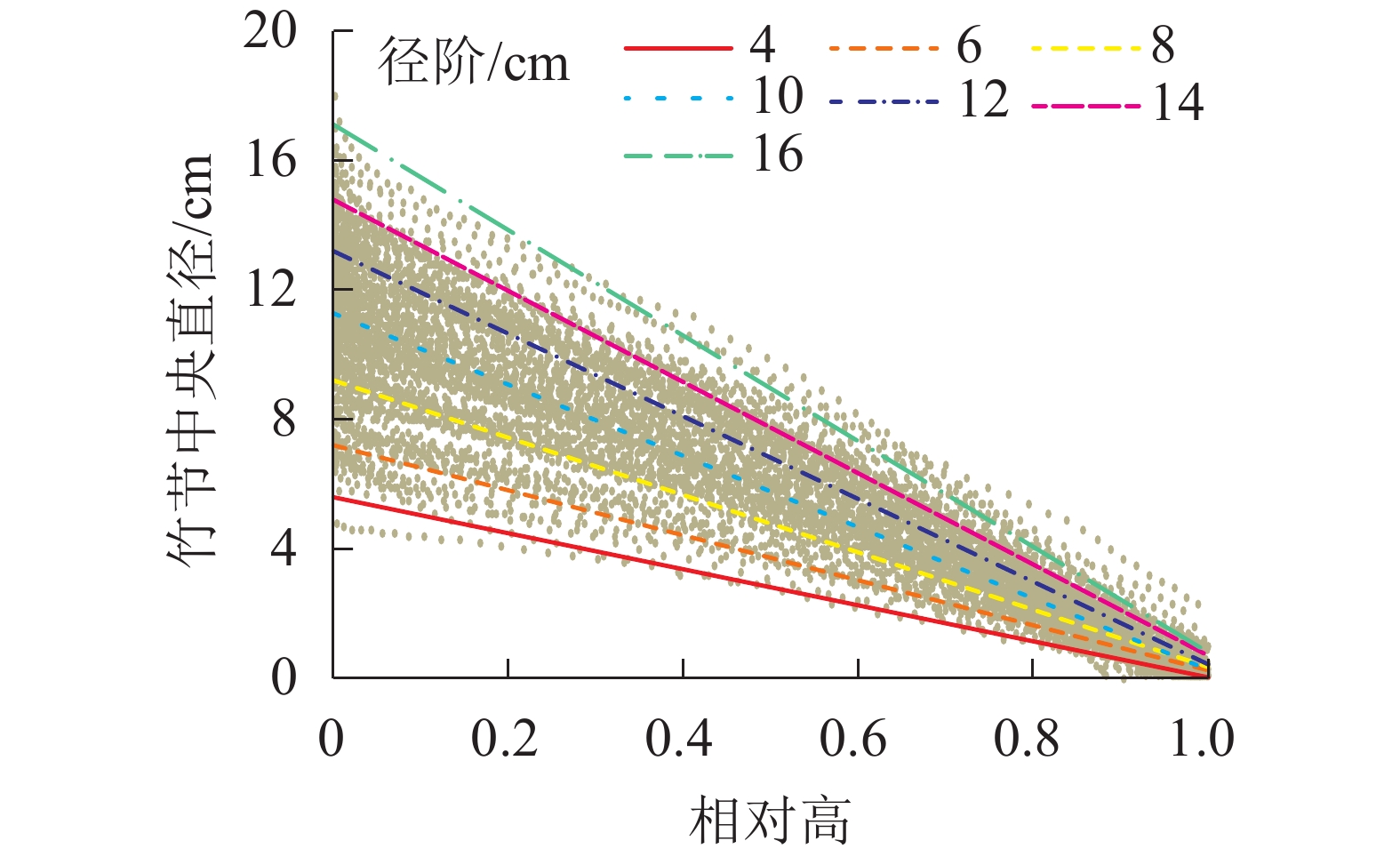
竹节中央直径可反映竹节的大小。从图5可见:随着竹节相对高的增加,竹节中央直径逐渐下降,呈明显的线性负相关关系。不同径阶毛竹竹节中央直径与相对高的关系拟合直线见图6,拟合参数见表4。经检验,拟合方程均达显著水平(P<0.01)。利用拟合方程,可推算不同径阶毛竹在某一相对高对应的理论竹节中央直径。从图6可以看出:随着径阶的增大,拟合函数斜率逐渐变小。但不论毛竹径阶怎么变化,竹秆相对高每增加10%,竹节中央直径约下降10%。
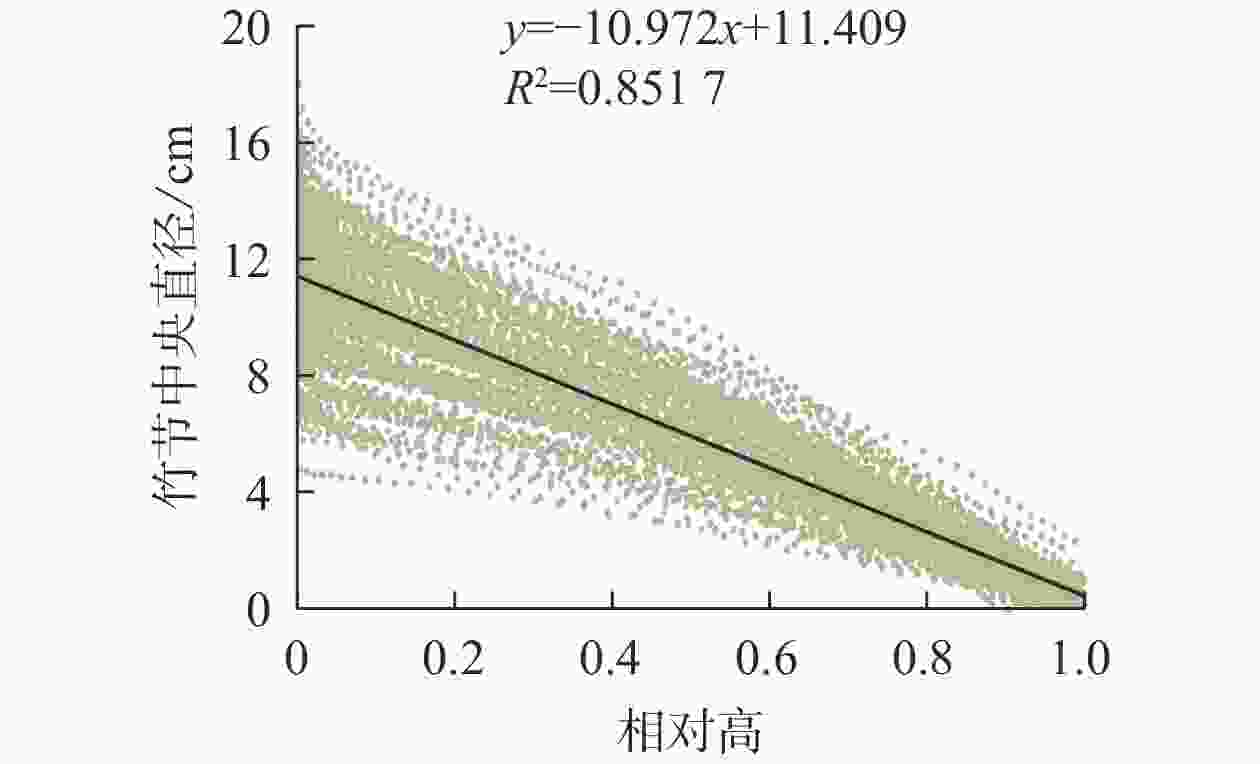
图 5 毛竹竹节中央直径与相对高的线性关系
Figure 5. Relationship between relative height and central diameter of bamboo node
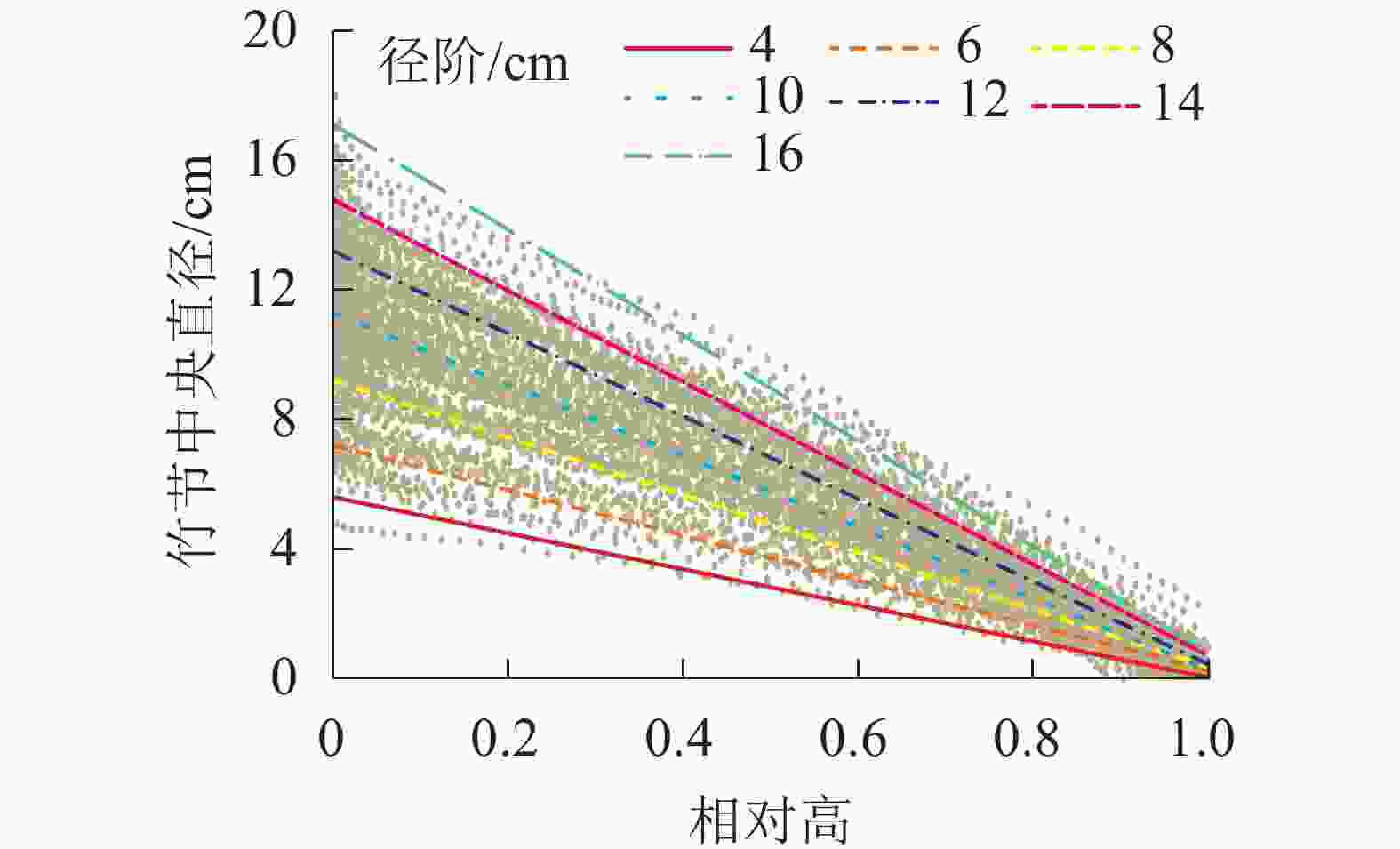
图 6 不同径阶毛竹竹节中央直径在竹秆上的分布
Figure 6. Distribution of central diameter of node on culm in different diameter class
表 4 不同径阶毛竹竹节中央直径分布特征拟合结果
Table 4. Fitting results of central diameter of node on culm in different diameter class
径阶/cm a b R2 4 −5.57 5.60 0.94 6 −6.95 7.20 0.96 8 −8.85 9.21 0.97 10 −11.01 11.28 0.97 12 −12.75 13.19 0.98 14 −14.08 14.79 0.98 16 −16.31 17.11 0.99 说明:拟合函数为y=ax+b,其中x为相对高,y为某相对高度 所对应的竹节直径,a和b为函数所对应的参数。R2为 拟合决定系数 -
为分析毛竹秆形结构因子之间的相互关系,选择11个具有代表性的秆形结构因子进行相关分析。由表5可知:竹节数与胸径、竹高呈显著正相关,相关系数分别为0.667 (P<0.01)和0.640 (P<0.01)。胸径与1/2高壁厚、1/2高节长呈显著正相关,相关系数分别为0.756 (P<0.01)和0.416 (P<0.01),说明竹子越粗壮,1/2高对应的壁厚越厚、节长越长。胸径与胸高节号呈显著负相关,相关系数为−0.551 (P<0.01),说明竹子越粗壮,达到胸高所需的节数越少,即胸高以下的平均节长越长。竹节数和1/2高节号呈显著正相关,相关系数为0.739 (P<0.01),主要原因是竹节长在竹秆上的分布具有对称性(图3)。1/2高节长和最长竹节长的相关系数为0.979 (P<0.01),主要原因是竹节长自基部往梢部有先增加后减少的趋势,且大约在1/2高时竹节最长(图3)。
表 5 毛竹秆形结构因子的相关性分析
Table 5. Correlation analysis of factor of culm form
指标 竹高 胸径 基部壁厚 胸高壁厚 1/2高壁厚 竹节数 1/2高节号 胸高节号 1/2高节长 胸高节长 最长竹节长 竹高 1 胸径 0.865** 1 基部壁厚 0.715** 0.764** 1 胸高壁厚 0.800** 0.875** 0.804** 1 1/2高壁厚 0.665** 0.756** 0.651** 0.801** 1 竹节数 0.640** 0.667** 0.575** 0.625** 0.466** 1 1/2高节号 0.442** 0.444** 0.401** 0.360** 0.256** 0.739** 1 胸高节号 −0.527** −0.551** −0.362** −0.583** −0.513** −0.076 0.326** 1 1/2高节长 0.686** 0.416** 0.386** 0.423** 0.381** 0.029 −0.146 −0.543** 1 胸高节长 0.465** 0.401** 0.289** 0.362** 0.333** −0.109 −0.376** −0.675** 0.684** 1 最长竹节长 0.706** 0.443** 0.411** 0.451** 0.406** 0.043 −0.138 −0.570** 0.979** 0.701** 1 说明:**表示相关极显著(P<0.01) -
由表6可知:前4个主因子含有原始数据90.66%的信息,第1主因子解释了秆形结构27.33%的方差,第2、3、4主因子分别解释了秆形结构26.00%、22.64%和14.69%的方差。与第1主因子相关性较大的是1/2高壁厚、基部壁厚和胸高壁厚,而壁厚因子两两之间相关性较高,均达显著水平(P<0.05,表5),表明第1主因子是反映壁厚的综合因子。与第2主因子相关性较大是1/2高节长和最长竹节长,而最长竹节长与1/2高节长之间的相关性达0.982(表5),表明第2主因子是反映1/2高节长的综合因子。与第3主因子相关性最大的是1/2高节号,其次是竹节数,而1/2高节号与竹节数之间的相关性达0.739,表明第3主因子是反映竹节数的综合因子。与第4主因子相关性最大的是胸高节号,其次是胸高节长和胸径,表明第4主因子是反映胸高处秆形结构的综合因子。因子分析结果表明:壁厚因子、1/2高节长、竹节数和胸高处秆形因子是反映毛竹秆形结构的主要因子。
表 6 毛竹秆形结构因子分析
Table 6. Factor analysis of culm form factor
指标 第1主因子 第2主因子 第3主因子 第4主因子 竹高 0.464 0.586 0.598 0.232 胸径 0.640 0.248 0.562 0.360 胸高节长 0.156 0.624 −0.155 0.608 基部壁厚 0.793 0.270 0.343 0.002 胸高壁厚 0.772 0.220 0.417 0.311 1/2高壁厚 0.865 0.169 0.172 0.206 胸高节号 −0.358 −0.325 0.051 −0.827 1/2高节号 0.222 −0.111 0.882 −0.410 竹节数 0.348 −0.065 0.832 0.036 1/2高节长 0.195 0.951 0.001 0.176 最长竹节长 0.220 0.942 0.010 0.201 特征值 3.000 2.860 2.490 1.610 贡献率 27.330 26.000 22.640 14.690 累计贡献率/% 27.330 53.330 75.970 90.670 -
稳定性是毛竹秆形结构的特点之一。毛竹的竹节数相对稳定,集中在53~67节,平均59节。抛物线可以较好地拟合竹节长与相对高在竹秆上的分布,但不同径阶毛竹最长竹节的相对高没有明显差异,均约在1/2竹高处。这种稳定性主要是由遗传结构和生长特点决定的[12],竹秆中部居间分生组织较两端分生组织活动期长、生长量多,因此中部竹节节间较长,基部和梢部节间较短[4, 13-14]。
-
差异性是毛竹秆形结构的另一个特点。随着径阶的增大,竹节数和最长竹节长有增大的趋势,这与周芳纯[9]的研究结论一致。胸径越大,胸高以下平均竹节长越长,汪阳东[12]的研究也得到相近的结论。差异性则主要受母竹大小和地理生态因素的影响[15-17]。在出笋前期,母竹消耗了大量的营养物质,导致在出笋中后期的竹笋营养匮乏,生长竞争压力增大[18-20],其生长发育受到一定程度影响。此外,母竹发挥其整合作用,优先将营养供给较大的竹子,较小的竹子由于营养竞争其秆形结构的生长受到了影响[21]。
林木的生长性状不仅与物种遗传特性有关,同时也受环境条件的影响[5, 22-23]。在出笋成竹期和孕笋期,气候因子异常,降水量少、气温偏低、寒流持续时间过长等影响竹子的正常生长发育,也会导致毛竹的秆形生长异常,秆形生长量明显变小[16-17, 24-25]。本研究方差分析表明:竹节数、1/2高节长、最长竹节长、基部壁厚、1/2高壁厚、高和胸径在10个区域间差异显著。相关分析表明:年均降水量、年均气温与各秆形结构因子均呈正相关,即随着年均降水量和年均气温的增加,毛竹相同位置对应的秆形指标均有逐渐增加的趋势。这与周文伟[26]的研究结论相似,该研究表明:降水量的增加能促进毛竹的生长,水热条件较高的庆元比水热条件较低的安吉的毛竹产量高。实际上,较好的水热条件可以降低毛竹之间的竞争,促进毛竹的生长[16, 27-28]。可见,年均降水量和年均气温是影响毛竹秆形生长的重要因素。
-
毛竹竹节数服从正态分布,竹节长与竹节相对高呈抛物线关系,且不同径阶毛竹最长竹节的相对高没有明显差异,均约在1/2竹高处。竹节中央直径与竹节所处相对高呈线性负相关关系。随着径阶的增大,最长竹节长和竹节数有逐渐增大的趋势。壁厚因子、1/2高节长、竹节数和胸高处秆形因子是反映毛竹秆形结构的主要结构因子。
On the structure characteristics of culm form of Phyllostachys edulis in Zhejiang Province
-
摘要:
目的 解析秆形结构因子之间的关系及其主导因子,并以竹节所处高度与竹高的比值(即相对高,取值为0~1)代替实际高,分析秆形结构因子及其依赖于相对高的分布规律,可以辅助判断毛竹Phyllostachys edulis生长发育状况。 方法 在浙江省不同区域共选择10个县市(区)设置毛竹林调查样地,测量样竹的竹节数、竹节长和中央直径等秆形结构因子,用抛物线和直线拟合秆形结构因子依赖于相对高在竹秆上的分布规律,通过相关性分析解析秆形结构因子之间的关系,应用因子分析解析秆形结构因子中的主要因子。 结果 毛竹竹节数集中在53~67节,符合正态分布,95%置信水平下毛竹竹节数平均数的置信区间为(58.1, 60.4);竹节长和竹节中央直径依赖于相对高分布分别具有抛物线和直线分布特征。竹秆高度每增加10%,竹节中央直径约下降10%。随着径阶的增大,最长竹节长有逐渐增大的趋势,但最长竹节的相对高没有明显差异,均为0.47~0.52;胸径与竹节数、1/2高节长、最长竹节长和胸高竹节长呈极显著正相关(P<0.01),与胸高节号呈极显著负相关(P<0.01)。 结论 稳定性和差异性并存是毛竹秆形结构因子的主要特点。不同径阶毛竹最长竹节的相对高没有明显差异,均约在1/2竹高处;竹子越粗壮,节数越多,1/2高节长越长,胸高以下平均竹节长越长;壁厚因子、1/2高节长、竹节数和胸高处秆形因子是反映毛竹秆形结构的主要因子。图6表6参28 Abstract:Objective With an analysis of the relationship between stalk structure factors and the dominant factors in stalk structure factors as well as an investigation into the stalk structure factors of culm and their distribution rules dependent on relatively high with the actual high replaced by the ratio of bamboo joint height and bamboo high (i. e. relative high, value range 0−1), this study is aimed to reveal the structure characteristics of culm form so as to better evaluate the growth and development of bamboo (Phyllostachys edulis). Method With 10 counties and cities (districts) of Zhejiang Province selected to set up sample plots of moso bamboo forest, the sample bamboo trees were cut down to measure the number of bamboo nodes, the length of bamboo nodes and the central diameter of bamboo nodes. Then the distribution patterns of culm form factors depending on relative height were fitted by parabola and line before the relationship between culm form factors was investigated with the correlation analysis whereas the main culm structure factors were studied with the factor analysis. Result The number of bamboo bamboo knots was concentrated in 53−67 sections, consistent with the normal distribution, and the confidence interval of the average bamboo knots at the 95% confidence level is (58.1, 60.4). The length of nodes and the central diameter of nodes depended on the relative height distribution with parabola and straight line distribution respectively while the central diameter of bamboo nodes decreased by 10% with a 10% increase of culm height. With the increase of diameter grades, the length of longest bamboo node tended to increase gradually, but there is no significant variation in the relative height of the longest bamboo node ranging between 0.47 and 0.52, and DBH was positively correlated with the number of bamboo nodes, length of node in half height, the length of longest node and the length of node in chest height (P< 0.01), while negatively correlated with the number of nodes in chest height (P<0.01). Conclusion The culm form of bamboo was featured with both stability and variation but there was no significant variation in the relative height of the longest bamboo nodes with different diameter class, generally at half height. Also, the bigger the bamboo was, the larger the number of nodes was, the longer the bamboo nodes were, and the longer the average length below the chest height was. The wall thickness factor, the length of nodes at half height, the number of bamboo nodes and the culm factor in the height of the chest are the main structural factors reflecting the culm structure. [Ch, 6 fig. 6 tab. 28 ref.] -
Key words:
- Phyllostachys edulis /
- culm form structure /
- distribution law /
- factor analysis
-
图 4 不同径阶毛竹竹节长在竹秆上的分布
Figure 4 Distribution of length of bamboo node on culm in different diameter class
图 5 毛竹竹节中央直径与相对高的线性关系
Figure 5 Relationship between relative height and central diameter of bamboo node
图 6 不同径阶毛竹竹节中央直径在竹秆上的分布
Figure 6 Distribution of central diameter of node on culm in different diameter class
表 1 研究区概况和样地样竹调查
Table 1. Study area overview and sample setting
地名 纬度
(N)经度
(E)年均气
温/℃年均降
水量/mm样地数/
个样竹数/
株余姚 30°03′ 121°09′ 16.2 1 361 8 24 临安 30°23′ 118°51′ 16.4 1 628 4 12 诸暨 29°43′ 120°32′ 16.3 1 373 6 18 安吉 30°38′ 119°40′ 15.8 1 420 4 12 宁海 29°29′ 121°25′ 16.4 1 480 6 19 泰顺 27°30′ 119°42′ 17.9 1 670 4 12 黄岩 28°38′ 121°17′ 17.0 1 676 4 12 武义 28°54′ 119°48′ 17.9 1 546 6 20 常山 28°51′ 118°30′ 16.3 1 700 6 18 庆元 27°27′ 119°30′ 17.4 1 760 4 12  下载: 导出CSV
下载: 导出CSV
表 2 毛竹秆形结构因子描述统计特征
Table 2. Description statistical characteristics of factor of culm form
指标 竹节数 1/2高节号 胸高节号 1/2高节长/cm 最长竹节长/cm 胸高竹节长/cm 极小值 39 18 6 17.30 18.30 11.70 极大值 76 39 16 46.00 46.00 30.60 均值 59 28 9 34.45 35.43 22.27 标准差 7.38 3.4 1.79 5.35 5.23 3.38 偏度 −0.30 −0.33 1.29 −0.22 −0.32 −0.10 峰度 −0.09 0.38 2.44 −0.20 −0.02 0.78 变异系数 0.12 0.12 0.20 0.16 0.15 0.15 地区显著性 P<0.05 P=0.08 P=0.32 P<0.05 P<0.05 P=0.06 指标 基部壁厚/mm 胸高壁厚/mm 1/2高壁厚/mm 竹高/m 胸径/cm 极小值 8.35 4.93 3.81 7.20 4.20 极大值 25.76 14.52 10.29 20.14 15.30 均值 16.61 10.32 6.53 14.56 10.08 标准差 3.21 2.01 1.10 2.64 2.34 偏度 0.26 −0.16 0.18 −0.23 −0.18 峰度 0.06 −0.40 0.63 −0.07 −0.44 变异系数 0.19 0.19 0.17 0.18 0.23 地区显著性 P<0.05 P=0.83 P<0.05 P<0.05 P<0.05
下载: 导出CSV
表 3 不同径阶毛竹竹节长分布特征拟合结果
Table 3. Fitting results of distribution of length of node on culm in different diameter class
径阶/cm a b c R2 ML/cm MRH 4 −68.54 0.52 23.71 0.87 24.40 0.48 6 −103.39 0.49 33.20 0.79 33.45 0.49 8 −96.19 0.49 33.42 0.77 33.32 0.51 10 −97.87 0.47 34.82 0.80 34.76 0.50 12 −102.09 0.47 36.65 0.85 36.60 0.49 14 −107.00 0.47 39.41 0.88 39.16 0.51 16 −111.59 0.46 40.87 0.92 42.40 0.50 说明:拟合函数为y=−a(x−b)2+c,其中x为相对高,y为某相对 高所对应的竹节长度,a和b为函数所对应的参数。R2为 拟合决定系数;ML为实测平均最长竹节长;MRH为实 测最长竹节长对应的相对高
下载: 导出CSV
表 4 不同径阶毛竹竹节中央直径分布特征拟合结果
Table 4. Fitting results of central diameter of node on culm in different diameter class
径阶/cm a b R2 4 −5.57 5.60 0.94 6 −6.95 7.20 0.96 8 −8.85 9.21 0.97 10 −11.01 11.28 0.97 12 −12.75 13.19 0.98 14 −14.08 14.79 0.98 16 −16.31 17.11 0.99 说明:拟合函数为y=ax+b,其中x为相对高,y为某相对高度 所对应的竹节直径,a和b为函数所对应的参数。R2为 拟合决定系数
下载: 导出CSV
表 5 毛竹秆形结构因子的相关性分析
Table 5. Correlation analysis of factor of culm form
指标 竹高 胸径 基部壁厚 胸高壁厚 1/2高壁厚 竹节数 1/2高节号 胸高节号 1/2高节长 胸高节长 最长竹节长 竹高 1 胸径 0.865** 1 基部壁厚 0.715** 0.764** 1 胸高壁厚 0.800** 0.875** 0.804** 1 1/2高壁厚 0.665** 0.756** 0.651** 0.801** 1 竹节数 0.640** 0.667** 0.575** 0.625** 0.466** 1 1/2高节号 0.442** 0.444** 0.401** 0.360** 0.256** 0.739** 1 胸高节号 −0.527** −0.551** −0.362** −0.583** −0.513** −0.076 0.326** 1 1/2高节长 0.686** 0.416** 0.386** 0.423** 0.381** 0.029 −0.146 −0.543** 1 胸高节长 0.465** 0.401** 0.289** 0.362** 0.333** −0.109 −0.376** −0.675** 0.684** 1 最长竹节长 0.706** 0.443** 0.411** 0.451** 0.406** 0.043 −0.138 −0.570** 0.979** 0.701** 1 说明:**表示相关极显著(P<0.01)
下载: 导出CSV
表 6 毛竹秆形结构因子分析
Table 6. Factor analysis of culm form factor
指标 第1主因子 第2主因子 第3主因子 第4主因子 竹高 0.464 0.586 0.598 0.232 胸径 0.640 0.248 0.562 0.360 胸高节长 0.156 0.624 −0.155 0.608 基部壁厚 0.793 0.270 0.343 0.002 胸高壁厚 0.772 0.220 0.417 0.311 1/2高壁厚 0.865 0.169 0.172 0.206 胸高节号 −0.358 −0.325 0.051 −0.827 1/2高节号 0.222 −0.111 0.882 −0.410 竹节数 0.348 −0.065 0.832 0.036 1/2高节长 0.195 0.951 0.001 0.176 最长竹节长 0.220 0.942 0.010 0.201 特征值 3.000 2.860 2.490 1.610 贡献率 27.330 26.000 22.640 14.690 累计贡献率/% 27.330 53.330 75.970 90.670
下载: 导出CSV
-
[1] 范叶青, 周国模, 施拥军, 等. 坡向坡位对毛竹林生物量与碳储量的影响[J]. 浙江农林大学学报, 2012, 29(3): 321 − 327. FAN Yeqing, ZHOU Guomo, SHI Yongjun, et al. Relationship of slope aspect and position on biomass and carbon storage in a Phyllostachys edulis stand [J]. J Zhejiang A&F Univ, 2012, 29(3): 321 − 327. [2] 周燕, 刘彩霞, 徐秋芳, 等. 毛竹入侵森林对固氮微生物群落结构和丰度的影响[J]. 植物营养与肥料学报, 2018, 24(4): 1047 − 1057. ZHOU Yan, LIU Caixia, XU Qiufang, et al. Effects of moso bamboo invasion on nitrogen-fixing microbial community structure and abundance in forest [J]. J Plant Nutr Fert, 2018, 24(4): 1047 − 1057. [3] 崔凯. 毛竹茎秆快速生长的机理研究[D]. 北京: 中国林业科学研究院, 2011. CUI Kai. The Mechanism Research of Fast-Growing Culms of Phyllostachys heterocycla[D]. Beijing: Chinese Academy of Forestry, 2011. [4] 曹永慧, 萧江华, 陈双林, 等. 竹阔混交林阔叶树下土壤养分对毛竹生长的影响[J]. 南京林业大学学报(自然科学版), 2007, 31(6): 43 − 47. CAO Yonghui, XIAO Jianghua, CHEN Shuanglin, et al. The effect of soil nutrient around broad-leaved trees on Phyllostachys edulis growth in the mixed forest [J]. J Nanjing For Univ Nat Sci Ed, 2007, 31(6): 43 − 47. [5] 郑进烜, 董文渊, 陈冲, 等. 海子坪天然毛竹林叶面积指数及秆形结构[J]. 林业科技开发, 2009, 23(3): 44 − 48. ZHENG Jinxuan, DONG Wenyuan, CHEN Chong, et al. A study on leaf area and culm-form structure of natural Phyllostachys pubescens in Haiziping [J]. J China For Sci Technol, 2009, 23(3): 44 − 48. [6] 熊文愈, 丁祖福, 李又芬. 竹类植物的居间分生组织与节间生长(Ⅰ)秆茎的居间分生组织与节间生长[J]. 林业科学, 1980, 16(2): 81 − 89. XIONG Wenyu, DING Zufu, LI Youfen. Intercalary meristem and internodal elongation of bamboo plants [J]. Sci Silv Sin, 1980, 16(2): 81 − 89. [7] CUI Kai, HE Caiyun, ZHANG Jianguo, et al. Temporal and spatial profiling of internode elongation-associated protein expression in rapidly growing culms of bamboo [J]. J Proteome Res, 2012, 11(4): 2492 − 2507. [8] 茂玲, 李霞, 杨杨, 等. 木本竹与草本竹节间生长规律比较[J]. 西南林业大学学报(自然科学), 2019, 39(3): 40 − 47. MAO Ling, LI Xia, YANG Yang, et al. Comparative study on the internode growth regularity of woody and herbaceous bamboos [J]. J Southwest For Univ Nat Sci, 2019, 39(3): 40 − 47. [9] 周芳纯. 毛竹秆形结构的研究[J]. 南京林产工业学院学报, 1981, 5(1): 16 − 69. ZHOU Fangchun. Studies on the structure of culm form of Phyllostachys pubescens [J]. J Nanjing Technol Coll For Prod, 1981, 5(1): 16 − 69. [10] 谢芳. 毛竹节间性状及其海拔效应研究[J]. 江西农业大学学报, 2002, 24(1): 86 − 89. XIE Fang. A study on the main internode characters of moso bamboo and its altitude effect [J]. Acta Agric Univ Jiangxi, 2002, 24(1): 86 − 89. [11] 甘代奎. 毛竹纸浆林秆形结构变化规律研究[J]. 湖北林业科技, 2009(2): 34 − 36. GAN Daikui. Study on rule in change of trunk shape structure for pulp forest of Phyllostachys pubescens Mazel ex H. de Lehaie [J]. Hubei For Sci Technol, 2009(2): 34 − 36. [12] 汪阳东. 竹子秆形生长和变异的研究进展[J]. 竹子研究汇刊, 2001, 24(4): 28 − 32. WANG Yangdong. Current research on bamboo culm form [J]. J Bamboo Res, 2001, 24(4): 28 − 32. [13] 卜柯丽, 傅卢成, 王灵杰, 等. 毛竹茎秆快速生长期PeATG1/PeATG4基因表达分析[J]. 浙江农林大学学报, 2020, 37(1): 43 − 50. BU Keli, FU Lucheng, WANG Lingjie, et al. Analysis of PeATG1/PeATG4 gene expression in Phyllostachys edulis during rapid growth [J]. J Zhejiang A&F Univ, 2020, 37(1): 43 − 50. [14] WANG H Y, CUI Kai, HE Caiyun, et al. Endogenous hormonal equilibrium linked to bamboo culm development [J]. Genet Mol Res GMR, 2015, 14(3): 11312 − 11323. [15] 叶诚业, 张遵强. 毛竹[M]. 北京: 农村读物出版社, 1985. [16] 汪阳东, 韦德煌. 气象因素对毛竹秆形生长变异的影响[J]. 竹子研究汇刊, 2002, 21(1): 46 − 52. WANG Yangdong, WEI Dehuang. The effect of weather factors on the culm growth of moso bamboo [J]. J Bamboo Res, 2002, 21(1): 46 − 52. [17] 鲁顺保, 饶玮, 彭九生, 等. 立地条件对毛竹生物量的影响研究[J]. 浙江林业科技, 2008, 28(4): 22 − 27. LU Shunbao, RAO Wei, PENG Jiusheng, et al. Effect of site conditions on biomass of Phyllostachys heterocycla var. pubescens [J]. J Zhejiang For Sci Technol, 2008, 28(4): 22 − 27. [18] 郑郁善, 洪伟, 邱尔发. 毛竹出笋退笋规律的研究[J]. 林业科学, 1998, 34(1): 73 − 77. ZHENG Yushan, HONG Wei, QIU Erfa. Study on the regulation of shoot emergency and degradation of Phyllostachys heterocycla var. pubescens [J]. Sci Silv Sin, 1998, 34(1): 73 − 77. [19] 李睿, 钟章成, 维尔格 M J A. 毛竹(Phyllostachys pubescens)竹笋群动态的研究[J]. 植物生态学报, 1997, 21(1): 53 − 59. LI Rui, ZHONG Zhangcheng, WERGER M J A. Studies the dynamics of the bamboo shoots in Phyllostachys pubescens [J]. Acta Phytoecol Sin, 1997, 21(1): 53 − 59. [20] 岳晋军, 袁娜, 李海峰, 等. 竹类植物退笋现象研究[J]. 竹子学报, 2018, 37(3): 25 − 31. YUE Jinjun, YUAN Na, LI Haifeng, et al. Studies on the abortion of bamboo shoots [J]. J Bamboo Res, 2018, 37(3): 25 − 31. [21] ALPERT P. Effects of clonal integration on plant plasticity in Fragaric chilaaensis [J]. Plant Ecol, 1999, 141(1): 99 − 106. [22] 林元震. 林木基因型与环境互作的研究方法及其应用[J]. 林业科学, 2019, 55(5): 142 − 151. LIN Yuanzhen. Research methodologies for genotype by environment interactions in forest trees and their applications [J]. Sci Silv Sin, 2019, 55(5): 142 − 151. [23] RAYMOND C A. Genotype by environment interactions for Pinusradiata in New South Wales [J]. Tree Genet Genomes, 2011, 7(4): 819 − 833. [24] 郭宝华, 刘广路, 范少辉, 等. 不同生产力水平毛竹林碳氮磷的分布格局和计量特征[J]. 林业科学, 2014, 50(6): 1 − 9. GUO Baohua, LIU Guanglu, FAN Shaohui, et al. Distribution patterns and stoichiometry characteristics of C, N, P in Phyllostachys edulis forests of different productivity levels [J]. Sci Silv Sin, 2014, 50(6): 1 − 9. [25] SHI Yongjun, XU Lin, ZHOU Yufeng, et al. Quantifying driving factors of vegetation carbon stocks of Moso bamboo forests using machine learning algorithm combined with structural equation model [J]. For Ecol Manage, 2018, 429(1): 406 − 413. [26] 周文伟. 降水对毛竹林生长的影响分析[J]. 竹子研究汇刊, 1991, 10(1): 33 − 39. ZHOU Wenwei. Effect of precipitation on growth of Phyllostachys pubescens forest [J]. J Bamboo Res, 1991, 10(1): 33 − 39. [27] 杨帆, 汤孟平. 毛竹林立地与结构的关系及其对生物量的影响[J]. 浙江农林大学学报, 2020, 37(5): 823 − 832. YANG Fan, TANG Mengping. Relationship between site and structure and its influence on biomass inPhyllostachys edulis forest [J]. J Zhejiang A&F Univ, 2020, 37(5): 823 − 832. [28] 张雷, 杨光耀, 黎祖尧, 等. 不同产地厚竹秆形结构比较[J]. 竹子学报, 2017, 36(1): 19 − 24. ZHANG Lei, YANG Guangyao, LI Zuyao, et al. Form and structure of bamboo culm of Phyllostachys edulis ‘Pachyloen’ growing in different habitats [J]. J Bamboo Res, 2017, 36(1): 19 − 24. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200796
 点击查看大图
点击查看大图
计量
- 文章访问数: 2047
- HTML全文浏览量: 626
- PDF下载量: 62
- 被引次数: 0
