






-
云南松Pinus yunnanesis是松科Pinaceae松属Pinus常绿针叶乔木,生长快,材质好,耐干旱瘠薄,天然更新能力强,是西南地区荒山造林先锋树种和主要的用材树种[1],具有很高的经济开发价值。然而,因遗传改良滞后,云南松优质高效人工林的发展受到严重制约。有研究[2]表明,云南松树高、通直度、结实量等性状受基因连锁或功效基因的作用较大,受环境的影响较小,有较高的遗传稳定性。同时,群体内存在许多生长、形质和材性兼优的家系、个体,尤其是生长性状的遗传变异潜力巨大,只要通过有效的良种选育就能发掘这些优良基因型[3]。花粉研究是研究遗传改良的重要内容,对于云南松花粉,除了花粉形态[4]、花粉成分[5]研究外,尚未见有关花粉生活力方面的研究报道。本研究通过对云南松花粉储藏温度及萌发条件的探究,旨在为云南松杂交育种工作的顺利开展提供技术依据。
-
云南松花粉采自四川省会理县太平镇云南松人工林。
-
2014年4月下旬采集成熟的云南松雄球花,置于光滑纸面上,在室内摊晾收集,花粉混合均匀后装入牛皮纸袋中带回实验室。
-
将花粉分装在清洁干燥带瓶盖的青霉素小瓶中,标记,分别置于25 ℃,5 ℃,-20 ℃条件下储藏。储藏初期隔24 h测定3种储藏温度下花粉的生活力,后期可根据情况延长测定间隔时间,直到花粉失去生活力(花粉生活力低于50%)。
-
采用碘-碘化钾染色法、α-萘酚-联苯胺法、次甲基蓝染色法等3种方法测定花粉生活力,筛选出能快速准确测定花粉生活力的方法。测定时,先将低温条件下的云南松花粉在常温下放置0.5 h,再用染色剂处理,重复3个·处理-1,观察清晰视野5个·重复-1,统计花粉粒数≥30粒·视野-1,统计各视野花粉生活力,并计算各处理平均生活力。花粉生活力(%)=(染色花粉数/统计花粉总数)×100%。
-
设置培养液蔗糖质量浓度为0,50,80,100,120 g· L-1;硼酸质量浓度为0,100,300,500 mg· L-1;赤霉素质量浓度为0,50,100,150 mg· L-1;氯化钙质量浓度为0,50,100,150 mg· L-1。通过单因素实验,筛选出培养液各因素的最适质量浓度,再进行正交实验设计,筛选出培养液各因素最佳质量浓度组合。花粉采用液体培养法进行培养,先将解剖针和双凹片干燥灭菌,处理时用解剖针蘸取少量花粉散播在载有不同培养液的双凹片凹孔中,盖上盖玻片,置于25 ℃LHP-300H智能型人工气候箱中黑暗培养。3 h在显微镜下观测1次,重复3次·处理-1,观察清晰视野5个·重复-1,统计花粉粒数≥30粒·视野-1,当花粉管长度≥花粉直径时视为花粉萌发,观察至萌发率不再增加为止。统计各视野花粉生活力,并计算各处理平均生活力。花粉萌发率(%)=(花粉萌发数/统计花粉总数)×100%。
-
所得数据用Excel,SPSS 20进行统计分析。进行方差分析时,需对原始数据进行反正弦转换。
-
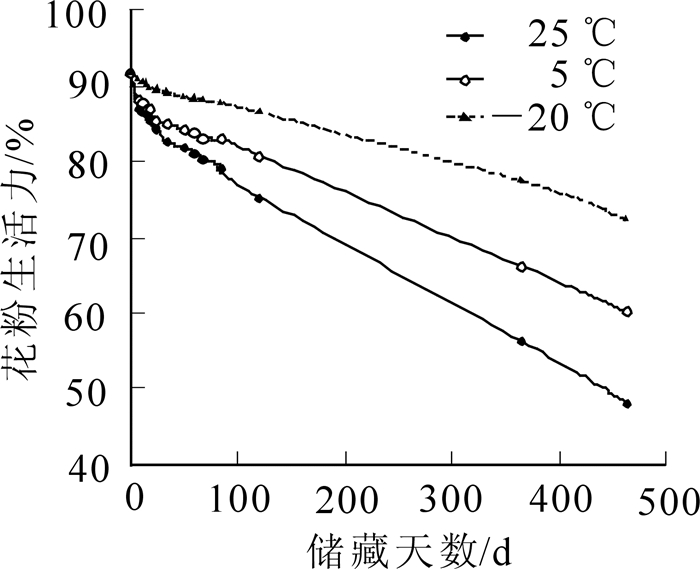
a-萘酚-联苯胺法染色不明显,次甲基蓝染色法在进行染色梯度界定时无固定标准,难统一,而经碘-碘化钾染色法测定结果稳定、易辨别、最接近萌发实验结果,因此,本研究以碘-碘化钾染色法测定结果作为论述依据。测定结果(图 1)显示:随着储藏天数的增加,3种储藏温度下云南松花粉生活力均下降,但储藏温度不同,花粉生活力下降的速率不同,其中25 ℃条件下花粉生活力下降较快,5 ℃条件和-20 ℃条件下花粉生活力下降趋势都较为平缓,总的花粉生活力下降趋势表现为25 ℃>5 ℃>-20 ℃。方差分析结果表明:储藏时间和储藏温度对花粉生活力的影响达极显著水平(F储藏时间=35.482>F0.01;F储藏温度=30.404>F0.01),且储藏温度的F值较储藏时间的F值大,说明储藏温度比储藏时间对花粉生活力的影响大。多重比较显示(表 1):3种储藏温度对云南松花粉生活力影响显著,均数大小表现为-20 ℃>5 ℃>25 ℃,即表明-20 ℃条件对花粉生活力的保持显著优于5 ℃条件和25 ℃条件,5 ℃条件显著优于25℃条件。

图 1 不同储藏温度下花粉生活力的比较
Figure 1. Comparison of pollen viability under different storage conditions
表 1 均衡子集表
Table 1. Balanced subsets table
储藏温度/℃ 均衡子集 1 2 3 25 63.377 9 5 65.636 4 -20 69.357 9 -
云南松花粉萌发过程见图 2。云南松花粉在培养24 h之前不萌发,培养24~72 h,萌发率急剧上升,72 h后萌发率趋于平稳,超过72 h以后新增花粉萌发数极少,可见,72 h后即可统计花粉萌发率。花粉粒萌动时萌发沟薄壁区呈透明状态并向两边突起,继而其中一端或两端伸长成为花粉管,花粉管生长后期会分支,甚至破裂。

图 2 云南松花粉萌发过程
Figure 2. Mination process of Pinus yunnanesis pollen
-
由图 3A可知:蔗糖对云南松花粉萌发存在极显著影响。花粉在清水中也能萌发,平均萌发率为9.02%,随着蔗糖质量浓度的增加,花粉萌发率呈先升高后降低的趋势,在蔗糖质量浓度为80 g· L-1时最高,为82.41%。蔗糖质量浓度超过80 g· L-1后花粉萌发率降低,可见最适蔗糖质量浓度为80 g· L-1。由图 3B可知:不同质量浓度的氯化钙对云南松花粉萌发率影响程度不同。当培养基氯化钙质量浓度为50 mg· L-1时,萌发率为13.06%;随氯化钙质量浓度的增加花粉萌发率逐渐升高,当氯化钙质量浓度升至100 mg· L-1时,萌发率达到最高,为56.98%;但随着氯化钙质量浓度继续增加,花粉萌发率下降。可见云南松花粉萌发的适宜氯化钙质量浓度是100 mg· L-1。由图 3C可知:硼酸质量浓度为300 mg· L-1时,花粉萌发率达到最大值18.23%,极显著高于对照和100 mg· L-1;而质量浓度为500 mg· L-1时,花粉萌发受到强烈抑制,萌发率仅为1.35%,极显著低于清水处理。结果表明:云南松花粉萌发的较适宜硼酸质量浓度为300 mg· L-1。由图 3D可知:当培养基赤霉素质量浓度为100 mg· L-1时,花粉萌发率达到最大值66.41%,极显著高于其他处理。蔗糖、氯化钙、硼酸、赤霉素等单因子对云南松花粉的萌发都存在极显著影响,添加后大部分萌发率比对照有显著增长,以蔗糖作用最明显。单因子实验结果显示最适浓度组合为80 g· L-1蔗糖+100 mg· L-1赤霉素+100 mg· L-1氯化钙+300 mg· L-1硼酸。

图 3 蔗糖、氯化钙、硼酸、赤霉素对云南松花粉萌发的影响
Figure 3. Effect of sucrose, CaCl2, H3BO3 and GA3 on in vitro pollen germinetion of Pinus yunnanesis
-
表 2结果表明:蔗糖、硼酸、赤霉素、氯化钙在一定质量浓度范围内共同作用能明显提高云南松花粉的萌发率。实验中各组处理对花粉萌发率的影响与对照组都存在极显著差异,其中处理1萌发率最高,达89.74%;其次为处理2,达77.65%。正交实验结果显示最优搭配为120 g· L-1蔗糖+150 mg· L-1赤霉素+100 mg· L-1氯化钙+100 mg· L-1硼酸,与单因子实验所得结果有差异,这说明各种营养因子对花粉萌发的影响是相互作用的。
表 2 花粉萌发L9(34)正交实验结果
Table 2. Pollen germination rate in orthogonal design experiment
处理编号 蔗糖/
(g·L-1)硼酸/
(mg·L-1)赤霉素/
(mg·L-1)氯化钙/
(mg·L-1)萌发率/% 1 120 100 150 100 89.74 aA 2 80 500 150 150 77.65 bB 3 80 300 100 100 54.69 cC 4 120 500 100 50 49.42 dD 5 100 300 150 50 46.15 eD 6 120 300 50 150 38.17 fE 7 100 500 50 100 34.66 gE 8 100 100 100 150 28.49 hF 9 80 100 50 50 15.09 iG 0(对照) 00 0 0 0 8.20 jH 说明:不同大写字母表示差异极显著(P<0.01);不同小写字母表示差异显著(P<0.05)。 -
研究表明:碘-碘化钾染色法能反应不同储藏温度下花粉生活力的变化趋势。染色法测定的花粉生活力略高于培养基法,这是因为染色时染色剂也能对未成熟、衰老和败育的花粉进行染色,而这类花粉不一定都具有受精能力,以致测定结果往往偏高。虽然培养基法是最能准确测定花粉生活力的方法,但是其耗时长,操作过程相对复杂,碘-碘化钾染色法能快速准确地测定云南松花粉的生活力。
报道显示[6-8]:银杉Cathaya argyrophylla等裸子植物的鲜花粉生活力均在90%以上。本研究通过碘-碘化钾染色法测定云南松鲜花粉的生活力为91.77%,与以上裸子植物结果相一致,都具有较高的鲜花粉生活力。本研究还表明:云南松花粉具有较高的耐储藏性,在25 ℃条件下储藏463 d时云南松花粉生活力为48.21%,在5 ℃条件下储藏463 d时为60.28%,在-20 ℃条件下463 d时为72.34%。储藏时间和储藏温度对于花粉生活力都有极显著的影响,但储藏温度影响更大。这是因为低温能降低花粉呼吸强度,减少自身营养物质的消耗。
-
蔗糖是许多植物花粉离体培养所必需的营养成分,一般认为蔗糖为花粉的萌发和花粉管的生长提供了能源和碳源,同时还能调节培养环境渗透压。蔗糖适宜质量浓度通常为100~150 g· L-1,云南松花粉萌发的最适宜蔗糖质量浓度为80 g· L-1,且总体情况是低质量浓度比高质量浓度对花粉萌发更有利。其原因可能是高质量浓度蔗糖造成花粉细胞质壁分离,使花粉粒脱水,从而影响花粉的萌发。钙离子是花粉管生长的基本因素。本研究中单因子氯化钙质量浓度对花粉萌发的影响极显著,低质量浓度时对花粉萌发率的影响不大,质量浓度增加能显著提高花粉萌发率,100 mg· L-1时萌发率达最大值56.98%。可能云南松花粉自身含有较少的钙离子,萌发时外源钙离子的适量补充可以促进其萌发。硼不仅可以增加糖的吸收、运转和代谢,形成花粉管顶端生长依赖的梯度,还可能作为一种相关因子影响关键酶活性,改变细胞壁延展性以至影响花粉的萌发、花粉管细胞壁的构建和花粉管的伸长[9]。一般认为,花粉内存在较多的钙,而缺乏硼,因硼的缺乏往往要从柱头和花柱内获得硼的补偿[10]。本研究中,云南松花粉萌发对硼需要量十分微小,在最适质量浓度300 mg· L-1时萌发率仅为18.23%,500 mg· L-1时即表现为强烈的抑制作用,可能花粉内含有较多的硼元素,能在一定程度上满足花粉萌发所需,但硼适量的补充也能提高花粉萌发率。虽然目前有关赤霉素等植物生长调节物质能促进花粉的萌发和花粉管生长的机制尚不清楚,但已有研究表明花粉自身富含生长素及赤霉素等生长调节物质,这些物质的多少对花粉自身的萌发、生长及储藏寿命均有较大的影响,对花粉管在花柱内生长完成受精及促进果实膨大也有重要的作用,因此,一定质量浓度的外源赤霉素处理能促进花粉萌发和花粉管生长[11]。在本研究中赤霉素对花粉萌发率存在极显著性影响,以100 mg· L-1最佳,萌发率可达66.41%。总体来说,云南松花粉生活力强,对培养基营养物质的需求量不高,单因子以蔗糖作用最明显,正交实验最适的液体培养基组合为120 g· L-1+150 mg· L-1赤霉素+100 mg· L-1氯化钙+100 mg· L-1硼酸。这可能是因为云南松花粉中自身含有的能量物质较多,适量补充能更好地满足萌发所需营养物质,高质量浓度的营养物质补充反而会抑制花粉的正常萌发。
在杂交授粉实际操作中,一般采用生活力不低于新鲜花粉生活力的50%的花粉进行授粉,因此,云南松花粉在25 ℃条件下储藏1 a之内可用于授粉,低温储藏能有效延缓花粉生活力下降的速度。在授粉时可以喷施一定质量浓度的蔗糖、赤霉素,促进花粉萌发和花粉管伸长,进而提高杂交育种成效。
Storage temperature and pollen viability of Pinus yunnanensis
-
摘要: 以云南松Pinus yunnanesis花粉为材料,通过设置3种储藏温度(25℃,5℃,-20℃)探讨花粉生活力随时间的变化规律;通过单因子实验和正交实验,探究不同质量浓度的蔗糖、硼酸、赤霉素、氯化钙单独或共同作用对花粉萌发的影响。结果表明:云南松花粉储藏0 d时花粉生活力为91.77%,随着储藏时间的增加3种储藏温度的花粉生活力均下降,下降速率表现为25℃ > 5℃ > -20℃,说明低温能增加花粉的耐储性。单因子萌发实验显示:蔗糖的最适质量浓度为80 g·L-1,萌发率为82.41%;赤霉素的最适质量浓度为100 mg·L-1,萌发率为66.41%;硼酸的最适质量浓度为300 mg·L-1,萌发率为18.23%;氯化钙的最适质量浓度为100 mg·L-1,萌发率为56.98%。正交实验显示:4种因素的最佳组合为120 g·L-1蔗糖+150 mg·L-1赤霉素+100 mg·L-1氯化钙+100 mg·L-1硼酸,萌发率可达89.74%。Abstract: To determine the storage temperature and pollen viability of Pinus yunnanensis, different storage conditions of 25℃, 5℃, and -20℃ were used to study variation in pollen viability. Different treatments of sucrose, H3BO3, GA3, and CaCl2 were used to determine the effects on pollen germination. Analysis employed orthogonal experiments alone as well as interactions. Results showed that pollen viability was 91.8% at 0 d and then decreased with greater storage time at temperatures of 25℃ > 5℃ > -20℃. Germination tests showed that the optimal concentration and the corresponding germination rate were as follows:sucrose-80 g·L-1 and 82.4%, GA3-100 mg·L-1 and 66.4%, H3BO3-300 mg·L-1 and 18.2%, and CaCl2-100 mg·L-1 and 57.0%. The best combination of four factors was 120 g·L-1 sucrose+150 mg·L-1 GA3+100 mg·L-1 H3BO3+100 mg·L-1 CaCl2 with a pollen germination rate of 89.7%. Meanwhile, above a certain concentration of the culture medium had inhibitory effects on pollen germination. Thus, low temperatures increased pollen storage time.
-
Key words:
- forest tree breeding /
- Pinus yunnanensis /
- pollen /
- storage temperature /
- viability /
- germination
-
图 1 不同储藏温度下花粉生活力的比较
Figure 1 Comparison of pollen viability under different storage conditions
图 3 蔗糖、氯化钙、硼酸、赤霉素对云南松花粉萌发的影响
Figure 3 Effect of sucrose, CaCl2, H3BO3 and GA3 on in vitro pollen germinetion of Pinus yunnanesis
表 1 均衡子集表
Table 1. Balanced subsets table
储藏温度/℃ 均衡子集 1 2 3 25 63.377 9 5 65.636 4 -20 69.357 9  下载: 导出CSV
下载: 导出CSV
表 2 花粉萌发L9(34)正交实验结果
Table 2. Pollen germination rate in orthogonal design experiment
处理编号 蔗糖/
(g·L-1)硼酸/
(mg·L-1)赤霉素/
(mg·L-1)氯化钙/
(mg·L-1)萌发率/% 1 120 100 150 100 89.74 aA 2 80 500 150 150 77.65 bB 3 80 300 100 100 54.69 cC 4 120 500 100 50 49.42 dD 5 100 300 150 50 46.15 eD 6 120 300 50 150 38.17 fE 7 100 500 50 100 34.66 gE 8 100 100 100 150 28.49 hF 9 80 100 50 50 15.09 iG 0(对照) 00 0 0 0 8.20 jH 说明:不同大写字母表示差异极显著(P<0.01);不同小写字母表示差异显著(P<0.05)。
下载: 导出CSV
-
[1] 黄瑞复.云南松的种群遗传与进化[J].云南大学学报(自然科学版), 1993, 15(1):50-63. HUANG Ruifu. The population genetics and evolution of Pinus yunnanensis[J]. J Yunnan Univ Nat Sci, 1993, 15(1):50-63. [2] 陈强, 常恩福, 董福美, 等.云南松天然优良林分疏伐营建母树林的研究[J].云南林业科技, 2000(3):1-8. CHEN Qiang, CHANG Enfu, DONG Fumei, et al. Study on establishment of seed production stand by thinning natural superior stands of Pinus yunnanensis[J]. J Yunnan For Sci Technol, 2000(3):1-8. [3] 张跃敏, 李根前, 李莲芳, 等.云南松生长特性及其促成培育[J].陕西林业科技, 2008(3):4-7. ZHANG Yuemin, LI Genqian, LI Lianfang, et al. The growth characteristics of Pinus yunnanensis and improvement measures[J]. J Shaanxi For Sci Technol, 2008(3):4-7. [4] 杨彩云, 杨貌仙.云南松花粉形态研究[J].云南植物研究, 1989, 11(1):71-74. YANG Caiyun, YANG Maoxian. Study on the pollen morphology of Pinus yunnanensis[J]. Acta Bot Yunnan, 1989, 11(1):71-74. [5] 范国栋, 刘嘉宝, 冯武.破壁云南松花粉成分分析及其安全性和功能评价[J].云南林业科技, 2001(1):54-57. FAN Guodong, LIU Jiabao, FENG Wu. Evaluation on ingredients, security and function of coat-broken Pinus yunnanensis pollen[J]. J Yunnan For Sci Technol, 2001(1):54-57. [6] 王红卫, 邓辉胜, 谭海明, 等.银杉花粉生命力及其变异[J].植物生态学报, 2007, 31(6):1199-1204. WANG Hongwei, DENG Huisheng, TAN Haiming, et al. Pollen viability and variation in Cathaya argyrophylla[J]. J Plant Ecol, 2007, 31(6):1199-1204. [7] 马建伟, 王军辉, 张守攻, 等.云杉花粉的贮藏及萌发研究[J].林业科学研究, 2012, 25(3):302-307. MA Jianwei, WANG Junhui, ZHANG Shougong, et al. Study on storage and bourgeoning of spruce pollen[J]. For Res, 2012, 25(3):302-307. [8] 孙蕾, 房用, 马玲, 等.黑松和赤松花粉取粉处理和干燥贮藏技术研究[J].食品科学, 2007, 28(7):189-191. SUN Lei, FANG Yong, MA Ling, et al. Study on production and storage technology of black pines and Japanese red pines pollens[J]. Food Sci, 2007, 28(7):189-191. [9] 龚明, 曹宗巽.钙和钙调素对花粉萌发和花粉管生长的调控[J].植物生理学通讯, 1995, 31(5):321-328. GONG Ming, CAO Zongxun. Regulation of calcium and calmodulin on pollen germination and growth of pollen tube[J]. Plant Physiol Commun, 1995, 31(5):321-328. [10] 张绍铃, 陈迪新, 康琅, 等.培养基组分及pH值对梨花粉萌发和花粉管生长的影响[J].西北植物学报, 2005, 25(2):225-230. ZHANG Shaoling, CHEN Dixin, KANG Lang, et al. Effects of medium components and pH on pollen germination and tube growth in pear (Pyrus pyrifolia)[J]. Acta Bot Boreal-Occident Sin, 2005, 25(2):225-230. [11] 张绍铃, 高付永, 陈迪新, 等.植物生长调节物质对丰水梨花粉萌发和花粉管生长的影响[J].西北植物学报, 2003, 23(4):586-591. ZHANG Shaoling, GAO Fuyong, CHEN Dixin, et al. The effects of plant growth regulating substances on pollen germination and tube gowth in Fengshui pear (Pyrus serotina)[J]. Acta Bot Boreal-Occident Sin, 2003, 23(4):586-591. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.01.010
 点击查看大图
点击查看大图
计量
- 文章访问数: 3671
- HTML全文浏览量: 732
- PDF下载量: 416
- 被引次数: 0
