




-
植被数量分析作为生态学研究方法中的重要组成部分,在反映生态规律上具有客观性及准确性的特点[1-2]。数量分类和排序在数量分析方法中使用频繁,能够合理地揭示植物种间、群落与环境间的生态关系,在植被分析中被广泛应用[3-4]。自组织特征映射网络属于人工神经网络,是在研究人脑神经元的生物行为基础之上,通过数学模型化而得到的人工信息处理模型[5]。自组织特征映射网络(self-organizing feature map,SOFM或SOM),其非线性拟合水平突出,适用于聚类分析以及研究数据信息的相关性[6]。SOM能够人工模拟人脑神经元的生物学行为,是一种高度非线性的人工信息处理模型[7]。而植物物种、植物群落与环境间的生态关系是非线性的[8],通过非监督自主学习的方式,SOM能够解决复杂的非线性问题,在生态学研究中具有一定的优势和应用潜力[9]。宝华玉兰Magnolia zenii 1933年由郑万钧在宝华山发现[10],开花时素雅秀丽,芬芳馥郁,是一种极佳的观花植物。现阶段其成年植株的数量以及野生幼苗都存在严重的缺失[11],成年个体仅有34株[12],野生种仅在宝华山国家森林公园内有分布。宝华玉兰具有植株散生、种群数量较低、种间竞争存在较大压力[13]、分布范围小的特点[14],1999年被列为国家Ⅱ级保护植物,2004被列为极危种(CR)[15-16]。为此,通过自组织特征网络对群落进行数量分类与排序,探究宝华玉兰生存群落的分布格局、其野生种群的分布规律以及与环境因子的关系,以期为保护和管理宝华玉兰种群野生种质资源提供理论依据和新的方法。
-
江苏省镇江句容市宝华山(31°37'~32°19'N,118°58'~119°58'E),山体坐落于宁镇山脉的中段。1981年江苏省批准设立了江苏宝华山省级自然保护区;1996年批准设立了江苏宝华山国家森林公园。宝华山位于北亚热带季风气候区,水热充沛,全年降水量为961.0 mm,季节性显著,年平均气温为15.4 ℃,地带性土壤为黄棕壤[17],土层较厚,适宜亚热带植物生长[18]。常绿落叶阔叶混交林是宝华山主要的地带性植被,区域内的植物种类丰富,共有维管植物168科642属1 232种[19],主要优势树种包括青冈Cyclobalanopsis glauca,紫楠Phoebe sheareri,冬青Ilex chinensis,栓皮栎Quercus variabilis,朴树Celtis sinensis,麻栎Quercus acutissima,建始槭Acer henryi等。
-
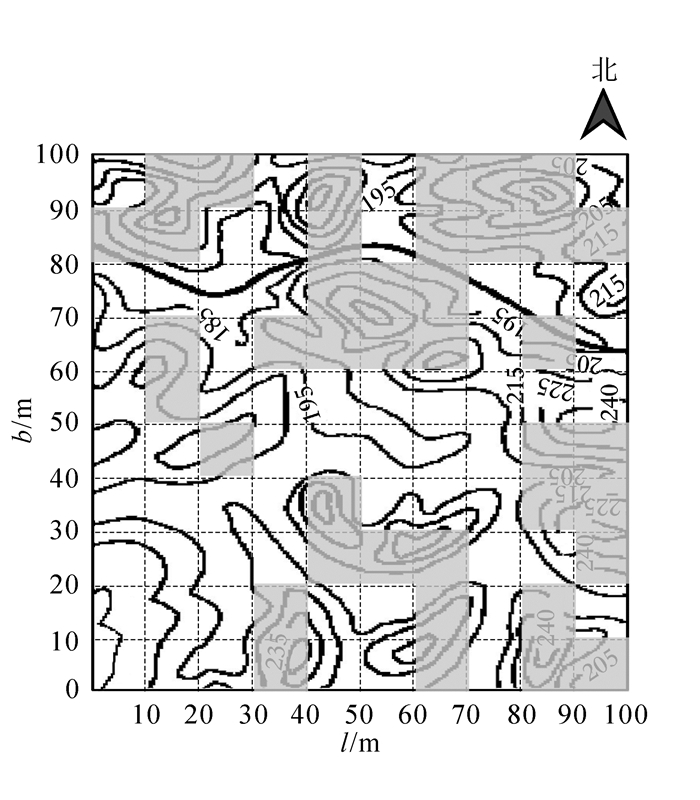
在宝华山1万m2的固定样地中,选择宝华玉兰天然种群的分布区域,共设置了40个大样方(10 m × 10 m)(图 1中灰色区域代表样方位置),并设置灌木样方2个(5 m × 5 m)·样方-1,草本样方4个(1 m × 1 m)·样方-1。测量群落中乔木树种(胸径>1 cm)的胸径、树高、株数;灌木层测量树高、株数(胸径<1 cm的小乔木或幼苗);统计草本层(包含蕨类植物)和层间植物的植物种类以及生长高度、盖度[20]。

图 1 宝华玉兰生存群落40个样方分布图
Figure 1. 40 samples of Magnolia zenii living communities
-
使用手持全球定位系统(GPS)测量海拔、经纬度;使用罗盘仪测量坡度、坡位以及坡向;凋落物层厚度使用土钎及钢直尺测量;土壤原位pH计采集土壤pH值;数字等级用于代表坡位和坡向[21]:上坡位1,中坡位为2,下坡位为3,坡向以正北为0°,隔45°设置1个等级,顺时针旋转分成不同的等级,序号代表数字等级(表 1)。
表 1 8个坡向等级
Table 1. Eight levels of aspect
序号 坡向 方位角 1 北坡 0°~ 22.5°,337.5°~360.0° 2 东北坡 22.5°~67.5° 3 西北坡 292.5°~337.5° 4 东坡 67.5°~112.5° 5 西坡 247.5°~292.5° 6 东南坡 112.5°~157.5° 7 西南坡 202.5°~247.5° 8 南坡 157.5°~202.5° -
对宝华山40个样方中维管植物的科、属、种进行数量统计。物种统计中,蕨类植物和被子植物依据秦仁昌系统(1978)以及哈钦松系统(1973)。
-
计算公式为[22]:VIV1=[d+p+f]/3×100%;VIV2=[d+h+f]/3×100%;VIV3=[h+c+f]/3×100%。其中:VIVi代表重要值,i=1代表乔木层,i=2代表灌木层,i=3代表草本层;d代表相对密度;h代表相对高度;p代表相对优势度;c代表相对盖度;f代表相对频度。
-
SOM模型由单层神经元网络组成,实质是一种从高维空间信息数据到低维空间的非线性映射[23],使用无监督学习,产生一个有序的拓扑输出从而呈现样方间相似性。SOM由输出层及输入层(竞争层)2层结构组成,其中输入层包含植被数据信息中每个变量(物种)的单元(神经元)[24]。输出层每个单元都通过加权连接与输入层中的每个单元连接,连接强度由权重向量确定。输出单元的横向交互确保了学习的竞争性,能够适应不同位置的不同输入。根据特征映射确定输出空间的最佳匹配单元,其相应的权重向量(最佳匹配单元在输出空间的映射坐标)根据邻近神经元进行更新调整,直至样本的分布显示出规律性,形成稳定的聚集状态。因此,相似的样方应与输出层中临近的单元相关联,而不相似的样方与输出层中其他地方的远处单元相关联[25-26]。使用MATLAB 6.1开发的SOM工具箱编程实现SOM的聚类和排序。物种数据矩阵训练图根据k-均值法进行聚类分析,采用Davies-Bouldin指数(DBI)来确定聚类是否有效。用色度对比度实现数据单元之间的欧几里得距离的U矩阵。SOM的计算方法前人研究较多[27-29],本文不再赘述。
-
在40个样方中,维管植物共计62科108属140种;其中蕨类植物共计4科5属5种。群落中科的组成以壳斗科Fagaceae,蔷薇科Rosaceae,榆科Ulmaceae,禾本科Gramineae,百合科Liliaceae等为主。其中乔木层共有植物39种,灌木层共计48种,草本层共计53种。单科单属共计39个,占总科数的27.86%,群落中寡属种占据比例较大。
-
在所调查的宝华玉兰生存群落中,表 2中仅列出重要值>2.00%优势种。乔木层的植物物种中,优势种共计9种。宝华玉兰的种群数量优势不明显,相对密度仅为1.32%。其相对频度与紫楠,油茶Camellia oleifera,建始槭相同,分布较为均匀,但相对优势度只有0.97%,说明宝华玉兰的相对新生的比例较低,更新速度缓慢。乔木层中紫楠重要值排第1位,是群落主要的建群种,占据主要的生存空间与资源。组成宝华玉兰生存群落中的主要灌木(重要值>2.00%)物种共9种。其中紫楠、建始槭、油茶、老鸦柿Diospyros rhombifolia和青冈等为乔木树种,并且在其对应的乔木层中的重要值靠前,属于优势物种。在将来可能为其乔木增加优势度,与宝华玉兰形成竞争。同时,宝华玉兰在灌木层中的重要值排名靠后,0.33%的相对密度和0.88%的相对频度说明群落中宝华玉兰的幼株较少,种群更新慢,幼株在群落中处于劣势地位。
表 2 宝华山宝华玉兰生存群落重要值
Table 2. Importance values of Magnolia zenii living communities in Baohua Mountain
层次 重要值排名 树种 相对密度/相对高度 相对频度 相对优势度/相对盖度 重要值/% 乔木层 1 紫楠Phoebe sheareri 51.07 6.80 77.56 45.15 2 建始槭Acer henryi 11.06 6.80 10.51 9.46 3 青冈Cyclobalanopsis glauca 8.91 6.12 3.74 6.26 4 红枝柴Meliosma oldhamii 3.55 5.44 2.71 3.90 5 油茶Camellia oleifera 3.55 6.80 0.25 3.53 6 宝华玉兰Magnolia zenii 1.32 6.80 0.97 3.03 7 枫香树Liquidambar formosana 1.40 5.44 2.04 2.96 8 琅玡榆Ulmus chenmoui 1.40 5.44 0.26 2.37 9 枳椇Hovenia acerba 1.32 4.76 0.24 2.11 灌木层 1 紫楠Phoebe sheareri 36.18 8.77 31.36 25.44 2 建始槭Acer henryi 56.18 8.77 61.36 42.11 3 络石Trachelospermum jasminoides 10.41 8.77 9.60 9.59 4 油茶Camellia oleifera 4.43 8.77 6.69 6.63 5 青冈Cyclobalanopsis glauca 5.46 8.77 3.84 6.02 6 老鸦柿Diospyros rhombifolia 4.25 7.89 4.77 5.64 7 卫矛Euonymus alatus 4.29 7.02 4.02 5.11 8 野蔷薇Rosa multiflora 0.98 6.14 0.36 2.49 9 薜荔Ficus pumila 2.01 3.51 0.84 2.12 22 宝华玉兰Magnolia zenii 2.89 2.63 0.67 2.07 草本层 1 阔叶山麦冬Liriope platyphylla 14.17 4.55 27.09 15.27 2 堇菜Viola verecunda 5.96 2.27 33.04 13.76 3 淡竹叶Lophatherum gracile 16.55 4.55 14.21 11.77 4 中国石蒜Lycoris chinensis 15.43 11.36 3.19 10.00 5 天葵Semiaquilegia adoxoides 11.48 2.27 11.29 8.35 6 贯众Cyrtomium fortunei 3.92 11.36 1.76 5.68 7 沿阶草Ophiopogon bodinieri 2.83 11.36 0.50 4.90 8 长柄山蚂蝗Podocarpium podocarpum 3.73 6.82 1.21 3.92 9 知风草Eragrostis ferruginea 5.37 4.55 1.32 3.75 10 钩腺大戟Euphorbia sieboldiana 4.47 4.55 0.83 3.28 11 渐尖毛蕨Cyclosorus acuminatus 1.94 6.82 0.61 3.12 12 点腺过路黄Lysimachia hemsleyana 2.68 2.27 2.09 2.35 13 蘘荷Zingiber mioga 1.79 4.55 0.22 2.19 14 求米草Oplismenus undulatifolius 3.58 2.27 0.50 2.12 宝华玉兰种群的常见伴生草本(重要值>2.00%)物种共14种。草本中以百合科Liliaceae以及鳞毛蕨科Dryopteridaceae居多,其余多为单科单属。单科单属虽然在科属比例上占优势,丰富了整个群落的多样性,但数量较少,容易被替代。
-
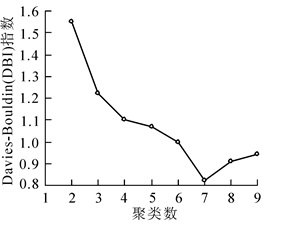
依据k-均值聚类规则DBI指数,SOM将调查样方总共分为7块区域,对应了宝华玉兰7个不同的群落类型(图 2)。

图 2 DBI指数随NOC值变化图
Figure 2. Change trands of Davies-Bouldin index based on number of clusters
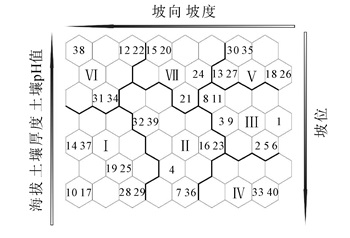
经SOM学习训练后挑选出量化误差和拓扑误差最低的7×8的拓扑结构为最终输出层,得到40个样方的得到拓扑映射图(图 3)。①群落Ⅰ:紫楠+油茶+红枝柴-红枝柴+老鸦柿+油茶-阔叶山麦冬+渐尖毛蕨。②群落Ⅱ:油茶+建始槭+红枝柴-油茶+紫楠+建始槭-堇菜+阔叶山麦冬+络石。③群落Ⅲ:紫楠+青冈+枳椇-青冈+紫楠-堇菜+沿阶草+淡竹叶。④群落Ⅳ:青冈+老鸦柿+紫楠-青冈+紫楠-阔叶山麦冬+淡竹叶+中国石蒜。⑤群落Ⅴ:老鸦柿+宝华玉兰+青冈-老鸦柿+青冈-阔叶山麦冬+淡竹叶。⑥群落Ⅵ:建始槭+紫楠+老鸦柿-紫楠+建始槭-贯众+络石+天葵。⑦群落Ⅶ:紫楠+建始槭+青冈-紫楠+红枝柴+老鸦柿-阔叶山麦冬+贯众+络石。

图 3 宝华玉兰生存群落40个样方拓扑映射图
Figure 3. Topology map of 40 samples of Magnolia zenii living communities
-
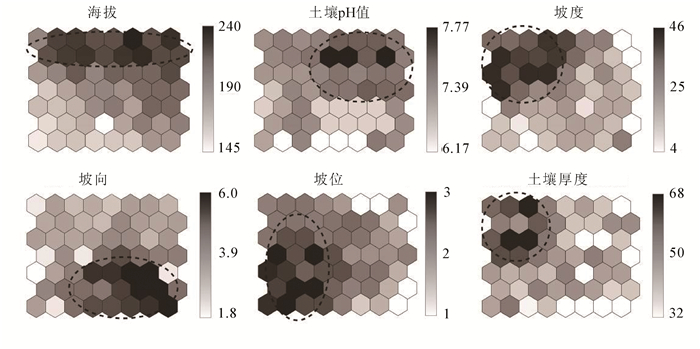
SOM映射上的环境因子平均值呈现了在排序空间上环境的变化梯度,颜色深浅与平均值大小相关,颜色越浅则值越小(图 4)。排序图中纵轴下方到上方的过程主要表明了海拔的逐渐升高以及坡位从上坡位向下坡位的逐渐过渡,但由于海拔变化范围小,对群落的影响不明显,所以海拔不是主要的影响因子;坡度最大值在左侧,从左至右坡度逐渐变缓;坡向为1~6,主要以阴坡和半阴坡为主,右侧达到最大值,为半阳坡;土壤厚度在排序图左侧达到最大值;土壤pH值在排序上显示在右侧中上部,变化范围相较于坡向、坡度不明显。通过SOM排序,环境因子平均值在图上的变化能够得出,坡向和坡度对宝华玉兰生存群落分布格局具有主导作用,是主要的环境因子,而海拔、土壤pH值以及土壤厚度的作用较小。

图 4 SOM排序空间的环境梯度
Figure 4. Environmental gradients of SOM ordination space
综合群落类型分布和环境因子的梯度变化结果,排序图由下至上主要表明了海拔、土壤厚度、土壤pH值、坡位的变化;排序图从左侧至右侧逐渐变为缓坡,坡向由阴到半阳的变化趋势。纵轴和横轴的梯度变化反映了宝华玉兰生存群落空间分布的环境变化。
-
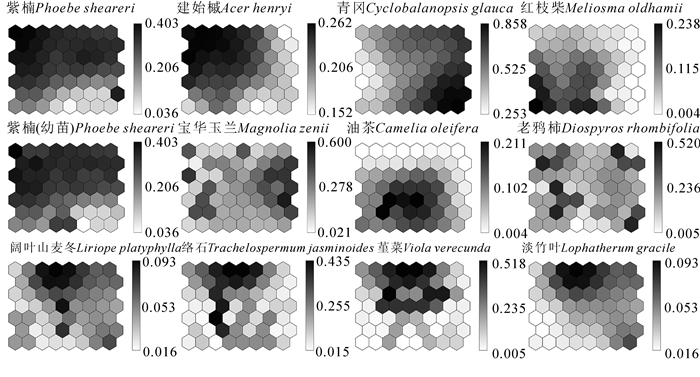
图 5(图例数字代表重要值)是宝华玉兰生存群落中乔木层、灌木层、草本层优势种以及宝华玉兰在排序图中的分布情况。根据SOM的分类结果,群落中优势种分布中心与相对应的群落类型分布之间相似程度较高。虽然紫楠分布最广泛,但显示在优势种排序图中的分布中心对应是群落Ⅵ类型,与该类型的灌木优势种类以及分布位置相同,建始槭的分布中心与该类型乔木优势种类分布位置相似,而青冈的分布中心在图中对应的是群落Ⅳ类型,络石和群落Ⅶ,油茶和群落Ⅱ也是具有相似的分布位置,能够表明群落类型的分布情况在一定程度上是由优势种的分布所决定的。红枝柴、阔叶山麦冬、淡竹叶分布范围广泛但也有相对集中的分布中心。宝华玉兰在图中分布区域较为均匀,但在最左侧和最右侧有少量集中的现象。将图 5的优势种在排序图中集中分布的区域结合图 4环境因子在排序图上的梯度变化,可以综合分析环境因子对群落分布格局的影响。

图 5 优势种在SOM排序图上的梯度分布
Figure 5. Gradient distributions of dominant species on the SOM ordination diagram
宝华玉兰在调查范围内分布较为均匀,但在排序图中在最左侧和最右侧有少量的集中,而左侧主要为较陡的阴坡,右侧主要为较缓半阳坡。阴坡主要是以紫楠、建始槭为优势种,有少量的宝华玉兰伴生。分析原因可能是由于宝华玉兰耐荫性相较紫楠和建始槭等都要弱,因此宝华玉兰出现了主要分布在半阳坡的现象。而紫楠的成年株和幼苗都较多地分布在湿冷的阴坡,建始槭也是优势种,占据了群落的主要环境资源,种间竞争强。而最右侧的青冈主要分布在排序图 5右下角,对宝华玉兰影响较小,所以反映在SOM排序图上则是宝华玉兰更多集中分布于图中的右半部分,而左半部分的分布数量较少。分析原因可能是由于宝华玉兰虽然为喜阳树种,但在种子萌发及幼苗时期需要阴凉的环境,随着幼树生长至成年植物耐荫性逐渐变弱,因此不同生长期需要不同的生境可能导致了宝华玉兰的濒危现状。
-
根据群落重要值分析,群落中的紫楠、建始槭等处于绝对优势地位,占据了群落主要环境资源,而宝华玉兰野生种群成年植株较少,并且幼株数量稀缺,种群更新慢,不利于未来种群增长和发展。通过SOM对宝华玉兰生存群落的分类和排序,将40个样方划分为了7个不同的群落类型,在拓扑排序图上不同群落类型间有着明显的分布范围和不同优势种分布,紫楠等优势种分布的集中区域与对应的群落类型的乔木或灌木优势种分布具有高度的相似性,说明了群落的分布格局在一定程度上是由优势种的分布情况所决定的,能够合理反映群落间生态关系。
非生物环境因素能够影响群落结构以及分布格局[30]。通过对宝华玉兰生存群落空间的环境因子在拓扑图上的排序,分析了群落优势种的分布特点以及宝华玉兰的分布情况。排序图上的结果表明:宝华玉兰天然种群分布于坡度较缓的半阳坡,而紫楠、建始槭等群落优势种分布较广,在陡坡和缓坡均有分布,特别是紫楠的成株及幼苗在阴坡至半阳坡也有大量分布。在实际的调查过程中发现,群落多处于山坡或是沟谷地带。坡度和坡向的变化对群落分布环境中的水热资源分配产生了重要作用,对宝华玉兰种群的分布产生了影响。在坡度较陡的地区岩石裸露,常见以紫楠为优势种的常绿阔叶林,宝华玉兰仅有极少成年株分布。分析原因可能是由于紫楠的萌生性较强,且耐荫耐旱能力都较高,能够适应恶劣的生境条件。宝华玉兰是明显的直根系和深根系,成株由于根系较长,能够从岩石下的土层或地下径流获得养分和水分,而其幼苗生长困难,难以存活。在坡度较缓的区域常见以建始槭、枫香等为优势种组成的落叶阔叶,宝华玉兰作为伴生种在群落中有一定分布,但林下其幼苗稀少。推测可能是由于群落的郁闭度较高,林下透光度较差,宝华玉兰种子和幼苗在生长初期需要阴凉的环境,但随着幼苗生长耐阴性逐渐减弱,无法从群落环境中获得足够的光照,不利于后期生长,加上种间竞争激烈,结合上述原因最终导致了宝华玉兰的濒危现状。分类和排序结果能够揭示宝华玉兰种群分布与环境之间的关系,说明SOM分类和排序结果具有一定的合理性,能够解释群落与环境之间的生态规律。
邻域函数是SOM和其他聚类算法的重要区别特征[31]。在拓扑结构限制下的训练和学习能够自动筛选和优化分类方案,并且将分类结果可视化,呈现样方之间的相似性,相比较于传统的聚类算法具有一定优势[32]。SOM可以同时分类和排序,确定主要环境因子并找到样方对应的生态梯度,而且可以保证客观性和准确性,主成分分析法(PCA)等其他分析方法目前无法做到这一点。基于SOM对宝华玉兰生存群落的分类排序研究,SOM在植物群落的类型划分具有合理性,能准确反映群落特征等优点,并有助于探索环境因子影响下的群落分布格局,能够为珍稀濒危植物物种保护提供新的思路和方法。
Classification and ordination of a Magnolia zenii community based on a self-organizing feature map
-
摘要: 为了解国家Ⅱ级保护植物宝华玉兰Magnolia zenii的种群分布规律及其濒危原因,进一步揭示宝华玉兰野生种群与群落及环境之间的生态关系,应用自组织特征映射网络(SOM)对宝华玉兰生存群落进行了排序和分类,以期为保护和管理宝华玉兰种群野生种质资源提供理论依据和新的方法。结果如下:①群落结构分析表明,宝华玉兰天然更新缺乏,且紫楠Phoebe sheareri,建始槭Acer henryi等优势种占据了主要生存空间与资源,影响了宝华玉兰种群增长和发展,使其在群落中处于不稳定地位;②SOM将40个样方进行聚类分析,得到7个群落类型。不同群落类型之间的物种组成存在差异,界限清晰,排序结果与优势种实际分布情况吻合,说明SOM的分类结果具有合理性;③环境因子可视化分析揭示了宝华玉兰分布于坡度较缓的半阳坡,种群分布主要受到坡向和坡度2个环境因子的影响,由于宝华玉兰自身生长过程中对环境要求的特殊性和不适应性,造成了其濒危现状;④SOM具有高度非线性拟合的特点,且能够同时进行排序和分类,并将分类结果可视化,呈现样方之间的相似性,说明SOM应用于群落分类和排序,反映种群与群落及环境之间的生态关系是合适的。Abstract: To understand the population distribution of the National Grade Ⅱ protected plant, Magnolia zenii, and reasons for its endangered status; to reveal the ecological relationship between the M. zenii community and the environment; and to provide a theoretical basis and new methods for protecting and managing the wild germplasm resources of M. zenii, a self-organizing feature map (SOM) network was used to sort and classify the living communities of M. zenii. 40 samples (quadrats) of 10 m×10 m for forest, 5 m×5 m for shrub land and 1 m×1 m for grass land were set up and species data was recorded in each sample. The classification and ordination was carried out by use of the NNTool box in MATLAB(6.1). Results of the community structure analysis showed that (1) natural regeneration of M. zenii was lacking with the dominant species, Phoebe sheareri and Acer henryi, occupying the main living space and resources. (2) The SOM clustered 40 quadrats of plant communities to obtain seven community types. Species composition of the different community types varied, the boundaries were clear, and the ranking results were consistent with actual distribution of the dominant species. (3) Visual analysis of the environmental factors showed that M. zenii was distributed on the semi-sunny and gentle slopes. Population distribution was mainly affected by two environmental factors:slope and slope direction. (4) The SOM had a highly nonlinear fitting, could sort and classify at the same time, and could visualize the classification results to show similarity between quadrats. The study suggests that the dominant species occupy the main environmental resources of the community, which affects the growth of M. zenii population, furthermore, the special requirements of its growth process and being unable to adapt caused its endangered status. Additionally, SOM is an effective quantitative technique in plant ecology, and it is applicable to reflect the ecological relationship of population, community, and the environment.
-
Key words:
- botany /
- classification /
- ordination /
- self-organizing map /
- Magnolia zenii
-
图 2 DBI指数随NOC值变化图
Figure 2 Change trands of Davies-Bouldin index based on number of clusters
图 3 宝华玉兰生存群落40个样方拓扑映射图
Figure 3 Topology map of 40 samples of Magnolia zenii living communities
图 5 优势种在SOM排序图上的梯度分布
Figure 5 Gradient distributions of dominant species on the SOM ordination diagram
表 1 8个坡向等级
Table 1. Eight levels of aspect
序号 坡向 方位角 1 北坡 0°~ 22.5°,337.5°~360.0° 2 东北坡 22.5°~67.5° 3 西北坡 292.5°~337.5° 4 东坡 67.5°~112.5° 5 西坡 247.5°~292.5° 6 东南坡 112.5°~157.5° 7 西南坡 202.5°~247.5° 8 南坡 157.5°~202.5°  下载: 导出CSV
下载: 导出CSV
表 2 宝华山宝华玉兰生存群落重要值
Table 2. Importance values of Magnolia zenii living communities in Baohua Mountain
层次 重要值排名 树种 相对密度/相对高度 相对频度 相对优势度/相对盖度 重要值/% 乔木层 1 紫楠Phoebe sheareri 51.07 6.80 77.56 45.15 2 建始槭Acer henryi 11.06 6.80 10.51 9.46 3 青冈Cyclobalanopsis glauca 8.91 6.12 3.74 6.26 4 红枝柴Meliosma oldhamii 3.55 5.44 2.71 3.90 5 油茶Camellia oleifera 3.55 6.80 0.25 3.53 6 宝华玉兰Magnolia zenii 1.32 6.80 0.97 3.03 7 枫香树Liquidambar formosana 1.40 5.44 2.04 2.96 8 琅玡榆Ulmus chenmoui 1.40 5.44 0.26 2.37 9 枳椇Hovenia acerba 1.32 4.76 0.24 2.11 灌木层 1 紫楠Phoebe sheareri 36.18 8.77 31.36 25.44 2 建始槭Acer henryi 56.18 8.77 61.36 42.11 3 络石Trachelospermum jasminoides 10.41 8.77 9.60 9.59 4 油茶Camellia oleifera 4.43 8.77 6.69 6.63 5 青冈Cyclobalanopsis glauca 5.46 8.77 3.84 6.02 6 老鸦柿Diospyros rhombifolia 4.25 7.89 4.77 5.64 7 卫矛Euonymus alatus 4.29 7.02 4.02 5.11 8 野蔷薇Rosa multiflora 0.98 6.14 0.36 2.49 9 薜荔Ficus pumila 2.01 3.51 0.84 2.12 22 宝华玉兰Magnolia zenii 2.89 2.63 0.67 2.07 草本层 1 阔叶山麦冬Liriope platyphylla 14.17 4.55 27.09 15.27 2 堇菜Viola verecunda 5.96 2.27 33.04 13.76 3 淡竹叶Lophatherum gracile 16.55 4.55 14.21 11.77 4 中国石蒜Lycoris chinensis 15.43 11.36 3.19 10.00 5 天葵Semiaquilegia adoxoides 11.48 2.27 11.29 8.35 6 贯众Cyrtomium fortunei 3.92 11.36 1.76 5.68 7 沿阶草Ophiopogon bodinieri 2.83 11.36 0.50 4.90 8 长柄山蚂蝗Podocarpium podocarpum 3.73 6.82 1.21 3.92 9 知风草Eragrostis ferruginea 5.37 4.55 1.32 3.75 10 钩腺大戟Euphorbia sieboldiana 4.47 4.55 0.83 3.28 11 渐尖毛蕨Cyclosorus acuminatus 1.94 6.82 0.61 3.12 12 点腺过路黄Lysimachia hemsleyana 2.68 2.27 2.09 2.35 13 蘘荷Zingiber mioga 1.79 4.55 0.22 2.19 14 求米草Oplismenus undulatifolius 3.58 2.27 0.50 2.12
下载: 导出CSV
-
[1] 姚帅臣, 王景升, 丁陆彬, 等.拉萨河谷草地群落的数量分类与排序[J].生态学报, 2018, 38(13):4779-4788. YAO Shuaichen, WANG Jingsheng, DING Lubin, et al. Quantitative classtion and ordination of grassland communities in Lhasa River Valley[J]. Acta Ecol Sin, 2018, 38(13):4779-4788. [2] 张峰, 张金屯.我国植被数量分类和排序研究进展[J].山西大学学报(自然科学版), 2000, 23(3):278-282. ZHANG Feng, ZHANG Jintun. Research progress of numerical classification and ordination of vegetation in China[J]. J Shanxi Univ Nat Sci Ed, 2000, 23(3):278-282. [3] 段后浪, 赵安, 姚忠.排序法在植物群落与环境关系研究中的应用述评[J].热带亚热带植物学报, 2017, 25(2):202-208. DUAN Houlang, ZHAO An, YAO Zhong. Overview of ordination methods application in relationship between plant community and environment[J]. J Trop Subtrop Bot, 2017, 25(2):202-208. [4] 庾晓红, 李贤伟, 白降丽.我国植被数量分析方法的研究概况和发展趋势[J].生态学杂志, 2005, 24(4):448-451. YU Xiaohong, LI Xianwei, BAI Jiangli. Research progress and development trend on numerical analysis of vegetation in China[J]. Chin J Ecol, 2005, 24(4):448-451. [5] 武伟.人工神经网络的范畴解释及其应用[D].南京: 南京大学, 2016. WU Wei. The Categorical Explanation and Application of Aetificial Neural Networks[D]. Nanjing: Nanjing University, 2016. [6] 杨黎刚, 苏宏业, 张英, 等.基于SOM聚类的数据挖掘方法及其应用研究[J].计算机工程与科学, 2007, 29(8):133-136. YANG Ligang, SU Hongye, ZHANG Ying, et al. A method of data mining based on SOM clustering and its application[J]. Comput Eng Sci, 2007, 29(8):133-136. [7] 吴红艳.基于自组织特征映射网络的聚类算法研究[D].重庆: 重庆大学, 2006. WU Hongyan. Clustering Algorithm Research based on Self-Organiziing Feature Map Network[D]. Chongqing: Chongqing University, 2006. [8] 刘金福, 朱德煌, 兰思仁, 等.戴云山黄山松群落与环境的关联[J].生态学报, 2013, 33(18):5731-5736. LIU Jinfu, ZHU Dehuang, LAN Siren, et al. Association between environment and community of Pinus taiwanensis in Daiyun Mountain[J]. Acta Ecol Sin, 2013, 33(18):5731-5736. [9] STANKIEWICZ A, KOSIBA P. Advances in ecological modelling of soil properties by self-organizing feature maps of natural environment of Lower Silesia (Poland)[J]. Acta Soc Bot Pol, 2009, 78(2):167-174. [10] 秋哲光.宝华山的宝华玉兰[J].植物杂志, 1989(4):7. QIU Zheguang. The Magnolia zenii in Baohua Mountain[J]. J Plant, 1989(4):7. [11] 王剑伟, 张光富, 陈会艳.特有珍稀植物宝华玉兰种群分布格局和群落特征[J].广西植物, 2008, 28(4):489-494. WANG Jianwei, ZHANG Guangfu, CHEN Huiyan. Population pattern and community characteristics of endemic and rare plant Magnolia zenii in Baohuashan National Forest Park[J]. Guihaia, 2008, 28(4):489-494. [12] 徐惠强, 郝日明, 姚志刚, 等.珍稀树种小叶银缕梅和宝华玉兰自然现状及其就地保护研究[J].江苏林业科技, 2001, 28(5):19-21. XU Huiqiang, HAO Riming, YAO Zhigang, et al. Present natural situation and in situ conservation of rare and endangered trees Magnolia zenii Cheng and Parrotia subaequalis[J]. J Jiangsu For Sci Technol, 2001, 28(5):19-21. [13] 蒋国梅, 孙国, 张光富, 等.濒危植物宝华玉兰种内与种间竞争[J].生态学杂志, 2010, 29(2):201-206. JIANG Guomei, SUN Guo, ZHANG Guangfu, et al. Intra-and interspecific competition of endangered plant Magnolia zenii[J]. Chin J Ecol, 2010, 29(2):201-206. [14] 陈云霞, 南程慧.珍稀濒危植物宝华玉兰遗传多样性的ISSR分析[J].四川农业大学学报, 2016, 34(4):445-449. CHEN Yunxia, NAN Chenghui. ISSR analysis on genetic diversity of endangered plant Magnolia zenii[J]. J Sichuan Agric Univ, 2016, 34(4):445-449. [15] 汪松, 解焱.中国物种红色名录(第1卷:红色名录)[M].北京:高等教育出版社, 2004:309-468. [16] 于永福.中国野生植物保护工作的里程碑:《国家重点保护野生植物名录(第1批)》出台[J].植物杂志, 1999(5):4. YU Yongfu. Milestones for conservation of wild plants in China-Cata of wild plants under key state protection (first batch)[J]. J Plant, 1999(5):4. [17] 安树青, 林向阳, 洪必恭.宝华山主要植被类型土壤种子库初探[J].植物生态学报, 1996, 20(1):41-50. AN Shuqing, LIN Xiangyang, HONG Bigong. A preliminary study on the soil seed banks of the dominant vegetation forms on Baohua Mountaion[J]. Acta Phyoecol Sin, 1996, 20(1):41-50. [18] 汤诗杰, 彭志, 汤庚国.宝华山南京椴群落的特征分析[J].扬州大学学报(农业与生命科学版), 2008, 29(1):90-94. TANG Shijie, PENG Zhi, TANG Gengguo. A community characteristics analysis on Tilia miqueliana in Baohua Mt[J]. J Yangzhou Univ Agric Life Sci Ed, 2008, 29(1):90-94. [19] 张帆航.句容宝华山国家森林公园维管植物区系研究[D].南京: 南京农业大学, 2010. ZHANG Fanhang. A Floristic Study of Vascular Plant on MT. Baohua National Forest Park in Jurong[D]. Nanjing: Nanjing Agricultural University, 2010. [20] 李林峰, 张金屯, 周兰, 等.自组织特征映射网络在芦芽山自然保护区青杄林分类和排序中的应用[J].林业科学, 2014, 50(5):1-7. LI Linfeng, ZHANG Jintun, ZHOU Lan, et al. Application of self-organizing feature map to classification and ordination of Picea wilsonii forest in Luya Mountain Nature Reserve[J]. Sci Silv Sin, 2014, 50(5):1-7. [21] 苏日古嘎, 张金屯, 张斌, 等.松山自然保护区森林群落的数量分类和排序[J].生态学报, 2010, 30(10):2621-2629. Suriguga, ZHANG Jintun, ZHANG Bin, et al. Numerical classification and ordination of forest communities in the Songshan National Nature Reserve[J]. Acta Ecol Sin, 2010, 30(10):2621-2629. [22] 从睿, 张开文, 谢梦梦, 等.江苏宝华山红柴枝群落组成结构及多样性分析[J].四川农业大学学报, 2017, 35(2):213-219. CONG Rui, ZHANG Kaiwen, XIE Mengmeng, et al. Community composition structure and diversity of Meliosma oldhamii maxim in the Baohua Mountain of Jiangsu Province[J]. J Sichuan Agric Univ, 2017, 35(2):213-219. [23] 李春华, 李宁, 史培军.自组织特征映射神经网络原理和应用研究[J].北京师范大学学报(自然科学版), 2006, 42(5):543-547. LI Chunhua, LI Ning, SHI Peijun. The principle and application of self-organizing mapping network in the cluster of cultivated land use pressure in China[J]. J Beijing Norm Univ Nat Sci, 2006, 42(5):543-547. [24] ZHANG Jinyun, DONG Yiru, XI Yuexiang. A comparison of SOFM ordination with DCA and PCA in gradient analysis of plant communities in the midst of Taihang Mountains, China[J]. Ecol Inform, 2008, 3(6):367-374. [25] 张金屯, 杨洪晓.自组织特征人工神经网络在庞泉沟自然保护区植物群落分类中的应用[J].生态学报, 2007, 27(3):1005-1010. ZHANG Jintun, YANG Hongxiao. Application of self-organizing neural networks to classification of plant communities in Pangquangou Nature Reserve, North China[J]. Acta Ecol Sin, 2007, 27(3):1005-1010. [26] 苏日古嘎, 张金屯, 田世广, 等.自组织特征映射网络在北京松山自然保护区山地草甸数量分析中的应用[J].植物生态学报, 2010, 34(7):811-818. SURI Guga, ZHANG Jintun, TIAN Shiguang, et al. Application of self-organizing map to quantitative analysis of mountain meadow in the Songshan Nature Reserve of Beijing, China[J]. Chin J Plant Ecol, 2010, 34(7):811-818. [27] 李林峰.芦芽山自然保护区青杄林的数量生态学研究[D].北京: 北京师范大学, 2014: 43-45. LI linfeng. Quantitative Ecology of Picea wilsohii Forest in the Luya Mountain Nature Reserve[D]. Beijing: Beijing Normal University, 2014: 43-45. [28] 张钦弟, 张金屯, 苏日古嘎, 等.庞泉沟自然保护区华北落叶松林的自组织特征映射网络分类与排序[J].生态学报, 2011, 31(11):2990-2998. ZHANG Qindi, ZHANG Jintun, Suriguga, et al. Self-organizing feature map classification and ordination of Larix principis-rupprechtii forest in Pangquangou Nature Reserve[J]. Acta Ecol Sin, 2011, 31(11):2990-2998. [29] 张金屯, 孟东平, 席跃翔.自组织神经影射网络排序及其在植物群落分析中的应用[J].北京林业大学学报, 2008, 30(1):1-5. ZHANG Jintun, MENG Dongping, XI Yuexiang. Ordination of self-organizing feature map neural networks and its application in the study of plant communities[J]. J Beijing For Univ, 2008, 30(1):1-5. [30] GILBERT B, LECHOWICZ M J. Neutrality, niches, and dispersal in a temperate forest understory[J]. Proc Nat Acad Sci USA, 2004, 101(20):7651-7656. [31] 徐步云, 倪禾.自组织神经网络和K-means聚类算法的比较分析[J].新型工业化, 2014, 4(7):63-69. XU Buyun, NI He. Comparative analysis of SOM and K-means clustering[J]. J New Ind, 2014, 4(7):63-69. [32] 张春平, 何平, 冀花存, 等.渝东地区药用保护植物金荞麦群落数量分类和排序研究[J].西南大学学报(自然科学版), 29(8):107-113. ZHANG Chunping, HE Ping, JI Huacun, et al. A study on the numerical classification and ordination of medicinal conservation plant Fagopyrum cymosum communities in eastern Chongqing[J]. J Southwest Univ Nat Sci Ed, 2007, 29(8):107-113. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.02.006
 点击查看大图
点击查看大图
计量
- 文章访问数: 3903
- HTML全文浏览量: 1032
- PDF下载量: 126
- 被引次数: 0
