








-
毛竹Phyllostachys edulis林是中国南方重要的森林类型,具有生长快、周期短、产量高、用途广等特点,对稳定调节碳平衡和护坡也有积极作用[1-2]。空间结构是森林生长过程的驱动因子,对森林未来的发展具有决定性作用[3]。毛竹林的空间结构指毛竹个体之间的相互关系,包括毛竹空间分布格局、竞争指数和年龄隔离度等[4]。近年来,对毛竹林空间结构的研究日益增多[5-8]。研究表明:与距离有关的空间结构指数可以精确描述毛竹林的结构特征,对分析和调控毛竹林结构与功能关系至关重要[9]。目前,对毛竹林空间结构的研究主要集中于某地如浙江省龙游县溪口镇、浙江省天目山自然保护区等地的典型毛竹林的空间结构特征[5-6]、动态变化分析[8-10]及非空间结构因子的关系[10-12]等,对大尺度上不同地区的毛竹林空间结构差异性研究尚少有报道。浙江省是中国毛竹主产区。根据第8次全国森林资源连续清查结果(2009-2013),全国毛竹林面积443.01万hm2,浙江省毛竹林面积71.6万hm2,占全国的16.16%,全国排名第3位。开展浙江省毛竹林空间结构特征研究,对精准掌握毛竹林的结构信息,提高全省毛竹林经营决策水平具有重要意义。
-
浙江省位于27°01′~31°10′N,118°01′~123°08′E,面积10.55万km2。年均气温为16.0~19.0 ℃,四季分明,光照充足,雨水充沛,是亚热带湿润季风气候。浙江有“七山一水二分田”之说,山地和丘陵占74.63%,比较适合毛竹林的生长。毛竹广泛分布于海拔为400~800 m的丘陵、低山山麓地带。
-
为增强调查样地的代表性,根据浙江省森林资源一类调查的系统抽样样地中毛竹林样地分布较多的区域,确定调查地点为浙江省南部的丽水市庆元县和温州市泰顺县、西部的衢州市常山县,中部的金华市武义县和绍兴市诸暨市,东部的宁波市宁海县、余姚市和台州市黄岩区,北部的湖州市安吉县、杭州市临安区共10个县市(区),选择少受人为干扰的近自然毛竹林典型地段,在不同坡向(东、南、西、北)和不同坡位(上坡、中坡、下坡)设置样地,样地面积为10 m × 10 m,共设置54个样地。其中,在庆元县、泰顺县、黄岩区、安吉县各设置了4个样地,在常山县、武义县、临安区、诸暨市、宁海县各设置了6个样地,余姚市设置了8个样地。各样地用全球定位系统(GPS)进行定位,用罗盘仪测量样地边界,调查每株毛竹的x和y坐标(m)、年龄、胸径、竹高、冠幅、生长状态等因子;同时,记录样地内的诸如海拔、坡向、坡度、坡位、腐殖质厚度、土层厚度等立地因子。
-
聚集指数是检验种群空间分布格局的常用指数[3]。计算公式为:
$$ R = \frac{{\frac{1}{N}\sum\limits_{i = 1}^N {{r_i}} }}{{\frac{1}{2}\sqrt {\frac{F}{N}} }}。 $$ (1) 式(1)中:ri表示第i株毛竹到其最近邻毛竹的距离;F表示样地面积;N表示样地内毛竹总株数;R是林分聚集指数。若R>1,那么这块样地的毛竹林属于均匀分布;R=1,则属于随机分布;R<1,则属于聚集分布。
-
采用基于Voronoi图的Hegyi竞争指数[14]。计算公式为:
$$ {I_{{\rm{C}}i}} = \sum\limits_{j = 1}^{{n_i}} {\frac{{{d_i}}}{{{d_i}{L_{ij}}}}}。 $$ (2) 式(2)中:di是第i株对象竹的胸径;dj是第j株相邻竹的胸径;Lij是第i株对象竹到第j株相邻竹的距离;ni是基于Voronoi图的所有相邻竹株数;ICi是第i株对象竹的竞争指数。取样地内全部竹子竞争指数的平均值作为林分竞争指数。计算公式为:
$$ {I_{\rm{C}}} = \frac{1}{N}\sum\limits_{i = 1}^N {{I_{{\rm{C}}i}}}。 $$ (3) 式(3)中:IC为林分竞争指数;N为样地内毛竹总株数;ICi为第i空间结构单元中对象竹的竞争指数。
-
本研究采用全混交度描述毛竹林的年龄隔离度[15]。全混交度的计算公式为:
$$ {M_{{{\rm{C}}_i}}}{\rm{ = }}\frac{1}{2}\left( {{D_i} + \frac{{{c_i}}}{{{n_i}}}} \right){M_i}。 $$ (4) 式(4)中:MCi表示第i空间结构单元中对象竹的全混交度;Di表示空间结构单位的Simpson指数,${D_i} = 1 - \sum\limits_{j = 1}^{{S_i}} {{p^2}_j} $,Di的取值范围为[0, 1];ci表示对象竹的最近邻竹中成对相邻竹不是同一竹龄的个数;ni表示最近邻竹的株数;$\frac{{{c_i}}}{{{n_i}}}$表示最近邻竹的年龄隔离度;Mi表示简单混交度,${M_i} = \frac{1}{{{n_i}}}\sum\limits_{j = 1}^{{n_i}} {{V_{i, j}}} $,如果第i对象竹和第j最近邻竹年龄相同,vi, j=0,否则等于1。取样地内全部竹子年龄隔离度平均值作为林分年龄隔离度,计算公式为:
$$ M = \frac{1}{N}\sum\limits_{i = 1}^N {{M_{{{\rm{C}}_i}}}} 。 $$ (5) 式(5)中:M表示林分年龄隔离度;N表示样地内毛竹总株数;MCi表示第i空间结构单元中对象竹的年龄隔离度。
-
由于毛竹内部中空,本研究采用可测量不规则形状物体体积的排水法测定毛竹竹秆材积。首先,将水注入定制水桶内至水龙头齐平处,排出桶内多余的水。然后,将竹秆劈成40 cm左右长的竹条,缓慢放入桶内,用铁桶收集由水龙头排出的水,用电子提秤称取总竹条排水量。最后计算毛竹竹秆材积。公式如下:
$$ {V_秆} = \frac{{{M_水}}}{\rho }。 $$ (6) 式(6)中:V秆为竹秆材积(dm3),M水为排水质量(kg);ρ为水的比重(ρ=1 g·cm-3)。
-
对10个县市(区)毛竹林空间结构因子进行单因素方差分析,并对差异显著(P<0.05)的空间结构因子进行多重比较。方差分析采用SPSS 20.0完成。空间结构因子的区域变化分析采用ArcGIS 10.2完成。
-
浙江省毛竹林空间结构指数的统计指标如表 1所示。以不同地区空间结构指数统计指标的平均值作为全省毛竹林空间结构指数。可以看出,3个空间结构指数中,竞争指数的均值为5.62,最大值为8.81,最小值为2.88;年龄隔离度的均值为0.58,最大值为0.84,最小值为0.30;聚集指数的均值为0.94,最大值为1.24,最小值为0.73。竞争指数的变异系数最大,达0.25,聚集指数的变异系数最小,为0.13。
表 1 浙江省毛竹林空间结构指数描述统计特征
Table 1. Descriptive statistical characteristics of spatial structure index of moso bamboo forest in Zhejiang Province
地区 空间结构指数 样地数 均值 中值 极小值 极大值 标准差 峰度 偏度 变异系数 竞争指数 8 6.47 6.71 4.75 8.68 1.20 0.74 0.48 0.19 余姚 年龄隔离度 8 0.62 0.62 0.52 0.73 0.07 -0.97 0.18 0.12 聚集指数 8 0.84 0.84 0.73 1.00 0.09 -0.47 0.46 0.11 竞争指数 6 6.40 6.56 5.52 7.35 0.65 -0.11 0.00 0.10 临安 年龄隔离度 6 0.51 0.51 0.34 0.65 0.11 0.57 -0.38 0.21 聚集指数 6 0.97 0.96 0.92 1.07 0.06 2.52 1.49 0.06 竞争指数 6 6.36 6.25 5.57 7.57 0.68 2.15 1.16 0.11 诸暨 年龄隔离度 6 0.60 0.64 0.36 0.71 0.12 4.58 -2.01 0.21 聚集指数 6 0.97 0.97 0.89 1.03 0.05 -0.59 -0.25 0.05 竞争指数 4 5.96 5.81 5.18 7.03 0.90 -3.53 0.43 0.15 安吉 年龄隔离度 4 0.47 0.47 0.44 0.50 0.03 -5.98 0.00 0.07 聚集指数 4 0.88 0.86 0.83 0.97 0.07 2.24 1.49 0.07 竞争指数 6 5.89 5.43 3.82 8.81 2.04 -1.63 0.54 0.35 宁海 年龄隔离度 6 0.56 0.53 0.47 0.68 0.09 -1.62 0.58 0.16 聚集指数 6 0.97 0.97 0.76 1.24 0.18 -0.38 0.38 0.18 竞争指数 4 4.97 4.73 4.54 5.88 0.61 3.64 1.88 0.12 黄岩 年龄隔离度 4 0.71 0.72 0.55 0.84 0.14 -3.25 -0.37 0.19 聚集指数 4 0.98 0.99 0.94 1.00 0.03 3.47 -1.82 0.03 竞争指数 6 4.97 5.12 3.37 6.72 1.16 0.07 0.13 0.23 武义 年龄隔离度 6 0.55 0.59 0.30 0.66 0.13 3.97 -1.90 0.24 聚集指数 6 0.92 0.89 0.76 1.20 0.15 3.38 1.57 0.16 竞争指数 6 4.89 4.72 2.95 7.44 1.64 -0.52 0.54 0.34 常山 年龄隔离度 6 0.60 0.61 0.45 0.71 0.10 -1.11 -0.43 0.16 聚集指数 6 0.96 0.93 0.80 1.17 0.13 -0.25 0.65 0.14 竞争指数 4 5.23 5.51 3.50 6.40 1.23 2.39 -1.26 0.24 泰顺 年龄隔离度 4 0.64 0.66 0.55 0.70 0.07 1.12 -1.21 0.11 聚集指数 4 0.94 0.92 0.90 1.02 0.05 3.37 1.80 0.05 竞争指数 4 3.99 3.59 2.88 5.91 1.32 2.94 1.60 0.33 庆元 年龄隔离度 4 0.53 0.54 0.38 0.64 0.11 0.50 -0.72 0.21 聚集指数 4 1.03 1.02 0.83 1.24 0.19 -4.30 0.11 0.19 竞争指数 54 5.62 5.57 2.88 8.81 1.38 -0.30 0.00 0.25 平均 年龄隔离度 54 0.58 0.58 0.30 0.84 0.11 0.20 -0.29 0.19 聚集指数 54 0.94 0.93 0.73 1.24 0.12 0.59 0.64 0.13 -
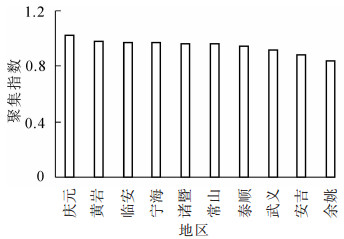
根据表 1可知:庆元的毛竹林聚集指数最大,为1.03,属于均匀分布。其他地区的聚集指数均小于1,属于聚集分布。全省毛竹林聚集指数平均值为0.94,属于聚集分布。从图 1可见:全省毛竹林聚集指数分布趋势不明显。聚集指数方差分析结果表明:不同地区毛竹林聚集指数没有显著性差异(P>0.05)(表 2)。
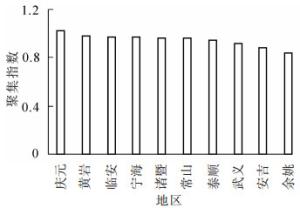
图 1 不同地区毛竹林聚集指数
Figure 1. Aggregation index of moso bamboo forests in different regions
表 2 不同地区毛竹林空间结构指数方差分析
Table 2. Analysis of variance of spatial structure index of moso bamboo forest in different regions
方差来源 离差平均和 自由度 均方 F 显著性水平 地区间 0.155 9 0.017 1.31 0.259 聚集指数 地区内 0.578 44 0.013 总数 0.733 53 地区间 0.191 9 0:021 2:069 0.054 年龄隔离度 地区内 0.450 44 0:01 总数 0.641 53 地区间 32:376 9 3.597 2:296 0:033 竞争指数 地区内 68:943 44 1.567 总数 101:319 53 -
根据表 1和图 2可知:年龄隔离度最大的地区是位于浙江省东部区域的黄岩地区;年龄隔离度最小的地区是位于浙江省北部区域的安吉地区,全省毛竹林年龄隔离度分布趋势不明显。年龄隔离度方差分析的结果表明:不同地区毛竹林年龄隔离度没有显著性差异(P>0.05)(表 2)。
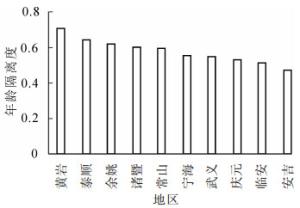
图 2 不同地区毛竹林年龄隔离度
Figure 2. Age mingling of moso bamboo forests in different regions
-
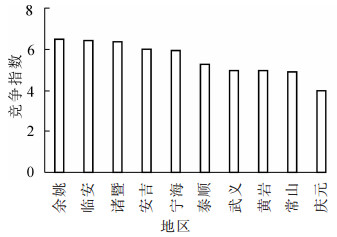
由表 1和图 3可知:竞争指数最小的是位于浙江省南部区域的庆元地区,竞争指数最大的是位于浙江省北部区域的余姚地区。从地理位置上看,浙江省南北部区域的竞争指数存在一定的差异,北部区域的竞争指数大于南部区域。
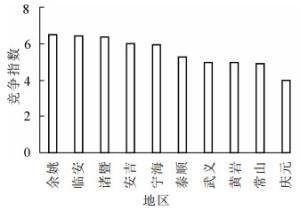
图 3 不同地区毛竹林竞争指数
Figure 3. Competition index of moso bamboo forests in different regions
竞争指数方差分析结果如表 2所示。不同地区毛竹林竞争指数存在显著差异(P<0.05)。因此,进一步对10个县(市、区)毛竹林竞争指数进行多重比较。多重比较的结果如图 4所示:余姚与庆元、常山、武义毛竹林竞争指数存在显著差异(P<0.05),与黄岩、泰顺、宁海、安吉、诸暨、临安毛竹林竞争指数差异不显著。临安、诸暨与庆元、常山毛竹林竞争指数存在显著差异(P<0.05),与黄岩、武义、泰顺、宁海、安吉毛竹林竞争指数差异不显著。诸暨与庆元、常山毛竹林竞争指数存在显著差异(P<0.05),与黄岩、武义、泰顺、宁海、安吉毛竹林竞争指数差异不显著。安吉、宁海与庆元毛竹林竞争指数存在显著差异(P<0.05),与常山、黄岩、武义、泰顺、宁海毛竹林竞争指数差异不显著。泰顺与庆元、常山、黄岩、武义毛竹林竞争指数差异不显著。武义与庆元、常山、黄岩毛竹林竞争指数差异不显著。黄岩与庆元、常山毛竹林竞争指数差异不显著。常山与庆元毛竹林竞争指数差异不显著。

图 4 不同地区毛竹林竞争指数多重比较
Figure 4. Multiple comparison of competition index of moso bamboo forests in different regions
-
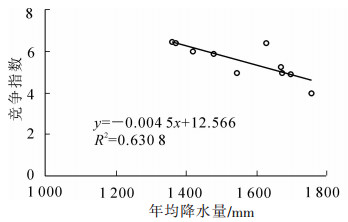
SHI等[15]研究发现:年均降水量和年均气温对毛竹林结构具有一定的影响。由图 5和图 6可知:不同地区竞争指数与年均降水量和年均气温显著相关(P<0.01),随着年均降水量和年均气温的增加,竞争指数减小。周文伟[16]研究表明:降水量的增加能促进毛竹的生长,水热条件较高的庆元比水热条件较低的安吉的毛竹产量高。实际上,较好的水热条件可以降低毛竹之间的竞争,从而促进毛竹的生长。可见,年均降水量和年均气温是影响空间结构的重要因素。
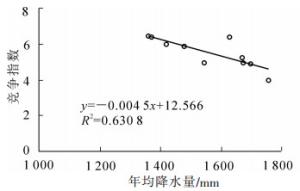
图 5 竞争指数与年均降水量的关系
Figure 5. Relationship between competition index and mean annual precipitation of moso bamboo forest in Zhejiang Province
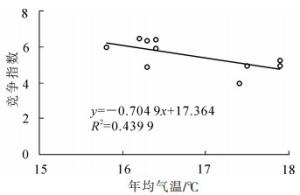
图 6 竞争指数与年均气温的关系
Figure 6. Relationship between competition index and mean annual temperature of moso bamboo forest in Zhejiang Province
-
为提高毛竹林经营水平,研究者通常关注立竹度、年龄结构和树种组成等结构因子[7]。本研究表明:浙江省不同地区的毛竹林竞争指数存在较大差异。不同地区毛竹竹秆材积与竞争指数存在相反的变化趋势(图 4和图 7)。因此,调控毛竹林的竞争关系在毛竹林经营中具有不可忽视的作用。为提高竹秆材积产量,在北部地区更需要采取合理的空间结构调控措施,降低毛竹林竞争强度,以促进毛竹生长,提高毛竹产量。
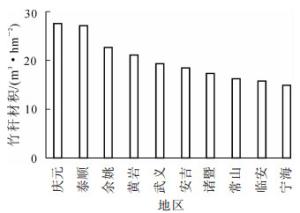
图 7 不同地区毛竹林竹秆材积
Figure 7. Stem volume of moso bamboo forest in different regions
-
本研究以浙江省少受人为干扰的近自然毛竹林为研究对象,研究浙江省不同地区毛竹林空间结构特征及其差异性。得出以下主要结论:①浙江省近自然毛竹林空间结构因子的统计特征表明,10个地区毛竹林竞争指数为2.88~8.81,平均为5.62,余姚毛竹林竞争指数最大,临安毛竹林竞争指数次之,庆元毛竹林竞争指数最小;毛竹林年龄隔离度为0.30~0.84,平均为0.58,黄岩毛竹林年龄隔离度最高,余姚次之,武义最低;毛竹林聚集指数为0.73~1.24,平均为0.94,宁海毛竹林聚集指数最大,庆元次之,余姚最小。浙江省近自然毛竹林空间分布格局以聚集分布为主。②浙江省近自然毛竹林空间结构因子的差异性分析结果表明:浙江省不同地区毛竹林竞争指数存在显著差异,聚集指数和年龄隔离度不存在显著差异。空间结构指数存在一定的区域变化趋势,从北到南竞争指数存在逐渐减小的趋势。
Spatial structure characteristics of close-to-nature Phyllostachys edulis forests in Zhejiang Province
-
摘要:
目的 研究浙江省不同地区毛竹Phyllostachys edulis林空间结构特征及其差异性。 方法 以浙江省不同地区近自然生长的毛竹林为研究对象,共设置54个样地,采用聚集指数、竞争指数和年龄隔离度3个空间结构指数。 结果 浙江省不同地区毛竹林竞争指数为2.88~8.81,其中余姚地区最大,庆元地区最小;年龄隔离度为0.30~0.84,其中黄岩地区最大,武义地区最小;聚集指数为0.73~1.24,其中宁海地区最大,余姚地区最小。浙江省不同地区毛竹林年龄隔离度和聚集指数没有显著差异,而不同地区间竞争指数存在显著差异(P < 0.05)。 结论 浙江省毛竹林空间分布格局以聚集分布为主。空间结构指数存在一定的区域变化趋势,从北到南竞争指数逐渐减小。 Abstract:Objective Spatial structure characteristics and differences of moso bamboo (Phyllostachys edulis) forests in different regions of Zhejiang Province were studied. Method The close-to-nature moso bamboo forests in different regions of Zhejiang Province were taken as the research objects, 54 sample plots were set up, and spatial structure indices (aggregation index, competition index and age mingling) were used to analyze spatial structure characteristics and differences of moso bamboo forests in different regions of Zhejiang Province. Result The mean competition index of moso bamboo forest in different regions of Zhejiang Province is 2.88-8.81, among which Yuyao is the largest and Qingyuan is the smallest. The mean age mingling is 0.30-0.84, the largest in Huangyan and the smallest in Wuyi. The mean aggregation index is 0.73-1.24, the largest in Ninghai area and the smallest in Yuyao area. There was no significant difference in age mingling and aggregation index among different regions in Zhejiang Province, whereas there was significant difference in competition index among different regions in Zhejiang Province(P < 0.05). Conclusion The spatial distribution pattern of moso bamboo forests in Zhejiang Province was mainly aggregated distribution. The spatial structure index has a certain regional variation trend, and the competition index decreases gradually from north to south. -
Key words:
- forest ecology /
- Zhejiang Province /
- Phyllostachys edulis /
- aggregation index /
- competition index /
- age mingling
-
图 1 不同地区毛竹林聚集指数
Figure 1 Aggregation index of moso bamboo forests in different regions
图 3 不同地区毛竹林竞争指数
Figure 3 Competition index of moso bamboo forests in different regions
图 4 不同地区毛竹林竞争指数多重比较
Figure 4 Multiple comparison of competition index of moso bamboo forests in different regions
图 5 竞争指数与年均降水量的关系
Figure 5 Relationship between competition index and mean annual precipitation of moso bamboo forest in Zhejiang Province
图 6 竞争指数与年均气温的关系
Figure 6 Relationship between competition index and mean annual temperature of moso bamboo forest in Zhejiang Province
表 1 浙江省毛竹林空间结构指数描述统计特征
Table 1. Descriptive statistical characteristics of spatial structure index of moso bamboo forest in Zhejiang Province
地区 空间结构指数 样地数 均值 中值 极小值 极大值 标准差 峰度 偏度 变异系数 竞争指数 8 6.47 6.71 4.75 8.68 1.20 0.74 0.48 0.19 余姚 年龄隔离度 8 0.62 0.62 0.52 0.73 0.07 -0.97 0.18 0.12 聚集指数 8 0.84 0.84 0.73 1.00 0.09 -0.47 0.46 0.11 竞争指数 6 6.40 6.56 5.52 7.35 0.65 -0.11 0.00 0.10 临安 年龄隔离度 6 0.51 0.51 0.34 0.65 0.11 0.57 -0.38 0.21 聚集指数 6 0.97 0.96 0.92 1.07 0.06 2.52 1.49 0.06 竞争指数 6 6.36 6.25 5.57 7.57 0.68 2.15 1.16 0.11 诸暨 年龄隔离度 6 0.60 0.64 0.36 0.71 0.12 4.58 -2.01 0.21 聚集指数 6 0.97 0.97 0.89 1.03 0.05 -0.59 -0.25 0.05 竞争指数 4 5.96 5.81 5.18 7.03 0.90 -3.53 0.43 0.15 安吉 年龄隔离度 4 0.47 0.47 0.44 0.50 0.03 -5.98 0.00 0.07 聚集指数 4 0.88 0.86 0.83 0.97 0.07 2.24 1.49 0.07 竞争指数 6 5.89 5.43 3.82 8.81 2.04 -1.63 0.54 0.35 宁海 年龄隔离度 6 0.56 0.53 0.47 0.68 0.09 -1.62 0.58 0.16 聚集指数 6 0.97 0.97 0.76 1.24 0.18 -0.38 0.38 0.18 竞争指数 4 4.97 4.73 4.54 5.88 0.61 3.64 1.88 0.12 黄岩 年龄隔离度 4 0.71 0.72 0.55 0.84 0.14 -3.25 -0.37 0.19 聚集指数 4 0.98 0.99 0.94 1.00 0.03 3.47 -1.82 0.03 竞争指数 6 4.97 5.12 3.37 6.72 1.16 0.07 0.13 0.23 武义 年龄隔离度 6 0.55 0.59 0.30 0.66 0.13 3.97 -1.90 0.24 聚集指数 6 0.92 0.89 0.76 1.20 0.15 3.38 1.57 0.16 竞争指数 6 4.89 4.72 2.95 7.44 1.64 -0.52 0.54 0.34 常山 年龄隔离度 6 0.60 0.61 0.45 0.71 0.10 -1.11 -0.43 0.16 聚集指数 6 0.96 0.93 0.80 1.17 0.13 -0.25 0.65 0.14 竞争指数 4 5.23 5.51 3.50 6.40 1.23 2.39 -1.26 0.24 泰顺 年龄隔离度 4 0.64 0.66 0.55 0.70 0.07 1.12 -1.21 0.11 聚集指数 4 0.94 0.92 0.90 1.02 0.05 3.37 1.80 0.05 竞争指数 4 3.99 3.59 2.88 5.91 1.32 2.94 1.60 0.33 庆元 年龄隔离度 4 0.53 0.54 0.38 0.64 0.11 0.50 -0.72 0.21 聚集指数 4 1.03 1.02 0.83 1.24 0.19 -4.30 0.11 0.19 竞争指数 54 5.62 5.57 2.88 8.81 1.38 -0.30 0.00 0.25 平均 年龄隔离度 54 0.58 0.58 0.30 0.84 0.11 0.20 -0.29 0.19 聚集指数 54 0.94 0.93 0.73 1.24 0.12 0.59 0.64 0.13  下载: 导出CSV
下载: 导出CSV
表 2 不同地区毛竹林空间结构指数方差分析
Table 2. Analysis of variance of spatial structure index of moso bamboo forest in different regions
方差来源 离差平均和 自由度 均方 F 显著性水平 地区间 0.155 9 0.017 1.31 0.259 聚集指数 地区内 0.578 44 0.013 总数 0.733 53 地区间 0.191 9 0:021 2:069 0.054 年龄隔离度 地区内 0.450 44 0:01 总数 0.641 53 地区间 32:376 9 3.597 2:296 0:033 竞争指数 地区内 68:943 44 1.567 总数 101:319 53
下载: 导出CSV
-
[1] 周国模, 姜培坤.不同植被恢复对侵蚀型红壤活性碳库的影响[J].水土保持学报, 2004, 18(6):68-70. ZHOU Guomo, JIANG Peikun. Changes in active organic carbon of erosion red soil by vegetation recovery[J]. J Soil Water Conserv, 2004, 18(6):68-70. [2] 杨麒麟, 李柏.滑坡区毛竹根系生长分布及其护坡效果研究[J].长江科学院院报, 2017, 34(10):45-49. YANG Qilin, LI Bai. Growth distribution of bamboo root system in landslide area and its slope protection effect[J]. J Yangtze River Sci Res Inst, 2017, 34(10):45-49. [3] PRETZSCH H. Analysis and modeling of spatial stand structures: methodological considerations based on mixed beech-larch stands in Lower Saxony[J]. For Ecol Manage, 1997, 97(3):237-253. [4] 汤孟平, 陈永刚, 徐文兵, 等.森林空间结构分析[M].北京:科学出版社, 2013. [5] 黄丽霞, 袁位高, 黄建花, 等.不同经营方式下毛竹林的林分空间结构比较研究[J].浙江林业科技, 2008, 28(3):48-51. HUANG Lixia, YUAN Weigao, HUANG Jianhua, et al. Comparative study on spatial structure of Phyllostachys heterocycla var. pubescens stand with different management[J]. J Zhejiang For Sci Technol, 2008, 28(3):48-51. [6] 邓英英, 汤孟平, 徐文兵, 等.天目山近自然毛竹纯林的竹秆空间结构特征[J].浙江农林大学学报, 2011, 28(2):173-179. DENG Yingying, TANG Mengping, XU Wenbing, et al. Spatial structure of bamboo culm of an almost natural, pure Phyllostachys pubescens forest in Mount Tianmu[J]. J Zhejiang A&F Univ, 2011, 28(2):173-179. [7] 汤孟平, 徐文兵, 陈永刚, 等.毛竹林空间结构优化调控模型[J].林业科学, 2013, 49(1):120-125. TANG Mengping, XU Wenbing, CHEN Yonggang, et al. Spatial structure optimizing adjustment and control model of Phyllostachys edulis stand[J]. Sci Silv Sin, 2013, 49(1):120-125. [8] 唐思嘉, 汤孟平, 赵赛赛, 等.天目山毛竹竞争空间格局的动态分析[J].浙江农林大学学报, 2018, 35(2):199-208. TANG Sijia, TANG Mengping, ZHAO Saisai, et al. Competitive spatial patterns for moso bamboo on Mount Tianmu[J]. J Zhejiang A&F Univ, 2018, 35(2): 199-208. [9] 汤孟平, 唐守正, 雷相东, 等.两种混交度的比较分析[J].林业资源管理, 2004(4):25-27. TANG Mengping, TANG Shouzheng, LEI Xiangdong, et al. Comparison analysis on two minglings[J]. For Resour Manage, 2004(4):25-27. [10] 仇建习, 汤孟平, 沈利芬, 等.近自然毛竹林空间结构动态变化[J].生态学报, 2014, 34(6):1444-1450. QIU Jianxi, TANG Mengping, SHEN Lifen, et al. Dynamic analysis of spatial structure in a close-to-nature Phyllostachys edulis stands[J]. Acta Ecol Sin, 2014, 34(6):1444-1450. [11] 仇建习, 汤孟平, 沈利芬, 等.天目山近自然毛竹林空间结构与胸径的关系[J].生态学报, 2015, 35(12):4081-4088. QIU Jianxi, TANG Mengping, SHEN Lifen, et al. The relationship between spatial structure and DBH of close-to-nature Phyllostachys edulis stands in Tianmu Mountain[J]. Acta Ecol Sin, 2015, 35(12):4081-4088. [12] 汤孟平, 徐文兵, 陈永刚, 等.天目山近自然毛竹林空间结构与生物量的关系[J].林业科学, 2011, 47(8):1-6. TANG Mengping, XU Wenbing, CHEN Yongguang, et al. Relationship between spatial structure and biomass of a close-to-nature Phyllostachys edulis stand in Tianmu Mountain[J]. Sci Silv Sin, 2011, 47(8):1-6. [13] 汤孟平, 陈永刚, 施拥军, 等.基于Voronoi图的群落优势树种种内种间竞争[J].生态学报, 2007, 27(11):4707-4716. TANG Mengping, CHEN Yonggang, SHI Yongjun, et al. Intraspecific and interspecific competition analysis of community dominant plant populations based on Voronoi diagram[J]. Acta Ecol Sin, 2007, 27(11):4707-4716. [14] 汤孟平, 娄明华, 陈永刚, 等.不同混交度指数的比较分析[J].林业科学, 2012, 48(8):46-53. TANG Mengping, LOU Minghua, CHEN Yonggang, et al. Comparative analyses on different mingling indices[J]. Sci Silv Sin, 2012, 48(8): 46-53. [15] SHI Yongjun, LIN Xu, ZHOU Yufeng, et al. Quantifying driving factors of vegetation carbon stocks of moso bamboo forests using machine learning algorithm combined with structural equation model[J]. For Ecol Manage, 2018, 429:406-413. [16] 周文伟.降水对毛竹林生长的影响分析[J].竹子研究汇刊, 1991, 10(2): 33-39. ZHOU Wenwei. Effect of precipitation on growth of Phyllostachys pubescens forest[J]. J Bamboo Res, 1991, 10(2): 33-39. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.02.005
 点击查看大图
点击查看大图
计量
- 文章访问数: 2846
- HTML全文浏览量: 663
- PDF下载量: 77
- 被引次数: 0
