





-
竹材作为天然可再生、绿色环保的生物质纤维材料,具有强度高、生长周期短等优势,被广泛应用于家具、建筑结构、工具用具等不同领域[1-9]。其中,毛竹Phyllostachys edulis由于种植面积广、成材速度快、蓄积量高、价格低廉等优点,是工程材料中利用较为普遍的竹种之一,可用于制造竹胶合板、竹木复合材料、竹集成材、室内装饰材等产品[10-15]。然而,由于竹材的特异性,竹黄、竹青和竹肉性能存在一定差异。在实际生产中一般需对原竹进行去青、去黄处理,造成毛竹材利用率低等问题。研究毛竹材各部分性能特性,对合理利用毛竹材具有一定应用价值。如竹篾-单板复合材料,是一种利用去青原竹,沿径向剖分制备竹篾,串联成竹帘与单板复合,制备而成的新型竹木复合材料。针对竹材构造的特殊性,采用适当改性与处理方法[16],充分合理利用竹青与竹黄,成为目前研究的重点,对于提高竹材利用率具有一定价值[17-18]。袁晶等[19]研究了维管束分布及结构对慈竹Neosinocalamus affins、花竹Bambusa albo-lineata及绿竹Dendrocalamopsis oldhami压缩性能的影响,发现竹材顺纹压缩性能与维管束分布密度呈正相关。维管束的分布密度是影响竹材压缩性能的结构因素之一。陈秋艳等[20]对绿竹不同部位物理化学特性的研究表明:竹材不同部位中纤维素含量从大到小依次为竹肉、竹青、竹黄;且竹肉的纤维素结晶度大于竹青和竹黄。竹青、竹黄表面结构较为致密,两者表面密度均大于竹肉的表面密度。竹材梯度结构对其力学性能也有一定影响,当竹黄受拉,竹青受压时,竹材的弯曲韧性最好;当竹黄受压,竹青受拉时,竹材弯曲模量最佳[21]。夏旭光等[22]对不同竹龄竹材的不同部位进行力学性能测试表明:竹材力学性能随竹龄增加呈增强趋势;竹材各单层顺纹抗压弹性模量呈非线性变化,竹青单层与竹黄单层模量相差较大,两者模量比值最大值达201%,比值最小值为173%。综上所述,竹材特殊结构与化学组成对各部位性能影响显著[23],直接影响了竹木复合材料制备工艺及性能。本研究拟通过纤维显微特征观察、物理力学性能以及干缩性能测试,分析、评价毛竹材不同部位性能差异,以期为毛竹材高效、合理利用提供基础数据。
-
毛竹采伐于安徽省金寨县青山镇(31.43°N,115.90°E),竹龄为3~4年生。选取15株胸径接近、无霉变等的毛竹,截取原竹距地面1.5~3.5 m的竹筒制作试件。
-
纤维形态的测试采用离析法进行。首先,将毛竹进行纵向剖分和横向截断,获得一定长度与宽度的竹片,再将竹黄、竹肉和竹青分开;然后,将各部分劈成小火柴棍状,分别放入不同试管中。试管中加入V(质量分数为30%过氧化氢)∶V(质量分数为95%冰醋酸)=1∶1的离析液[24],60 ℃水浴加热,直至试样边缘有纤维开始离析为止。最后,将试液倒出并将试样用清水洗涤至无气味,再往试管中加入适量清水摇匀纤维,用解剖针挑取纤维到玻璃载玻片上,用光学显微镜进行纤维形态特征参数测量[25]。
同时,将毛竹锯切成小块状,通过微波加热法对其软化后,用滑走式切片机将毛竹试块切片,利用光学显微镜观察纤维鞘区域、薄壁细胞区域。
-
为研究竹材不同部位对其力学性能的贡献,将试件分为4类:A组试件为去除竹青的试件,尺寸为160 mm×10 mm×6 mm(纵向×弦向×径向);B组试件为去除竹黄的试件,尺寸为160 mm×10 mm×8 mm;C组试件为竹青和竹黄均去除的试件,尺寸为160 mm×10 mm×4 mm;D组试件为原竹片,尺寸为160 mm×10 mm×t mm (t为原竹分片后的自然厚度)。4组试件的抗弯强度和弹性模量测量采用3点弯曲方法进行,详细试验步骤依据GB/T 15780−1995《竹材物理力学性质试验方法》[26]。
顺纹抗压强度测试试件也分为4组:A组试件为去除竹青的试件,尺寸为20 mm×20 mm×6 mm(纵向×弦向×径向);B组试件为去除竹黄的试件,尺寸为20 mm×20 mm×8 mm;C组试件为竹青和竹黄均去除的试件,尺寸为20 mm×20 mm×4 mm;D组试件为原竹片,尺寸为20 mm×20 mm×t mm(t为原竹分片后的自然厚度)。顺纹抗压强度测试依据GB/T 15780−1995《竹材物理力学性质实验方法》[27]。
-
将毛竹加工成10 mm×10 mm×t mm(t为原竹分片后的自然厚度,纵向×弦向×径向)大小的试件(共计150个);然后,采用“逐级削减法”将毛竹试块进行剖分、打磨,制得尺寸为10 mm×10 mm×2 mm的毛竹竹黄片、尺寸为10 mm×10 mm×4 mm的竹肉片和尺寸为10 mm×10 mm×4 mm的竹青片(每类试件各50个,划线标号);最后,按照GB/T 15780−1995《竹材物理力学性质试验方法》进行气干干缩率和全干干缩率测试。
-
由表1可知:毛竹竹肉的纤维长度及宽度均最大,分别为1.88 mm和15.15 μm。t检验结果表明:毛竹竹肉与竹黄、竹青的纤维长度和宽度差异极显著(P<0.01)。
表 1 毛竹材各部位纤维平均尺寸
Table 1. Mean size of fiber in different parts of Ph. edulis
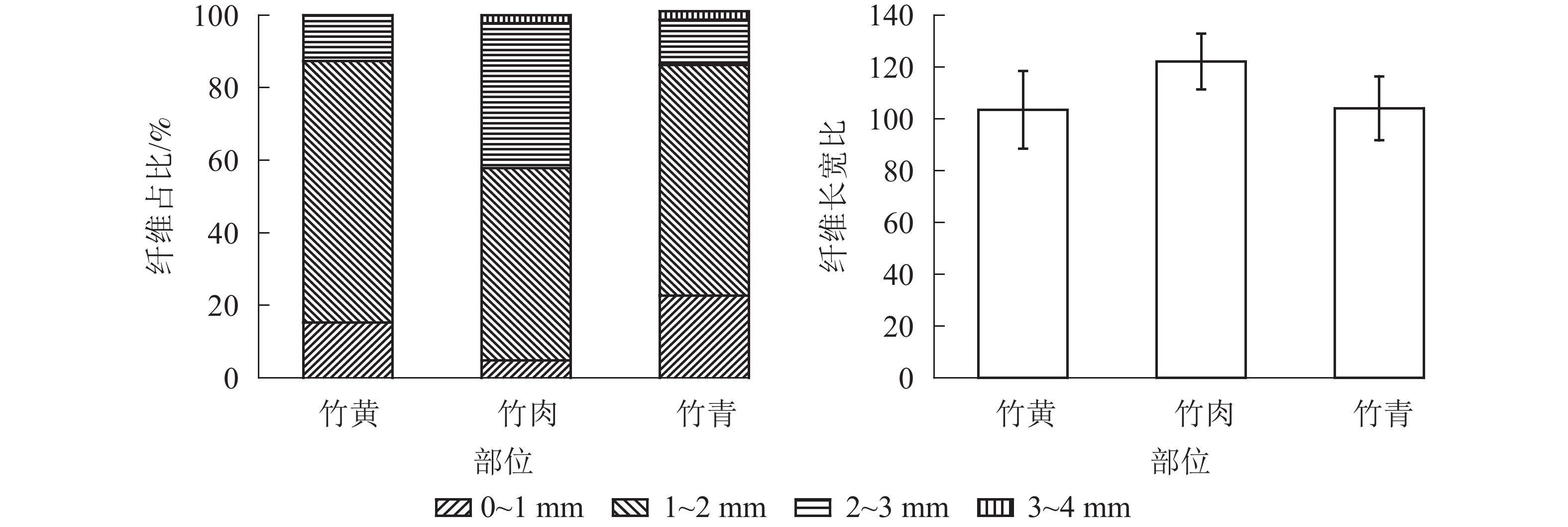
部位 长度/mm 宽度/μm 竹黄 1.48±0.45 b 14.01±2.75 b 竹肉 1.88±0.56 a 15.15±3.53 a 竹青 1.43±0.53 b 13.40±3.47 b 说明:数值为平均值±标准差。不同字母表示部位间差异极 显著(P<0.01) 毛竹不同部位的纤维长度分布频率见图1。竹黄、竹肉及竹青中,长度为1~2 mm的纤维占比最高,分别为72.0%、53.0%和63.5%。竹黄中,0~1、2~3 mm的纤维占比分别为15.3%、12.7%,未见3~4 mm长度的纤维。竹肉中,0~1、2~3以及3~4 mm的纤维占比分别为4.8%、40.0%和2.3%。竹青中,0~1、2~3以及3~4 mm的纤维占比分别为22.7%、12.5%和1.3%。竹肉的纤维长宽比大于竹青和竹黄,且竹青和竹黄的纤维长宽比较为接近。
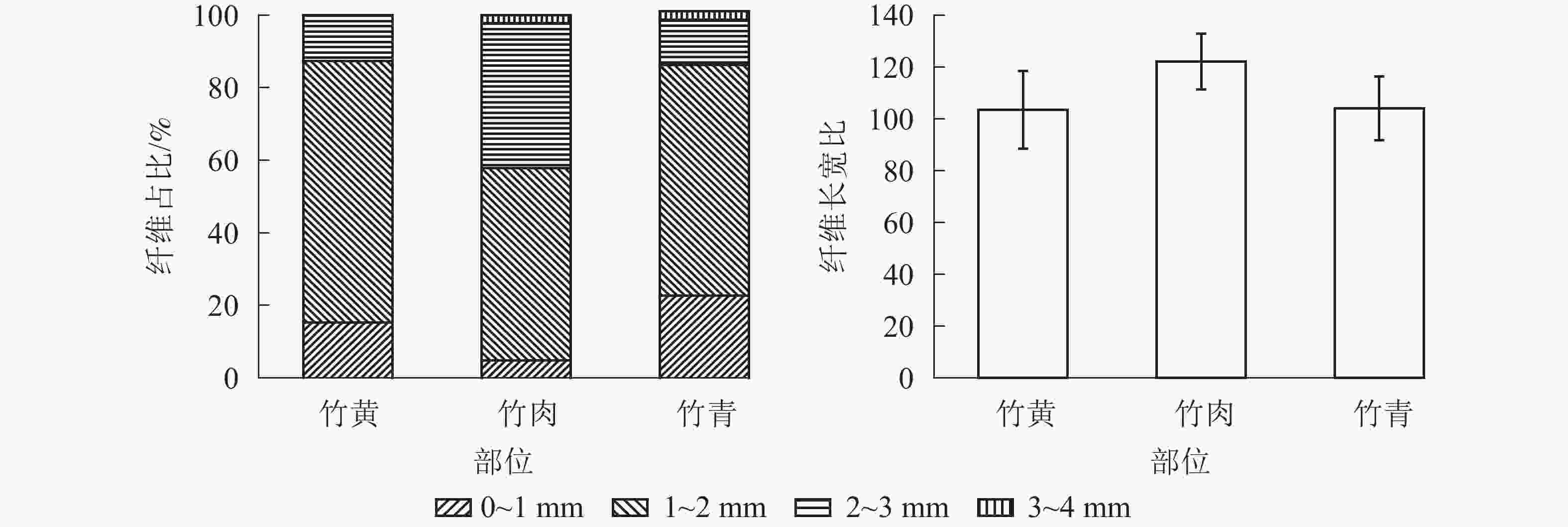
图 1 毛竹不同部位纤维长度分布频率和纤维长宽比
Figure 1. Fiber length distribution frequency and the ratio of fiber length to width in different parts of Ph. edulis
纸浆造纸过程中,纤维长度和长宽比是衡量纤维性能的重要指标。一般而言,随着纤维长度变长,其纸浆性能越佳[28]。同时,毛竹材纤维长度、宽度以及长宽比等参数亦与毛竹材结构、力学性能等指标有密切关系[29-30]。因此,工业化利用中应充分考虑不同部位纤维尺寸,并合理利用。
由图2可知:竹青维管束占比最大,其次为竹肉和竹黄。就薄壁细胞而言,竹黄部位薄壁细胞占比较大,为69.45%,竹青中薄壁细胞含量最小。竹黄中,维管束的面积较大,其均值为2.41×105 μm2,竹肉部位维管束平均面积为1.96×105 μm2,竹青部位的维管束平均面积为1.72×105 μm2。同时,竹黄部位维管束分布也更为稀疏[31]。由于维管束和薄壁细胞占比不同,以及维管束形态与尺寸差异,竹青、竹黄以及竹肉的力学性能差异极显著(P<0.01)[20, 32]。
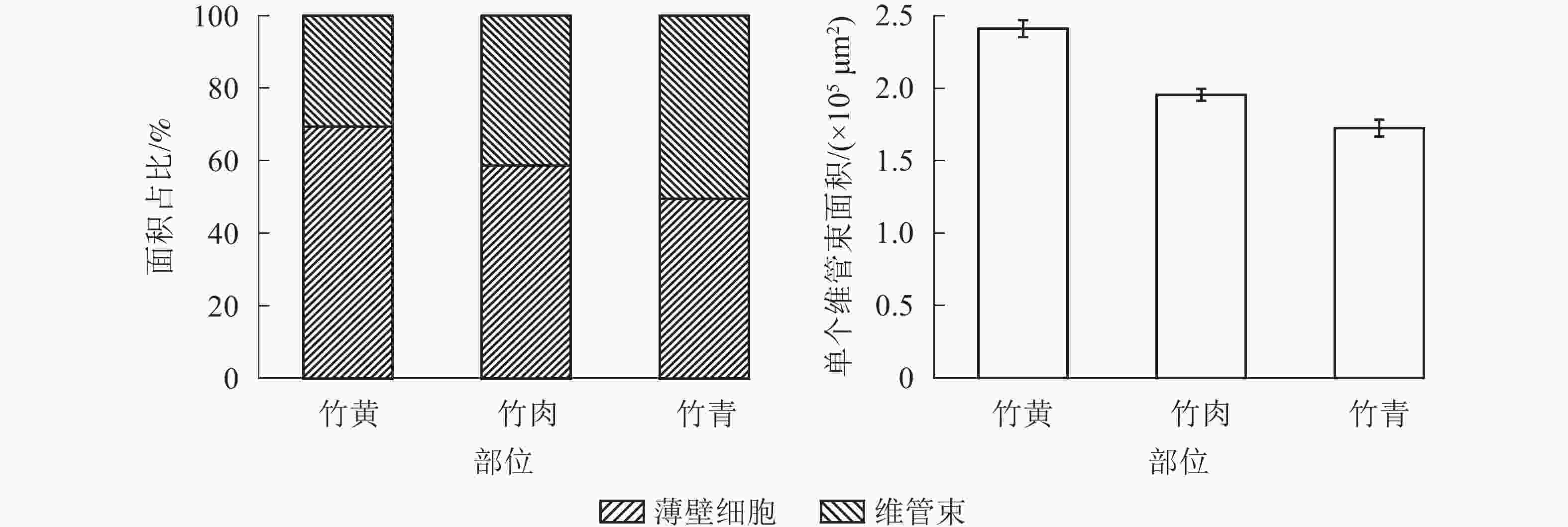
图 2 不同部位毛竹的维管束与薄壁细胞面积比例及单个维管束的面积
Figure 2. Area ratio of vascular bundle and parenchyma tissue in different parts of Ph. edulis and the area of single vascular bundle
-
从表2可见:毛竹不同试件的抗弯强度从小到大依次为C、A、D、B。原竹片(D)的抗弯强度为109.13 MPa,去青试件(A)抗弯强度骤减至57.77 MPa,去青去黄试件(C)的抗弯强度为48.19 MPa,而去黄试件(B)抗弯强度为115.28 MPa,较原竹片抗弯强度提高了5.5%。弹性模量呈现同样的变化趋势。由此可知,竹青对竹材的抗弯强度贡献最大,其次为竹肉和竹黄。竹材的梯度结构表现为维管束占比从竹黄到竹青呈上升趋势,薄壁细胞占比呈下降趋势[32]。因此,竹青在弯曲过程中,由于受拉侧(竹青)纤维较多,弯曲应变较大,弯曲模量表现较低值,对于弯曲强度贡献较大。竹黄由于薄壁细胞较多,弯曲过程中弯曲应变较小,弯曲弹性模量表现出较大值[20, 33-34]。
表 2 毛竹不同试件的抗弯强度和弹性模量
Table 2. Bending strength and elastic modulus of different Ph. edulis samples
试件 抗弯强度 弹性模量 平均值/MPa 标准差/MPa 变异系数/% 平均值/GPa 标准差/GPa 变异系数/% A 57.77 8.19 14.2 3.26 0.53 16.20 B 115.28 11.10 9.6 8.39 0.83 9.80 C 48.19 7.40 15.5 2.26 0.74 32.80 D 109.13 11.36 10.4 7.91 0.95 12.00 -
由表3可见:原竹片(D)顺纹抗压强度为59.54 MPa,去黄试件(B)顺纹抗压强度为59.36 MPa。去黄后,试件的顺纹抗压强度差异不明显,然而,去青试件(A)顺纹抗压强度明显下降,下降至38.35 MPa。因此,竹青对于毛竹的顺纹抗压强度具有较大贡献。去青试件顺纹抗压强度比去青去黄试件(C)略大,说明竹黄顺纹抗压强度较竹肉稍大。因此,竹制抗压构件设计制造过程中,合理利用竹黄对提高竹材利用率具有一定意义。
表 3 不同毛竹试件顺纹抗压强度
Table 3. Compressive strength of different Ph. edulis samples
试件 平均厚度/mm 顺纹抗压强度 均值/MPa 标准差/MPa 变异系数/% A 5.21 38.35 4.05 10.56 B 7.08 59.36 5.65 9.51 C 4.21 35.11 2.15 6.12 D 8.56 59.54 6.36 10.68 -
毛竹的干缩性对其加工利用具有重要影响,亦是衡量竹材产品性能及其稳定性的重要指标[35]。由于竹材中无横向组织,且径向和弦向干缩差异较大,竹材在加工利用时极易出现开裂或翘曲等问题[36]。
由表4和表5可知:从全湿状态至气干状态的过程中(即气干干缩率),竹黄的径向干缩率最大,为4.14%。3个方向的气干干缩率从大到小依次为径向、弦向、纵向;竹青变化规律亦如此。然而,竹肉和毛竹材3个方向的气干干缩率从大到小依次为弦向、径向、纵向。竹青、竹黄、竹肉以及毛竹的全干干缩率从大到小依次为径向、弦向、纵向,与前人研究结果一致[37]。无论气干干缩率还是全干干缩率,竹黄径向与弦向干缩率差值均最大,易产生翘曲变形等问题。因此,在后续加工利用中,合理配置竹黄结构对提高其结构稳定性至关重要。
表 4 毛竹竹黄、竹肉、竹青与毛竹气干干缩率测试结果
Table 4. Results of air-dry shrinkage rate for different Ph. edulis samples
部位 气干干缩率/% 平均值 标准差 变异系数 弦向 径向 纵向 体积 弦向 径向 纵向 体积 弦向 径向 纵向 体积 竹黄 1.55 4.14 0.44 7.47 0.20 0.23 0.01 1.02 10.34 8.26 11.01 17.25 竹肉 2.89 2.77 0.43 7.94 0.32 0.19 0.02 0.93 9.87 9.11 10.28 10.72 竹青 3.83 4.05 0.42 10.10 0.41 0.43 0.01 0.98 12.24 14.67 10.11 8.29 毛竹 3.15 2.11 0.43 6.37 0.43 0.18 0.03 0.78 10.26 15.24 13.56 9.43 表 5 毛竹竹黄、竹肉、竹青与毛竹全干干缩率测试结果
Table 5. Results of total-dry shrinkage rate for different Ph. edulis samples
部位 全干干缩率/% 平均值 标准差 变异系数 弦向 径向 纵向 体积 弦向 径向 纵向 体积 弦向 径向 纵向 体积 竹黄 2.58 6.60 1.03 8.76 0.28 0.46 0.03 1.21 14.27 10.66 13.28 12.21 竹肉 4.99 6.81 0.84 8.80 0.36 0.64 0.01 0.95 13.72 10.92 9.92 13.46 竹青 7.57 7.98 0.63 13.70 0.99 0.55 0.04 1.53 11.98 13.29 8.91 12.18 毛竹 5.25 5.76 0.81 11.39 0.63 0.59 0.03 0.95 9.89 14.31 12.21 13.62 -
毛竹不同部位的纤维形态及部分物理性能存在一定差异。毛竹竹黄、竹肉与竹青的纤维长度和宽度差异显著,且不同长度的纤维占比和纤维长宽比亦存在着差异。毛竹材利用过程中需要根据应用领域的不同,合理选择相应部位,以进一步提高利用效率。竹青对竹材抗弯强度与抗弯弹性模量贡献最大。气干干缩率和全干干缩率在竹材不同部位、不同方向上均存在一定差异。未来可开展不同剖篾状态下,竹黄、竹青含量对竹质、木竹复合工程材料力学性能、尺寸稳定性等性能影响规律的研究,对提高竹材利用率具有一定意义。
Differences in fiber morphology and partial physical properties in different parts of Phyllostachys edulis
-
摘要:
目的 探明毛竹Phyllostachys edulis竹青、竹黄及竹肉不同部位的纤维形态、力学性能以及干缩性能等差异,为毛竹材高效利用提供基础数据。 方法 通过纤维离析与显微观察、力学性能与尺寸稳定性测试,分析比较毛竹材不同部位性能差异。 结果 毛竹材竹黄、竹肉与竹青不同部位中,纤维长度和宽度以及纤维占比差异极显著(P<0.01)。竹青和竹黄的纤维长宽比较为接近,且极显著小于竹肉(P<0.01)。竹青密布维管束,对毛竹材抗弯强度、弹性模量贡献最为大,其次为竹肉和竹黄。就顺纹抗压强度而言,从大到小依次为竹青、竹黄、竹肉。竹材横向干缩性明显大于纵向,全干干缩率从大到小依次为径向、弦向、纵向。竹材不同部位中,径向和弦向气干干缩率的大小关系略有差异。 结论 毛竹材不同部位性能差异明显,竹黄抗压力学性能优于竹肉,可将竹黄保留用于制备新型竹木复合材料,有助于提高竹材利用率。图2表5参37 Abstract:Objective With an investigation of the differences of fiber morphology, mechanical properties and dry shrinkage properties in different parts of moso bamboo(Phyllostachys edulis), this study is aimed to provide basic data for the efficient utilization of moso bamboo. Method With fiber segregation, microscopic observation as well as tests of the mechanical properties and dimensional stability, a comparative analysis was conducted of the performance differences. Result The fiber length, width, and fiber proportion parameters were significantly different in different parts of bamboo (P<0.01). The length-width ratio of fiber in bamboo outer layer was similar to that in the inner layer, but significantly lower than that in the middle layer(P<0.01). Because of the dense vascular bundle distribution, the bamboo outer layer had the most significant contribution to the flexural strength and elastic modulus of bamboo samples, followed by the bamboo middle layer and then the bamboo inner layer. In terms of compression strength parallel to grain, the outer layer outperforms the inner layer and the middle layer (with a successive decrease). The horizontal dry shrinkage of bamboo was significantly greater than that of the longitudinal, and the total dry shrinkage was the largest in radial direction, followed by that of tangential direction and longitudinal direction. In different parts of the bamboo, the air-drying shrinkage rate in radial direction is slightly different from that in the tangential direction. Conclusion The properties of different parts of bamboo are obviously different, and the compressive property of the inner layer is better than that of the middle layer and with a certain structural design, the inner layer can be retained to prepare new bamboo-wood composite materials, which is conducive to improving the utilization rate of bamboo. [Ch, 2 fig. 5 tab. 37 ref.] -
Key words:
- wood science /
- Phyllostachys edulis /
- fiber morphology /
- mechanical property /
- bamboo outer layer /
- bamboo inner layer
-
图 1 毛竹不同部位纤维长度分布频率和纤维长宽比
Figure 1 Fiber length distribution frequency and the ratio of fiber length to width in different parts of Ph. edulis
图 2 不同部位毛竹的维管束与薄壁细胞面积比例及单个维管束的面积
Figure 2 Area ratio of vascular bundle and parenchyma tissue in different parts of Ph. edulis and the area of single vascular bundle
表 1 毛竹材各部位纤维平均尺寸
Table 1. Mean size of fiber in different parts of Ph. edulis
部位 长度/mm 宽度/μm 竹黄 1.48±0.45 b 14.01±2.75 b 竹肉 1.88±0.56 a 15.15±3.53 a 竹青 1.43±0.53 b 13.40±3.47 b 说明:数值为平均值±标准差。不同字母表示部位间差异极 显著(P<0.01)  下载: 导出CSV
下载: 导出CSV
表 2 毛竹不同试件的抗弯强度和弹性模量
Table 2. Bending strength and elastic modulus of different Ph. edulis samples
试件 抗弯强度 弹性模量 平均值/MPa 标准差/MPa 变异系数/% 平均值/GPa 标准差/GPa 变异系数/% A 57.77 8.19 14.2 3.26 0.53 16.20 B 115.28 11.10 9.6 8.39 0.83 9.80 C 48.19 7.40 15.5 2.26 0.74 32.80 D 109.13 11.36 10.4 7.91 0.95 12.00
下载: 导出CSV
表 3 不同毛竹试件顺纹抗压强度
Table 3. Compressive strength of different Ph. edulis samples
试件 平均厚度/mm 顺纹抗压强度 均值/MPa 标准差/MPa 变异系数/% A 5.21 38.35 4.05 10.56 B 7.08 59.36 5.65 9.51 C 4.21 35.11 2.15 6.12 D 8.56 59.54 6.36 10.68
下载: 导出CSV
表 4 毛竹竹黄、竹肉、竹青与毛竹气干干缩率测试结果
Table 4. Results of air-dry shrinkage rate for different Ph. edulis samples
部位 气干干缩率/% 平均值 标准差 变异系数 弦向 径向 纵向 体积 弦向 径向 纵向 体积 弦向 径向 纵向 体积 竹黄 1.55 4.14 0.44 7.47 0.20 0.23 0.01 1.02 10.34 8.26 11.01 17.25 竹肉 2.89 2.77 0.43 7.94 0.32 0.19 0.02 0.93 9.87 9.11 10.28 10.72 竹青 3.83 4.05 0.42 10.10 0.41 0.43 0.01 0.98 12.24 14.67 10.11 8.29 毛竹 3.15 2.11 0.43 6.37 0.43 0.18 0.03 0.78 10.26 15.24 13.56 9.43
下载: 导出CSV
表 5 毛竹竹黄、竹肉、竹青与毛竹全干干缩率测试结果
Table 5. Results of total-dry shrinkage rate for different Ph. edulis samples
部位 全干干缩率/% 平均值 标准差 变异系数 弦向 径向 纵向 体积 弦向 径向 纵向 体积 弦向 径向 纵向 体积 竹黄 2.58 6.60 1.03 8.76 0.28 0.46 0.03 1.21 14.27 10.66 13.28 12.21 竹肉 4.99 6.81 0.84 8.80 0.36 0.64 0.01 0.95 13.72 10.92 9.92 13.46 竹青 7.57 7.98 0.63 13.70 0.99 0.55 0.04 1.53 11.98 13.29 8.91 12.18 毛竹 5.25 5.76 0.81 11.39 0.63 0.59 0.03 0.95 9.89 14.31 12.21 13.62
下载: 导出CSV
-
[1] 郝际平, 申国臣, 田黎敏, 等. 毛竹力学强度标准值和设计值的确定方法[J]. 安徽农业大学学报, 2020, 47(1): 56 − 61. HAO Jiping, SHEN Guochen, TIAN Limin, et al. Methods for determination of mechanical strength standard value and design value of bamboo [J]. J Anhui Agric Univ, 2020, 47(1): 56 − 61. [2] 刘娇, 周爱萍, 盛宝璐, 等. 温度对重组竹短期受压蠕变性能的影响[J]. 林业工程学报, 2021, 6(2): 64 − 69. LIU Jiao, ZHOU Aiping, SHENG Baolu, et al. Effect of temperature on short-term compression creep property of bamboo scrimber [J]. J For Eng, 2021, 6(2): 64 − 69. [3] 闵余晨, 宋莎莎, 费本华. 竹编在家具设计中的创新应用研究[J]. 林产工业, 2020, 57(4): 47 − 50. MIN Yuchen, SONG Shasha, FEI Benhua. Research on innovative application of bamboo weaving in furniture products [J]. China For Prod Ind, 2020, 57(4): 47 − 50. [4] 吴婕妤, 陈红, 费本华, 等. 浙江地区竹编工艺特色概述[J]. 林产工业, 2020, 57(3): 61 − 64. WU Jieyu, CHEN Hong, FEI Benhua, et al. Study on bamboo weaving process characteristics in Zhejiang Province [J]. China For Prod Ind, 2020, 57(3): 61 − 64. [5] 李海涛, 宣一伟, 许斌, 等. 竹材在土木工程领域的应用[J]. 林业工程学报, 2020, 5(6): 1 − 10. LI Haitao, XUAN Yiwei, XU Bin, et al. Bamboo application in civil engineering field [J]. J For Eng, 2020, 5(6): 1 − 10. [6] 娄志超, 袁成龙, 李延军, 等. 饱和蒸汽热处理对竹束化学成分和结晶度的影响[J]. 林业工程学报, 2020, 5(2): 29 − 35. LOU Zhichao, YUAN Chenglong, LI Yanjun, et al. Effect of saturated steam treatment on the chemical composition and crystallinity properties of bamboo bundles [J]. J For Eng, 2020, 5(2): 29 − 35. [7] 张雨湉, 陈芝贤, 费本华, 等. 中国不同地域竹编工艺的发展现状[J]. 世界竹藤通讯, 2018, 16(6): 37 − 41. ZHANG Yutian, CHEN Zhixian, FEI Benhua, et al. Development status of bamboo weaving process in different regions of China [J]. World Bamboo Rattan, 2018, 16(6): 37 − 41. [8] 吴婕妤, 陈红, 吴智慧, 等. 四川地区竹编工艺特性研究[J]. 竹子学报, 2020, 39(1): 90 − 94. WU Jieyu, CHEN Hong, WU Zhihui, et al. Bamboo weaving technology in Sichuan Province [J]. J Bamboo Res, 2020, 39(1): 90 − 94. [9] 吴为, 徐伟, 康雨, 等. 非木质材料在茶家具中的应用和设计实践[J]. 林业机械与木工设备, 2020, 48(12): 54 − 58, 64. WU Wei, XU Wei, KANG Yu, et al. Application and design practice of non-wood materials in tea furniture [J]. For Mach Woodworking Equip, 2020, 48(12): 54 − 58, 64. [10] WANG Xinzhou, CHENG Dali, HUANG Xianai, et al. Effect of high-temperature saturated steam treatment on the physical, chemical, and mechanical properties of moso bamboo[J]. J Wood Sci, 2020, 66: 52. doi: 10.1186/s10086-020-01899-8. [11] 曹钰, 王新洲, 李延军, 等. 高温油热处理对竹材淀粉含量及防霉性能的影响[J]. 林业工程学报, 2020, 5(2): 109 − 115. CAO Yu, WANG Xinzhou, LI Yanjun, et al. Effect of high temperature oil heat treatment on the starch content and mold-resistant property of bamboo [J]. J For Eng, 2020, 5(2): 109 − 115. [12] 李媛媛, 张双燕, 王传贵, 等. 毛竹采伐剩余物的化学成分、纤维形态及纸浆性能[J]. 浙江农林大学学报, 2019, 36(2): 219 − 226. LI Yuanyuan, ZHANG Shuangyan, WANG Chuangui, et al. Chemical composition, fiber morphology, and pulping properties of logging residues inPhyllostachys edulis [J]. J Zhejiang A&F Univ, 2019, 36(2): 219 − 226. [13] 吴杉杉, 吴婧怡. 新形势下如何更好对竹文化体育资源进行保护性开发[J]. 林产工业, 2020, 57(10): 97 − 99. WU Shanshan, WU Jingyi. How to take better protective development of bamboo culture and sports resources under the new situation [J]. China For Prod Ind, 2020, 57(10): 97 − 99. [14] 刘国苹. 传统竹家具制造工艺对现代生产工艺的影响[J]. 林产工业, 2020, 57(2): 95 − 97. LIU Guoping. The influence of traditional bamboo furniture manufacturing technology on modern production technology [J]. China For Prod Ind, 2020, 57(2): 95 − 97. [15] 王晶, 吴燕, 王雨欣, 等. 透明竹材的研究进展及其家居产品设计[J]. 家具, 2021, 42(2): 52 − 56. WANG Jing, WU Yan, WANG Yuxin, et al. Research progress of transparent bamboo and home product design [J]. Furniture, 2021, 42(2): 52 − 56. [16] 许艳青, 陈红, 杨峰. 低温等离子体处理对竹丝装饰材表面性能的影响[J]. 家具, 2019, 40(4): 21 − 27. XU Yanqing, CHEN Hong, YANG Feng. Effects of low temperature plasma treatment on the surface properties of bamboo filament decorative materials [J]. Furniture, 2019, 40(4): 21 − 27. [17] 赵鹤, 林金国, 陈礼辉. 竹青竹黄加工利用关键问题的探讨[J]. 家具, 2014, 35(6): 11 − 14, 43. ZHAO He, LIN Jinguo, CHEN Lihui. The key issues about comprehensive utilization of the outer and the inner of bamboo [J]. Furniture, 2014, 35(6): 11 − 14, 43. [18] WANG Fuli, SHAO Zhuoping. Study on the variation law of bamboo fibers’ tensile properties and the organization structure on the radial direction of bamboo stem[J]. Ind Crops Prod, 2020, 152(4): 112521. doi: 10.1016/j.indcrop.2020.112521. [19] 袁晶, 张雪霞, 余雁, 等. 维管束分布及结构对竹材宏观压缩性能的影响[J]. 中南林业科技大学学报, 2019, 39(6): 121 − 127. YUAN Jing, ZHANG Xuexia, YU Yan, et al. Effects of the distribution and structure of vascular bundles on the compressive properties of bamboo [J]. J Cent South Univ For Technol, 2019, 39(6): 121 − 127. [20] 陈秋艳, 魏璐, 苗庆显, 等. 绿竹不同部位竹材的物理化学特性研究[J]. 中国造纸学报, 2016, 31(3): 1 − 6. CHEN Qiuyan, WEI Lu, MIAO Qingxian, et al. Physical and chemical properties of different parts of green bamboo [J]. Trans China Pulp Pap, 2016, 31(3): 1 − 6. [21] 马弘帅. 竹材梯度结构对弯曲力学性能的影响[J]. 世界竹藤通讯, 2020, 18(3): 50 − 54. MA Hongshuai. The influence of bamboo gradient structure on flexure modulus [J]. World Bamboo Rattan, 2020, 18(3): 50 − 54. [22] 夏旭光, 姚文斌, 俞伟鹏, 等. 非均匀竹材各单层顺纹抗压弹性模量的测定[J]. 竹子学报, 2018, 37(4): 49 − 55. XIA Xuguang, YAO Wenbin, YU Weipeng, et al. The measurement of the parallel grain compression MOE of each layer of non-uniform bamboo [J]. J Bamboo Res, 2018, 37(4): 49 − 55. [23] GUTIERREZ M, MALUK C. Mechanical behaviour of bamboo at elevated temperatures-experimental studies [J]. Eng Struct, 2020, 220: 110997. doi: 10.1016/j.engstruct.2020.110997. [24] 李权, 林金国, 刘主凰, 等. 福建主要竹材纤维化学性质的差异分析[J]. 竹子研究汇刊, 2014, 33(2): 5 − 10. LI Quan, LIN Jinguo, LIU Zhuhuang, et al. The difference in fiber properties of major bamboo species in Fujian Province [J]. J Bamboo Res, 2014, 33(2): 5 − 10. [25] LIAN Caiping, LIU Rong, LUO Junji, et al. The morphological characteristics and classification of vascular parenchyma cells in bamboo [Phyllostachys edulis (Carr.) J. Houz. ] [J]. Holzforschung, 2020, 74(9): 829 − 838. [26] 严彦, 刘焕荣, 张秀标, 等. 毛竹材性差异对胶合竹层板应力分级的影响[J]. 安徽农业大学学报, 2017, 44(2): 260 − 264. YAN Yan, LIU Huanrong, ZHANG Xiubiao, et al. The effect of Phyllostachys pubescens f. lutea Wen property on the E-Rated classification of laminated bamboo board [J]. J Anhui Agric Univ, 2017, 44(2): 260 − 264. [27] 陈冠军, 袁晶, 余雁, 等. 竹材顺纹抗压性能的种间差异及其影响因子研究[J]. 木材加工机械, 2018, 29(6): 23 − 27. CHEN Guanjun, YUAN Jing, YU Yan, et al. Study on interspecific differences of the compression performance parallel to the grain of bamboo and its influencing factors [J]. Wood Process Mach, 2018, 29(6): 23 − 27. [28] 陈丽娟, 张朝燕, 赖永平, 等. 四川6种丛生竹的纤维形态研究[J]. 西北农林科技大学学报(自然科学版), 2015, 43(5): 79 − 86. CHEN Lijuan, ZHANG Chaoyan, LAI Yongping, et al. Fiber morphology of six sympodial bamboos in Sichuan [J]. J Northwest A&F Univ Nat Sci Ed, 2015, 43(5): 79 − 86. [29] 陈铭, 郭琳, 郑笑, 等. 中国15个主产区毛竹纤维形态比较[J]. 南京林业大学学报(自然科学版), 2018, 42(6): 7 − 12. CHEN Ming, GUO Lin, ZHENG Xiao, et al. Comparison of cell morphology of moso bamboo fibers from fifteen main producing regions in China [J]. J Nanjing For Univ Nat Sci Ed, 2018, 42(6): 7 − 12. [30] 甘小洪, 陈凤, 林树燕, 等. 慈竹不同变异类型的纤维形态研究[J]. 南京林业大学学报(自然科学版), 2013, 37(4): 99 − 104. GAN Xiaohong, CHEN Feng, LIN Shuyan, et al. Fiber morphology of different variation types of Neosinocalamus affinis (Rendle) Keng f. [J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(4): 99 − 104. [31] LIAN Caiping, LIU Rong, ZHANG Shuqin, et al. Ultrastructure of parenchyma cell wall in bamboo (Phyllostachys edulis) culms [J]. Cellulose, 2020, 27(13): 7321 − 7329. [32] 刘焕荣, 杨晓梦, 张秀标, 等. 竹展平板拉伸剪切胶合性能[J]. 林业工程学报, 2021, 6(1): 68 − 72. LIU Huanrong, YANG Xiaomeng, ZHANG Xiubiao, et al. The tensile shear bonding property of flattened bamboo sheet [J]. J For Eng, 2021, 6(1): 68 − 72. [33] 岳祥华, 左奇玉, 张双燕. 油浴热处理对竹材干缩性和力学性能的影响[J]. 世界竹藤通讯, 2020, 18(1): 11 − 15. YUE Xianghua, ZUO Qiyu, ZHANG Shuangyan. Influence of oil heat treatment on shrinkage and mechanical properties of bamboo [J]. World Bamboo Rattan, 2020, 18(1): 11 − 15. [34] CHEN Meiling, YE Ling, LI Hui, et al. Flexural strength and ductility of moso bamboo [J]. Constr Build Mater, 2020, 246: 118418. doi: 10.1016/j.conbuildmat.2020.118418. [35] 崔敏, 殷亚方, 姜笑梅, 等. 不同竹龄毛竹材物理性质的差异分析[J]. 福建林学院学报, 2010, 30(4): 338 − 343. CUI Min, YIN Yafang, JIANG Xiaomei, et al. Variation analysis of physical characteristics in Phyllostachy pubescens stem at different growth ages [J]. J Fujian Coll For, 2010, 30(4): 338 − 343. [36] ZHANG Wenbo, TIAN Genlin, POLLE A, et al. Comparative characterization of ethanol organosolv lignin polymer from bamboo green, timber and yellow [J]. Wood Sci Technol, 2018, 52(5): 1331 − 1341. [37] 高珊珊, 郑仁红, 吴晓丽, 等. 4种大径丛生竹材的密度和干缩性研究[J]. 福建林学院学报, 2010, 30(3): 270 − 274. GAO Shanshan, ZHENG Renhong, WU Xiaoli, et al. Study on density and shrinkage of the 4 species of big sympodial bamboo [J]. J Fujian Coll For, 2010, 30(3): 270 − 274. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200649
 点击查看大图
点击查看大图
计量
- 文章访问数: 2564
- HTML全文浏览量: 776
- PDF下载量: 89
- 被引次数: 0
