





-
竹子具备高强度、高韧性等优点,并且生长速度快,可快速成材,被广泛应用于建筑、家具和造纸等领域[1−3]。作为21世纪最具开发潜力的生物质资源,竹材具有不可比拟的生态、经济以及社会效益[4],对促进碳达峰、碳中和具有极大的意义。竹材被认为是以薄壁组织为基体,嵌入维管束作为增强相的纤维增强生物复合材料[5−6]。维管束起着承担力学强度的骨架作用,基本组织填充其余部分,起着传递载荷的作用[7]。竹材优异的力学性能主要与纤维鞘组织比量密切相关[8]。目前,竹产业主要生产传统竹胶合板、竹席/竹帘胶合板、竹集成材和重组竹等或新型工程竹复合材料[9]、复合吸波材料[10]和竹缠绕复合管[11]等,大多以毛竹Phyllostachys edulis等大径级的竹材为原料[12−13],小径级的竹材应用较少。竹材的力学性能决定其利用价值与领域[14],竹材顺纹抗压强度作为评估竹材不同用途的适用性的重要指标[15−16],具有测试方便快速等优点[17]。小径级竹材物理力学性能测试并无相关标准,相关科学研究较少,基本上都参照GB/T 15780—1995《竹材物理力学性质试验方法》[18−19],缺乏评价小径级竹材的系统的物理力学实验方法。
本研究对不同长径比试件进行顺纹抗压试验,探索适合测试小径级竹材顺纹抗压强度的方法,为发掘小径级竹材工业应用提供科学依据[20]。同时利用基于YOLO深度学习算法开发的维管束识别模型对小径级竹材纤维鞘的组织比量和维管束密度作出统计并对其与抗压强度的关系进行线性拟合。通过引入人工智能用于竹材基础性质的研究,不仅可以解决人工处理耗时费力以及容易出错等问题,相较于传统图像处理方法还能提高处理精度和准确性。
-
样品采自安徽省六安市金寨县。随机选取健康、完好无缺陷的3~4年生处于成熟期的竹株,采集胸径在50 mm以下的苦竹Pleioblastus amarus、篌竹Phyllostachys nidularia、水竹Phyllostachys heteroclada和早园竹Phyllostachys propinqua各5株,平均直径分别为12.04、30.44、19.92、33.02 mm,平均壁厚分别为2.38、3.90、3.24、4.58 mm。从离地约0.5 m处向上截取2.0~4.0 m竹段作为试样,并对试样进行标号带回实验室待用。
-
用游标卡尺测量试样同一点在相互垂直的2个方向的直径,求其平均值。沿顺纹方向按上述平均直径的1.0、1.5和2.0倍长度截制不同长径比试件,试件两端面平整并相互平行,端面应与顺纹方向垂直(图1)。参照 GB/T 15780—1995《竹材物理力学性质试验方法》[18]调整试样含水率。

图 1 试件
Figure 1. Samples
-
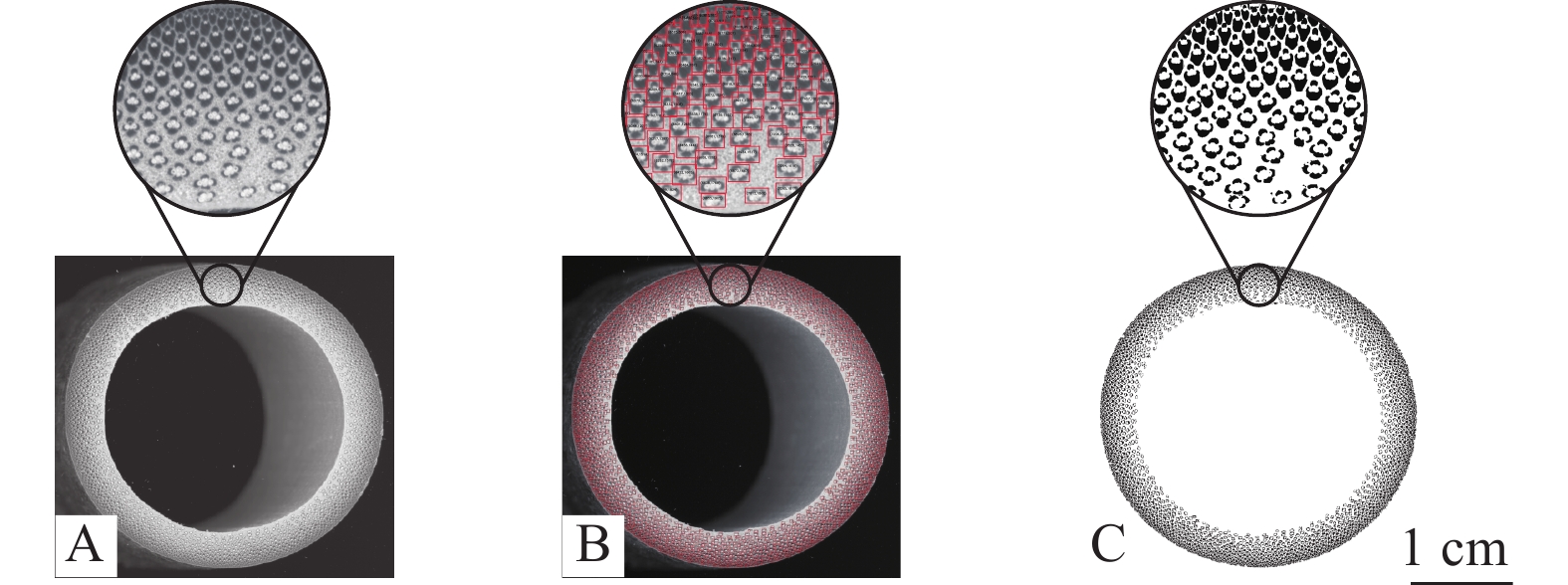
对截断后的横截面用不同目数的砂纸砂光处理,最后一次精砂目数为320目,直至触摸手感光滑,既保证精确识别纤维鞘和薄壁细胞之间的分界线,又可以利用砂光产生的碎屑填充维管束中的大导管及韧皮部等孔隙,避免大导管等干扰计算机识别[21]。利用高清扫描仪对符合要求的试件横截面进行扫描,用局部聚类算法对图片进行二值化处理,采用经过训练的模型进行识别,检测维管束并统计纤维鞘总面积[22](图2)。

图 2 高清灰度图(A)、识别每个维管束的检测图(B)和利用局部聚类算法获得的二值化图(C)
Figure 2. HD grayscale image (A), detection image identifying each vascular bundle (B) and binarized image obtained using local clustering algorithm (C)
-
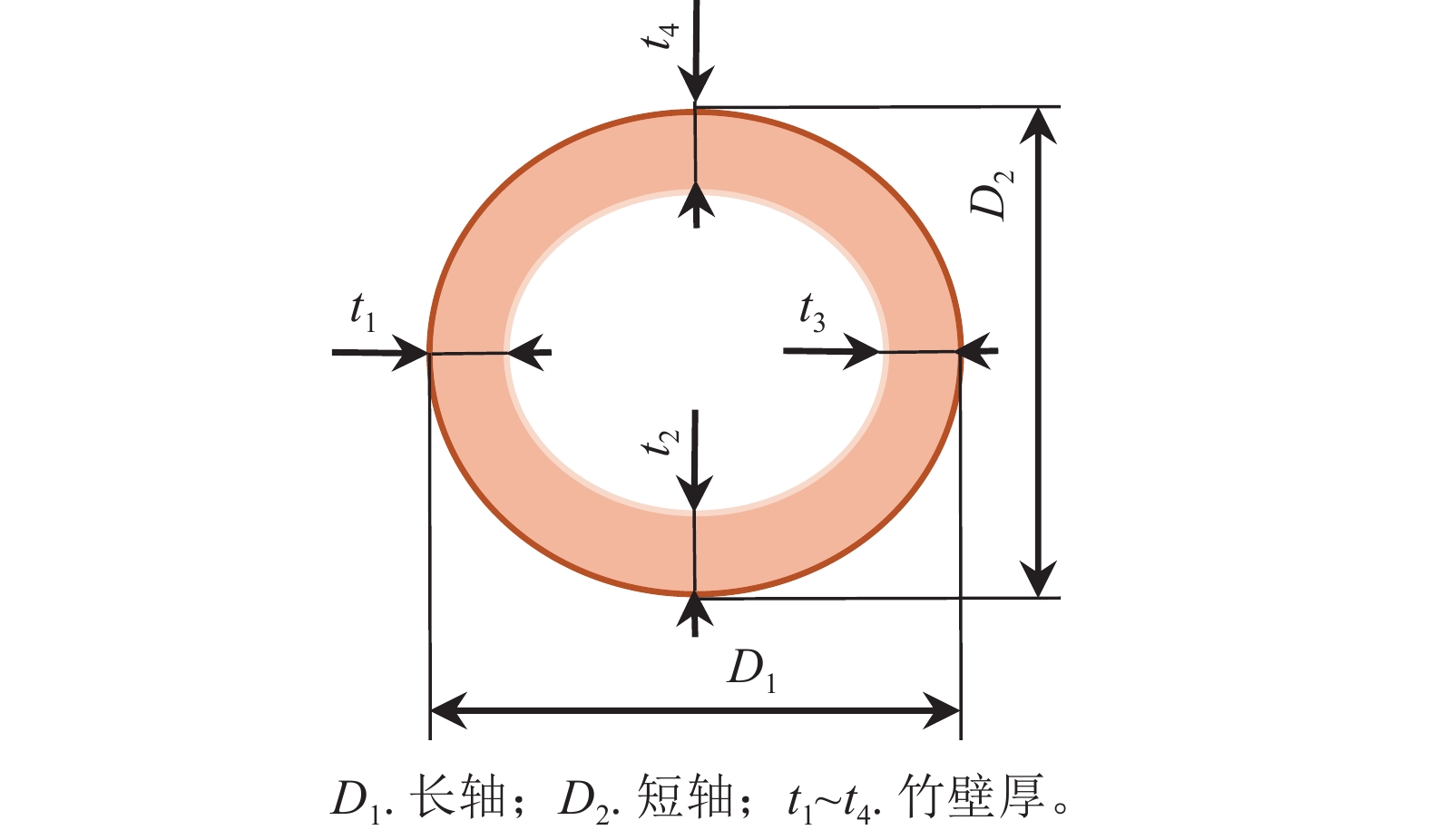
①尺寸测量。如图3所示:在扫描计算完的试件两端用游标卡尺测其相互垂直的2个方向长轴(D1)和短轴(D2),以同样方法测定试件两端竹壁厚(t1、t2、t3、t4)取其均值(t),准确至0.01 mm,计算试件两端面横截面面积均值。将横截面近似为椭圆计算。②顺纹抗压试验。将试件垂直放置于万能力学试验机球面滑动支座中心位置,调整下支座,从相互垂直的2个方向观察,使试件横截面与压头表面平行,施压方向和纤维方向平行。施加预荷载不大于10.0 N,调整放好试件。沿试件轴向以1 mm·min−1的速度均匀加载,在(90±30) s内破坏,记录试件破坏的最大载荷,精确至0.1 N。③含水率测定。抗压性能试验结束后,立即清理试件上易剥落物,进行称量,准确至0.001 g。测定含水率。测定方法依据GB/T 15780—1995《竹材物理力学性质试验方法》。④计算。顺纹抗压强度按近似椭圆管状件计算。采用$ \sigma_{\text{12}}\text{=}\sigma_{{w}}\left[\text{1+0.045}\left({w}-\text{12}\right)\right] $将含水率为w的顺纹抗压强度$ \sigma_{{w}}$换算为试件含水率为12%的顺纹抗压强度$\sigma_{{12}} $,准确至0.01 MPa。$ \sigma_{{w}}\text{=}\dfrac{\text{2}{{F}}_{\text{max}}}{\text{π}t\left({{D}}_{\text{1}}\text{+}{{D}}_{\text{2}}{-}{2t}\right)} $。其中:$ {{F}}_{\text{max}} $为试件破坏最大载荷(N);$ {t} $为试件平均壁厚(mm);$ {{D}}_{\text{1}} $为试件横截面长轴(mm);$ {{D}}_{\text{2}} $为试件横截面短轴(mm)。

图 3 尺寸测量示意图
Figure 3. Dimension measurement schematic
-
由表1可见:4种小径级竹材的顺纹抗压强度较为优异,采用2.0倍长径比试件测试,早园竹顺纹抗压强度最大,为82.91 MPa,水竹顺纹抗压强度最小,为67.01 MPa。在不同长径比的试件中,顺纹抗压强度测试结果平均值相差不大。方差齐性检验表明:4种小径级竹材的不同长径比试件的顺纹抗压强度差异不显著(P>0.05),故进行单因素ANOVA检验。检验结果显示:除苦竹外,其余3种小径级竹材的不同长径比试件对顺纹抗压强度没有产生显著影响(P>0.05)。然而,苦竹不同长径比试件对顺纹抗压强度产生显著影响(F=3.342,P=0.040<0.05)。进行LSD多重比较分析苦竹不同长径比试样之间的差异是否显著,结果显示:苦竹的1.0倍长径比试件与其他2种试件之间差异不显著,而1.5与2.0倍试件之间产生显著差异(P=0.012<0.05)。这可能是由于苦竹平均直径仅为12.04 mm,直径太小并且制备设备落后,对试件造成了不同程度的破坏。
表 1 顺纹抗压测试结果
Table 1. Test results of compressive strength
竹种 长径比 顺纹抗压强度 方差齐性
检验P单因素
ANOVA检验平均值/
MPa标准差/
MPa变异系
数/%F P 苦竹 1.0 71.64 ab 3.80 5.30 0.456 3.342 0.040 1.5 70.27 b 3.20 4.55 2.0 72.65 a 3.89 5.35 篌竹 1.0 74.57 a 8.02 10.76 0.989 0.239 0.788 1.5 74.01 a 7.68 10.38 2.0 73.19 a 7.59 10.37 水竹 1.0 67.09 a 5.75 8.57 0.776 0.222 0.801 1.5 67.78 a 6.01 8.87 2.0 67.01 a 5.12 7.64 早园竹 1.0 83.29 a 9.52 11.43 0.528 0.010 0.990 1.5 83.06 a 8.19 9.86 2.0 82.91 a 7.76 9.36 说明:不同小写字母表示同一竹种不同处理之间LSD检验差异显著(P<0.05)。 除去由于试件制备中引起的破坏而产生的测量误差,不同长径比的试样对顺纹抗压强度没有产生显著影响。根据表1结果所示:除苦竹外,其他3种竹材均是2.0倍长径比的试件测试的顺纹抗压强度的变异系数最小。其中,篌竹、早园竹和水竹2.0倍长径比试件的顺纹抗压强度变异系数分别为10.37%、9.36%和7.64%。苦竹的3种试件变异系数均较小,1.5倍长径比试件变异系数为4.55%。变异系数越小,说明数据越稳定。早园竹和篌竹的标准差在一定范围内随着长径比的增加而减少。具体而言,2.0倍长径比试件的标准差分别为7.76和7.59 MPa。水竹的2.0倍长径比试件最小,为5.12 MPa。苦竹标准差是1.5倍长径比试件最小,为3.20 MPa。标准差越小,试验数据的离散程度越小。综上所述,2.0倍长径比的试件测试的数据比较稳定,可靠性较好,变异系数小,意味着需要准备的最小试件数量就最少,整个试验的操作就相对容易,试验数据更加准确。因此,2.0倍长径比的试件用于测试小径级圆竹顺纹抗压强度最好。
-
从长径比为2.0的试样的维管束参数 (表2) 可以看出:4种竹材的纤维鞘组织比量的标准偏差均较小,说明同一竹种不同竹株不同部位的纤维鞘组织比量相差不大。其中篌竹纤维鞘组织比量最大,为35.64%,最小的是水竹,为33.05%。苦竹的维管束分布密度最大,达7.94 个·mm−2,早园竹最小,为5.77 个·mm−2。
表 2 4种小径级竹材长径比为2.0试样的维管束参数
Table 2. Vascular bundle parameters of four species of small diameter bamboo at the length-to-diameter ratio of 2.0
竹种 纤维鞘体积分数/% 维管束分布密度/(个·mm−2) 平均值 标准偏差 平均值 标准偏差 苦竹 34.61 2.18 7.94 0.73 篌竹 35.64 1.51 6.32 0.92 水竹 33.05 2.15 7.01 0.31 早园竹 34.65 5.53 5.77 1.07 如图4所示:纤维鞘体积分数 (y) 与顺纹抗压强度 (x) 之间呈线性正相关,即纤维鞘体积分数越大,顺纹抗压强度越大。基于最小二乘法对其进行线性拟合,得到线性方程:y = 260.44x−18.26,R2=0.60,r=0.778,P<0.001。维管束分布密度和顺纹抗压强度之间的相关性并不强,可能原因是不同竹种的维管束面积以及类型不同。换而言之,不同的竹种即使维管束分布密度相同,也有可能单位面积内所含的纤维含量不同。

图 4 顺纹抗压强度与纤维鞘体积分数和维管束分布密度之间的分布规律
Figure 4. Distribution pattern between the compressive strength and the fiber volume fraction and distribution density of vascular bundles
-
本研究结果发现:4种竹材的顺纹抗压强度较为优异,高于同为圆竹形态测试的4年生毛竹的基部试件的顺纹抗压强度 (48.54 MPa)[23]。 竹子之所以具有轻质且高强度的特性,是由于其薄壁中空和功能梯度结构等优势。竹壁的构造包括表皮、维管束和基本组织等3种组织[24],其中富含二氧化硅的表皮起着保护竹子抵御外界环境破坏的作用,维管束则承担提供力学强度的骨架作用,基本组织则填充其余部分并起着传递载荷的作用[7]。竹材作为一种典型的单向纤维增强的生物质复合材料,其机械性能主要取决于呈梯度结构分布的维管束的机械特性,如纤维鞘的体积分数和维管束的分布密度等[21, 25]。纤维密度是表征竹子强度性能的良好指标[26],纤维鞘体积分数正是纤维密度的定量化表述。已有研究对不同纤维鞘体积分数的毛竹进行了顺纹抗压测试,发现顺纹抗压强度和模量随纤维鞘体积分数增加而线性增加的规律[27]。同样,具有梯度结构的竹材在抗弯强度和模量中随着纤维体积分数的增加而增加[28]。此外,针对不同纤维鞘体积分数的毛竹样品的拉伸测试发现,纤维鞘体积分数和拉伸强度之间存在明显的线性关系,并且根据混合定律得出的纤维和基本组织的拉伸强度和MOE分别为581.7 MPa、40.40 GPa、19.0 MPa和0.22 GPa[29]。纤维的力学性能远高于薄壁细胞,纤维在毛竹的抗拉性能中起着决定性作用[29]。这可以归因于纤维和薄壁细胞在结构和化学成分等方面的不同,竹纤维的细胞壁几乎呈实心状,而薄壁细胞则具有较大的细胞腔和较薄的细胞壁[30],竹纤维和薄壁细胞在化学组成和晶体结构也有一定的差异[31]。因此,纤维鞘体积分数是影响竹材的顺纹抗压强度的重要因素之一。然而,竹子的维管束类型是多样的,包括双断腰型、断腰型、紧腰型、开放型和半开放型等5种维管束类型[32],不同竹种维管束的面积各异,即使同一类型维管束在不同位置或者不同竹种中的面积也不尽相同[33],因此,在研究不同种竹材的力学性能中探究维管束分布密度对力学性能的影响意义不大。综上所述,了解竹材结构的影响因素是非常必要的,这有助于掌握竹材的材料特性,充分发挥机械性能。
-
基于YOLO深度学习算法开发的竹材维管束模型应用于小径级竹材维管束数量和纤维鞘面积的测定,可以达到快速且准确的效果。小径级竹材的顺纹抗压性能不亚于大径级的竹材,早园竹顺纹抗压强度高达82.91 MPa,因此,小径级竹材具有较高的开发潜力。纤维鞘体积分数极显著地正向影响顺纹抗压强度。纤维鞘体积分数(x)与顺纹抗压强度(y)关系表述为一元一次方程:$ {y}\text{=260.44}{x}-\text{18.26} $。而维管束密度与顺纹抗压强度相关性较小。对于胸高直径50 mm以下的竹材,测试顺纹抗压强度时,建议采用纵向长度为平均外径2.0倍的试件,采用上述试验方法操作并控制试件在(90±30) s内压溃。另外,对于较小直径竹材制备试件时需避免对竹材造成损伤。
Compressive strength of small-diameter bamboo and its influencing factors
-
摘要:
目的 探究测试小径级圆竹顺纹抗压强度的方法,以及纤维鞘体积分数和维管束分布密度对顺纹抗压强度的影响。 方法 以苦竹Pleioblastus amarus、篌竹Phyllostachys nidularia、水竹Phyllostachys heteroclada和早园竹Phyllostachys propinqua 等4种小径级竹材(胸高直径50 mm以下)为研究对象,采用圆竹形式的试件探究小径级竹材的顺纹抗压性能测试方法,测试不同长径比(试件长度与直径的比值)试件对顺纹抗压强度的影响。同时,利用基于YOLO深度学习算法开发竹材维管束检测模型对维管束数量和纤维鞘面积进行测定,以探索其对顺纹抗压强度的影响。 结果 不同长径比试件测试结果并无显著性差异,以数据稳定性为准,则2.0倍长径比试件测试结果较为合理。采用该长径比试件测试,早园竹顺纹抗压强度最大,为82.91 MPa,水竹顺纹抗压强度最小,为67.01 MPa。篌竹纤维鞘组织比量最大,为35.64%,水竹最小,为33.05%。苦竹的维管束分布密度最大,达7.94 个·mm−2,早园竹最小,为5.77 个·mm−2。将不同种竹材作为整体对象的研究表明:纤维鞘体积分数正向影响顺纹抗压强度,而维管束分布密度对顺纹抗压强度的影响较小。 结论 小径级竹材顺纹抗压强度的测试宜采用2.0倍长径比试件,控制试件在(90±30) s内压溃。该试验选用的小径级竹材的顺纹抗压性能较为优异,纤维鞘体积分数(y)与顺纹抗压强度(x)之间存在$ {y}\text{=260.44}{x}{-}\text{18.26} $线性相关性。图4表2参33 Abstract:Objective This study is aimed to investigate the testing method for determining the compressive strength of small-diameter bamboo culms as well as the impact of fiber sheath volume fraction and distribution density of vascular bundles on it. Method Four species of small-diameter bamboo, namely Pleioblastus amarus, Phyllostachys nidularia, Phyllostachys heteroclada, and Phyllostachys propinqua, with a diameter at breast height of less than 50 mm, were selected as research subjects before bamboo culm samples were utilized to investigate the compressive strength testing method for small-diameter bamboo and to examine the impact of varying length-to-diameter ratios on compressive strength. At the same time, a bamboo vascular bundle detection model based on the YOLO deep learning algorithm was employed to determine the number of vascular bundles and fiber sheath area so as to investigate their influence on compressive strength. Result There were no significant differences in the test results among specimens with different length-to-diameter ratios and the test results for the specimen with a length-to-diameter ratio of 2.0 were more reasonable. Of specimens at the length-to-diameter ratio of 2.0, Phyllostachys propinqua exhibited the highest compressive strength at 82.91 MPa while Phyllostachys heteroclada demonstrated the lowest strength at 67.01 MPa. The volume fraction of fiber sheath was highest in Phyllostachys nidularia at 35.64% and lowest in Phyllostachys heteroclada at 33.05%. The density of vascular bundles in Pleioblastus amarus was highest at 7.94 pieces·mm−2, while that of Phyllostachys propinqua was the lowest at 5.77 pieces·mm−2. Studies that treated various species of bamboo as a unified entity have shown that the positive effect of the volume fraction of fiber sheath on compressive strength was significant while the influence of vascular bundle distribution density on compressive strength was relatively minor. Conclusion A specimen with a length-to-diameter ratio of 2.0 is an ideal choice for testing the compressive strength of small-diameter bamboo and the specimen should be controlled for a period of (90±30) seconds before it collapses. Furthermore, the small-diameter bamboo selected for this experiment exhibited excellent compressive performance while there was a direct correlation between the volume fraction of fiber sheath (y) and compressive strength (x), as shown in the equation y=260.44x−18.26. [Ch, 4 fig. 2 tab. 33 ref.] -
图 2 高清灰度图(A)、识别每个维管束的检测图(B)和利用局部聚类算法获得的二值化图(C)
Figure 2 HD grayscale image (A), detection image identifying each vascular bundle (B) and binarized image obtained using local clustering algorithm (C)
图 4 顺纹抗压强度与纤维鞘体积分数和维管束分布密度之间的分布规律
Figure 4 Distribution pattern between the compressive strength and the fiber volume fraction and distribution density of vascular bundles
表 1 顺纹抗压测试结果
Table 1. Test results of compressive strength
竹种 长径比 顺纹抗压强度 方差齐性
检验P单因素
ANOVA检验平均值/
MPa标准差/
MPa变异系
数/%F P 苦竹 1.0 71.64 ab 3.80 5.30 0.456 3.342 0.040 1.5 70.27 b 3.20 4.55 2.0 72.65 a 3.89 5.35 篌竹 1.0 74.57 a 8.02 10.76 0.989 0.239 0.788 1.5 74.01 a 7.68 10.38 2.0 73.19 a 7.59 10.37 水竹 1.0 67.09 a 5.75 8.57 0.776 0.222 0.801 1.5 67.78 a 6.01 8.87 2.0 67.01 a 5.12 7.64 早园竹 1.0 83.29 a 9.52 11.43 0.528 0.010 0.990 1.5 83.06 a 8.19 9.86 2.0 82.91 a 7.76 9.36 说明:不同小写字母表示同一竹种不同处理之间LSD检验差异显著(P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 4种小径级竹材长径比为2.0试样的维管束参数
Table 2. Vascular bundle parameters of four species of small diameter bamboo at the length-to-diameter ratio of 2.0
竹种 纤维鞘体积分数/% 维管束分布密度/(个·mm−2) 平均值 标准偏差 平均值 标准偏差 苦竹 34.61 2.18 7.94 0.73 篌竹 35.64 1.51 6.32 0.92 水竹 33.05 2.15 7.01 0.31 早园竹 34.65 5.53 5.77 1.07
下载: 导出CSV
-
[1] KAUR P J. Bamboo availability and utilization potential as a building material [J]. Forestry Research and Engineering: International Journal, 2018, 2(5): 240 − 242. [2] YUAN Tiancheng, WANG Xinzhou, LIU Xiaorong, et al. Bamboo flattening technology ebables efficient and value-added utilization of bamboo in the manufacture of furniture and engineered composites [J/OL]. Composites Part B: Engineering, 2022, 242: 110097[2023-02-28]. doi:10.1016/j.compositesb.2022.110097. [3] SHAMSURI M A, MAIN N M. Review on the paper making process from bamboo as a paper product [J]. Progress in Engineering Application and Technology, 2021, 2(1): 965 − 971. [4] 李佳, 顾蕾, 朱玮强, 等. 浙江省安吉县CCER竹林经营碳汇交易项目经济效益分析[J]. 浙江农林大学学报, 2018, 35(4): 581 − 588. LI Jia, GU Lei, ZHU Weiqiang, et al. Economic benefit of carbon sequestration project of CCER bamboo forest management in Anji Country, Zhejiang Province [J]. Journal of Zhejiang A&F University, 2018, 35(4): 581 − 588. [5] DAI Fukuan, WANG Ziwei, WANG Hankun, et al. Vascular bundle characteristics and mechanical properties of Dendrocalamus sinicus [J/OL]. Construction and Building Materials, 2023, 363: 129858[2023-02-28]. doi: 10.1016/j. conbuildmat. 2022.129858. [6] MA Xinxin, LUO Zhiqiang, JI Conghui, et al. Flexural creep behaviors of bamboo subjected to different gradient variation directions and relative humidity [J/OL]. Industrial Crops and Products, 2022, 179: 114679[2023-02-28]. doi: 10.1016/j.indcrop.2022.114679. [7] YU Yanglun, HUANG Yuxiang, ZHANG Yahui, et al. The reinforcing mechanism of mechanical properties of bamboo fiber bundle-reinforced composites [J]. Polymer Composites, 2019, 40(4): 1463 − 1472. [8] LI Hongbo, SHEN Shengping. The mechanical properties of bamboo and vascular bundles [J]. Journal of Materials Research, 2011, 26(21): 2749 − 2756. [9] LIU Yanyan, HUANG Dongsheng, ZHU Junjie. Experimental investigation of mixed-mode I/II fracture behavior of parallel strand bamboo [J/OL]. Construction and Building Materials, 2021, 288: 123127[2023-02-28]. doi: 10.1016/j.conbuildmat.2021.123127. [10] ZHAO Xiaoxiao, YAN Jing, HUANG Ying, et al. Magnetic porous CoNi@C derived from bamboo fiber combined with metal-organic-framework for enhanced electromagnetic wave absorption [J]. Journal of Colloid and Interface Science, 2021, 595: 78 − 87. [11] CHEN Meiling, WENG Yun, SEMPLE K, et al. Sustainability and innovation of bamboo winding composite pipe products [J/OL]. Renewable and Sustainable Energy Reviews, 2021, 144: 110976[2023-02-28]. doi: 10.1016/j.rser.2021.110976. [12] 张亚慧, 黄宇翔, 于文吉, 等. 我国竹产业的发展历程、现状及趋势[J]. 中国人造板, 2019, 26(6): 32 − 36. ZHANG Yahui, HUANG Yuxiang, YU Wenji, et al. Development, current situation and trend of bamboo industry in China [J]. China Wood-Based Panels, 2019, 26(6): 32 − 36. [13] 林敏, 龚媛媛, 石金明, 等. 江西的竹产业发展现状与建议[J]. 能源研究与管理, 2019(3): 1 − 5. LIN Min, GONG Yuanyuan, SHI Jinming, et al. Development status and suggestion of bamboo industry in Jiangxi [J]. Energy Research and Management, 2019(3): 1 − 5. [14] CHEN Lin, YU Zixuan, FEI Benhua, et al. Study on performance and structural design of bamboo helmet [J/OL]. Forests, 2022, 13(7): 1091[2023-02-28]. doi: 10.3390/f13071091. [15] 夏雨, 牛帅红, 李延军, 等. 常压高温热处理对红竹竹材物理力学性能的影响[J]. 浙江农林大学学报, 2018, 35(4): 765 − 770. XIA Yu, NIU Shuaihong, LI Yanjun, et al. Physical and mechanical properties of Phyllostachys iridescins under normal pressure and heat temperature [J]. Journal of Zhejiang A&F University, 2018, 35(4): 765 − 770. [16] BAHTIAR E T, IMANULLAH A P, HERMAWAN D, et al. Structural grading of three sympodial bamboo culms (Hitam, Andong, and Tali) subjected to axial compressive load [J]. Engineering Structures, 2019, 181: 233 − 245. [17] 陈冠军, 袁晶, 余雁, 等. 竹材顺纹抗压性能的种间差异及其影响因子研究[J]. 木材加工机械, 2018, 29(6): 23 − 27. CHEN Guanjun, YUAN Jing, YU Yan, et al. Study on interspecific differences of the compression performance parallel to the grain of bamboo and its influencing factors [J]. Forestry and Grassland Machinery, 2018, 29(6): 23 − 27. [18] 中国木材标准化技术委员会. 竹材物理力学性质试验方法: GB/T 15780—1995[S]. 北京: 中国标准出版社, 1995. National Technical Committee on Timber of Standardization Administrator of China. Testing Methods for Physical and Mechanical Properties of Bamboos: GB/T 15780−1995 [S]. Beijing: Standards Press of China, 1995. [19] 蔡绍祥, 夏雨, 徐冰洁, 等. 小径级圆竹家具用材力学性能及其设计改良研究[J]. 林产工业, 2020, 57(9): 32 − 36. CAI Shaoxiang, XIA Yu, XU Bingjie, et al. Study on mechanical properties and furniture design improvement of small diameter round bamboo [J]. China Forest Products Industry, 2020, 57(9): 32 − 36. [20] 代福宽, 潘怀志, 王传贵. 小径级竹材抗弯性能测试方法[J]. 世界竹藤通讯, 2020, 18(3): 45 − 49, 54. DAI Fukuan, PAN Huaizhi, WANG Chuangui. Test method of bending properties for small diameter bamboo [J]. World Bamboo and Rattan, 2020, 18(3): 45 − 49, 54. [21] LI Jing, XU Haocheng, YU Yan, et al. Intelligent analysis technology of bamboo structure. PartⅠ: The variability of vascular bundles and fiber sheath area [J/OL]. Industrial Crops and Products, 2021, 174: 114163[2023-02-28]. doi: 10.1016/j.indcrop.2021.114163. [22] 黎静, 黄汉霄, 石俊利, 等. 竹环中维管束分布密度和纤维鞘组织比量纵向变异研究[J]. 世界竹藤通讯, 2019, 17(2): 7 − 11. LI Jing, HUANG Hanxiao, SHI Junli, et al. A study of longitudinal variation of vascular bundles distribution density and fiber sheath tissue proportion in internode of moso bamboo [J]. World Bamboo and Rattan, 2019, 17(2): 7 − 11. [23] 张文福. 圆竹性能评价及其帚化加工技术的研究[D]. 北京: 中国林业科学研究院, 2012. ZHANG Wenfu. Mechanical Properties and Brooming Processing of Bamboo-culm [D]. Beijing: Chinese academy of forestry, 2012. [24] AKINBADE Y, HARRIES K A. Is the rule of mixture appropriate for assessing bamboo material properties? [J/OL]. Construction and Building Materials, 2021, 267: 120955[2023-02-28]. doi: 10.1016/j.conbuildmat.2020.120955. [25] 李荣荣, 贺楚君, 彭博, 等. 毛竹材不同部位纤维形态及部分物理性能差异[J]. 浙江农林大学学报, 2021, 38(4): 854 − 860. LI Rongrong, HE Chujun, PENG Bo, et al. Differences in fiber morphology and partial physical properties in different parts of Phyllostachys edulis [J]. Journal of Zhejiang A&F University, 2021, 38(4): 854 − 860. [26] LO T Y, CUI Hongzhi, LEUNG H C. The effect of fiber density on strength capacity of bamboo [J]. Materials Letters, 2004, 58(21): 2595 − 2598. [27] ZHANG Xuexia, LI Jinghao, YU Zixuan, et al. Compressive failure mechanism and buckling analysis of the graded hierarchical bamboo structure [J]. Journal of Materials Science, 2017, 52: 6999 − 7007. [28] HABIBI M K, SAMAEI A T, GHESHLAGHI B, et al. Asymmetric flexural behavior from bamboo’s functionally graded hierarchical structure: underlying mechanisms [J]. Acta Biomaterialia, 2015, 16: 178 − 186. [29] SHAO Zhuoping, FANG Changhua, HUANG Shengxia, et al. Tensile properties of Moso bamboo (Phyllostachys pubescens) and its components with respect to its fiber-reinforced composite structure [J]. Wood Science and Technology, 2010, 44: 655 − 666. [30] WANG Hankun, ZHANG Xuexia, JIANG Zehui, et al. A comparison study on the preparation of nanocellulose fibrils from fibers and parenchymal cells in bamboo (Phyllostachys pubescens) [J]. Industrial Crops and Products, 2015, 71: 80 − 88. [31] REN Wenting, ZHU Jiawei, GUO Fei, et al. Structural evolution of cellulose from bamboo fibers and parenchyma cells during ionic liquid pretreatment for enhanced hydrolysis [J]. Biomacromolecules, 2022, 23(5): 1938 − 1948. [32] DARWIS A, ISWANTO A H. Morphological characteristics of Bambusa vulgaris and the distribution and shape of vascular bundles therein [J]. Journal of the Korean Wood Science and Technology, 2018, 46(4): 315 − 322. [33] XU Haocheng , LI Jing, MA Xingxing , et al. Intelligent analysis technology of bamboo structure. Part Ⅱ: The variability of radial distribution of fiber volume fraction [J/OL]. Industrial Crops and Products, 2021, 174: 114164[2023-02-28]. doi: 10.1016/j.indcrop.2021.114164. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230207
 点击查看大图
点击查看大图

计量
- 文章访问数: 1875
- HTML全文浏览量: 559
- PDF下载量: 40
- 被引次数: 0
