




-
种子是植物天然更新的主要途径。在特定的时间和空间范围内从母树上散落的种子被称为种子雨,它是森林土壤种子库的重要组成部分[1−2]。树种能否更新取决于该树种能否从种子成长为成树,其中种子的传播则是构建植物群落的第一步[3]。扩散方式[4]、树高[5]、种实大小[6]及种实产量[7]等母树自身因素,气候[8]、动物和昆虫[9]、地形[10]等环境因素均会影响种子的扩散与传播,这些因素最终会对整个种群的结构、发展方向及树木的空间分布产生重要影响[11−13]。另外,种子雨具有明显的时间动态。冯倩倩等[14]指出白桦Betula platyphylla种子落种经历了起始期、高峰期和末尾期3个散种阶段。窦丽娜等[15]通过连续9 a对西双版纳望天树Parashorea chinensis林的种子雨动态监测,发现该树种存在明显的年际变化。种子是森林更新的基础,它们的空间分布模式是由其大树的空间分布格局、种子产量和种子域共同决定的,并且对森林的持续发展至关重要[16−17]。当散种量较大时,其种子呈现聚集分布模式,当散种量较小时,其种子呈现随机或均匀分布模式。因此,深入研究种子的时空分布特征以及它们的萌发行为,有助于更好地理解森林的变化规律。
天山云杉Picea schrenkiana var. tianschanica是新疆特有的植物,它们在新疆的分布范围极广,储量丰富,不仅是新疆山区森林生态系统的核心,而且还可以为新疆的绿洲灌溉农业、生态安全提供强有力的支撑[18−19]。本研究以新疆天山云杉纯林为对象,调查其种子雨的时间动态、空间分布格局和种子萌发特征等,旨在了解天山云杉林的自然更新规律,预测森林群落的演替趋势,为保护天山森林生态系统的物种多样性提供科学依据。
-
研究区位于新疆农业大学实习林场(43°16′~43°26′N, 86°46′~86°57′E),海拔为1 700~3 200 m, 林区总面积为15.07万 hm2,该区地处天山北麓的头屯河上游,地形南高北低,地形切割较为剧烈,坡度为10°~40°。年降水量为500~600 mm,雨季(5—8月)降雨量约占年降水量的60%;年均气温为3 ℃,1月平均气温为−10 ℃,7月平均气温为14 ℃,年日照时长>1 300 h,属于温带大陆性气候[20]。区内森林主要分布在北、东北和西北坡,森林覆盖率为56.64%,是以天山云杉为建群种的纯林,另外还有天山花楸Sorbus tianschanica、天山桦Betula tianschanica等树种。
-
2020年在研究区设置3块面积为1 hm2(100 m×100 m)的天然林永久固定监测样地。3个监测样地水源充足,林木资源丰富,处于成熟林阶段,地势较平缓,海拔为2 100~2 300 m。样地基本概况:样地1,坡度8°,海拔2 256.2 m,平均树高19.25 m,平均胸径24.05 cm;样地2,坡度12°,海拔2 278.8 m,平均树高18.70 m,平均胸径23.13 cm;样地3,坡度11°,海拔2 161.8 m,平均树高24.15 m,平均胸径28.59 cm。采用机械布局的方法在3块样地中设置种子雨收集器,以10 m×10 m的样方为单位,在样方中心设置1个种子雨收集器,每个样地共100个收集器(图1)。种子收集器面积为1 m2,网目为1 mm的尼龙网。分别于2021、2022年9月至次年2月,每隔7 d收集1次种子雨。
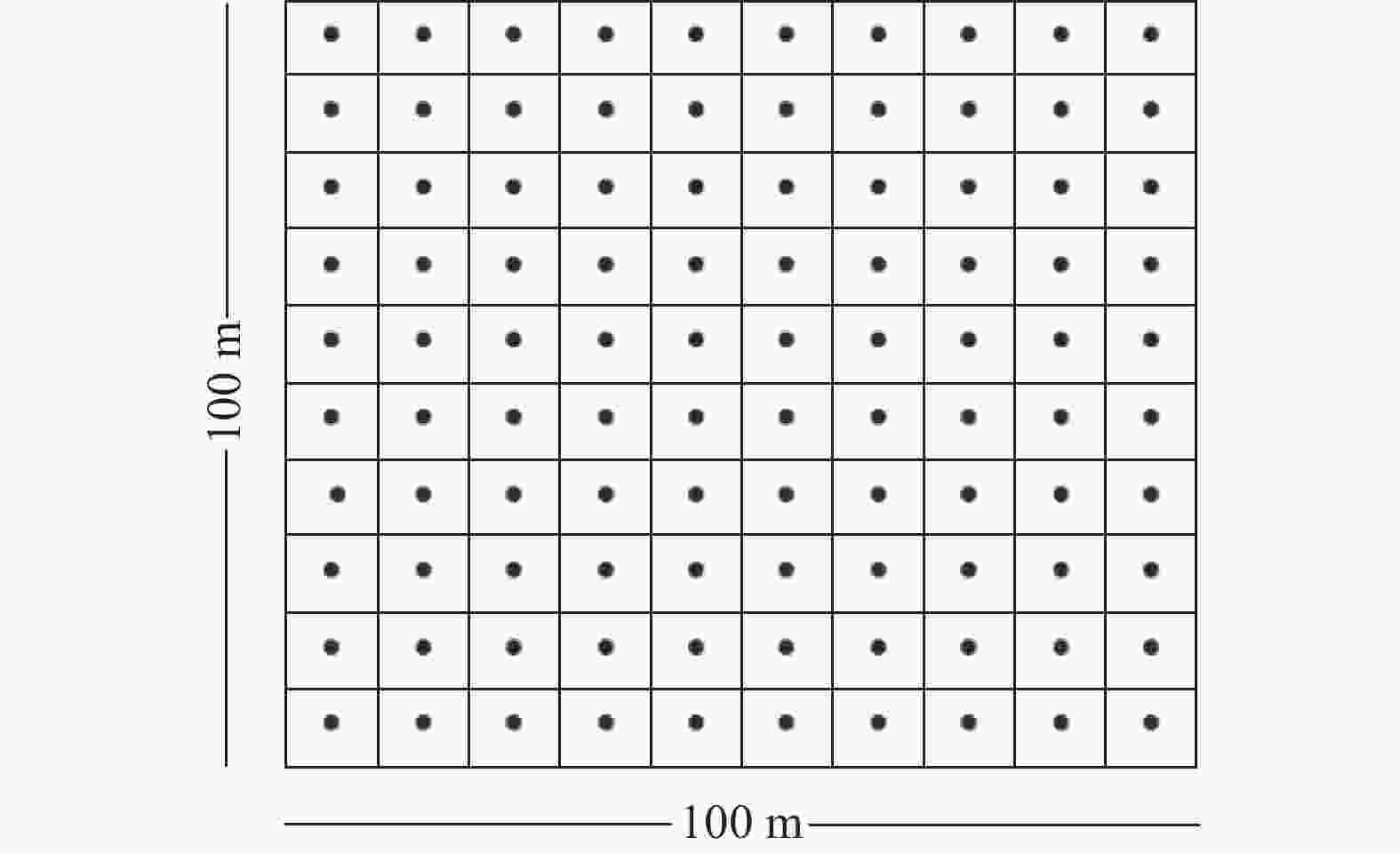
图 1 种子雨收集器
Figure 1. Location of seed collector
-
对收集的种子样品进行分拣、鉴定、计数和记录。将分拣出来的种子分为完整种子和虫蛀种子。对天山云杉种子采用四分法计数, 将挑选出来的种子样品放置在玻璃板或光滑的桌面上,用分样板先将种子横向混合,之后纵向混合,如此混合5次,使种子混合均匀。然后摊平,种子厚度不超过1 cm,沿对角线将样品分成4个三角形,再取2个对顶三角形内的样品继续按以上方法分取。随后分别提取测定样品8份,从中随机选取100粒为1组,共8个重复。分别计算种子雨密度[14]和种子千粒重。$U = {N}/V $。其中:U为种子雨密度,N为收集器中收集到的种子数量,V为种子收集器的面积。
-
根据GB 2772—1999《林木种子检验规程》规定,将每个收集器收集的天山云杉种子分别置于培养皿中,放置在(30±2) ℃的培养箱内进行发芽试验。从放置种子的第2 天起开始记录发芽数目,连续21 d 无种子萌发时停止发芽,分别计算种子发芽势和发芽率[21]。$ {P_0} = ({N_0}/N) \times 100\% $。其中:P0为种子发芽率;N0为发芽种子数;N为种子总数。$ P_{1}=\left(N_{1} / N\right) \times 100 \% $。其中:P1为种子发芽势,N1为发芽高峰期发芽总数。
-
种子雨的空间分布类型分为聚集分布、随机分布和均匀分布[22],对收集到的种子进行种子密度分析,并采用方差/均值(Cx)、负二项指数(K)、聚集度指标(I)、Cassie R M指标(CA)4 种空间分布类型评价方法判别天山云杉种子雨空间分布格局[23],Cx<1为均匀分布,Cx=1为随机分布,Cx>1为聚集分布;K<0为非聚集分布,K>0为聚集分布;I<0为均匀分布,I=0为随机分布,I>0为聚集分布;CA<0为均匀分布,CA=0为随机分布,CA>0为聚集分布。
将所测定的数据输入到Excel,利用SPSS 19.0对数据进行非参数检验(Kruskal-Wallis检验法)和差异显著性检验,使用Origin 2022制图。
-
2021年9月至2022年2月,收集2021年天山云杉种子,3个监测样地共收集到91粒种子,且所有种子均未萌发。2022年9月至2023年2月下旬收集2022年天山云杉种子,共收集到139 257粒种子。天山云杉2021年的种子雨密度为0.46 粒·m−2,2022年的种子雨密度为696.29 粒·m−2,说明天山云杉种子的散种量存在明显的“大小年”现象。
-
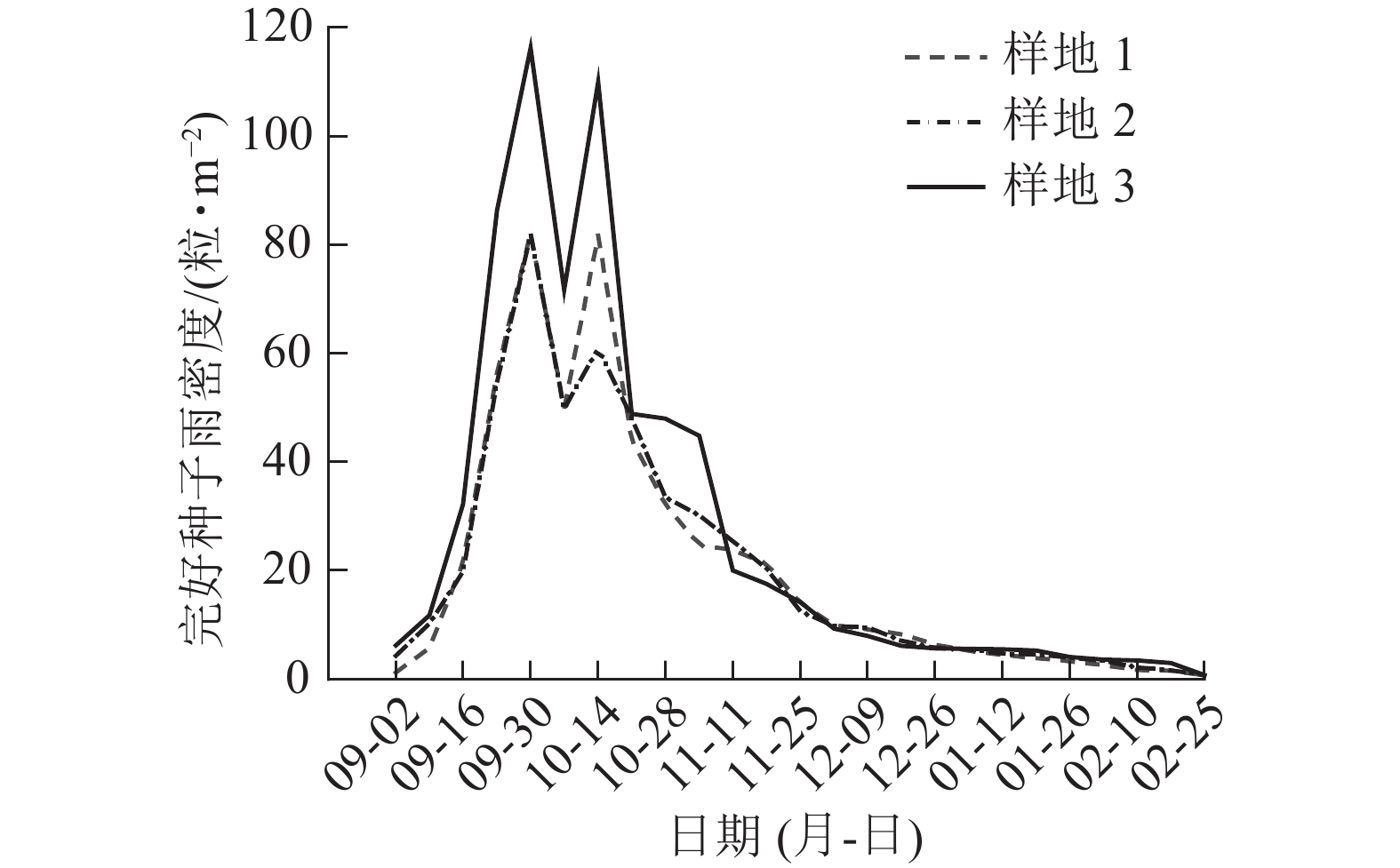
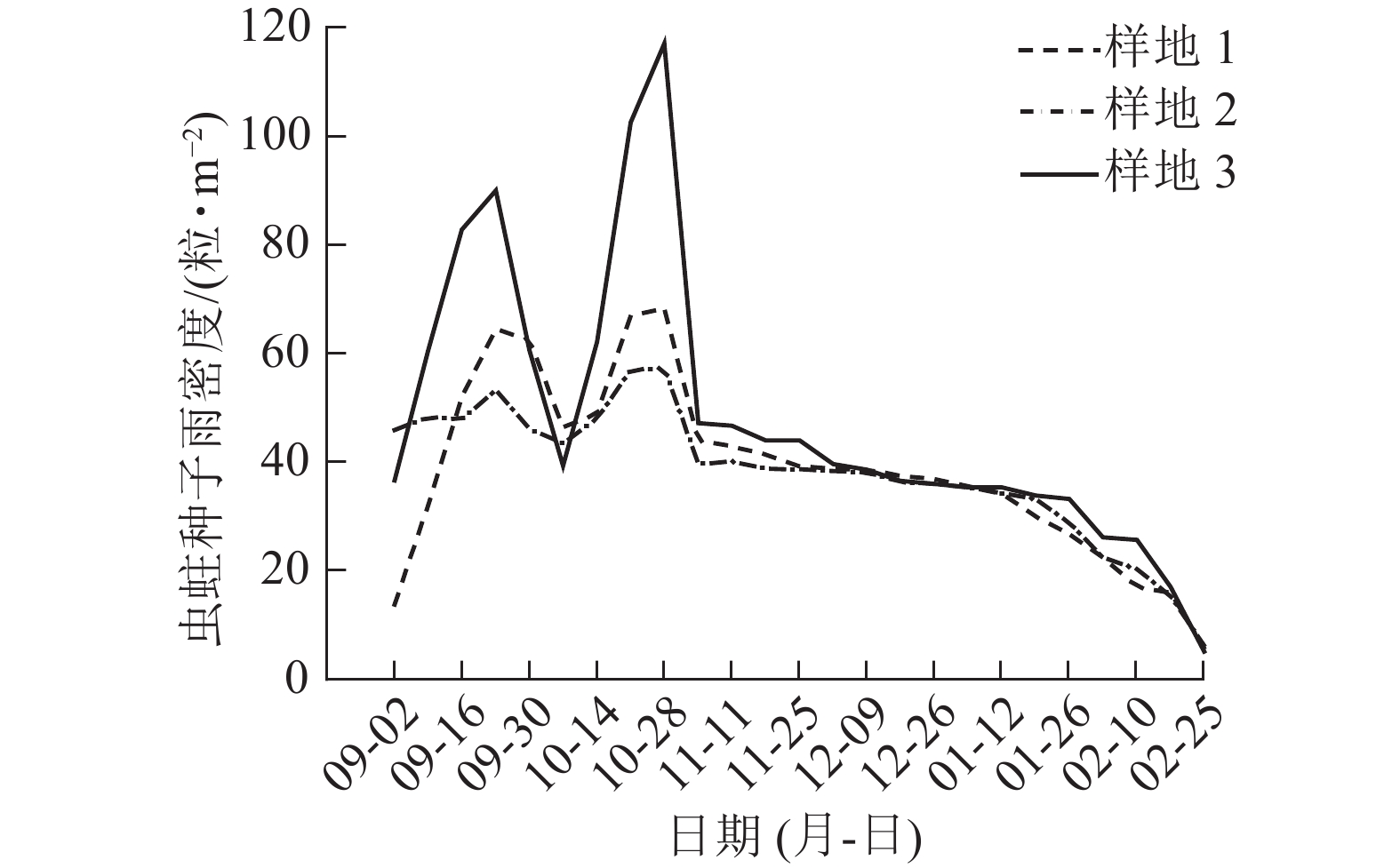
因2021年收集的天山云杉种子过少,仅对2022年种子雨进行时间动态分析。2022年天山云杉种子雨随着时间的变化呈现规律性变化(图2):9月2日开始有种子掉落,9月24日至10月21日种子雨散种量急剧上升,绝大多数种子均在这个时间内散落;10月21日后至次年2月,种子雨散种量开始下降并趋于平稳。虫蛀种子数量为86.00~108.97粒·m−2 (图3),在9月24日和10月21日虫蛀种子数量到达峰值。依据种子散种量可知:9月2—16日为种子散种起始期,9月24日—10月21日为散种高峰期,10月21日后种子散种量进入末尾期。

图 2 各样地完好种子雨密度的时间动态
Figure 2. Temporal dynamics of intact seed rain density in 3 plots

图 3 各样地虫蛀种子雨密度的时间动态
Figure 3. Time dynamics of insect-eaten seed rain density in 3 plots
-
使用4种不同的空间分布类型评价方法对种子雨的空间分布特征进行了分析(表1):Cx、K、I和CA的平均值分别达到了2 047.01、1.02、2 046.01以及0.99。所有样地4个指标值均远大于临界值,表明该林区天山云杉种子扩散时聚集程度很高。各样地CA从大到小依次为样地3、样地1、样地2。
表 1 2022年9月—2023年2月各监测样地下种子雨的空间分布格局
Table 1. Spatial distribution pattern of underground seed rain in the monitoring samples from September of 2022 to January of 2023
监测样地 Cx K I CA 排序 数值 分布格局 数值 分布格局 数值 分布格局 数值 分布格局 1 1 232.50 A 1.05 A 1 231.50 A 0.95 A 2 2 1 055.20 A 1.16 A 1 054.20 A 0.86 A 3 3 3 853.33 A 0.85 A 3 852.33 A 1.17 A 1 均值 2 047.01 1.02 2 046.01 0.99 说明:A. 聚散分布。 -
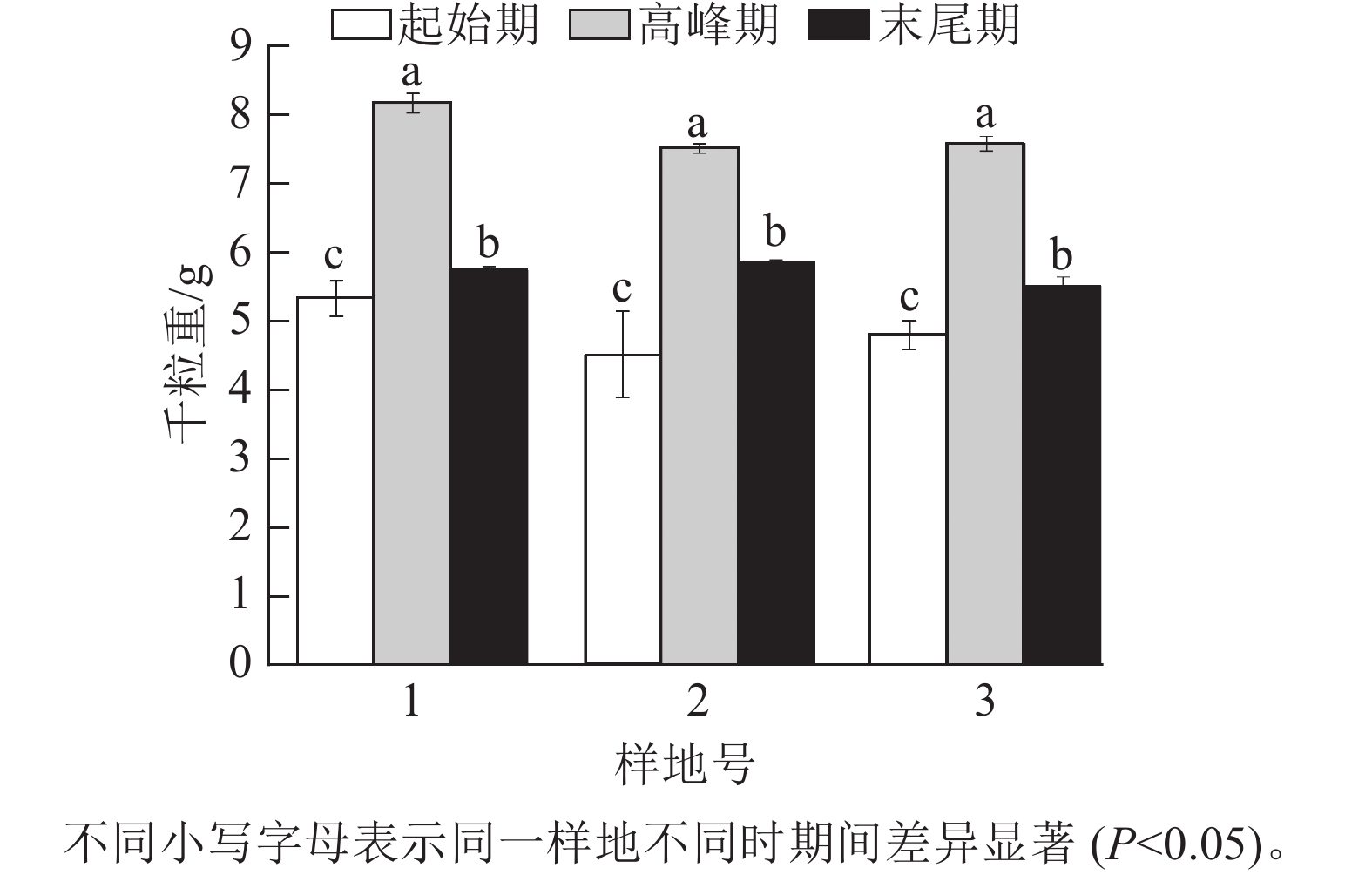
由图4可以看出:天山云杉的平均种子雨千粒重为(6.116±0.182) g。不同时期,3个样地高峰期种子的千粒重都显著高于初始期和末尾期,高峰期的种子千粒重分别为(8.173±0.141)、(7.519±0.067)和(7.588±0.109) g。3个样地的平均天山云杉种子千粒重从大到小依次为样地1、样地3、样地2,平均种子千粒重分别为(6.417±0.151)、(5.967±0.146)和(5.963±0.248) g。

图 4 各样地不同时期种子的千粒重
Figure 4. 1 000-grain seed mass of seeds in different times
-
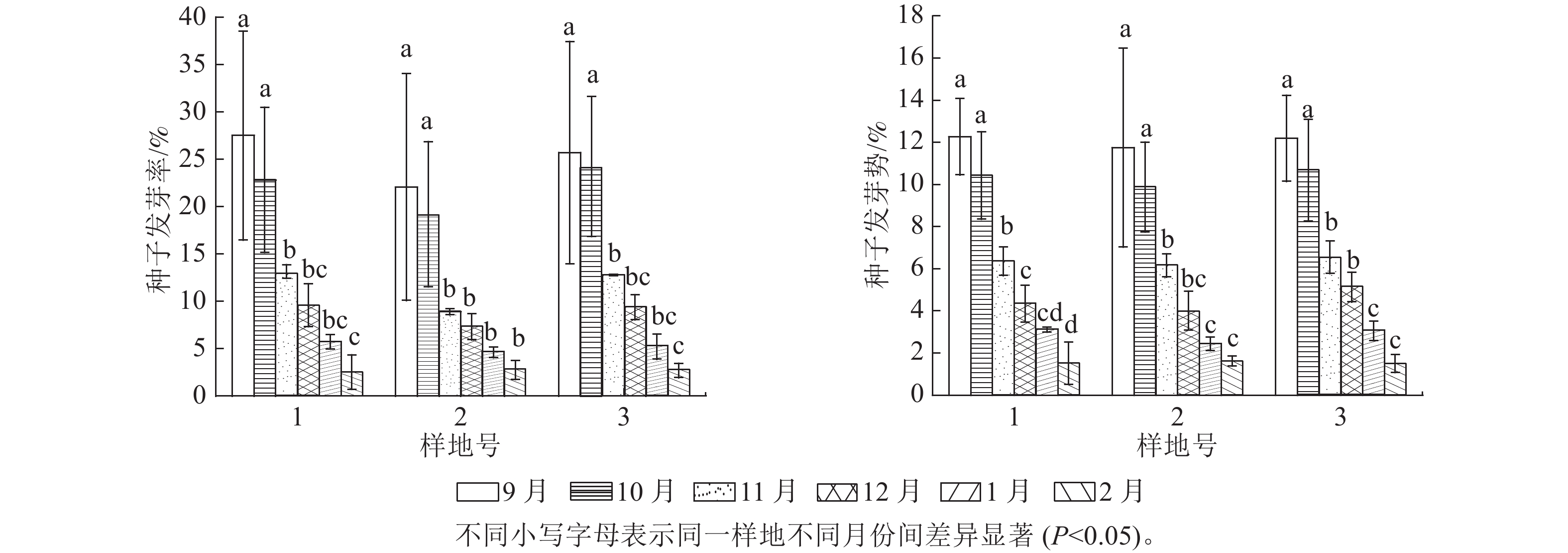
9、10和11月收集到种子第4天开始萌发,12、1和2月收集到种子萌发有所变化,第6天才开始萌发。由图5可见:不同时间收集的天山云杉种子发芽率差异显著(P<0.05),其中9月和10月收集的种子平均发芽率差异不显著,分别为(25.08±11.57)%和(22.08±7.57)%,但二者均显著高于其他月份收集的种子平均发芽率,次年2月种子平均发芽率最低,为(2.66±1.19)%。不同的样地发芽率从大到小依次为样地1、样地3、样地2。9、10和11月收集到的种子发芽势在第8天达最高,12、1和2月收集到的种子发芽势,在第10天才达最高。不同时间收集的天山云杉种子发芽势差异显著(P<0.05),其中,9月和10月收集的种子平均发芽势差异不显著,分别为(12.07±2.86)%和(10.33±2.20)%,但显著高于其他月份收集的种子平均发芽势,次年2月平均发芽势最低,为(1.55±0.55)%。不同的样地的发芽势从大到小依次为样地1、样地3、样地2。

图 5 各样地种子发芽率和发芽势
Figure 5. Seed germination rate and potential in 3 plots
-
种子是森林更新的起点,它们的数量和生长潜力对森林恢复至关重要[17]。天山云杉2021年的种子雨密度为0.46 粒·m−2,2022年的种子雨密度为696.29 粒·m−2,表明天山云杉种子的散种量存在较大的年际变化,存在“大小年”,但明确其年际的周期性变化仍需继续观测[24]。时间动态是种子雨的主要特征之一[22],天山云杉的种子雨整个过程经历了起始期、高峰期和末尾期3个阶段,即9月2—24日为种子散种起始期、9月24日至10月21日为散种高峰期,10月21日后种子散种进入末尾期。这与冯倩倩等[14]研究的白桦种子落种时期结果相似。出现双峰及峰谷是由9月底降雨引起的。本研究发现:天山云杉从10月下旬到次年2月仍然有种子下落,并且趋于平稳,这与油松Pinus tabuliformis[25]种子下落持续时间一致,说明后期天山云杉种子雨主要为林冠截获的种子。本研究中,虫蛀种子数量为 86.09~108.97粒·m−2。天山云杉林以纯林为主,树种单一,病虫害发生率高,因此在天山云杉林生态恢复时,应营造混交林。
-
种子雨的空间分布可以揭示种群对环境的适应能力[16],种子雨的散种量对物种的空间分布格局存在影响。在对赤皮青冈Cyclobalanopsis gilva[26]的研究中发现:当散种量较大时,种子呈现聚集分布模式,当散种量较小时,种子呈现随机或均匀分布模式。枫桦Betula costata[27]种子分布格局表现为高度聚集,且这种高强度聚集分布与林分郁闭度和林分密度有着密切的联系,种子无法进行远距离传播。本研究运用4种方法对2022年天山云杉的种子雨进行了空间分布特征分析,发现天山云杉种子的空间分布特征均呈现聚集分布,说明该树种的空间格局与种子产量、林分密度等有关。
-
种子质量变化和幼苗更新是森林演替的重要过程[4]。本研究结果表明:天山云杉种子的千粒重与其分布规律相吻合,其平均种子雨千粒重为(6.116±0.182) g,且高峰期的天山云杉种子千粒重显著高于初始期和末尾期的天山云杉种子千粒重。这与落叶松Larix gmelinii种子千粒重[3]情况相似。
发芽率与发芽势是反映种子质量优劣的主要指标之一,杨彬等[28]对木麻黄Casuarina equisetifolia种子进行了8个月发芽试验,发现种子雨的萌发率不仅取决于种子本身的萌发能力,还与种子的抗老化能力和种子雨收集间隔期长短等有关。程福山等[29]研究发现:枫桦10月种子雨达到峰值,但9月种子发芽率显著高于其他月份种子发芽率,这可能与种子储存蛋白质水平、植物的生长状况以及气候因素等有关。在本研究中,不同月份之间种子萌发能力不同,9月和10月收集的种子发芽势与发芽率最高,且显著高于其他月份收集的种子平均发芽势和平均发芽率,并且9月各样地种子发芽率和发芽势排列顺序为:样地1>样地3>样地2。可见,云杉种子萌发能力不同,可能与种子收集时期有关。
-
天山云杉种子的散种量存在较大的年际变化,且具有明显的时间分布特征,9月下旬至10月中旬为种子雨散种高峰期;另外,高峰期的天山云杉种子千粒重显著高于初始期和末尾期的天山云杉种子千粒重;而且9月和10月收集的种子的萌发率最高。3个样地中,3号样地种子雨散种最多,质量最佳,萌发最高。
Spatiotemporal dynamic distribution of seed rain and seed germination characteristics of Picea schrenkiana var. tianschanica in Xinjiang
-
摘要:
目的 研究新疆天山云杉Picea schrenkiana var. tianschanica 种子雨时空动态分布规律和种子萌发状况,为预测森林群落的演替趋势,并为保护天山森林生态系统的物种多样性提供科学依据。 方法 在林场设置3块面积为1 hm2天然林永久固定监测样地,采用机械布局的方法以10 m×10 m的样方为单位,在样方中心设置1个种子雨收集器,每个样地共100个收集器。采用方差/均值(Cx)、负二项指数(K)、聚集度指标(I)、Cassie R M指标(CA)等4种空间分布类型评价方法判别种子雨空间分布状态。此外,进行种子萌发试验,分别测定种子千粒重、发芽势和发芽率。 结果 9月下旬至10月中旬散种量较高;2022年的种子雨总量极显著高于2021年的种子雨总量;Cx为2 047.01、K为1.02、I为2 046.01、CA为0.99,4个指标均显示种子空间分布呈聚集分布;各样地高峰期的天山云杉种子千粒重显著高于初始期和末尾期的天山云杉种子千粒重;而种子雨初始期和高峰期的种子平均发芽势与平均发芽率差异不显著,但都显著高于末尾期的种子的平均发芽势和平均发芽率。 结论 天山云杉种子的散种量存在较大的年际变化,且种子空间分布呈聚集分布,落种高峰期为9月下旬至10月中旬,此时种子质量最大,平均发芽率和发芽势最高。图5表1参29 Abstract:Objective In order to provide scientific basis for predicting the succession trend of forest community and protecting the species diversity of ecosystem in Tianshan, this study focus on the spatiotemporal distribution of seed rain and seed germination of Picea schrenkiana var. tianschabica (Tianshan spruce) in Tianshan, Xinjiang. Method Based on three sample plots, 1 hm2 each set in the Xinjiang agricultural university experimental forest farm, and a seed rain collector was set in the center of each 10 m×10 m sample plot by mechanical layout method, with a total of 100 collectors in each sample plot. Using variance/mean (Cx), negative binomial index (K), aggregation index David (I), and Cassie R M, index (CA), 4 evaluation methods to distinguish the spatial distribution state of seed rain. In addition, the seed germination experiment 1 000-grain seed mass, germination energy and germination rate were measured respectively. Result The peak period of seed falling of Picea schrenkiana var. tianschanica was from late September to mid-October. The total amount of seed rain in 2022 was significantly higher than that in 2021. The seeds of Tianshan spruce presented aggregation distribution at the spatial level based on 4 indexes, Cx reaching 2047.01, K reaching 1.02, I reaching 2 046.01, and CA reaching 0.99 respectively. The 1 000-grain mass of spruce seed in the peak period was significantly higher than in the initial stage and the end stage. There was no significant in seed gernination energy and gernination rates between the initial period and period, but were significantly higher than the ending time of the rain. [ Conclusio n ] The interannual variation characteristics of seed rain of Tianshan spruce obviously showed an aggregated distribution. The peak period of seed rain was from late September to mid-October, the 1 000-grain mass, the germination energy and average germination rate were the best during this period. [Ch, 5 fig. 1 tab. 29 ref.] -
Key words:
- Picea schrenkiana var. tianschanica /
- seed rain /
- spatial distribution /
- seed germination
-
图 3 各样地虫蛀种子雨密度的时间动态
Figure 3 Time dynamics of insect-eaten seed rain density in 3 plots
表 1 2022年9月—2023年2月各监测样地下种子雨的空间分布格局
Table 1. Spatial distribution pattern of underground seed rain in the monitoring samples from September of 2022 to January of 2023
监测样地 Cx K I CA 排序 数值 分布格局 数值 分布格局 数值 分布格局 数值 分布格局 1 1 232.50 A 1.05 A 1 231.50 A 0.95 A 2 2 1 055.20 A 1.16 A 1 054.20 A 0.86 A 3 3 3 853.33 A 0.85 A 3 852.33 A 1.17 A 1 均值 2 047.01 1.02 2 046.01 0.99 说明:A. 聚散分布。  下载: 导出CSV
下载: 导出CSV
-
[1] 黄雍容, 马祥庆, 庄凯, 等. 福建闽清福建青冈天然林种子雨和种子库[J]. 热带亚热带植物学报, 2010, 18(1): 68 − 74. HUANG Yongrong, MA Xiangqing, ZHUANG Kai, et al. Seed rain and seed bank of Cyclobalanopsis chungii forest in Minqing, Fujian Province [J]. Journal of Tropical and Subtropical Botany, 2010, 18(1): 68 − 74. [2] 于顺利, 郎南军, 彭明俊, 等. 种子雨研究进展[J]. 生态学杂志, 2007, 26(10): 1646 − 1652. YU Shunli, LANG Nanjun, PENG Mingjun, et al. Research advances in seed rain [J]. Chinese Journal of Ecology, 2007, 26(10): 1646 − 1652. [3] 张凌宇, 董灵波, 陈莹, 等. 大兴安岭中部天然次生林种子雨动态[J]. 应用生态学报, 2020, 31(12): 4035 − 4041. ZHANG Lingyu, DONG Lingbo, CHEN Ying, et al. Seed rain dynamics of natural secondary forest in the central part of Greater Xing’ an Mountains, China [J]. Chinese Journal of Applied Ecology, 2020, 31(12): 4035 − 4041. [4] 陈智平, 张涛, 赵万奎, 等. 黄土高原子午岭林区油松林种子雨强度及时空动态特征[J]. 中国沙漠, 2020, 40(3): 85 − 93. CHEN Zhiping, ZHANG Tao, ZHAO Wankui, et al. Seed rain intensity and spatiotemporal dynamics of Pinus tabulaeformis forest in Ziwuling forest area of the Loess Plateau [J]. Journal of Desert Research, 2020, 40(3): 85 − 93. [5] 李永霞, 安毅鹏, 侯萧棐, 等. 藏东南巨柏土壤种子库的时空分布格局[J]. 西部林业科学, 2021, 50(4): 7 − 12. LI Yongxia, AN Yipeng, HOU Xiaofei, et al. Spatial and temporal distribution pattern of soil seed bank of Cupressus gigantea in southeast Tibet [J]. Western Forestry Science, 2021, 50(4): 7 − 12. [6] 吴友贵, 朱志成, 吴倩倩, 等. 极危植物百山祖冷杉的种子雨[J]. 植物研究, 2023, 43(5): 711 − 719. WU Yougui, ZHU Zhicheng, WU Qianqian, et al. The seed rain of critically endangered plant Abies beshanzuensis [J]. Plant Research, 2023, 43(5): 711 − 719. [7] ZHU Chenxi, LIU Zhigang, YU Yangyang, et al. Seed composition and seasonal dynamics of Chinese fir plantation [J]. The Journal of Applied Ecology, 2018, 29(5): 1515 − 1522. [8] 包明慧, 方中平, 胡来庭, 等. 千岛湖片段化生境中木本植物种子雨基本特征及其影响因子[J]. 广西植物, 2023, 43(9): 1600 − 1610. BAO Minghui, FANG Zhongping, HU Laiting, et al. Basic characteristics and influencing factors of seed rain of woody plant in fragmented habitats in the Thousand Island Lake [J]. Guihaia, 2023, 43(9): 1600 − 1610. [9] 王通, 郑枭, 庞春梅, 等. 天目山常绿落叶阔叶混交林种子雨特征及其与群落组成的关系[J]. 生态学报, 2024, 44(2): 780 − 792. WANG Tong, ZHENG Xiao, PANG Chunmei, et al. Characteristics of seed rain and its relationship with community composition in the evergreen and deciduous broad-leaved mixed forest of Tianmu Mountain [J]. Acta Ecologica Sinica, 2024, 44(2): 780 − 792. [10] LU Yanlei, ZHANG Wenhui, YANG Yangbin, et al. Seed rain, soil seed bank and seedling regeneration of oak on different slopes in the middle part of Qinling Mountains [J]. Journal of Applied Ecology, 2019, 30(6): 1965 − 1973. [11] 邓自发, 谢晓玲, 王启基, 等. 高寒小嵩草草甸种子库和种子雨动态分析[J]. 应用与环境生物学报, 2003, 9(1): 7 − 10. DENG Zifa, XIE Xiaoling, WANG Qiji, et al. Dynamic analysis of seed rain and seed bank in Kobresia pygmaea meadow [J]. Chinese Journal of Applied &Environmental Biology, 2003, 9(1): 7 − 10. [12] 肖静, 黄力, 杨超, 等. 缙云山常绿阔叶林种子雨组成及其时空动态[J]. 林业科学, 2019, 55(7): 163 − 169. XIAO Jing, HUANG Li, YANG Chao, et al. Composition and spatial-temporal variation of the seed rain in an evergreen broad-leaved forest on Jinyun Mountain [J]. Scientia Silvae Sinicae, 2019, 55(7): 163 − 169. [13] 夏虹, 夏春燕, 宋海燕, 等. 重庆金佛山3种喀斯特森林群落木本植物种子雨、土壤种子库与幼苗更新[J]. 林业科学, 2022, 58(1): 1 − 11. XIA Hong, XIA Chunyan, SONG Haiyan, et al. Seed rain, soil seed bank, and seedling regeneration of woody plants in three karst forest communities in Jinfo Mountain, Chongqing [J]. Scientia Silvae Sinicae, 2022, 58(1): 1 − 11. [14] 冯倩倩, 周梅, 赵鹏武, 等. 大兴安岭南段不同林龄白桦种子雨与地表种子库研究[J]. 林业资源管理, 2019(4): 74 − 79. FENG Qianqian, ZHOU Mei, ZHAO Pengwu, et al. Study on seed rain and surface seed bank of Betula alba at different ages in the southern Daxing’ anling Mountains [J]. Forest Resources Management, 2019(4): 74 − 79. [15] 窦丽娜, 张文富, 邓晓保, 等. 西双版纳望天树林种子雨9年动态[J]. 生物多样性, 2018, 26(9): 919 − 930. DOU Li’na, ZHANG Wenfu, DENG Xiaobao, et al. Nine-year seed rain dynamics in Parashorea chinensis forest Xishuangbanna, Southwest China [J]. Biodiversity Science, 2018, 26(9): 919 − 930. [16] SAASTAMOINEN M, BOCEDI G, COTE J, et al. Genetics of dispersal [J]. Biological Reviews, 2018, 93(1): 574 − 599. [17] 沈泽吴, 吕楠, 赵俊. 山地常绿落叶阔叶混交林种子雨的地形格局[J]. 生态学报, 2004, 24(9): 1981 − 1987. SHEN Zewu, LÜ Nan, ZHAO Jun. The topographic pattern of seed rain of a mountain mixed evergreen and deciduous forest community [J]. Acta Ecologica Sinica, 2004, 24(9): 1981 − 1987. [18] 马琪瑶, 王振锡, 曲延斌, 等. 不同郁闭度天山云杉林分形维数特征[J]. 中南林业科技大学学报, 2022, 42(5): 65 − 73. MA Qiyao, WANG Zhenxi, QU Yanbin, et al. Study on fractal dimension characteristics ofPicea schrenkiana var. tianschanica forests with different canopy densities [J]. Journal of Central South University of Forestry and Technology, 2022, 42(5): 65 − 73. [19] 房园, 梁中, 张毓涛, 等. 天山云杉森林生态系统的水源涵养能力海拔梯度变化特征[J]. 生态环境学报, 2023, 32(9): 1574 − 1584. FANG Yuan, LIANG Zhong, ZHANG Yutao, et al. Water conservation capacity of Picea schrenkiana var. tianschanica forest ecosystem in mountains [J]. Chinese Journal of Eco-Environment, 2023, 32(9): 1574 − 1584. [20] 童海麦, 潘存德, 赵善超, 等. 天山云杉天然更新幼苗性状与微生境因子的关系[J]. 森林与环境学报, 2023, 43(2): 160 − 168. TONG Haimai, PAN Cunde, ZHAO Shanchao, et al. Relationship between microhabitat factors and natural regeneration seedling traits of Picea schrenkiana var. tianschanica [J]. Journal of Forestry and Environment, 2023, 43(2): 160 − 168. [21] ZHAO Wei, QI Yanni, ZHANG Jianping, et al. Comprehensive evaluation of salt tolerance during the germination period of flax resources [J]. Plant Research, 2019, 39(6): 955 − 963. [22] 李云红, 田松岩, 沃晓棠, 等. 伴生东北红豆杉针阔混交林种子雨时空动态[J]. 中南林业科技大学学报, 2022, 42(5): 109 − 118. LI Yunhong, TIAN Songyan, WO Xiaotang, et al. Spatial-temporal dynamics of the seed rain in coniferous and broad-leaved mixed forests of Taxus cuspidata [J]. Journal of Central South University of Forestry and Technology, 2022, 42(5): 109 − 118. [23] DAVID F N, MOORE P G. Notes on contagious distributions in plant populations [J]. Annals of Botany, 1954, 18(1): 47 − 53. [24] COMITA L S, HUBBELL S P. Local neighborhood and species’ shade tolerance influence survival in a diverse seedling bank [J]. Ecology, 2009, 90(2): 328 − 334. [25] 马晓勇, 李鑫. 灵空山油松林内凋落物与油松幼苗空间分布相关关系研究[J]. 山西师范大学学报(自然科学版), 2019, 33(3): 79 − 85. MA Xiaoyong, LI Xin. Study on the correlation between litter and the spatial distribution of Pinus tabulaeformis seedlings in Lingkongshan forest [J]. Journal of Shanxi Normal University (Natural Science Edition), 2019, 33(3): 79 − 85. [26] 陈国兴. 不同生境赤皮青冈种子萌发及幼苗高的生长规律研究[J]. 四川林业科技, 2022, 43(4): 117 − 123. CHEN Guoxing. Study on seed germination and seedling height in different habitats of Cyclobalanopsis ligustris [J]. Sichuan Forestry Science and Technology, 2022, 43(4): 117 − 123. [27] 程福山, 刘志宇, 吴蒙嘉, 等. 云冷杉天然次生林种子雨时空分布特征[J]. 东北林业大学学报, 2021, 49(6): 16 − 18, 52. CHENG Fushan, LIU Zhiyu, WU Mengjia, et al. Temporal and spatial distribution of deed rain in natural spruce-fir mixed forest [J]. Journal of Northeast Forestry University, 2021, 49(6): 16 − 18, 52. [28] 杨彬, 王玉, 郝清玉. 木麻黄海防林种子雨的时空动态[J]. 热带亚热带植物学报, 2019, 27(4): 367 − 375. YANG Bin, WANG Yu, HAO Qingyu. Spatial and temporal dynamics of seed rain of Casuarina equisetifolia coastal protection forest [J]. Journal of Tropical and Subtropical Botany, 2019, 27(4): 367 − 375. [29] 程福山, 周末, 吴蒙嘉, 等. 云冷杉针阔混交林枫桦种子雨时空分布及种子萌发特性研究[J]. 北京林业大学学报, 2020, 42(12): 32 − 39. CHENG Fushan, ZHOU Mo, WU Mengjia, et al. Spatiotemporal distribution and seed germination characteristics of Betula costata seed rain in the spruce-fir mixed forest [J]. Journal of Beijing Forestry University, 2020, 42(12): 32 − 39. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230502
 点击查看大图
点击查看大图
图(5) / 表(1)
计量
- 文章访问数: 1387
- HTML全文浏览量: 490
- PDF下载量: 37
- 被引次数: 0
