




 下载:
下载:
-
生物的总初级生产力(gross primary productivity)是指单位时间内生物(主要是指绿色植物)通过光合作用途径所固定的光合产物量或有机碳总量,又称为总第一性生产力或总生态系统生产力(gross ecosystem productivity)[1-2]。随着遥感技术的发展,对总初级生产力的区域模拟成为可能。近年来,已经展开了基于遥感数据的总初级生产力模型模拟研究并建立了一系列模型,如全球生产率模型(GLO-PEM),区域尺度遥感参数模型(C-Fix),通量观测的水分利用率模型(EC-LUE),中分辨率成像光谱仪(MODIS-PSN),植被光合模型(VPM),总初级生产力(TG)和植被指数(VI)模型。植被光合作用模型(VPM)作为一种改进的光能利用效率模型已成功地被用来估算森林和高寒草地等生态系统总初级生产力并取得了很好的效果[3-7]。毛竹Phyllostachys edulis是中国最重要的竹林资源类型,占全国竹林面积的70%左右,以其生长快、周期短等特有的结构和功能特性使其有别于其他类型的森林生态系统,在维护生态平衡方面发挥了重要作用[8]。在全球森林面积急剧下降的情况下,竹林却以每年3%左右的速度在递增。这意味着竹林将是一个不断增加的碳汇[9],对其总初级生产力的研究具有重要的意义。2010年,全球首座毛竹林碳通量观测塔在浙江省安吉县山川乡建成,这为毛竹林碳通量的准确估计提供了基础,并为遥感估测和涡度相关数据计算碳通量的交叉验证提供了可能。本研究拟运用VPM模型对浙江省安吉县山川毛竹林2011年的总初级生产力进行估算,同时分析其季相变化,以便探讨模型的适用性,了解毛竹林的物质循环和能量流动过程。
-
安吉县位于浙江省西北部,30°23′~30°53′N,119°14′~119°35′E,县域三面环山,中部为谷底平原,西南高东北低。属亚热带季风气候,年平均气温12.2~15.6 ℃,年降水量1 100~1 900 mm。土壤主要有红壤、黄壤、岩性土、潮土、水稻土等5类,山地土壤主要为红壤、黄壤、岩性土,其中以红壤面积最大。安吉县植被在区域上划属亚热带常绿阔叶林亚区,中亚热带常绿阔叶林北部亚地带。安吉县是“中国竹子之乡”,竹林面积居全国前列、浙江首位,特别是毛竹林资源丰富,毛竹林覆盖率达29.3%,面积占全县森林面积的41.2%,占全省毛竹林面积近十分之一[10]。研究区位于安吉县山川乡,全球首座毛竹林碳通量观测塔在此建成,通量塔总高40 m,主要通过辐射传感器、光三维超声风速仪、二氧化碳(CO2)水汽分析仪等科学仪器,全自动、全天候采集竹林不同冠层的二氧化碳浓度等竹林生态系统的宏观信息,以此观测、记录毛竹林的固碳功能
-
毛竹林的碳通量以及相关数据由数据采集器自动、连续采集存储,并在线计算30 min的汇总或平均数据。在实际的涡度通量观测中,由于传感器物理属性的局限性、坐标系选择不恰当以及夜间湍流混合不均匀等会造成观测数据的丢失或误差,因此需要对通量数据进行质量控制。本研究主要采用了二维坐标旋转和WPL校正,在EdiRe软件中完成。
在通量观测中,由于仪器、降水等原因影响,引起数据无效,因此须剔除不合格数据,包括:①二氧化碳通量观测数据通常在一个阈值范围内,据研究,森林生态系统的阈值为-3~3 mg·m-2·s-1[11]。本研究沿用该数据;?譺?訛剔除湍流不充分情况下的数据,即摩擦风速u*<0.2 m·s-1,并对处理后的资料进行数据插补。
经过以上数据处理后,全年完整的经过数据插补的0.5 h通量数据(ENE)被用来进行下一步的处理。净生态系统碳交换量ENE是总初级生产力(PGP)与生态系统呼吸(Re)之差。一般假设夜间涡度相关技术观测的ENE等于Re,利用夜间的ENE与夜间温度Ta之间的关系,以及Van’t Hoff方程得到ENE和Ta的关系,结合白天的温度数据估计白天的Re。PGP等于白天的Re减去ENE。
$${{\text{P}}_{\text{GP}}}\text{=}{{\text{R}}_{\text{e}}}\text{-}{{\text{E}}_{\text{NE}}}\text{ }$$ (1) 本研究中使用的温度数据Ta和光合有效辐射数据PAR从通量观测系统的梯度系统获得。这些数据同样以0.5 h 1次的频率进行测定并记录。其中,PAR数据由梯度系统测得的短波有效辐射数据乘以系数0.5获得[12]。为了匹配模型使用的MODIS(8 d时间分辨率)数据,逐日光合有效辐射被累积成8 d的总和值,对温度数据求8 d的平均值。
-
遥感数据使用美国宇航局/中分辨率影像辐射度计(National Aeronautics and Space Administration/Moderate Resolution Imaging Spectroradiometer,NASA/MODIS)陆地产品组按照统一算法开发的2011年共计46幅8 d最大值合成的陆地表面反照率产品(MOD09A1),空间分辨率为500 m,空间位置在全球正弦曲线投影SIN(sinusoidal project)系统中的编号为h28v05,数据格式为EOS-HDF。该数据可以从USGS(United States Geological Survey)上下载。对于下载的MODIS数据,用ENVI软件将坐标转换为UTM坐标系下WGS84坐标,并使用其中的蓝(459~479 nm),红(620~670 nm),NIR(841~875 nm)和SWIR(1 628~1 652 nm)4个波段的数据进行植被指数的计算。研究表明:使用单个像元和使用3×3,5×5像元进行反照率数据提取对于计算植被指数影响差异不显著[13]。因此,基于研究地点的经纬度信息,从MOD09A1产品中提取通量塔所在位置的单个像元的反照率数据,参照以下公式进行植被指数归一化(normalized difference vegetation index,INDV),增强植被指数(enhance vegetation index,IEV)和陆地表面水分指数(land surface water index,ILSW)的计算:
$$\begin{align} & {{I}_{NDV}}=\frac{{{P}_{NIR}}-{{\operatorname{P}}_{red}}}{{{P}_{NIR}}-{{\operatorname{P}}_{red}}}; \\ & {{I}_{EV}}=2.5\times \frac{{{P}_{NIR}}-{{\operatorname{P}}_{red}}}{{{P}_{NIR}}+1+6.0\times {{\operatorname{P}}_{red}}+7.5\times {{P}_{blue}}}; \\ & {{I}_{LSW}}={{P}_{NIR}}-{{P}_{SWIR}}{{P}_{NIR}}+{{P}_{SWIR}} \\ \end{align}$$ (2) 其中:ρ表示相应波段的地表反照率,下标NIR,red,blue和SWIR分别代表近红外、红、蓝和短波红外波段。INDV在高植被区易饱和、低植被区易受土壤背景影响,为了克服这种缺点,IEV引入蓝光波段来降低大气影响,同时,气溶胶对IEV的影响也不如对INDV的影响显著[14]。IEV对植被冠层变化敏感,包括冠层叶面积指数ILA,冠层类型和冠层结构;而INDV对叶绿素变化比较敏感; ILSW对于叶片含水量变化比较敏感,短波红外光谱波段对植物含水量和土壤湿度的变化敏感,而近红外光谱波段对这种变化表现的敏感性较小,通过两者的联系来反演植被叶片含水量[15]。
-
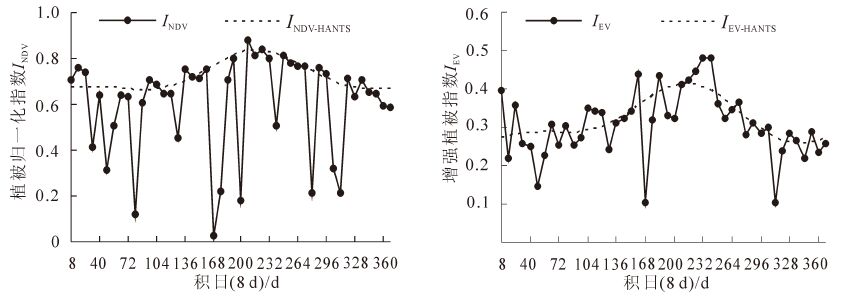
通过遥感手段获取的植被指数数据表征植被的生长状态。MOD09A1数据虽然已用最大值合成法(MVC)进行8 d数据合成,可以消除部分噪音污染,保证合成结果为周期内最接近现实情况的影像,但是由于算法本身的缺陷,仍然会受“云”等不良大气噪声的影响。这些残留噪声造成植被指数总趋势“衰退”[16]。本研究采用时间序列谐波分析法(harmonic analysis of time series,HANTS)对INDV和IEV数据进行异常值处理(主要是消除云层的影响)。由于ILSW受云层影响较小,且ILSW数值波动较大,异常值判别比较困难,因此未对ILSW进行异常值处理。通过HANTS对INDV和IEV数据进行处理后,使得全年46个数据呈现一定的周期性,保持全年数据的平滑状态。

图 1 HANTS方法处理前后INDV和IEV时间序列对比图
Figure 1. Comparison of normalized difference vegetation index,enhance vegetation index sequence before and after HANTS
-
Xiao等[17]提出了植被光合作用模型(vegetation photosynthesis model,VPM)用来估算植被生长季节——植被光合有效周期总初级生产力。VPM模型适用于站点尺度上单一植被类型的生态系统[18],一些专家和学者应用VPM模型实现了对于高寒草甸、温带草原、森林、高寒、高寒沼泽生态系统8 d时间分辨率、500 m空间分辨率总初级生产力的准确估测[3-7, 18]。模型中使用了2个改进的遥感植被指数,即增强植被指数IEV和陆地表面水分指数ILSW来反映冠层(PAV)吸收的有效辐射和叶子的年龄,模型基本构成如下:
$$\begin{align} & {{\text{P}}_{\text{GP}}}\text{=}{{\varepsilon }_{g}}\times {{F}_{PARPAV}}\times {{P}_{AR}}; \\ & \varepsilon ={{\varepsilon }_{0}}\times {{T}_{scalar}}\times {{W}_{\sec lar}}\times {{P}_{scalar}} \\ \end{align}$$ (3) 其中:PAR是光合有效辐射,FPARPAV代表光合有效辐射被冠层叶绿素吸收的部分,εg是光能利用效率。参数ε0表示表观量子效率或最大光能利用效率,Tscalar,Wscalar,Pscalar分别是温度、水分和物候对最大光能利用效率的影响函数。
在VPM模型中,FPAR被分为2个部分,一部分是冠层中含有叶绿素可以进行光合作用的部分FPARPAV,另一部分是不能进行光合作用的部分FPARnPAV(如枯叶、树枝等)[19]。在FPAR中只有FPARPAV部分对碳通量的估算起决定作用。在植被生长期间FPARPAV可以被看作是IEV的线性函数,且系数α被设置为1。表达式如下:
$${{\text{F}}_{\text{PARPAV}}}\text{=}\alpha \times {{I}_{EV}}$$ (4) Tscalar是温度对最大光能利用率的影响函数,采用陆地生态系统模型(TEM)的算法[6]:
$${{T}_{scalar}}=\frac{(T-{{T}_{max}})(T-{{T}_{max}})}{(T-{{T}_{min}})(T-{{T}_{max}})-{{(T-{{T}_{opt}})}^{2}}}$$ (5) 其中:Tmin,Tmax,Topt分别是植物进行光合作用的最低、最高和最适温度,T为某一时刻的气温。当空气温度低于最低光合作用时,Tscalar为0。在本文中,参考生长季IEV达到最大值时的温度以及研究资料,最适温度取值为25 ℃,研究发现当温度超过25 ℃时生长最快,超过25 ℃,光合累积减少;Tmin,Tmax分别设为-5 ℃和40 ℃[20]。
Wscalar表征水分对光合作用的影响,在VPM模型中被表示为地表水分指数的函数:
$${{W}_{scalar}}=\frac{1+{{I}_{LSW}}}{1+{{I}_{LSW}}_{\max }}$$ (6) 其中: ILSWmax是单个像元内植被生长季的最大ILSW值。本研究中ILSW最大值为0.282 25。
Pscalar表征物候状况对光合作用的影响,其计算与叶片的寿命有关,具有物种特异性[7]。由于毛竹林整年均有叶片覆盖,所以Pscalar被设置为1。
最大的光能利用效率因植被类型的不同而呈现显著的差异[21]。对于特定的植被类型可以通过文献调查或利用瞬时净生态系统碳交换数据(NEE)和光量子通量密度数据(PPFD)的关系获得。本研究毛竹林最大光能利用效率取值来源于杨爽的硕士论文,和本研究为同一数据源,其值为0.002 8 mg·μmol-1,由Michaelis-Menten模型拟合得出[19]。
-
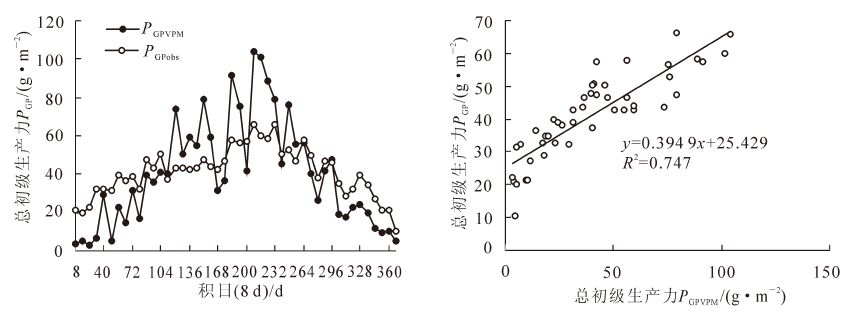
将由通量塔获取的温度和光合有效辐射整合成8 d数据,结合由MODIS影像计算获得的植被指数数据来驱动VPM模型,获得2011年、时间分辨率为8 d的毛竹林总初级生产力的估测值(PGPVPM),并和由通量塔数据获取的总初级生产力(PGPobs)进行对比验证。由图 2可以看出:利用VPM模型估测的总初级生产力和涡度相关通量观测的总初级生产力在研究时间段内季节动态一致。
VPM模型估算的2011年毛竹林总初级生产力为1 848.54 g·m-2,通量塔数据获得的2011年毛竹林总初级生产力为1 899.69 g ·m-2,相对误差为2.69%。尽管VPM模型估算的总初级生产力和通量塔获得的总初级生产力在总量上相对误差较小,在季节动态上趋势一致,但是VPM模型估算的总初级生产力(PGPVPM)并不能很好地与涡度协方差技术获得的总初级生产力(PGPobs)在细节上相匹配。在整个生长季节,PGPVPM与PGPobs的差异随着温度升高以及植被生长,逐渐增大,在夏天达到峰值,之后逐渐降低。PGPVPM与PGPobs线性关系的决定系数为0.747,低于同一模型在森林、农田以及草原上的估算准确度[3-7]。

图 2 利用VPM模型估算的基于MODIS的总初级生测量的总初级生产力(PGPobs)
Figure 2. Comparison of season changes in gross primary production from carbon flux site (PGPobs) and
-
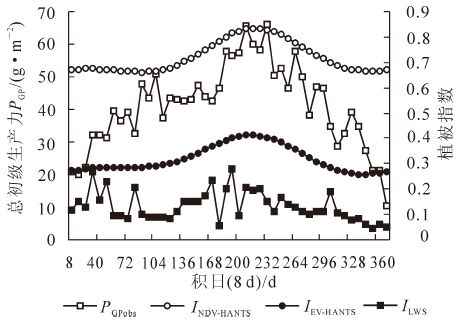
图 3是2011年山川站毛竹林INDV,IEV和ILSW的季节动态。在数值上INDV最大,其次是IEV,ILSW的值最小。这3个植被指数都在7月达到峰值,全年变化趋势与总初级生产力保持一致。植被指数与总初级生产力的判定系数为: IEV(R2=0.69)>INDV(R2=0.62)>ILSW(R2=0.12)。非生长季ILSW的显著升高源于研究区域冬季的积雪覆盖,积雪覆盖改变了地表反射率[22]。IEV与INDV相比,与通量塔数据获取的总初级生产力有更好的线性关系,IEV与总初级生产力的决定系数(R2)在0.69,而INDV为0.62,因此对总初级生产力的解释程度来看,IEV比INDV具有更好的解释结果。

图 3 毛竹林植被指数(IEV,INDV,ILSW)和通量塔获得的总初级生产力
Figure 3. Vegetation indices(IEV,INDV,ILSW)and the gross primary profuctionof the carbon flux site
-
本研究应用集成MODIS卫星影像数据和地面通量站点的观测数据的VPM模型来估算毛竹林总初级生产力。估测结果表明,对于毛竹林生态系统,IEV和总初级生产力(PGP)仍然具有更强的相关性,而INDV和PGP的相关性则相对较弱,表明IEV在表征地表植被覆盖状况方面优于INDV,进一步说明使用IEV代替INDV进行PGP估测的优势。VPM模型估测的PGP在季节变化趋势上和通量站点数据获得的PGP保持一致,在全年累积总量上接近,但是估测值和观测值之间仍然存在一定的差异,尤其是在生长季节。
尺度扩展问题是通量研究的热点和难点问题,通量塔的测量范围是有限的,通常为通量塔高度的100倍[23]。遥感模型的应用能帮助我们将单个站点的研究推导到区域尺度,遥感模型估算结果和实测数据的比较是验证和校正模型的第1步。限于时间等因素,本研究仅应用VPM模型对毛竹林的总初级生产力进行了估测和检验,并没有将模型进行尺度外推,且对模型的误差来源没有进行深入探讨,研究中的一些结论和观点还有待于进一步验证。根据目前在研究过程中遇到问题的一些理解和思考,认为在以下一些问题上需要进一步的完善和发展。
本研究仅利用VPM模型对毛竹林的总初级生产力(PGP)进行了估测,和通量塔观测数据进行了交叉验证,分析了毛竹林总初级生产力在2011年的季节和年际变化规律,但是并未对模型进行扩展研究。对VPM模型有效性的验证仅利用了2011年的观测资料,基于毛竹林生长的一些特殊性,叶龄、竹龄、叶位等均对毛竹的光合作用能力和碳汇能力有巨大的影响,并且毛竹存在大小年之分,对毛竹林生态系统碳通量的监测应充分考虑这些因素,需要长期数据的检验和分析,才能对毛竹林生态系统的碳循环有更深刻的认识。
在整个模型的应用中,关键参数只有IEV和ILSW,结合了温度和光合有效辐射数据,忽略了毛竹林生长过程中的一些其他因素;虽然植被指数能很好地反映植被的生长状况和季节变化趋势,但是并不能很好地解释物理过程,如:植被的光合作用过程、生态系统呼吸过程、横向碳交换等,需要进一步增加相关解释因子。
VPM模型中,最大光能利用率是一个最重要的参数,由植被类型确定,且在植被生长的不同阶段,最大光能利用率不是固定不变的。本研究参照杨爽的研究结果,将最大光能利用率取为一个固定值,且该值在计算过程中并没有考虑海拔、温度和坡度等因素对最大光能利用率的影响,可能导致估算结果的误差。在模型中仅用温度和水分因子对最大光能利用效率进行了线性调节,最大光能利用率和调节因子的误差会带来总初级生产力估算的不确定性。 由于数据观测和采样误差、相关数据的缺乏,模型在模拟生态系统过程中的偏差,以及模拟技术和算法的不确定性,使得在观测、模拟区域或全球尺度的陆地生态系统生产力过程中存在着一定的不确定性。在本研究中并没有对误差来源进行分析、量化和定量化表达,如何对这些误差来源和不确定性进行分析将是进一步研究的重点。
总之,毛竹林作为森林生态系统一个重要类型和组成部分,对其碳汇能力的研究应进一步深化。VPM模型虽然已在森林、草地、农田等生态系统中得到很好的应用和验证,在毛竹林生态系统中也有一定的精度,但是要精确估测毛竹林的总初级生产力仍需要进一步的发展和细化参数。VPM模型的最终目的是以更高的时间和空间分辨率实现对于区域或全球不同生态系统总初级生产力的准确估测。
Gross primary production in Phyllostachys edulis based on MODIS
-
摘要: 通过集成中分辨率成像光谱仪(MODIS)卫星影像数据与地面通量台站的观测数据,基于遥感的植被光合模型(VPM),估测了浙江省安吉县山川乡2011年的毛竹Phyllostachys edulis林总初级生产力(PGP)。研究表明:VPM模型估测的PGP(PGPVPM)在季节变化趋势上和通量站点数据获得的PGP(PGPobs)保持一致,VPM模型估算的2011年毛竹林总初级生产力为1 848.54 g·m-2,通量塔数据获得的2011年毛竹林总初级生产力为1 899.69 g·m-2,相对误差为2.69%。在全年累积总量上接近,但是估测值和观测值之间仍然存在一定的差异,尤其是在生长季节,PGPVPM的值要高于PGPobs;VPM模型估测的PGP和通量塔数据获取的PGP之间的决定系数为0.747,相关系数为0.864;且时间序列的增强植被指数(IEV)比PGP的相关关系强于归一化植被指数(INDV)与PGP的关系。研究表明:VPM对于站点和区域尺度的毛竹林生态系统PGP的模拟具有很大的潜力。Abstract: To analyze gross primary production(PGP) of Phyllostachys edulis from Shanchuan Village in Anji County,Zhejiang Province in 2011,the vegetation photosynthesis model (VPM) which was developed by integrating MODIS(Moderate-resolution Imaging Spectroradiometer)and flux measurements was used. Results showed that PGPVPM(1 848.54 g·m-2)was consistent with results from Shanchuan carbon flux site(1 899.69 g·m-2) having a relative tolerance of 2.69%. Though total PGP was similar,some differences occurred especially in the growing season,PGPVPM were higher than PGPobs. The correlation of PGPVPM to PGPobs was 0.747 and the correlation coefficient was 0.864. Further,time-series data for the IEV have a stronger linear relationship with the PGP than those for the Normalized Difference Vegetation Index. Results of this study demonstrate that the satellite-driven VPM has been potential for estimating site-level or regional Phyllostachys edulis PGP.
-
图 1 HANTS方法处理前后INDV和IEV时间序列对比图
Figure 1 Comparison of normalized difference vegetation index,enhance vegetation index sequence before and after HANTS
图 2 利用VPM模型估算的基于MODIS的总初级生测量的总初级生产力(PGPobs)
Figure 2 Comparison of season changes in gross primary production from carbon flux site (PGPobs) and
-
[1] 何学兆,周涛,贾根锁,等. 光合有效辐射总量及其散射辐射比例变化对森林GPP影响的模拟[J]. 自然资源学报,2011,26(4):619-634. HE Xuezhao,ZHOU Tao,JIA Gensuo,et al. Modeled effects of changes in the amount and diffuse fraction of PAR on forest GPP[J]. J Nat Resour,2011,26(4):619-634. [2] 于贵瑞,孙晓敏. 陆地生态系统通量观测的原理与方法[M]. 北京:高等教育出版社,2006. [3] WU Chaoyang,MUNGER J W,NIU Zheng,et al. Comparison of multiple models for estimating gross primary production using MODIS and eddy covariance data in Harvard Forest[J]. Remote Sens Environ,2010,114(12):2925-2939. [4] LI Zhengquan,YU Guirui,XIAO Xiangming,et al. Modeling gross primary production of alpine ecosystems in the Tibetan Plateau using MODIS images and climate data[J]. Remote Sens Environ,2007,107(3):510-519. [5] WANG Zheng,XIAO Xiangming,YAN Xiaodong. Modeling gross primary production of maize cropland and degraded grassland in northeastern China[J]. Agric For Meteorol,2010,150(9):1160-1167. [6] RAICH J W,RASTETTER E B,MELILLO J M,et al. Potential net primary productivity in South America:application of a global model[J]. Ecol Appl,1991,1(4):399-429. [7] 伍卫星,王绍强,肖向明,等. 利用MODIS影像和气候数据模拟中国内蒙古温带草原生态系统总初级生产力[J]. 中国科学D辑:地球科学,2008,38(8):993-1004. WU Weixing,WANG Shaoqiang,XIAO Xiangming,et al. Modeling gross primary production of a temperate grassland ecosystem in Inner Mongolia,China,using MODIS images and climate data[J]. Sci China Earth Sci,2008,38(8):993-1004. [8] 李雅红,江洪,原焕英,等. 西天目山毛竹林土壤呼吸特征及其影响因子[J]. 生态学报,2010,30(17):4590-4597. LI Yahong,JIANG Hong,YUAN Huanying,et al. Soil respiration and its regulating factors in the Phyllostachys edulis forest of west Tianmu Mountain,China[J]. Acta Ecol Sin,2010,30(17):4590-4597. [9] 王燕,王兵,赵广东,等. 我国毛竹林生态系统碳平衡研究进展[J]. 林业科技开发,2008,22(4):9-12. WANG Yan,WANG Bing,ZHAO Guangdong,et al. Progress studies on carbon budget in Phyllostachys edulis forest of China[J]. China For Technol,2008,22(4):9-12. [10] 潘春霞,李雪涛,吕玉龙. 安吉县毛竹资源及其生物量研究[J]. 浙江林业科技,2010,30(1):82-84. [11] 李春,何洪林,刘敏,等. China FLUX CO2通量数据处理系统与应用[J]. 地球信息科学,2008,10(5):557-565. LI Chun,HE Honglin,LIU Min,et al. The design and application of CO2 flux date processing system at China FLUX[J]. Geo-Inf Sci,2008,10(5):557-565. [12] XIAO Xiangming,ZHANG Qingyuan,HOLLINGER D,et al. Modeling gross primary production of an evergreen needleleaf forest using MODIS and climate data[J]. Ecol Appl,2005,15(3):954-969. [13] 刘敏. 基于RS和GIS的陆地生态系统生产力估算及不确定性研究[D]. 南京:南京师范大学,2008. LIU Min. Study on Estimation and Uncertainty of Terrestrial Ecosystem Productivity Based on RS and GIS:Take the Grassland Transect in Tibetan Plateau for Example[D]. Nanjing:Nanjing Normal University,2008. [14] 宋婷. 基于GIS和RS的青藏高原海北高寒草甸CO2通量的空间格局研究[D]. 南京:南京师范大学,2007. SONG Ting. Spatial Pattern Study of CO2 Fluxes of Haibei Alpine Meadow on the Qinghai-tibet Plateau Based on GIS and RS[D]. Nanjing:Nanjing Normal University,2007. [15] 严燕儿. 基于遥感模型和地面观测的河口湿地碳通量研究[D]. 上海:复旦大学, 2009. YAN Yaner. Carbon Flux in an Estuarine Wetland Estimated by Remote Sensing Model and Ground-based Observations[D]. Shanghai:Fudan University,2009. [16] XIAO Xiangming,HOLLINGER D,ABER J,et al. Satellite-based modeling of gross primary production in an evergreen needleleaf forest[J]. Remote Sens Environ,2004,89(4):519-534. [17] 王鹤松,贾根锁,冯锦明,等. 我国北方地区植被总初级生产力的空间分布与季节变化[J]. 大气科学, 2010,34(5):882-890. WANG Hesong, JIA Gensuo,FENG Jinming,et al. Spatial distribution and seasonality of gross primary production in northern China[J]. Chin J Atmos Sci,2010,34(5):882-890. [18] 王旭峰,马明国,李新,等. 遥感GPP模型在高寒草甸的应用比较[J]. 遥感学报,2012,16(4):751-763. WANG Xufeng,MA Mingguo,LI Xin,et al. Comparison of remote sensing based GPP models at an alpine meadow site[J]. J Remote Sens,2012,16(4):751-763. [19] 杨爽. 浙江安吉毛竹林生态系统CO2通量观测研究[D]. 临安:浙江农林大学,2012. YANG Shuang. The Study of CO2 in Anji Phyllostachys edulis Ecosystems[D]. Lin'an:Zhejiang A & F University, 2012. [20] 蔡小红. 崇义县毛竹种植气候区划[J]. 现代农业科技,2012(8):298-307. CAI Xiaohong. The climate division of Phyllostachys edulis in Chongyi[J]. Modern Agric Sci Technol,2012(8):298-307. [21] 赵育民,牛树奎,王军邦,等. 植被光能利用率研究进展[J]. 生态学杂志,2007,26(9):1471-1477. ZHAO Yumin,NIU Shukui,WANG Junbang,et al. Light use efficiency of vegetation:a review[J]. Chin J Ecol,2007,26(9):1471-1477. [22] -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.02.003
 点击查看大图
点击查看大图
图(3)
计量
- 文章访问数: 3771
- HTML全文浏览量: 698
- PDF下载量: 619
- 被引次数: 0
