




-
叶面积指数(LAI)为投影到水平基准面上的每单位地表面积上总绿叶面积的一半,是描述植被生长状况和开展陆面过程研究的重要参数[1]。传统获取LAI数据的地面测量方法只适合小范围区域的短期监测,而卫星遥感具有大面积和重复观测的特点,为获取大尺度长期LAI提供了有效的数据来源[2]。目前已经有多个全球范围、长时间序列LAI产品,如AVHRR LAI[3]、MODIS LAI[4]、GEOV2 LAI[5]、PROBA-V LAI[6]、VIIRS LAI[7]、GLASS LAI[8]、MUSES LAI[9]等。然而这些产品的空间分辨率大多较低,在反演过程中受到云雪覆盖、气溶胶、大气辐射以及传感器自身参数等因素影响,导致数据存在算法误差,数据集存在异常值、数据缺失和精度偏低等问题,限制了产品的应用研究[10]。
多源数据同化融合技术可以显著提高数据精度。基于集合卡尔曼滤波(EnKF)的数据同化算法具有整合不同来源和不同误差信息数据的能力,通过观测数据对模型模拟结果进行调整和校正,提高模拟数据精度[11]。LIU等[12]整合3种不同来源数据的时间、光谱和角度信息,利用EnKF技术将反演的LAI进行气候学的递归更新,可以填补缺失数据,获得更准确的LAI产品。李喜佳等[13]耦合辐射传输模型和动态模型,运用双集合卡尔曼滤波分别对林地、草地和农田的LAI进行同化,改善了模型预测能力。周红敏等[14]利用EnKF方法耦合神经网络模型和动态模型,将反演得到的Landsat高分辨率LAI数据与MODIS LAI融合生成30 m空间分辨率的LAI数据集,估算结果与地面实测数据一致性较高。然而,LAI产品在反演过程中输入数据和模型算法的不确定性导致其存在质量问题,如MODIS产品由主算法反演得到的高质量LAI在全球尺度上约占67%,其余为备用算法反演得到的低质量LAI[15]。主算法失效主要是由于残云或较差的大气校正,失效后采用备用算法,导致备用算法检索数据准确性较低[16]。
为了尽可能减少LAI产品由于观测不稳定造成的不确定性,本研究将多年LAI产品数据作为先验知识,利用产品的质量控制文件构建LAI背景库修正其低质量数据,利用混合像元分解方法对2017年的MODIS、VIIRS和PROBA-V的LAI数据进行降尺度处理,实现LAI时序数据的重建工作。利用3种LAI数据集和Sentinel-2反射率数据,结合EnKF技术耦合LAI动态模型和辐射传输模型生成同化LAI,最后对3种同化后的LAI数据进行赋权融合,达到减少不同来源数据差异、提高LAI产品估算精度和一致性的目的。
-
研究区位于黑龙江省尚志市帽儿山实验林场(45°14′~45°29′ N,127°29′~127°44′ E,图1),平均海拔为428.0 m,属于温带季风气候,年均气温为2.8 ℃,土壤为典型暗棕壤。植被属于长白山植物区系,主要针叶树种有红松Pinus koraiensis、落叶松Larix gmelinii、樟子松Pinus sylvestris var. mongolica等,主要阔叶树种有白桦Betula platyphylla、山杨Populusdavidiana、色木槭Acer mono、水曲柳Fraxinus mandshurica、胡桃楸Juglans mandshurica等[17]。
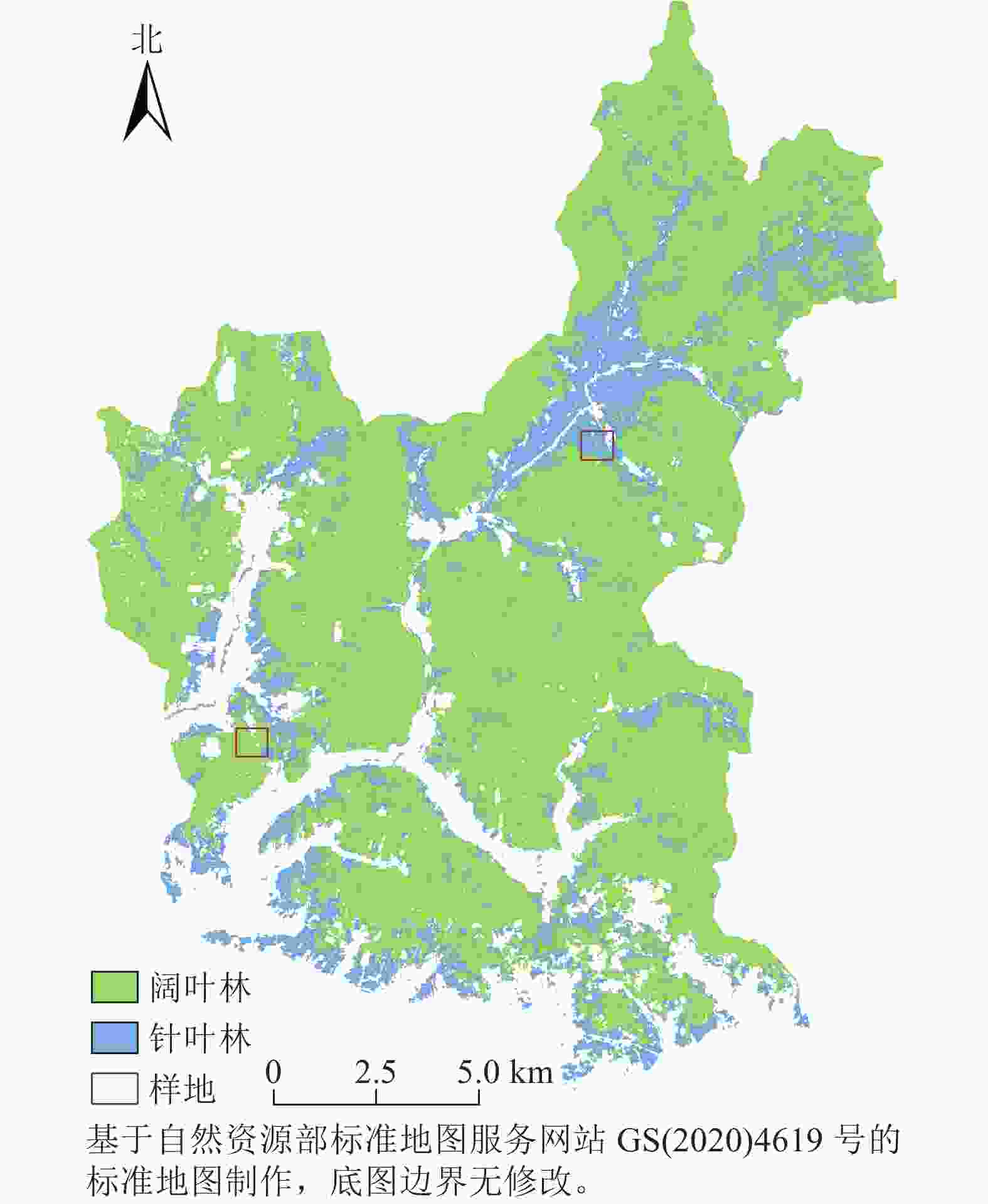
图 1 研究区和样地分布示意图
Figure 1. Geographical location and plot distribution of the study area
-
在研究区内根据植被类型选取2块1 km×1 km大样地(图1),每个大样地内包含5块100 m×100 m小样地。使用LAI-2200和TRAC冠层分析仪测量LAI[18−19]。使用GPS记录位置坐标。
-
考虑到研究区数据的可获取性,本研究选取了3种应用广泛的LAI产品(表1)。利用HEG软件,对MOD15A2H和VNP15A2H数据进行格式转换和投影变换。利用ArcGIS 10.2对CGLS 300m V1数据进行投影变换。
表 1 LAI产品数据及基本信息
Table 1. LAI product data and basic information
产品名称 传感器 空间分辨率/m 时间分辨率/d 投影方式 数据来源 MOD15A2H MODIS 500 8 正弦曲线 https://ladsweb.modaps.eosdis.nasa.gov/ VNP15A2H VIIRS 500 8 正弦曲线 https://ladsweb.modaps.eosdis.nasa.gov/ CGLS 300m V1 PROBA-V 300 10 简易圆柱 https://land.copernicus.eu/global/ -
选取2017年云层覆盖率低于10%的11幅Sentinel-2影像,利用Sen2cor工具进行大气校正得到L2A级数据,利用SNAP工具箱将数据重采样至10 m分辨率,并转换为与LAI产品一致的UTM WGS84投影。
-
采用2017年8月14日的Sentinel-2 MSIL2A影像,利用ENVI 5.3进行最大似然法分类[20]。定义了农田、阔叶林、针叶林、草地和其他5类训练样本,通过实地调查数据和30 m地表覆盖分类产品GLC_FCS30相结合的方式目视判读各样本点的类别,采用混淆矩阵的方法进行精度检验,计算得到的总体精度为79.34%,Kappa系数为0.7517,整体分类质量高。将分类后的影像掩膜提取生成10 m分辨率的阔叶林和针叶林土地覆盖图(图1)。
-
利用2014—2016年的MOD15A2H和VNP15A2H的质量控制数据集进行像元质量标识,采用时间加权插值算法建立两者的LAI背景库,基于背景库对2017年的MODIS LAI和VIIRS LAI进行数据重建,计算公式参考文献[15]。最后使用三次样条帽盖算法(LACC)[21]对初修正的2个数据集和PROBA-V LAI数据集进行平滑处理,去除异常值。
根据解混理论,假设每个粗分辨率像元的反射率是构成混合像元的每个土地覆盖类型反射率的线性组合,同理混合像元的LAI由不同类别的LAI及其在像元中所占的面积比例(即丰度)构成,基于Sentinel-2的10 m土地覆盖类型图来计算丰度,利用约束最小二乘法计算混合像元内不同类别的平均LAI值,并根据土地覆盖类型图分配到对应类别的像元上,从而得到10 m分辨率的高精度LAI数据[22]。
-
利用降尺度后的3种LAI数据集分别构建经验性的LAI动态模型,将模型预测的LAI作为参数,连同其他参数一起输入PROSAIL模型模拟森林冠层反射率。同时,将Sentinel-2的红光波段(Band 4)和近红外波段(Band 8)影像作为观测数据,通过EnKF算法比较观测数据集合和模拟反射率集合,对预测LAI进行校正。结合精度评价结果对同化后的3种LAI数据进行赋权融合。
-
LAI动态模型能模拟叶面积指数[23],描述LAI随时间的动态变化过程,计算公式参考文献[24]。PROSAIL模型是由叶片光学特性模型(PROSPECT)和冠层二向反射率模型(SAIL)组合成的耦合模型,参数设置参考文献[25−27]。PROSPECT模型输入参数为:叶片结构参数(N=1.4)、叶绿素含量(Cab=58 μg·cm−2)、叶片含水量(Cw=0.025 g·cm−2)和干物质含量(Cm=0.009 g·cm−2);SAIL模型输入参数为:叶面积指数(LAI)﹑土壤亮度参数(Psoil=0.25)、热点参数(Hspot=0.037)、太阳天顶角(tts=30)、观测天顶角(tto=0)和相对方位角(psi=0),叶片反射率和透射率采用PROSPECT模型输出数据,tts、tto和psi可通过遥感影像头文件获取。
-
EnKF是基于蒙特卡罗的集合预报方法估计预报误差统计量的数据同化技术[11],计算公式如下:
$$ {{\boldsymbol{A}}}_{t}^{\mathrm{f}}={\boldsymbol{M}}\left({{\boldsymbol{A}}}_{t-1}^{\mathrm{a}}\right)+\varepsilon \text{;} $$ (1) $$ {{\boldsymbol{A}}}_{t}^{\mathrm{a}}={{\boldsymbol{A}}}_{t}^{\mathrm{f}}+{{\boldsymbol{P}}}_{t}{{\boldsymbol{H}}}^{\mathrm{T}}({\boldsymbol{H}}{{\boldsymbol{P}}}_{t}{{\boldsymbol{H}}}^{\mathrm{T}}+{{\boldsymbol{R}}}_{t}{)}^{-1}({\boldsymbol{D}}-{\boldsymbol{H}}{{\boldsymbol{A}}}_{t}^{\mathrm{f}}) \text{;} $$ (2) $$ {{\boldsymbol{P}}}_{t}=\frac{1}{N-1}({{\boldsymbol{A}}}_{t}^{\mathrm{f}}-{\overline{{{\boldsymbol{A}}}_{t}^{\mathrm{f}}}})({{\boldsymbol{A}}}_{t}^{\mathrm{f}}-{\overline{{\boldsymbol{A}}_{t}^{\mathrm{f}}}}{)}^{\mathrm{T}} \text{;} $$ (3) $${\overline{{{\boldsymbol{A}}}_{t}^{\mathrm{a}}}}=\frac{1}{N}\sum _{i=1}^{N}{{\boldsymbol{A}}}_{{{it}}}^{\mathrm{a}} 。 $$ (4) 式(1)~(4)中:${\boldsymbol{A}}=({x}_{1}, {x}_{2}, \cdots , {x}_{N})\in {{{{{\boldsymbol{R}}}}}}^{n\times N}$,是包含N个成员的状态变量集合,x表示n维状态向量(LAI);M代表LAI动态模型,$ \varepsilon $为模型误差;${{\boldsymbol{A}}}_{t}^{\mathrm{f}}$表示t时刻模型预测的状态变量集合;a表示分析状态变量,T表示转置矩阵。${\boldsymbol{D}}\in {{{{{\boldsymbol{R}}}}}}^{m\times N}$代表扰动观测集合矩阵,是通过给定的m维观测向量${\boldsymbol{d}}\in {{{{{\boldsymbol{R}}}}}}^{m}$加上N个扰动构成,本研究中m=2。${\boldsymbol{P}}\in {{{{{\boldsymbol{R}}}}}}^{n\times n}$代表预测误差协方差矩阵,${\boldsymbol{R}}\in {{{{{\boldsymbol{R}}}}}}^{m\times m}$代表观测误差协方差矩阵,其算法与P相似,如式(3)所示。式(4)中,$\overline{{{\boldsymbol{A}}}_{t}^{\mathrm{a}}}$为状态最优估计值。H代表PROSAIL模型,因为该模型是非线性观测算子,不适用于标准EnKF,所以构建了扩展集$\widehat{{\boldsymbol{A}}}=[{{\boldsymbol{A}}}_{t}^{\mathrm{f}},\; h({{\boldsymbol{A}}}_{t}^{\mathrm{f}}\left)\right]$,将H转化成线性算子$\widehat{{\boldsymbol{H}}}$,此时P为扩展集协方差矩阵,故式(2)改写如下:
$$ {{\boldsymbol{A}}}_{t}^{\mathrm{a}}={{\boldsymbol{A}}}_{t}^{\mathrm{f}}+{\boldsymbol{P}}{\widehat{{\boldsymbol{H}}}}^{\mathrm{T}}(\widehat{{\boldsymbol{H}}}{\boldsymbol{P}}{\widehat{{\boldsymbol{H}}}}^{\mathrm{T}}+{{\boldsymbol{R}}}_{t}{)}^{-1}({\boldsymbol{D}}-\widehat{{\boldsymbol{H}}}{{\boldsymbol{A}}}_{t}^{\mathrm{f}})。 $$ (5) 本研究观测集合和第1次的LAI预测集合是通过添加N个扰动(符合均值为0,方差为1的标准正态分布的随机数)生成,其中集合数N取值为100。
-
对3种同化LAI数据的精度检验结果归一化处理后分别赋予权重,将3种同化LAI数据分别与各自的权重相乘得到融合LAI数据。除了相关系数外的指标数值的绝对值越大,精度越差,需要在归一化处理前对它们取倒数,计算公式如下:
$$ {Y}_{i,j}=\left|1/{Y}_{i,j}\right|,j=\mathrm{1, \;2},\; \cdots, \;n-1 \text{;} $$ (6) $$ {Y}_{i}=\frac{1}{n}\sum _{j=1}^{n}\left({Y}_{i,j}/\sum _{i=1}^{m}{Y}_{i,j}\right) 。 $$ (7) 式(6)~(7)中:n表示评价指标的个数,m表示LAI同化数据的个数,i=1$, \cdots , $ m,$ {Y}_{i,j} $表示第i个LAI同化数据对应的第j个指标的值,$ {Y}_{i} $表示第i个LAI同化数据在所有指标进行归一化后的值。
-
利用实测数据对3种同化LAI数据和融合LAI数据进行精度评价,评价指标选用相关系数(R)、均方根误差(ERMSE)、平均绝对误差(EMAE)和相对偏差(ERE),计算公式参考文献[28]。
-
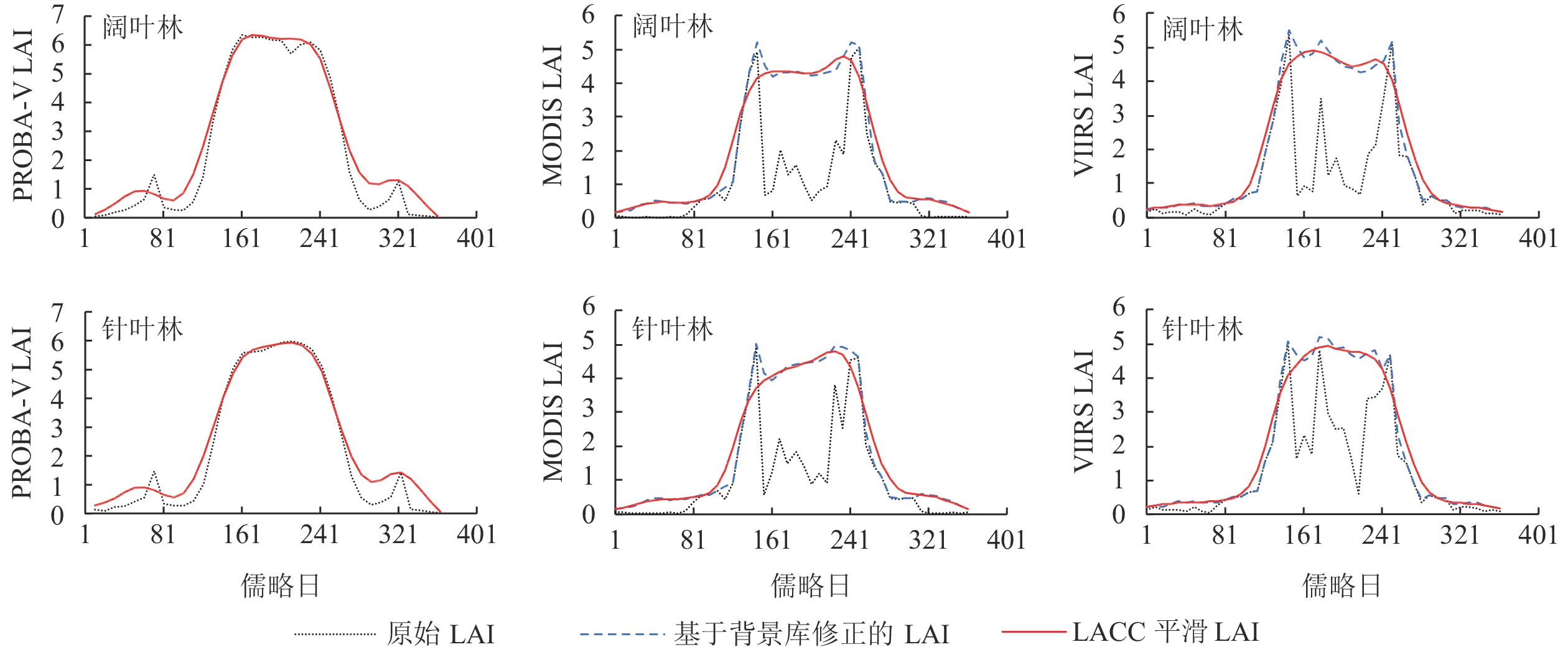
由图2可见:原始MODIS和VIIRS LAI时序曲线存在较为剧烈的跳跃现象,影响了其对植被物候特征的表征,而PROBA-V LAI时序曲线较为平滑,非生长季出现较小的跳跃现象。各产品都清晰表现了植被LAI夏季高、冬季低的生长特点。在植被LAI关键点缺失的情况下,MODIS和VIIRS产品基于背景库修正的方法,能够更好地重建LAI时序曲线,修正后的LAI与PROBA-V LAI差异较小,且时序曲线在形态上与PROBA-V也更加接近。修正后的LAI使用LACC算法进行平滑处理,平滑后的非生长季LAI均值都有所提高。在生长季的阔叶林VIIRS和PROBA-V LAI变化趋势相似;在生长季的针叶林MODIS和PROBA-V LAI变化趋势相似。在生长季,PROBA-V阔叶林的LAI高于针叶林。总体上,PROBA-V LAI均值高于MODIS和VIIRS。

图 2 3种LAI时序曲线在阔叶林和针叶林区域的变化情况以及重建效果
Figure 2. Changes and reconstruction effects of 3 LAI time series curves in broad-leaved forest and coniferous forest areas
-
由表2可见:3种LAI产品与实测数据的相关系数均较低,均方根误差、平均绝对误差和相对偏差值均较高,说明3种LAI产品与实测数据之间的一致性不高,没有明显的线性相关关系。经过EnKF同化后,3种LAI产品的相关系数均提升到0.49以上,MODIS LAI、VIIRS LAI和PROBA-V LAI的相关系数在阔叶林分别提高了0.57、0.52和0.57,在针叶林分别提高了0.52、0.32和0.40。3种同化后LAI数据的均方根误差均在0.40以下。其中,PROBA-V LAI在阔叶林和针叶林的均方根误差最小。VIIRS LAI在阔叶林的均方根误差小于MODIS LAI,而MODIS LAI在针叶林的均方根误差小于VIIRS LAI。相比同化前,VIIRS LAI在针叶林的均方根误差减小程度最低,仅减小了0.67,而VIIRS LAI在阔叶林的均方根误差减小程度最高,减小了1.69。对比3种同化后LAI数据的平均绝对误差,MODIS和VIIRS LAI在阔叶林的平均绝对误差最小,均为0.60,其中,VIIRS LAI在阔叶林的平均绝对误差相比同化前减小了1.11,减小程度最高。对比相对偏差,同化后除了MODIS LAI在阔叶林的相对偏差为负值,其他相对偏差都为正值。相比同化前,除了MODIS LAI在针叶林的偏差增大(同化前相对偏差=−0.72%,同化后相对偏差=8.34%),其他数据的偏差均有不同程度的降低。
表 2 3种LAI产品在阔叶林和针叶林数据同化前后的精度指标
Table 2. Accuracy indexes of 3 LAI products before and after data assimilation in broad-leaved and coniferous plots
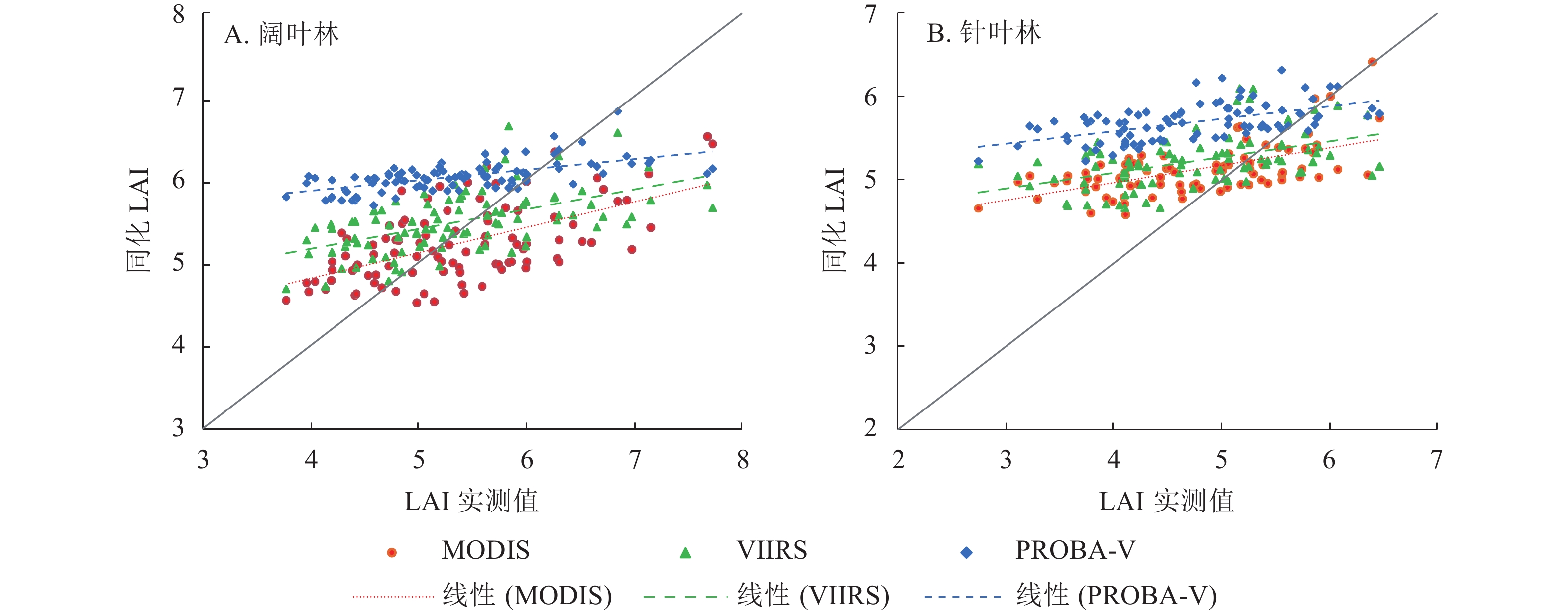
LAI产品 样地 同化前 同化后 R ERMSE EMAE ERE/% R ERMSE EMAE ERE/% MODIS 阔叶林 0.02 1.60 1.37 −23.75 0.59 0.37 0.60 −2.56 针叶林 0.07 1.46 1.30 −0.72 0.59 0.24 0.64 8.34 VIIRS 阔叶林 0.04 2.00 1.71 −28.62 0.56 0.31 0.60 2.29 针叶林 0.17 0.95 0.77 11.55 0.49 0.28 0.72 10.53 PROBA-V 阔叶林 0.05 1.20 1.04 15.24 0.62 0.14 0.86 12.49 针叶林 0.16 1.54 1.39 28.39 0.56 0.19 1.03 20.53 由图3可见:同化后LAI数据在阔叶林和针叶林从大到小依次为PROBA-V、VIIRS、MODIS,PROBA-V LAI具有明显的高估现象,与表2的相对偏差情况一致,与PROBA-V LAI原始数据就具有高估情况有关。同化后MODIS和VIIRS LAI数值相近,与两者原始数据就具有较高一致性有关。
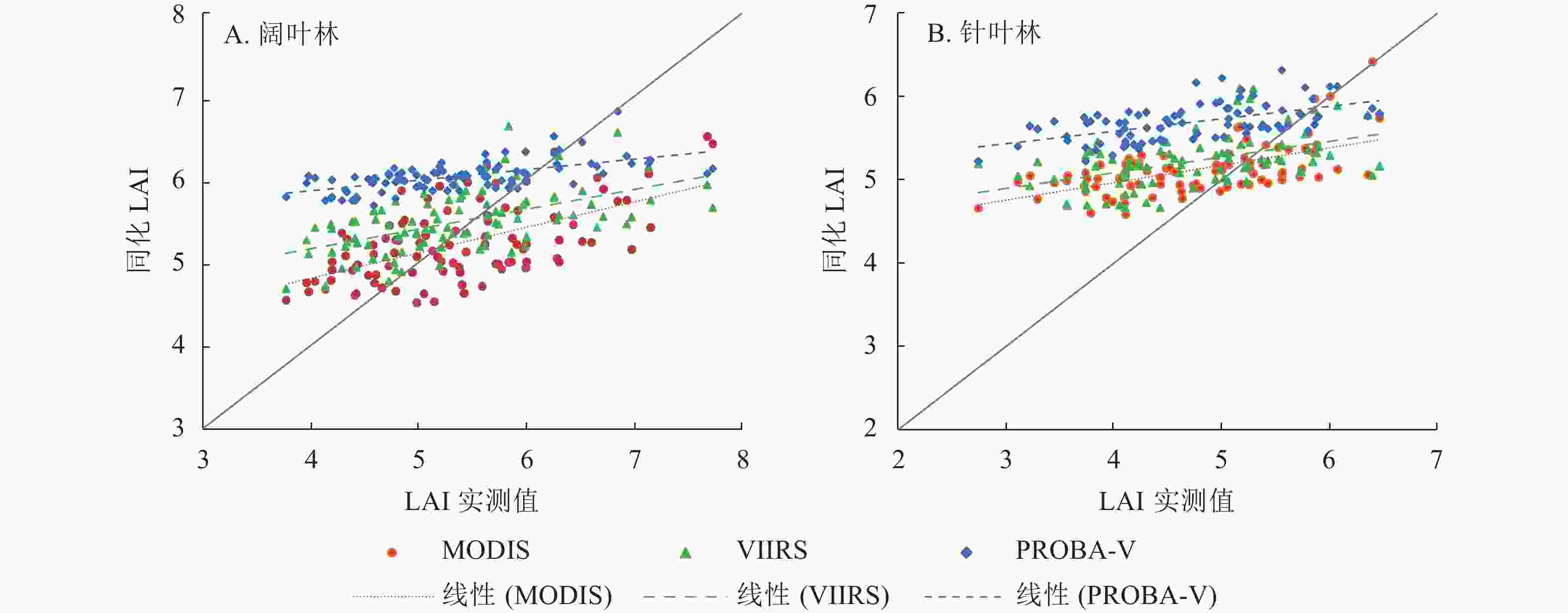
图 3 同化LAI与实测值的散点验证图
Figure 3. Scatter verification diagram of assimilated LAI and measured value
-
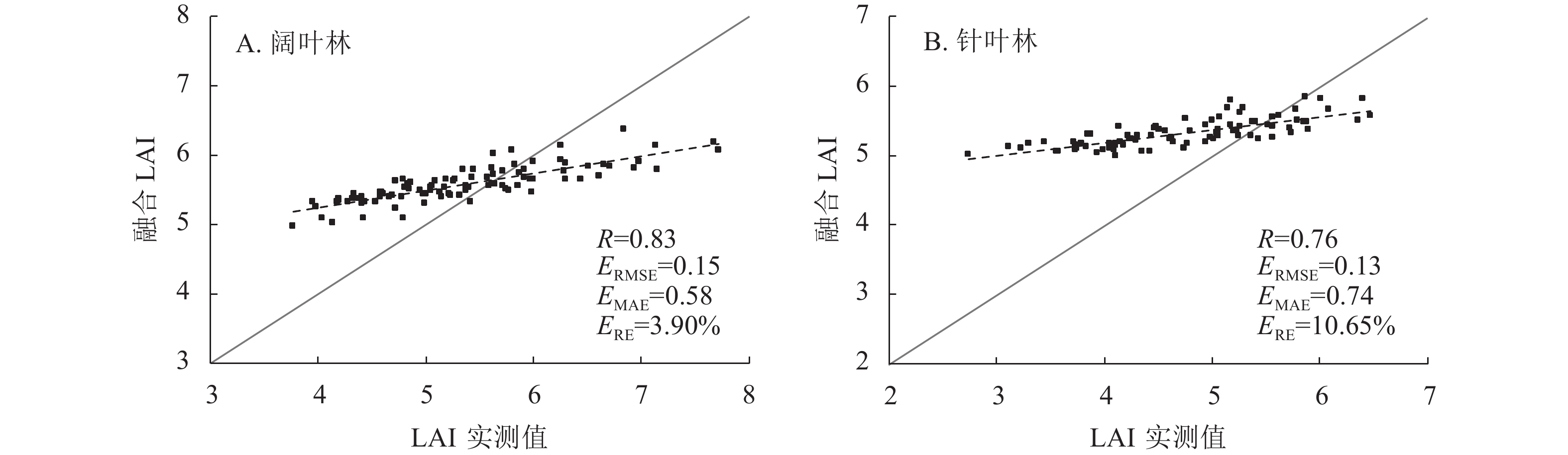
由图4可见:相比同化后LAI,融合后LAI与实测数据的相关性较高。融合LAI在阔叶林和针叶林的相关系数分别为0.83和0.76,均方根误差均在0.20以下,平均绝对误差和相对偏差趋势相同。在阔叶林,相比同化后的MODIS、VIIRS和PROBA-V LAI,融合后的LAI相关系数分别提高了0.24、0.27和0.21;对比同化后的MODIS和VIIRS LAI,融合后的LAI均方根误差分别降低了0.22和0.16;融合后的LAI平均绝对误差相比同化后的降低了0.02~0.28;融合后的LAI 相对偏差相比同化后的PROBA-V LAI,偏差降低程度最大,降低了8.59%。在针叶林,相比同化后的MODIS、VIIRS和PROBA-V LAI,融合后的LAI相关系数分别提高了0.17、0.27和0.20;均方根误差分别降低了0.11、0.15和0.06;对比同化后的PROBA-V LAI,融合后的LAI平均绝对误差降低了0.29;融合后的LAI相对偏差相比同化后的PROBA-V LAI,偏差降低程度最大,降低了9.88%。
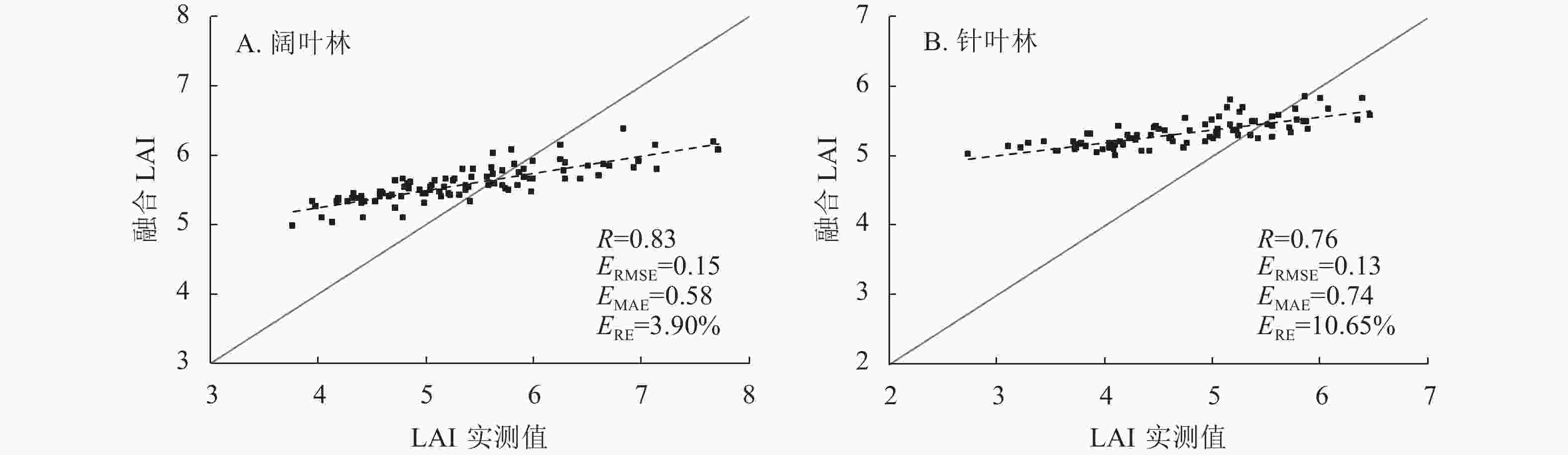
图 4 融合LAI与实测值的散点验证图
Figure 4. Scatter verification diagram of fused LAI and measured values
-
一般而言,LAI时序曲线是光滑且连续的,因此,当严重锯齿状的变化曲线中出现与正常趋势差距较大的拐点时,认为不符合植被正常生长规律[29]。MODIS和VIIRS LAI时序曲线存在剧烈的跳跃现象,PROBA-V LAI即使利用时间平滑和间隙填充技术,使得时序曲线较为平滑,但仍存在较多缺失值[30],因此在产品应用前进行重建至关重要。本研究对3种LAI产品进行重建,初步改善了数据集质量较差的问题,使得三者LAI时序曲线变化趋势趋于一致。由于LAI背景库值反映的是研究区LAI生长变化的平均趋势,受空间和时间因素的影响非常大,因此与实际值有一定的偏差。本研究利用混合像元分解法进行降尺度处理,由于粗分辨率像元存在异质性,会存在一定的不确定性,经过降尺度处理,样地内同质像元具有相同的LAI值。本研究表明:经过同化方法校正了LAI值,使得LAI产品与实测值具有较高的相关性,但存在低值高估和高值低估的现象。经过EnKF同化,3种LAI产品精度和一致性均有所提高,经过同化融合后的LAI数据,降低了单一LAI数据误差,可有效弥补地面实测数据缺乏的不足,但能否满足植被监测和地表模型输入参数的要求还需进一步验证。本研究通过将LAI数据降尺度后作为同化算法的输入数据,使融合LAI的精度得以提高,生成10 m空间分辨率LAI数据,然而由于缺少研究区长期有效的观测数据,生成数据的时间分辨率受限制,在后续研究中可以考虑加入其他反射率数据,以期得到高精度长时序的LAI产品。
-
本研究结果表明:3种原始LAI产品与实测数据的一致性均较差,精度较差;EnKF同化后的LAI更接近实测数据;相比针叶林,阔叶林LAI同化效果较好;总体上MODIS LAI同化效果最好,其次是PROBA-V LAI,VIIRS LAI在阔叶林和针叶林的同化效果存在较大差异。融合后的LAI精度优于同化后单一的LAI产品,其相关系数为0.83和0.76,比同化后LAI提升了0.17~0.27。同化融合后的LAI数据提高了单一LAI产品的精度,减小了不同源数据的差异,达到了充分利用多源遥感数据优势和提高产品精度的目的,可以为开展高精度LAI数据生成方法的研究提供参考。
Reconstruction and fusion correction method of multi-source LAI products in Maoershan forest based on ensemble Kalman filter
-
摘要:
目的 现有叶面积指数(LAI)产品大多存在分辨率低、数据异常和精度低等问题,难以满足某些应用需求。因此,本研究提出一种多源LAI数据的融合方法,以减少不同来源数据的差异并提高产品精度。 方法 以帽儿山实验林场的阔叶林和针叶林区域为研究区,基于2017年的MODIS、VIIRS和PROBA-V 的LAI产品,利用多年LAI数据作为先验知识建立LAI背景库修正低质量数据,对3种LAI数据集进行混合像元分解的降尺度处理,基于Sentinel-2反射率产品耦合集合卡尔曼滤波(EnKF)算法、LAI动态模型和辐射传输模型进行数据同化,最后对同化后的3种LAI数据进行赋权融合,使用实测数据进行精度评价。 结果 在阔叶林,同化后的MODIS、VIIRS和PROBA-V LAI与实测数据的相关系数分别为0.59、0.56和0.62,比原始数据提升了0.57、0.52和0.57;均方根误差分别为0.37、0.31和0.14,比原始数据减小了1.23、1.69和1.06。在针叶林,同化后的MODIS、VIIRS和PROBA-V LAI与实测数据的相关系数分别为0.59、0.49和0.56,比原始数据提升了0.52、0.30和0.40;均方根误差分别为0.24、0.28和0.19,比原始数据减小了1.22、0.67和1.35。通过融合方法,阔叶林LAI和针叶林LAI的相关系数分别为0.83和0.76,比同化后数据的相关性更高;均方根误差分别为0.15和0.13,比同化后数据的误差更小。 结论 通过数据同化提升了3种LAI产品精度,融合后LAI较同化后单一LAI具有更高的精度和可靠性。图4表2参30 -
关键词:
- 叶面积指数(LAI) /
- MODIS /
- VIIRS /
- PROBA-V /
- 重建 /
- 集合卡尔曼滤波(EnKF) /
- 数据融合
Abstract:Objective Most of the existing leaf area index (LAI) products have some problems, such as low resolution, abnormal data and low accuracy, which are difficult to meet the requirements of some applications. Therefore, this study proposes a method of fusing multi-source LAI data to reduce the differences of data from different sources and improve product accuracy. Method The broad-leaved forest and coniferous forest in Maoershan experimental forest farm were taken as the research area. Based on MODIS LAI, VIIRS LAI and PROBA-V LAI products in 2017, the LAI background database was established to correct low-quality data by using years of LAI data as prior knowledge, and 3 LAI data sets were downscaled by mixed pixel decomposition. Based on Sentinel-2 reflectivity product coupling ensemble Kalman filter (EnKF) algorithm, LAI dynamic model and radiative transfer model, data assimilation was carried out. Finally, 3 LAI data after assimilation were weighted and fused, and the accuracy was evaluated by using measured data. Result In broad-leaved forest, the correlation coefficients between the assimilated MODIS, VIIRS and PROBA-V LAI and the measured data were 0.59, 0.56 and 0.62, respectively, which were 0.57, 0.52 and 0.57 higher than the original data. The root mean square error (ERMSE) were 0.37, 0.31 and 0.14 respectively, which were 1.23, 1.69 and 1.06 lower than the original data. In coniferous forest, the correlation coefficients between the assimilated MODIS, VIIRS and PROBA-V LAI and the measured data were 0.59, 0.49 and 0.56, respectively, which were 0.52, 0.30 and 0.40 higher than the original data. ERMSE were 0.24, 0.28 and 0.19 respectively, which were 1.22, 0.67 and 1.35 lower than the original data. Through the fusion method, the correlation coefficients of LAI in broad-leaved forest and coniferous forest were 0.83 and 0.76 respectively, which were higher than the data after assimilation. ERMSE were 0.15 and 0.13, respectively, which were smaller than the error of the assimilated data. Conclusion Through data assimilation, the accuracy of 3 LAI products is improved, and the fused LAI data has higher accuracy and reliability than the single LAI data after assimilation. [Ch, 4 fig. 2 tab. 30 ref.] -
Key words:
- leaf area index (LAI) /
- MODIS /
- VIIRS /
- PROBA-V /
- reconstruction /
- ensemble Kalman filter (EnKF) /
- data fusion
-
图 1 研究区和样地分布示意图
Figure 1 Geographical location and plot distribution of the study area
图 2 3种LAI时序曲线在阔叶林和针叶林区域的变化情况以及重建效果
Figure 2 Changes and reconstruction effects of 3 LAI time series curves in broad-leaved forest and coniferous forest areas
图 3 同化LAI与实测值的散点验证图
Figure 3 Scatter verification diagram of assimilated LAI and measured value
图 4 融合LAI与实测值的散点验证图
Figure 4 Scatter verification diagram of fused LAI and measured values
表 1 LAI产品数据及基本信息
Table 1. LAI product data and basic information
产品名称 传感器 空间分辨率/m 时间分辨率/d 投影方式 数据来源 MOD15A2H MODIS 500 8 正弦曲线 https://ladsweb.modaps.eosdis.nasa.gov/ VNP15A2H VIIRS 500 8 正弦曲线 https://ladsweb.modaps.eosdis.nasa.gov/ CGLS 300m V1 PROBA-V 300 10 简易圆柱 https://land.copernicus.eu/global/  下载: 导出CSV
下载: 导出CSV
表 2 3种LAI产品在阔叶林和针叶林数据同化前后的精度指标
Table 2. Accuracy indexes of 3 LAI products before and after data assimilation in broad-leaved and coniferous plots
LAI产品 样地 同化前 同化后 R ERMSE EMAE ERE/% R ERMSE EMAE ERE/% MODIS 阔叶林 0.02 1.60 1.37 −23.75 0.59 0.37 0.60 −2.56 针叶林 0.07 1.46 1.30 −0.72 0.59 0.24 0.64 8.34 VIIRS 阔叶林 0.04 2.00 1.71 −28.62 0.56 0.31 0.60 2.29 针叶林 0.17 0.95 0.77 11.55 0.49 0.28 0.72 10.53 PROBA-V 阔叶林 0.05 1.20 1.04 15.24 0.62 0.14 0.86 12.49 针叶林 0.16 1.54 1.39 28.39 0.56 0.19 1.03 20.53
下载: 导出CSV
-
[1] CHEN J M, BLACK T A. Defining leaf area index for non-flat leaves [J]. Plant,Cell &Environment, 1992, 15(4): 421 − 429. [2] MYNENI R B, HOFFMAN S, KNYAZIKHIN Y, et al. Global products of vegetation leaf area and fraction absorbed PAR from year one of MODIS data [J]. Remote Sensing of Environment, 2002, 83(1/2): 214 − 231. [3] WU Wei, SUN Yin, XIAO Kun, et al. Development of a global annual land surface phenology dataset for 1982−2018 from the AVHRR data by implementing multiple phenology retrieving methods [J/OL]. International Journal of Applied Earth Observation and Geoinformation, 2021, 103: 102487[2023-12-10]. doi: 10.1016/j.jag.2021.102487. [4] ZOU Dongxiao, YAN Kai, PU Jiapin, et al. Revisit the performance of MODIS and VIIRS leaf area index products from the perspective of time-series stability [J]. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 2022, 15: 8958 − 8973. [5] VERGER A, SÁNCHEZ-ZAPERO J, WEISS M, et al. GEOV2: improved smoothed and gap filled time series of LAI, FAPAR and FCover 1 km copernicus global land products [J/OL]. International Journal of Applied Earth Observation and Geoinformation, 2023, 123: 103479[2023-12-10]. doi: 10.1016/j.jag.2023.103479. [6] FUSTER B, SÁNCHEZ-ZAPERO J, CAMACHO F, et al. Quality assessment of PROBA-V LAI, fAPAR and fCOVER collection 300 m products of copernicus global land service [J/OL]. Remote Sensing, 2020, 12(6): 1017[2023-12-10]. doi: 10.3390/rs12061017. [7] YAN Kai, PU Jiapin, PARK T, et al. Performance stability of the MODIS and VIIRS LAI algorithms inferred from analysis of long time series of products [J/OL]. Remote Sensing of Environment, 2021, 260: 112438[2023-12-10]. doi: 10.1016/j.rse.2021.112438. [8] MA Han, LIANG Shunlin. Development of the GLASS 250-m leaf area index product (version 6) from MODIS data using the bidirectional LSTM deep learning model [J/OL]. Remote Sensing of Environment, 2022, 273: 112985[2023-12-10]. doi: 10.1016/j.rse.2022.112985. [9] XIAO Zhiqiang, SONG Jinling, YANG Hua, et al. A 250 m resolution global leaf area index product derived from MODIS surface reflectance data [J]. International Journal of Remote Sensing, 2022, 43(4): 1409 − 1429. [10] XING Luqi, LI Xuejian, DU Huaqiang, et al. Assimilating multiresolution leaf area index of Moso bamboo forest from MODIS time series data based on a hierarchical Bayesian network algorithm [J/OL]. Remote Sensing, 2019, 11(1): 56[2023-12-10]. doi: 10.3390/rs11010056. [11] 李雪建, 毛方杰, 杜华强, 等. 双集合卡尔曼滤波LAI同化结合BEPS模型的竹林生态系统碳通量模拟[J]. 应用生态学报, 2016, 27(12): 3797 − 3806. LI Xuejian, MAO Fangjie, DU Huaqiang, et al. Simulating of carbon fluxes in bamboo forest ecosystem using BEPS model based on the LAI assimilated with dual ensemble Kalman filter [J]. Chinese Journal of Applied Ecology, 2016, 27(12): 3797 − 3806. [12] LIU Qiang, LIANG Shunlin, XIAO Zhiqiang, et al. Retrieval of leaf area index using temporal, spectral, and angular information from multiple satellite data [J]. Remote Sensing of Environment, 2014, 145: 25 − 37. [13] 李喜佳, 肖志强, 王锦地, 等. 双集合卡尔曼滤波估算时间序列LAI[J]. 遥感学报, 2014, 18(1): 27 − 44. LI Xijia, XIAO Zhiqiang, WANG Jindi, et al. Dual ensemble Kalman filter assimilation method for estimating time series LAI [J]. Journal of Remote Sensing, 2014, 18(1): 27 − 44. [14] 周红敏, 张国东, 王昶景, 等. 塞罕坝地区高空间分辨率叶面积指数时序估算与变化检测[J]. 遥感学报, 2021, 25(4): 1000 − 1012. ZHOU Hongmin, ZHANG Guodong, WANG Changjing, et al. Time series high-resolution leaf area index estimation and change monitoring in the Saihanba area [J]. Journal of Remote Sensing, 2021, 25(4): 1000 − 1012. [15] 张慧芳, 高炜, 施润和. 基于背景库的高质量LAI时间序列数据重建[J]. 遥感学报, 2012, 16(5): 986 − 999. ZHANG Huifang, GAO Wei, SHI Runhe. Reconstruction of high-quality LAI time-series product based on long-term historical database [J]. Journal of Remote Sensing, 2012, 16(5): 986 − 999. [16] XU Bao, PARK T, YAN Kai, et al. Analysis of global LAI/FPAR products from VIIRS and MODIS sensors for spatio-temporal consistency and uncertainty from 2012−2016 [J/OL]. Forests, 2018, 9(2): 73[2023-12-10]. doi: 10.3390/f9020073. [17] 魏萌, 范文义, 张海军, 等. 4-Scale几何光学模型冠层反射率模拟的空间尺度适用性[J]. 应用生态学报, 2023, 34(3): 605 − 613. WEI Meng, FAN Wenyi, ZHANG Haijun, et al. Spatial scale applicability of canopy reflectance simulation of 4-Scale geometric optical model [J]. Chinese Journal of Applied Ecology, 2023, 34(3): 605 − 613. [18] ROGERS C, CHEN Jingming, CROFT H, et al. Daily leaf area index from photosynthetically active radiation for long term records of canopy structure and leaf phenology [J/OL]. Agricultural and Forest Meteorology, 2021, 304: 108407[2023-12-10]. doi: 10.1016/j.agrformet.2021.108407. [19] WEN Yibo, ZHUANG Linlan, WANG Hezhi, et al. An Automated Hemispherical Scanner for monitoring the leaf area index of forest canopies [J/OL]. Forests, 2022, 13(9): 1355[2023-12-10]. doi: 10.3390/f13091355. [20] 贾玉洁, 刘云根, 杨思林, 等. 面向Sentinel-2A影像的大理市土地利用分类方法适用性研究[J]. 浙江农林大学学报, 2022, 39(6): 1350 − 1358. JIA Yujie, LIU Yungen, YANG Silin, et al. Applicability of land use classification method in Dali City based on Sentinel-2A image [J]. Journal of Zhejiang A&F University, 2022, 39(6): 1350 − 1358. [21] CHEN Jingming, DENG Feng, CHEN Mingzhen. Locally adjusted cubic-spline capping for reconstructing seasonal trajectories of a satellite-derived surface parameter [J]. IEEE Transactions Geoscience and Remote Sensing, 2006, 44(8): 2230 − 2238. [22] ZHAI Huan, HUANG Fang, QI Hang. Generating high resolution LAI based on a modified FSDAF model [J/OL]. Remote Sensing, 2020, 12(1): 150[2023-12-10]. doi: 10.3390/rs12010150. [23] 杨绍钦, 王翔, 许澄, 等. 基于MODIS时间序列数据的竹林地上生物量估算[J]. 浙江农林大学学报, 2022, 39(4): 734 − 741. YANG Shaoqin, WANG Xiang, XU Cheng, et al. Estimation of aboveground biomass of bamboo forest based on MODIS time series data [J]. Journal of Zhejiang A&F University, 2022, 39(4): 734 − 741. [24] JI Jiayi, LI Xuejian, DU Huaqiang, et al. Multiscale leaf area index assimilation for Moso bamboo forest based on Sentinel-2 and MODIS data [J/OL]. International Journal of Applied Earth Observation and Geoinformation, 2021, 104: 102519[2023-12-10]. doi: 10.1016/j.jag.2021.102519. [25] 于颖, 刘敏, 范文义, 等. 基于PROSPECT和4-scale模型的光化学植被指数尺度转换[J]. 北京林业大学学报, 2020, 42(10): 27 − 35. YU Ying, LIU Min, FAN Wenyi, et al. Scale conversion of photochemical reflectance index based on PROSPECT and 4-scale models [J]. Journal of Beijing Forestry University, 2020, 42(10): 27 − 35. [26] 王晓冰, 范文义. 叶面积指数与植被指数饱和点分析及验证[J]. 东北林业大学学报, 2023, 51(9): 83 − 94, 111. WANG Xiaobing, FAN Wenyi. Analysis and verification of saturation point of leaf area index and vegetation index [J]. Journal of Northeast Forestry University, 2023, 51(9): 83 − 94, 111. [27] 周杨凡, 郑兴明, 孙源, 等. GF-1与Sentinel-2在典型农作物LAI估算中的比较[J]. 遥感技术与应用, 2023, 38(3): 599 − 613. ZHOU Yangfan, ZHENG Xingming, SUN Yuan, et al. Comparison of GF-1 and Sentinel-2 for estimation of leaf area index in typical crops [J]. Remote Sensing Technology and Application, 2023, 38(3): 599 − 613. [28] 孟庆博, 刘艳丽, 鞠琴, 等. 基于多源数据同化融合的尼洋河降水时空分布特征[J]. 南水北调与水利科技, 2020, 18(3): 110 − 118. MENG Qingbo, LIU Yanli, JU Qin, et al. Analysis of temporal and spatial distribution characteristics of precipitation based on multi-source data assimilation and fusion in Niyang River Basin [J]. South-to-North Water Transfers and Water Science &Technology, 2020, 18(3): 110 − 118. [29] 周旻悦, 沈润平, 陈俊, 等. 基于像元质量分析和异常值检测的LAI时序数据S-G滤波重建研究[J]. 遥感技术与应用, 2019, 34(2): 323 − 330. ZHOU Minyue, SHEN Runping, CHEN Jun, et al. Filtering and reconstruction of LAI time series data by S-G filter based on pixel quality analysis and outlier detection [J]. Remote Sensing Technology and Application, 2019, 34(2): 323 − 330. [30] BROWN L A, MEIER C, MORRIS H, et al. Evaluation of global leaf area index and fraction of absorbed photosynthetically active radiation products over North America using Copernicus Ground Based Observations for Validation data [J/OL]. Remote Sensing of Environment, 2020, 247: 111935[2023-12-10]. doi: 10.1016/j.rse.2020.111935. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230601
 点击查看大图
点击查看大图

计量
- 文章访问数: 1400
- HTML全文浏览量: 529
- PDF下载量: 39
- 被引次数: 0
