


-
花叶矢竹Pseudosasa japonica f. akebonosuji原产日本,是矢竹的一个栽培变种,属于矮小型混生竹种,是一种优良的珍稀观赏竹种,株高2 m左右[1]。花叶矢竹在自然栽培过程中,叶色会发生变异,有全绿叶、全白叶和绿白相间条纹叶,而且条纹叶中的绿条纹和白条纹的颜色深浅、形状和出现的个数和位置都具有不确定性,还会随着栽培条件的变化而变化。随着转录组测序技术的发展,越来越多的物种转录组被测序。为了了解花叶矢竹不同叶片转录组中的基因表达差异情况,花叶矢竹的不同生长阶段和不同颜色的叶片转录组被测序,这为分析花叶矢竹叶片发育和叶色变异提供了重要基础数据。转座子(TEs或transposons)也可称为转座因子或转座元件,它是一段DNA序列,能从同一染色体的一个位置转移到另一个位置,或者从一条染色体转移到另一条染色体上[2-4]。依据不同的转座机制,可将转座子分为三大类:RNA类转座子(Ⅰ类转座子),DNA类转座子(Ⅱ类转座子)和Helitron转座子(Ⅱ类转座子中的滚筒式转座子)。RNA类转座子也可称为反转录转座子,是通过“复制和粘贴”的机制进行转座。该类转座子主要包括:长末端重复反转录转座子(LTRs),DIRs转座子,PLE转座子,自主的非长末端反转录转座子(LINEs)和非自主的非长末端反转录转座子(SINEs)。DNA转座子是以DNA为中介在基因组中通过“剪切和粘贴”机制进行转座[5]。该类转座子主要包括:TIR类转座子、Crypton转座子超家族、Maverick转座子。虽然Maverick转座子被归类为DNA类转座子,但是它的转座机制是滚环式,属于 Helitron转座子[6],内部含有一个负责链酶结构域的编码基因和复制起始的基序[7]。转座子在宿主基因组跳跃,会插入或脱离目的基因而关闭或恢复基因的表达活性[8-9]。有文献报道[10-11],转座子转座可导致叶色或花色变化,例如转座子Mutator插入到玉米Zea mays叶绿素合成以及光合作用有关基因中,引起玉米白化突变体;研究表明:转座子nDart1插入水稻Oryza sativa叶绿体蛋白酶基因OsClpP5,使基因被破坏形成了叶色为浅黄色的pyl-v突变株[12];康乃馨Dianthus caryophyllus花色由紫色变为深粉色就是由一个有活性的hAT型转座子Tdic101插入到F3′H(flavonoid 3′-hydroxylase,类黄酮3′-羟化酶)基因引起的[13];日本Iida实验室研究发现,蓝色花的矮牵牛Petunia hybrida突变为紫色花是由于转座子Tpn4插入到Pr基因,影响了该基因的正常表达,使之缺乏Na+/H+逆向转运蛋白,因而液泡内的pH值降低,突变株无法产生正常的亮蓝色,因此,蓝色的花冠檐上产生紫色的斑痕,形成嵌合花色[14]。本研究利用花叶矢竹不同颜色和不同发育阶段的10种叶片转录组数据,分析花叶矢竹不同颜色和不同发育阶段叶片的转座子种类、数量及其选择性表达特性等,这将为探索转座子是否参与花叶矢竹叶色变异奠定一定的基础。
-
本实验室具有花叶矢竹10种叶片材料的转录组数据。这10种叶片材料是根据叶片的颜色和长度来确定的。首先选择了2种极端颜色的叶片,即全绿叶(G)和全白叶(A);又分别从这2种叶片中选择了卷曲叶最里面一层叶芽的5个不同发育阶段,0.1~1.0 cm长的刚刚抽出的叶芽为第1发育阶段(G1和A1),叶芽随时间逐渐发育伸长,第2发育阶段(G2和A2)选择的叶芽长度是1.0~5.0 cm,长度5.0~7.0 cm的叶芽为第3发育阶段(G3和A3),长度7.0~10.0 cm的叶芽为第4发育阶段(G4和A4),长度10.0~12.0 cm的叶芽为第5发育阶段(G5和A5)。
-
将拼接后的绿叶5个发育阶段和白叶5个发育阶段的转录组序列提交到repeatmasker网站,利用Protein-based RepeatMasking预测其中的转座子序列(http://www.repeatmasker.org/cgi-bin/RepeatProteinMaskRequest),对预测到的转座子通过e-值进行筛选(e<0.000 01),根据不同转座子的类型,将筛选过的转座子进行分类分析。
-
按照转座子类型,将从转录组中预测到的每条转座子的FPKM值(expected number of fragments per kilobase of transcript sequence per millions base pairs sequenced,即每百万测序碱基中每千个转录子测序碱基中所包含的测序片段数)合并,除以每种类型转座子的数目,得到每种类型转座子的平均FPKM值。
转座子根据转座机制可分为RNA类转座子(Ⅰ类转座子)、DNA类转座子(Ⅱ类转座子)和Helitron转座子(Ⅱ类转座子中的滚筒式转座子),根据上述方法算出样本G1,G2,G3,G4,G5和A1,A2,A3,A4,A5各转录组RNA类转座子的FPKM平均值、DNA类转座子的FPKM平均值和Helitron转座子的FPKM平均值以及总的转座子FPKM平均值。
-
从预测筛选出的转座子中随机挑选10个转座子序列,用软件Primer 5设计引物,由生工生物工程(上海)股份有限公司合成。引物详细信息见表 1。
表 1 转座子验证引物
Table 1. Primers of transposons verification
转座子类型 上游引物(5'→3') 下游引物(5'→3') DNA/Academ GCTAAAGGCGGTAAGT CATCTGTGGCAATCAGTA DNA/EnSpm GGGCATACTAATCGTAAAG CCTCAAGCGTCAGTGTCT DNA/hAT CTCAGTCACCTCCTCTT CATCAATCTATCGCCAC DNA/MulE TGATACGGTCACTCTTACT ATATGGATCTGGCTTTT DNA/P CAGTCTTGGTCGCCTCG CGCCTCCATCTCCCTGT DNA/PIF-Harbing GACTTGGGCATGAGCACG CGTCGCCAAGGAGGACA LINE GCTCGATGGATTTAGTGG GCCATCATGGAGCACAT LTR/Copia CGGGTCAAGATAGAGG CACCGTCATCCGTAAT LTR/ERV TGCGCTGAAGCATTCG GCACGGTGGCATCAAG RC/Helitron ATGGTAGCCTGGATTTG CCTTTGATCTCCCTGAA 参照Trizol试剂(TaKaRa公司,日本)方法提取上述绿叶和白叶各5个发育阶段叶片的总RNA,然后用TaKaRa公司的反转录试剂盒(PrimeScriptTM Ⅱ 1st Strand cDNA Synthesis Kit)将提取的RNA反转录为cDNA。以cDNA作为模板,基因PheACT2为内参[15],参照SYBR Green I Master(TaKaRa公司,日本)说明书进行荧光定量聚合酶链式反应(qRT-PCR),20.0 μL的反应体系包括模板cDNA 2.0 μL,上下游引物各0.8 μL,灭菌的双蒸水6.4 μL,SYBR® Premix Ex TaqⅡ 10.0 μL。反应条件为:95 ℃,30 s;95 ℃,5 s,55 ℃,20 s,72 ℃ 20 s,共40个循环。
-
按样本转录组中的转座子FPKM比值大于4,差值大于2的原则,除了筛选出在绿叶5个发育阶段之间和白叶5个发育阶段之间显著性差异表达的转座子序列,还筛选出白叶和绿叶各对应生长时期相比显著性差异表达的转座子序列。
-
根据花叶矢竹叶片颜色的不同,可将其叶片分成3种形态:全绿叶、全白叶和花叶。全绿叶是整张叶片都是绿色的(图 1A);整张叶片都是白色的叶片是全白叶(图 1B);花叶是一张叶片上既有绿色也有白色,绿白相间,但是有的花叶绿色部分多于白色部分,就形成了白条纹叶,而有的花叶白色部分多于绿色部分,就形成了绿条纹叶(图 1C)。

图 1 花叶矢竹的叶片形态
Figure 1. Leaf form of Pseudosasa japonica f. akebonosuji
-
从转录组中共预测到57 890个转座子序列,筛选e值小于0.000 01的序列,得到1 627条转座子序列。根据不同转座子的类型,将这1 627个转座子进行分类分析,可将其分为3个大类,包括1 205个RNA转座子,占总转座子的74.06%;380个DNA转座子(23.36%)和42个Helitron转座子(2.58%)。RNA转座子的数量远远多于DNA转座子数量。共有22种类型转座子,其中RNA转座子9种,包括LINE,LTR/Copia,LTR/DIRS,LTR/ERV,LTR/Gypsy,LTR/Lenti,LTR/Ngaro,LTR/Pao和Other/subtelome;DNA转座子12种,包括DNA/Academ,DNA/Chapaev,DNA/EnSpm,DNA/Crypton,DNA/Ginger,DNA/hAT,DNA/Maverick,DNA/MuLE,DNA/Novosib,DNA/P,DNA/PIF-Harbing和DNA/Sola。Helitron转座子虽然被归为DNA类转座子,但由于它的转座机制是滚环式,所以单列为一类(表 2)。RNA转座子中数量最多的是LTR/Copia,占总转座子数的25.38%,其中转座子LINE,LTR/Gypsy和LTR/Pao也占了很高比例,分别为16.72%,15.18%和9.96%;DNA转座子中数量最多的是DNA/hAT(8.85%)。
表 2 花叶矢竹转录组中的转座子类型及数量
Table 2. Super-family and abundance of transposons in transcriptome of Pesudosasajaponica f. akebonosuji
转座子类型 数量/个 所占比例/% DNA/Academ 35 2.15 DNA/Chapaev 10 0.61 DNA/EnSpm 38 2.34 DNA/Crypton 1 0.06 DNA/Einger 1 0.06 DNA/hAT 144 8.85 DNA/Maverick 4 0.25 DNA/MuLE 77 4.73 DNA/Novosib 5 0.31 DNA/P 13 0.80 DNA/PIF-Harbing 51 3.13 DNA/Sola 1 0.06 LINE 272 16.72 LTR/Copia 413 25.38 LTR/DIRS 2 0.12 LTR/ERV 96 5.90 LTR/Gypsy 247 15.18 LTR/Lenti 7 0.43 LTR/Ngaro 3 0.18 LTR/Pao 162 9.96 Other/subtelome 3 0.18 RC/Helitron 42 2.58 DNA 380 23.36 RNA 1 205 74.06 RC 42 2.58 总和 1 627 100.00 -
转座子的表达丰度是检测转座子转座活性的一个重要指标。从表 3中可见:转座子总表达丰度平均值在绿叶5个不同发育阶段中上下浮动,而在白叶5个不同发育阶段中是随着发育逐渐减小的。
表 3 花叶矢竹绿叶和白叶各5种发育阶段中不同转座子的表达丰度平均值
Table 3. Different transposons’ average expression level in five developmental stages of green leaves and albino leaves of Pseudosasa japonica f. akebonosuji
转座子类型 绿叶 白叶 G1 G2 G3 G4 G5 A1 A2 A3 A4 A5 DNA/Academ 46.52 33.68 36.33 34.10 29.07 43.16 30.79 29.12 23.75 22.28 DNA/Chapaev 21.47 16.41 20.69 16.78 16.34 19.54 18.70 18.59 18.95 16.06 DNA/EnSpm 31.57 38.66 35.34 35.59 35.32 33.11 38.02 36.57 38.01 43.30 DNA/Crypton 20.56 12.82 18.50 10.81 12.45 21.71 9.73 16.11 15.10 10.94 DNA/*inger 18.51 18.03 19.62 22.99 28.90 29.14 22.05 28.99 20.30 26.52 DNA/hAT 34.47 29.66 31.55 29.45 27.34 32.82 28.65 29.03 16.64 25.12 DNA/Maverick 6.61 15.41 13.54 16.72 18.05 8.68 18.75 20.08 79.73 26.83 DNA/Mu 20.57 14.59 16.43 14.62 13.68 19.66 15.86 14.67 14.09 12.72 DNA/NovosibLE 21.76 21.83 19.78 21.47 20.86 21.18 22.20 24.14 24.06 19.88 DNA/P 18.02 43.16 40.24 51.92 61.69 20.30 49.31 57.43 65.97 74.25 DNA/PIF-Harbing 31.55 53.37 24.37 44.78 84.14 60.80 44.56 37.59 38.43 22.33 DNA/Sola 275.37 176.92 204.89 235.67 167.55 306.03 149.75 117.70 73.31 84.45 LINE 39.40 37.36 35.91 35.12 32.87 38.14 34.72 34.53 32.73 31.33 LTR/Copia 30.94 31.35 28.61 30.40 32.18 32.26 30.99 28.86 27.82 31.32 LTR/DIRS 14.95 19.80 11.65 16.11 14.56 14.07 15.86 15.57 16.93 19.17 LTR/ERV 37.72 38.19 35.24 36.87 36.74 38.34 36.22 35.74 32.58 32.37 LTR/Gypsy 31.95 28.19 28.56 28.29 28.90 32.60 28.88 27.66 26.89 25.01 LTR/Lenti 105.03 84.54 83.28 62.48 52.75 94.82 67.84 64.05 54.81 37.93 LTR/Ngaro 19.82 12.24 13.34 13.18 9.36 14.90 10.01 11.53 9.86 6.67 LTR/Pao 134.25 154.24 123.43 124.61 121.10 147.02 131.70 118.89 112.61 106.39 Other/subtelome 60.09 37.78 35.71 35.13 35.56 58.28 39.96 34.65 29.86 29.97 RC/Helitron 34.89 25.38 31.60 25.17 20.74 29.50 23.17 23.74 21.20 16.38 DNA 31.23 31.70 28.39 30.69 34.92 34.75 30.44 29.38 28.27 26.28 RNA 47.94 49.38 43.79 44.35 44.02 49.92 45.52 42.70 40.61 40.16 RC 34.89 25.38 31.60 25.17 20.74 29.50 23.17 23.74 21.20 16.38 总和 43.70 44.63 39.88 40.66 41.30 45.85 41.42 39.10 37.23 36.31 RNA转座子表达丰度平均值高于DNA转座子,RC/Helitron型转座子表达丰度平均值最小;不同类型转座子的表达丰度平均值也是不同的,RNA转座子9种类型中,LTR/Pao型转座子表达丰度平均值最高,在所有类型转座子中表达量居第2位;DNA转座子12种类型中,DNA/sola型转座子表达量显著高于其他类型转座子,并居于所有类型转座子中的第1位。LTR/Pao型转座子和DNA/sola型转座子的表达丰度平均值在绿叶和白叶5个发育阶段的趋势与转座子总表达丰度平均值的趋势相似。表达丰度平均值相对较高的转座子类型还有LTR/Lenti。但无论在绿叶还是白叶的5个发育阶段中,其表达丰度平均值的趋势均随着叶片发育成熟逐渐减小。
-
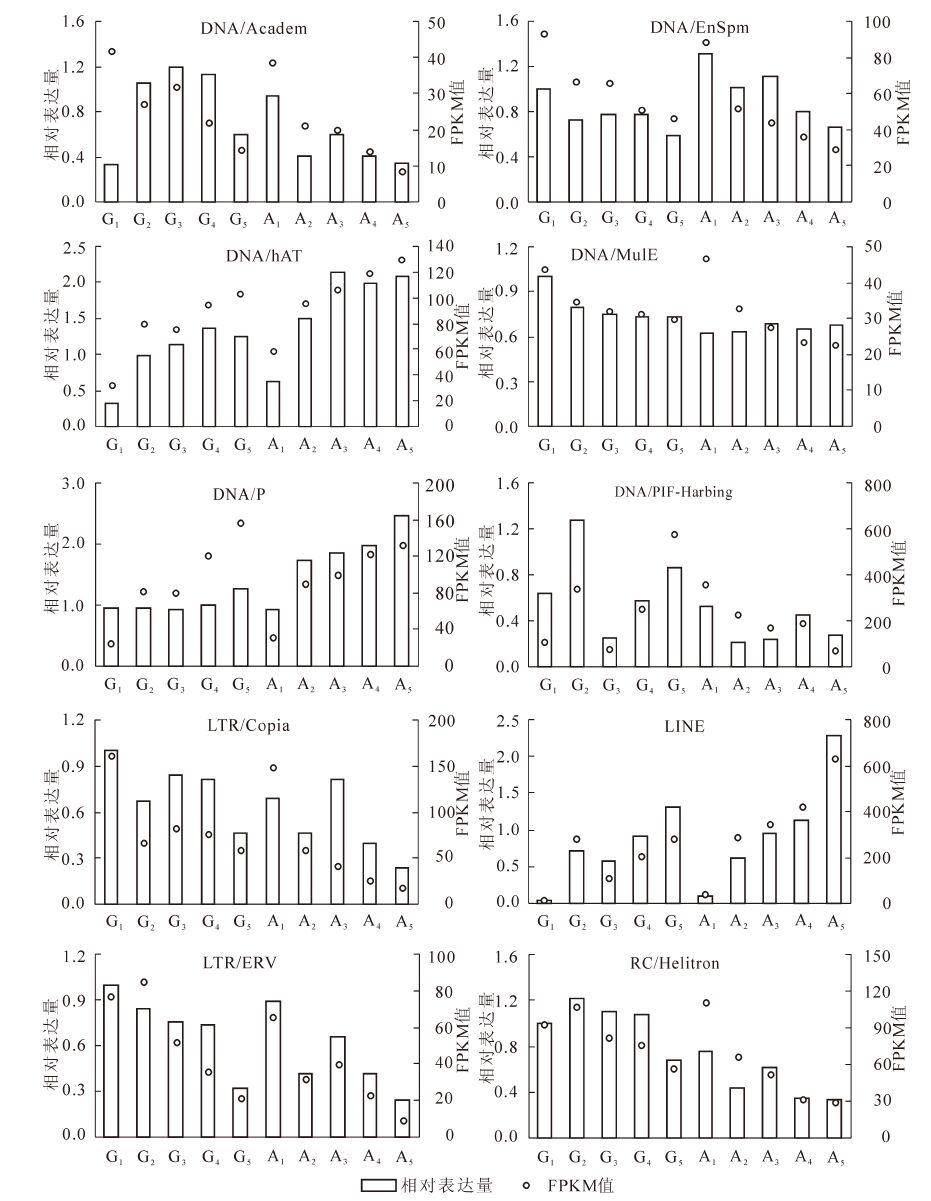
从1 627条转座子序列中,随机选出10条转座子序列进行荧光定量qRT-PCR验证,结果如图 2。可见,qRT-PCR所得到的转座子在各个样本中的相对表达量值,与其转录组数据的FPKM值的趋势基本上是一致的。

图 2 转座子的相对表达量与其相对应的FPKM值
Figure 2. Relative expression level and FPKM value of transposons
-
从1 627条转座子中,按转座子FPKM比值大于4,差值大于2的原则,筛选出绿叶5个发育阶段之间、白叶5个发育阶段之间以及绿叶和白叶对应发育阶段之间表达显著性差异的转座子序列。绿叶5个发育阶段之间表达存在显著性差异的有194个转座子,白叶5个发育阶段之间表达存在显著性差异的有181个转座子,绿叶和白叶相对应5个发育阶段之间表达存在显著性差异的有107个转座子。
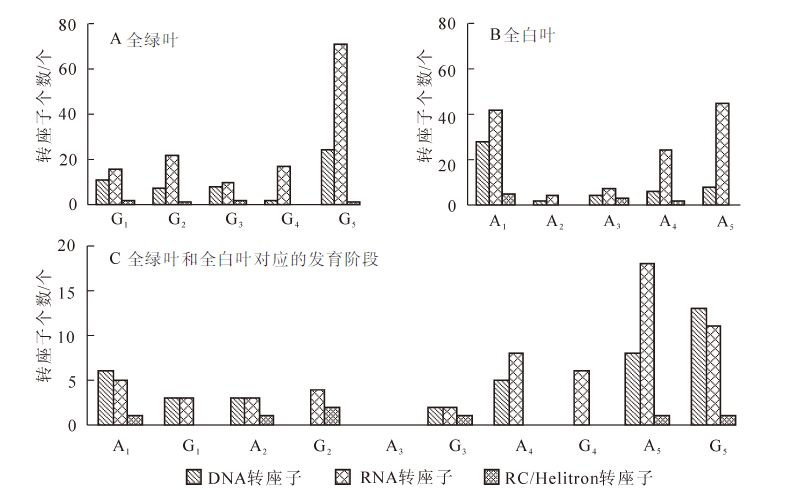
分别对绿叶中194个显著性差异表达转座子进行分析,在每个发育阶段高表达的转座子中既有DNA转座子也有RNA转座子(图 3A)。总的来看,G5发育阶段较高表达量的转座子明显多于其他发育阶段,RNA转座子明显多于DNA转座子。G1发育阶段,有29个转座子较高表达,有RNA转座子16个,DNA转座子11个以及2个RC/Helitron转座子。G2发育阶段有22个RNA转座子、7个DNA转座子和1个RC/Helitron转座子较高表达。G3发育阶段有10个RNA转座子、8个DNA转座子和2个RC/Helitron转座子较高表达,共20个。G4发育阶段共有17个RNA转座子、2个DNA转座子较高表达。G5发育阶段有71个RNA转座子、24个DNA转座子、1个RC/Helitron转座子较高表达,共96个,占绿叶中显著性差异表达转座子总数的49.48%。

图 3 花叶矢竹不同发育阶段显著性差异表达转座子的类型和数量
Figure 3. Type and number of transposons with significant difference expression level among different development stages of leaves of Pseudosasa japonic f. akebonosuji
分别对白叶中181个显著性差异表达转座子进行分析,A1发育阶段高表达的转座子多于其他的发育阶段(图 3B)。A1阶段有75个高表达转座子,占白叶中显著性差异表达转座子总数的41.44%,其中RNA转座子有42个,DNA转座子有28个,RC/Helitron转座子有5个。A2阶段只有6个高表达的转座子,RNA转座子有4个,DNA转座子有2个。A3阶段高表达转座子有14个,RNA转座子有7个,DNA转座子有4个。A4发育阶段有32个高表达转座子,24个RNA转座子,DNA转座子只有6个。A5发育阶段有53个高表达转座子,无RC/Helitron 转座子;RNA转座子有45个,8个DNA转座子。
绿叶和白叶的各对应的发育时期相比较,有107个转座子表达有显著性差异,依然是RNA转座子数量远多于DNA转座子。总体来说白叶中较高表达量的转座子数量多于绿叶。第1发育阶段白叶中较高表达的转座子数量有12个,绿叶中仅有6个较高表达转座子;第2发育阶段白叶有7个较高表达的转座子,绿叶中有6个;第3发育阶段白叶中没有较高表达的转座子,而绿叶中只有5个;第4发育阶段白叶中有13个较高表达的转座子,绿叶中有6个;第5发育阶段,白叶中的较高表达的转座子数为27个,绿叶中有25个较高表达的转座子(图 3C)。
-
自美国细胞遗传学家McCLINTOCK[16]在研究玉米遗传的过程中发现转座子起,越来越多的转座子被发现和研究。转座子在宿主基因组跳跃,插入或脱离目的基因从而关闭或恢复基因的表达活性[8-9],可见转座子在调控基因表达方面发挥着很大作用。本研究比对花叶矢竹10个转录组数据,分析了花叶矢竹中1 627条共22类转座子序列。结果表明:花叶矢竹转录组中的转座子含量丰富,种类繁多,其中RNA转座子占74.06%,DNA转座子占23.36%,Helitron转座子只占2.58%。RNA转座子中数量最多的类型是转座子LTR/Copia占25.38%,DNA转座子中数量最多的类型是转座子DNA/hAT。花叶矢竹转录组数据中RNA转座子的数量明显多于DNA转座子,并且转座子LTR/Copia明显多于其他类型的转座子。这与其他研究相似,即RNA转座子在生物中广泛存在[17],其中LTR/Copia型转座子在植物中分布最广[18]。
基因的选择性表达贯穿于生物体整个生长发育阶段,既具有组织特异性也具有时间特异性[19-21]。经荧光定量PCR验证,转座子在花叶矢竹中的表达也具有时间和空间的特异性(图 2)。本研究对花叶矢竹全绿叶和全白叶5个发育阶段之间显著性差异表达的转座子表达模式分析表明:无论在哪个发育阶段,RNA转座子数量都多于DNA转座子,不同发育时期高表达的转座子类型基本上不同。绿叶中第5发育阶段有较高表达量的转座子明显多于其他发育阶段,说明叶片后期成熟期间有大量的转座子表达,这些高表达转座子可能参与了叶片成熟过程;而白叶中高表达量的转座子主要出现在第1发育阶段,这些在白叶第1发育阶段高表达的转座子可能参与了白叶早期发育过程[10]。
通过对花叶矢竹全绿叶和全白叶相对应的5个发育阶段显著性差异表达的转座子表达模式分析,发现总体白叶中的高表达转座子数量高于绿叶中的转座子。在白叶中高表达的转座子可能参与了花叶矢竹叶色变异过程,也可能是逆境胁迫导致转座子转座激活。白叶缺乏叶绿素,无法进行正常的光合作用,再加上白叶的透光率较高,强光可能会导致白叶出现光损伤和光过度氧化,使白叶发育受到一定的胁迫[22],因此,这种胁迫可能导致白叶中大量转座子的转座激活。事实上,已有大量的研究表明,转座子在逆境胁迫下转座频率大大增加[23-24],如生活在黄河入海口的野大豆Glycine soja盐渍种群有多拷贝及多样性的Gypsy类转座子[25]。
Transposons expression analysis of transcriptome in Pseudosasa japonica f. akebonosuji
-
摘要: 以花叶矢竹Pseudosasa japonica f.akebonosuji 10种不同颜色和不同发育阶段的叶片转录组数据为基础,调查了花叶矢竹中转座子的种类、数量以及选择性表达特性。结果表明:花叶矢竹转录组中转座子类型丰富、数量繁多,其中RNA转座子明显多于DNA转座子,转座子LTR/Copia类型数量最多。绿叶的5个发育阶段中,转座子主要在第5发育阶段高表达,表明这些转座子可能参与了花叶矢竹叶片成熟过程。而在白叶5个发育阶段中,转座子主要在第1发育阶段高表达;对绿叶和白叶5个相对应的发育阶段分析表明,白叶中高表达的转座子多于绿叶,表明这些转座子可能参与了花叶矢竹叶色变异过程,也可能是逆境胁迫导致转座子转座激活。图3表3参25Abstract: To study the super-family, abundance, and expression patterns of transposons in Pseudosasa japonica f. akebonosuji, transcriptome data from 10 kinds of different colors and different leaf developmental stages were assembled. Transposons were identified by Protein-based RepeatMasking. Then a comparison analysis of transposons for the five corresponding developmental stages of green leaves and albino leaves was conducted. Results showed many different super-families of transposons in transcriptome with more RNA (74.06%) than DNA (23.36%) transposons and with LTR/Copia transposons (25.38%) the most abundant for all super-families. Among the five developmental stages of green leaves, the fifth developmental stage had the most high-level expressed transposons (49.48%). Also, among the five developmental stages of albino leaves, the first developmental stage had the most high-level expressed transposons. The comparison analysis of the five corresponding developmental stages of green and albino leaves showed more high-level expressed transposons in albino leaves (55.14%). Thus, in green leaves of P. japonica f. akebonosuji, transposons could be involved with leaf maturation; also to lay a foundation for whether transposons regulate leaf color variation, the green and albino leaf comparison revealed that transposons could influence leaf color variation with high expression of transposons due to stress from abnormal photosynthesis.[Ch, 3 fig. 3 tab. 25 ref.]
-
Key words:
- botany /
- Pseudosasa japonica f. akebonosuji /
- transposons /
- expression patterns
-
图 2 转座子的相对表达量与其相对应的FPKM值
Figure 2 Relative expression level and FPKM value of transposons
图 3 花叶矢竹不同发育阶段显著性差异表达转座子的类型和数量
Figure 3 Type and number of transposons with significant difference expression level among different development stages of leaves of Pseudosasa japonic f. akebonosuji
表 1 转座子验证引物
Table 1. Primers of transposons verification
转座子类型 上游引物(5'→3') 下游引物(5'→3') DNA/Academ GCTAAAGGCGGTAAGT CATCTGTGGCAATCAGTA DNA/EnSpm GGGCATACTAATCGTAAAG CCTCAAGCGTCAGTGTCT DNA/hAT CTCAGTCACCTCCTCTT CATCAATCTATCGCCAC DNA/MulE TGATACGGTCACTCTTACT ATATGGATCTGGCTTTT DNA/P CAGTCTTGGTCGCCTCG CGCCTCCATCTCCCTGT DNA/PIF-Harbing GACTTGGGCATGAGCACG CGTCGCCAAGGAGGACA LINE GCTCGATGGATTTAGTGG GCCATCATGGAGCACAT LTR/Copia CGGGTCAAGATAGAGG CACCGTCATCCGTAAT LTR/ERV TGCGCTGAAGCATTCG GCACGGTGGCATCAAG RC/Helitron ATGGTAGCCTGGATTTG CCTTTGATCTCCCTGAA  下载: 导出CSV
下载: 导出CSV
表 2 花叶矢竹转录组中的转座子类型及数量
Table 2. Super-family and abundance of transposons in transcriptome of Pesudosasajaponica f. akebonosuji
转座子类型 数量/个 所占比例/% DNA/Academ 35 2.15 DNA/Chapaev 10 0.61 DNA/EnSpm 38 2.34 DNA/Crypton 1 0.06 DNA/Einger 1 0.06 DNA/hAT 144 8.85 DNA/Maverick 4 0.25 DNA/MuLE 77 4.73 DNA/Novosib 5 0.31 DNA/P 13 0.80 DNA/PIF-Harbing 51 3.13 DNA/Sola 1 0.06 LINE 272 16.72 LTR/Copia 413 25.38 LTR/DIRS 2 0.12 LTR/ERV 96 5.90 LTR/Gypsy 247 15.18 LTR/Lenti 7 0.43 LTR/Ngaro 3 0.18 LTR/Pao 162 9.96 Other/subtelome 3 0.18 RC/Helitron 42 2.58 DNA 380 23.36 RNA 1 205 74.06 RC 42 2.58 总和 1 627 100.00
下载: 导出CSV
表 3 花叶矢竹绿叶和白叶各5种发育阶段中不同转座子的表达丰度平均值
Table 3. Different transposons’ average expression level in five developmental stages of green leaves and albino leaves of Pseudosasa japonica f. akebonosuji
转座子类型 绿叶 白叶 G1 G2 G3 G4 G5 A1 A2 A3 A4 A5 DNA/Academ 46.52 33.68 36.33 34.10 29.07 43.16 30.79 29.12 23.75 22.28 DNA/Chapaev 21.47 16.41 20.69 16.78 16.34 19.54 18.70 18.59 18.95 16.06 DNA/EnSpm 31.57 38.66 35.34 35.59 35.32 33.11 38.02 36.57 38.01 43.30 DNA/Crypton 20.56 12.82 18.50 10.81 12.45 21.71 9.73 16.11 15.10 10.94 DNA/*inger 18.51 18.03 19.62 22.99 28.90 29.14 22.05 28.99 20.30 26.52 DNA/hAT 34.47 29.66 31.55 29.45 27.34 32.82 28.65 29.03 16.64 25.12 DNA/Maverick 6.61 15.41 13.54 16.72 18.05 8.68 18.75 20.08 79.73 26.83 DNA/Mu 20.57 14.59 16.43 14.62 13.68 19.66 15.86 14.67 14.09 12.72 DNA/NovosibLE 21.76 21.83 19.78 21.47 20.86 21.18 22.20 24.14 24.06 19.88 DNA/P 18.02 43.16 40.24 51.92 61.69 20.30 49.31 57.43 65.97 74.25 DNA/PIF-Harbing 31.55 53.37 24.37 44.78 84.14 60.80 44.56 37.59 38.43 22.33 DNA/Sola 275.37 176.92 204.89 235.67 167.55 306.03 149.75 117.70 73.31 84.45 LINE 39.40 37.36 35.91 35.12 32.87 38.14 34.72 34.53 32.73 31.33 LTR/Copia 30.94 31.35 28.61 30.40 32.18 32.26 30.99 28.86 27.82 31.32 LTR/DIRS 14.95 19.80 11.65 16.11 14.56 14.07 15.86 15.57 16.93 19.17 LTR/ERV 37.72 38.19 35.24 36.87 36.74 38.34 36.22 35.74 32.58 32.37 LTR/Gypsy 31.95 28.19 28.56 28.29 28.90 32.60 28.88 27.66 26.89 25.01 LTR/Lenti 105.03 84.54 83.28 62.48 52.75 94.82 67.84 64.05 54.81 37.93 LTR/Ngaro 19.82 12.24 13.34 13.18 9.36 14.90 10.01 11.53 9.86 6.67 LTR/Pao 134.25 154.24 123.43 124.61 121.10 147.02 131.70 118.89 112.61 106.39 Other/subtelome 60.09 37.78 35.71 35.13 35.56 58.28 39.96 34.65 29.86 29.97 RC/Helitron 34.89 25.38 31.60 25.17 20.74 29.50 23.17 23.74 21.20 16.38 DNA 31.23 31.70 28.39 30.69 34.92 34.75 30.44 29.38 28.27 26.28 RNA 47.94 49.38 43.79 44.35 44.02 49.92 45.52 42.70 40.61 40.16 RC 34.89 25.38 31.60 25.17 20.74 29.50 23.17 23.74 21.20 16.38 总和 43.70 44.63 39.88 40.66 41.30 45.85 41.42 39.10 37.23 36.31
下载: 导出CSV
-
[1] 杨海芸,王晓芹,张宁,等. 日本花叶矢竹组织培养与叶色变异研究[J]. 竹子研究汇刊,2010,29(4):15-20. YANG Haiyun, WANG Xiaoqin, ZHANG Ning, et al. Tissue culture and leaf color variation of Pseudosasa japonica f. akebonosuji[J]. J Bamboo Res, 2010, 29(4):15-20. [2] McCLINTOCK B. Chromosome organization and genic expression[J]. Cold Spring Harb Symp Quant Biol, 1951, 16(2):13-47. [3] McCLINTOCK B. The significance of responses of the genome to challenge[J]. Science, 1984, 226(4676):792-801. [4] 高东迎,何冰,孙立华. 水稻转座子研究进展[J]. 植物学通报,2007,24(5):667-676. GAO Dongying,HE Bing,SUN Lihua. Research advances on transposable elements in rice[J]. Chin Bull Bot, 2007, 24(5):667-676. [5] 何承忠,陈宝昆,江涛,等. 植物转座子的研究与应用[J]. 西南农业学报,2004,17(3):399-403. HE Chengzhong, CHEN Baokun, JIANG Tao, et al. Application and research on transposons of plants[J]. Southwest China J Agric Sci, 2004, 17(3):399-403. [6] KAPITONOV V V,JURKA J. Self-synthesizing DNA transposons in eukaryotes[J]. Proc Natl Acad Sci, 2006, 103(12):4540-4545. [7] 马赑. 桑树全基因组转座子的鉴定及特征分析[D]. 重庆:西南大学,2014. MA Bi. Genome-wide Identification and Characterization of Transposable Elements in Mulberry (Morus notabilis)[D]. Chongqing:Southwest University, 2014. [8] 李军,李洪清,吴萍,等. 转座子在观赏植物嵌合花色形成中的应用[J]. 植物生理学通讯,2005,41(4):542-546. LI Jun, LI Hongqing, WU Ping, et al. Application of transposons in making variegated flower color in ornamental plants[J]. Plant Physiol Commun, 2005, 41(4):542-546. [9] 刘晓红,李叶峰,彭丽媛,等. 裂叶牵牛突变体的研究[J]. 安徽农业科学,2011,39(28):17214-17216. LIU Xiaohong, LI Yefeng, PENG Liyuan, et al. Study on the mutants of Pharbitis nil[J]. J Anhui Agric Sci, 2011, 39(28):17214-17216. [10] 王婷婷,翟立红,苏旭,等. Mutator诱导的玉米白化突变体插入位点的遗传分析及代谢途径的构建[J]. 中国农业科学,2010,43(22):4571-4578. WANG Tingting, ZHAI Lihong, SU Xu, et al. Genetic analysis and construction of metabolic pathway by mutator-mediated albino mutatant insertion sites in Zea mays L.[J]. Sci Agric Sin, 2010, 43(22):4571-4578. [11] SLOTKIN R,FREELING M,LISCH D. Mu killer causes the heritable inactivation of the mutator family of transposable elements in Zea mays[J]. Genetics, 2003, 165(2):781-797. [12] TSUGANE K, MAEKAWA M, TAKAGI K, et al. An active DNA transposon nDart causing leaf variegation and mutable dwarfism and its related elements in rice[J]. Plant J Cell Mol Boil, 2006, 45(1):46-57. [13] MOMOSE M, NAKAYAMA M, ITOH Y, et al. An activehAT transposable element causing bud mutation of carnation by insertion into the flavonoid 3'-hydroxylase gene[J]. Eur J Clin Invest, 1974, 4(4):247-252. [14] 李美茹,李洪清,孙梓健,等. 影响蓝色花着色的因素[J]. 植物生理学通讯,2003,39(1):51-55. LI Meiru, LI Hongqing, SUN Zijian, et al. Factors of influencing blue flower color[J]. Plant Physiol Commun, 2003, 39(1):51-55. [15] 齐飞艳,胡陶,彭镇华,等. 毛竹实时荧光定量PCR内参基因的筛选及成花基因PheTFL1表达分析[J]. 西北植物学报,2013,33(1):48-52. QI Feiyan, HU Tao, PENG Zhenhua, et al. Screening of reference genes used in qRT-PCR and expression analysis of PheTFL1 gene in moso bamboo[J]. Acta Bot Boreal-Occident Sin, 2013, 33(1):48-52. [16] McCLINTOCK B. The origin and behavior of mutable loci in maize[J]. Proc Nat Acad Sci, 1950, 36(6):344-355. [17] WICKER T, SABOT F, HUA-VAN A, et al. A unified classification system for eukaryotic transposable elements[J]. Nat Rev Genet, 2007, 8(12):973-982. [18] 仲晓芳. 菰(Zizania latifolia)Ty1/copia-like反转座子反转座酶序列的克隆和分析[D]. 长春:东北师范大学,2005. ZHONG Xiaofang. Isolation and Characterization of Ty1/copia-like Retrotransposons of Wild Rice (Zizania latifolia)[D]. Changchun:Northeast Normal University, 2005. [19] 郭红媛,贾举庆,张莉,等. 燕麦EST数据库中Ty1-copia和Ty3-gypsy型反转录转座子的频数分析[J]. 中国农业大学学报,2014,19(4):23-30. GUO Hongyuan, JIA Juqing, ZHANG Li, et al. Frequencies of Ty1-copia and Ty3-gypsy retroelements within the Avena EST database[J]. J China Agric Univ, 2014, 19(4):23-30. [20] HIROCHIKA H,SUGIMOTO K,OTSUKI Y,et al. Retrotransposons of rice involved in mutations induced by tissue culture[J]. Proc Nat Acad Sci, 1996, 93(15):7783-7788. [21] HIROCHIKA H. Activation of tobacco retrotransposons during tissue culture[J]. Embo J, 1993, 12(6):2521-2528. [22] 杨海芸. 花叶矢竹叶色变异机理研究[D]. 北京:北京林业大学,2015. YANG Haiyun. Study on Mechanism of Leaf Color Variation of Pseudosasa japonica f. akebonosuji[D]. Beijing:Beijing Forestry University, 2015. [23] DOMINGUES D S,CRUZ G M,METCALFE C J,et al. Analysis of plant LTR-retrotransposons at the fine-scale family level reveals individual molecular patterns[J]. BMC Gen, 2012, 13(4):1-14. [24] GRANDBASTIEN M A. Activation of plant retrotransposons under stress conditions[J]. Trends Plant Sci, 1998, 3(5):181-187. [25] 胡志昂,姜国强,邓馨,等. 野大豆种群转座子和转录因子的多样性和分子适应[J]. 植物生态学报,2007,31(5):952-959. HU Zhiang, JIANG Guoqiang, DENG Xin, et al. Molecular adaptation through diversity of retrotransposons and transcriptional factors in populations of wild soybean (Glycine soja)[J]. J Plant Ecol, 2007, 31(5):952-959. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2016.06.003
 点击查看大图
点击查看大图

计量
- 文章访问数: 3214
- HTML全文浏览量: 675
- PDF下载量: 518
- 被引次数: 0
