




 下载:
下载:
-
竹子属于禾本科Poaceae竹亚科Bambusoideae,热带和亚热带为其主要分布区。竹亚科又包括了木本的竹类和草本的竹类[1]。世界上的竹子约有88属1 400余种[2],其中草本竹类有28属,180余种。绝大多数草本竹类分布在拉丁美洲,非洲有5属,亚洲和大洋洲共有3属,中国仅台湾有1种草本竹即Leptaspis formosana,其他地区尚未见报道[3]。竹子的开花是一个复杂的生理现象,其开花类型多样,开花周期较长且难以预测。不同竹种的开花周期不同,即使同一竹种,不同地方的开花周期也不是固定不变的。另外,大多数竹种为一生一次性开花植物,随着开花结实,植株衰老死亡[4-7]。竹子的开花特性不仅给竹种分类鉴定、杂交育种等工作带来困难,竹林的大面积开花衰老还导致严重的经济损失和生态环境的破坏[8]。所以竹子的开花现象一直是生物学研究的热点和难点。迄今为止,关于竹子开花现象的描述也有很多,但大多数都是针对木本竹类, 对于草本竹开花现象的研究却未见报道。藓竺属Mniochloa为禾本科竹亚科莪利竹族Olyreae草本竹类,多分布在巴西、古巴等地。因与提灯藓属Mnium很相似,1908年,CHASE将它命名为Mniochloa,并归入黍族Paniceae。近年来,研究者把Mniochloa和Ekmanochloa归入竹亚科莪利竹族[9-10]。本课题组将原产于巴西的草本竹Mniochloa abersend引种至浙江临安,对其形态特征与开花生物学特性进行系统观察与研究,以期为全面了解竹子的开花生物学特性,并可为后续开展竹子杂交育种等研究工作提供参考。
-
Mniochloa abersend(图 1A)原产于巴西东北部的累西腓市,地处8°06′S, 34°53′W,海拔为2~55 m。该地四季如夏,气候炎热潮湿,属于热带草原气候。极端高温为38 ℃,极端低温15 ℃,年平均气温为26.8 ℃,年平均降水量2 459 mm,年平均日照时数2 464 h。

图 1 Mniochloa abersend开花过程
Figure 1. Flowering period of Mniochloa abersend
-
引种地位于浙江省临安市,地处30°14′N,119°42′E,海拔100~150 m。该地四季分明,气候温暖湿润,雨水充足,属于亚热带季风性气候。极端高温为41.9 ℃,极端低温-13.3 ℃,年平均气温15.8 ℃,年平均降水量1 628.0 mm,年平均日照时数为1 939.0 h。Mniochloa abersend于2013年11月引种至浙江农林大学东湖校园内。
-
为了研究Mniochloa abersend引种后的生长适应情况,将其分为4丛,20株·丛-1,株高30~40 cm,分别移栽在实验楼驯化室(恒温25 ℃),智能温室(最低温约0 ℃),塑料大棚(最低温约-4 ℃)和室外空地(最低温约-8 ℃)里。2013年11月至2015年9月,连续观察竹丛的生长适应情况,观察其形态学特征,并记录竹丛的萌蘖伸长、抽枝、长叶等生长状态。
-
为了研究M. abersend的开花生物学特性,自引种时开始,连续2 a从有开花迹象开始对智能温室的竹子进行不定期观察,记录其开花预兆、开花次序、开花周期及持续时间等开花生物学特性。在盛花期随机选取20根开花枝条进行定时定株观察。每天8:00-12:00观察竹子开花情况;同时,取下开花的枝条带回实验室,在OLYMPUS SZX12体视显微镜下对花序进行解剖,观察花序、小穗、小花数、颖片、内外稃、花药、柱头及子房等花器官构造,并用OLYMPUS C-5060相机拍照。
-
Mniochloa abersend为多年生草本植物,秆丛生,较细弱,高80~100 cm,基部直立,地面以上成熟秆每节长为6~9 cm,直径为2.0~5.0 mm,节稍突起而被白色髯毛,各节均有分枝,枝条为绿色,因环境变化偶尔会出现紫红色;须根系且根系发达,有根毛,有时下部节上生根;叶鞘长于或短于节间,边缘薄纸质,鞘口及边缘有繸毛,长为3.0~5.0 mm;叶柄较短;叶舌短而低平,呈纤毛状;叶片呈披针状,先端渐尖,基部收缩,长为3.0~7.5 cm,宽为4.0~7.0 mm;叶片较光滑,疏生纤毛。
引种在驯化室的竹子在经过2周的适应以后逐渐恢复生长,开始抽枝长叶。智能温室的竹子刚开始地上部分枝条受冻枯黄,过冬以后,从竹丛基部重新抽枝长叶,生长健壮。塑料大棚和室外空地的竹子叶片全部受冻脱落,竹丛干枯死亡,不再抽枝长叶。引种结果表明,该草本竹对低温的适应能力较差,在25 ℃的环境下生长良好,在0 ℃左右虽然地上部分会产生冻害,但次年从基部萌生新芽,在0 ℃以下的环境下则会发生冻害甚至死亡。
-
Mniochloa abersend具有较明显的开花预兆,当竹子即将开花时,枝条顶端出现变型小叶(图 1B),较正常叶片小,长约为2.5 cm,宽约为3.0 mm。此时花序已经开始发育(图 1C),叶鞘饱满,鞘口出现淡黄色的芒(图 1D)。
-
Mniochloa abersend在1月底出现开花迹象,初期开花枝条稀少,3月底进入盛花期。整个竹丛外侧枝条先开花;同一根枝条则主枝的顶端先开花,随后侧枝顶端开花;同一花序上,中部小花先开放。无柄小穗中,第2小花较第1小花先成熟(图 1E)。
-
1月底,Mniochloa abersend的枝条顶端出现变型小叶,即进入花期,叶鞘逐渐饱满,鞘口出现淡黄色的芒,3~4 d后,芒全部伸出,长约1.0 cm。随之小穗开始伸出,未盛开时孪生小穗成对贴伏于花序轴上,随着小穗伸出,上部小穗的稃片开裂,雌蕊羽状柱头伸出。5 d左右,整个花序全部伸出,雌蕊白色羽状柱头从上到下依次伸出(图 1F)。2~3 d后小花成熟开放,内外稃微启,花药逐渐伸出(图 1G)。花药刚伸出时是淡黄色,伸出过程中逐渐变成淡紫色,花丝随着慢慢伸出,从花药刚露出到花丝全部伸出约需2~3 h,全部伸出后不到1 h,花药从非花丝一端开裂,花粉随风散去,花药2室分开成“X”形状,之后花药枯萎,由淡紫色逐渐变成褐色(图 1H)。小花的开放时间大多数在上午,下午偶尔也有开放。另外,由于持续的雨水天气,有时小穗花药伸出的过程伴随着散粉。当花药完全伸出时,就已经完成散粉过程。该草本竹的开花时间较长,一直可以持续到7月。
-
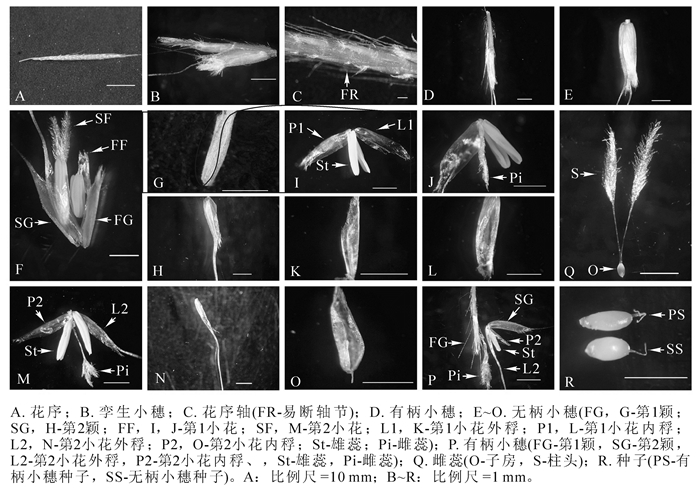
Mniochloa abersend的花序为总状花序,穗形(图 2A),单生于顶端,长约4.0 cm(包括芒1.0 cm在内,直立或微弯曲),宽约2.0 mm,微弯曲,乳黄色。孪生小穗着生于花序轴上,一无柄,一有柄(图 2B)。花序轴节长约1.5 mm,轴节间易断(图 2C),两侧具长短不一的纤毛。无柄小穗长约3.0 mm(图 2E和图 2F);第1颖(图 2G),长约2.8 mm,近先端边缘密具流苏状纤毛,长约0.5 mm,背部具3~5脉,背面稍粗糙,被微毛,无芒;第2颖与小穗等长,稍长于第1颖,舟形(图 2H),近先端边缘具流苏状纤毛,具1脉而呈脊,沿脊粗糙,先端有纤毛,脉延伸成芒,芒乳黄色,长15.0~18.0 mm;含2小花;第1小花雄性或两性(图 2I~J),外稃细长舟形(图 2K),长约2.0 mm,透明膜质,稍短于第1颖;内稃长卵圆形(图 2L),长约1.5 mm,透明膜质;雄蕊2枚,花丝极短,花药长约1.5 mm;雌蕊极小或不发育;第2小花两性(图 2M);外稃稍短于第1颖,先端有短纤毛,具一脉而伸出细弱而直的芒(图 2N),芒长约15.0 mm;内稃宽卵圆形(图 2O),短于外稃,透明膜质;雄蕊2枚,花药长约2.0 mm,通常比第1小花花药长;花柱长约1.5 mm,自基部分离为2枚,柱头长约1.5 mm,呈帚刷状,伸出小穗,长约1.0 mm,子房卵状圆形,长约0.3 mm(图 2Q)。有柄小穗较细小(图 2D和图 2P),基部有长短不一的纤毛,柄长约2.0 mm,约与小穗等长;第1小花缺;第2小花两性,具雄蕊1枚,花药长约1.5 mm;其他如无柄小穗第2小花。花序轴顶端有顶生小穗,与有柄小穗同形同性。

图 2 Mniochloa abersend花序结构
Figure 2. Inflorescence structure of Mniochloa abersend
-
Mniochloa abersend的大部分花序均有结实。种子未成熟时为淡黄色,成熟后为褐色。无柄小穗和有柄小穗种子形态稍有差异。无柄小穗种子短卵圆形,长约0.8 mm;有柄小穗种子细长梭形,长约1.0 mm(图 2R)。当种子成熟时,与花序自然分离,分离时仍被颖片及稃片包裹着(图 1I)。
-
Mniochloa abersend 为多年生草本竹类植物,外形较低矮,秆丛生,纤细而不木质化,没有复杂的分枝系统。虽然草本竹与大多数禾本科禾草外形上没有显著差异,但是在微观结构上,如叶片内部的解剖结构、叶片表皮细胞的形态等与木本竹类相同[3]。另外,Mniochloa abersend对低温的适应能力较差,在0 ℃以下的环境下会发生冻害甚至死亡,所以在引种时要充分考虑环境因素的影响。
竹类植物的花序对竹子的分类具有重要意义。McCLURE[11-12]关于竹子的无限制与有限制花序的理论为竹子分类研究奠定了基础。耿伯介[13]在此基础上进行了修改,指出:真花序为有限制的一次发生,具有总梗,上延即形成花序主轴。假花序为无限制地续次发生,无柄的假小穗在营养枝上呈簇状。根据花序的特点将木本竹类分为2个超族:一是具假花序的箣竹超族,如牡竹属Dendrocalamus,刚竹属Phyllostachys等;二是具真花序的北美箭竹超族,如香竹属Chimonocalamus,苦竹属Pleioblastus等。据报道,草本竹的花序均为真花序,目前已知的有圆锥花序和总状花序[3, 9]。本研究发现Mniochloa abersend的花序为穗形总状花序,为单次发生的真花序。
木本竹子的开花现象极为复杂。不同竹种在开花周期、开花次数、开花方式及表现等方面都不同。在开花周期上,毛竹Phllostachys edulis的开花周期甚长,可达60~70 a,而孝顺竹Bambusa multiplex间隔几年甚至年年开花;在开花次数上,毛竹一生只开花1次,Sasa veitchii 1 a开花2次[14],雷竹Ph. violascens则经常性开花。在开花方式及表现上,毛竹成片开花且开花后死亡,Sasa veitchii既有零星开花又有成片开花且通常开花后不死亡。雷竹是零星开花且在开花后2种情况并存。由此可见,木本竹子的开花情况难以预测。相反,草本竹的开花现象较为单一,具有每年开花以及整丛开花的特性,且开花后并不死亡,开花周期较固定,一般只相隔几个月,且花序都是真花序,罕见假小穗,花可两性、杂性或单性,这与禾草的开花生物学特性较为相似[3, 15-16]。但是,草本竹的一些开花特性与木本竹子也有相似之处。Mniochloa abersend在即将开花时会出现变型小叶,其枝条顶端先开花,同一花序中部小花先开放,且小花的开放时间大多数都在上午,这与麻竹Dendrocalamus latiflorus,绿竹Dendrocalamopsis oldhami和紫竹Phyllostachys nigra的开花特性是一致的[17-19]。草本竹类与木本竹类具有较近的亲缘关系,且每年开花结实,在研究其开花特性时相比木本竹类而言具有独特的优势,并可以为木本竹类的相关研究提供参考。
竹子是风媒传粉植物,由于外界环境因素导致传粉困难,结实率较低。此外,花粉不育或活力低也是导致其结实率较低的原因之一[20-22]。ABE等[14]对Sasa veitchii的2次开花现象以及其零星开花与成片开花与结实率的关系进行了统计分析,表明竹子的开花模式在一定程度上也影响了它的结实率。所以,Mniochloa abersend的每年开花及整丛开花的现象可能对其结实情况具有一定影响。但是,关于Mniochloa abersend的花粉活力、传粉授粉能力以及种子活力等都有待进一步深入研究。
Floral biology of Mniochloa abersend
-
摘要: 草本竹的开花现象鲜有报道。为了全面了解竹子的开花特性,以引种至浙江省临安市的原产于巴西的Mniochloa abersend为研究对象,首先对其形态特征和引种适应性进行了分析,之后采用定时定株的方法,对其开花预兆、开花次序、开花周期及持续时间、花序结构以及结实情况等开花生物学特性进行系统的观察与研究。结果表明:M. abersend为多年生丛生草本竹,植株矮小,不适宜在0 ℃以下环境下生长。开花前出现变型小叶,花期较长;花序属于穗形总状花序,为真花序;基本单位是孪生小穗,结实情况良好。M. abersend具有每年开花结实的特性。这将为竹子的杂交育种等研究提供参考。
图 2 参22 -
关键词:
- 植物学 /
- Mniochloa abersend /
- 开花生物学特性 /
- 花序 /
- 孪生小穗
Abstract: There is few research on the flowering phenomena of herbaceous bamboo. To fully understand the characteristics of bamboo flowering, the herbaceous bamboo Mniochloa abersend which was native to Brazil and introduced to Lin'an, Zhejiang was studied. The research examined the morphological characteristics and adaptability of Mniochloa abersend, and observed the selected plants at the stated hours to study their flowering biological characteristics including flowering omens, blooming sequence of florets, flowering period and duration, inflorescence structure as well as seed setting. The results demonstrated that M. abersend was perennial and tufted herbaceous bamboo, which grew not so well under 0 ℃ environment. Deform leaves would appear before flowering. Besides, M. abersend had long florescence, good seed production and spike-shaped raceme, belonging to the genuine inflorescence, whose basic units were twin spikes. In conclusion, M. abersend blossomed and set seeds annually, which could be a reference for crossing breeding and other research on bamboos.[Ch,2 fig. 22 ref. ]-
Key words:
- botany /
- Mniochloa abersend /
- flowering biological characteristics /
- inflorescence /
- twin spikes
-
[1] 郭振华, 李德铢.竹亚科系统学和生物地理学研究进展及存在的问题[J].云南植物研究, 2002, 24(4):431-438. GUO Zhenhua, LI Dezhu. Advances in the systematics and biogeography of the Bambusoideae (Gramineae) with remarks on some remaining problems [J]. Acta Bot Yunnan, 2002, 24(4): 431-438. [2] WU Zhengyi. Flora of China: Poaceae [J]. Beijing: Science Press, 2006: 22. [3] 耿伯介.世界竹亚科各属的考订(之六)[J].竹子研究汇刊, 1984, 3(2):2-21. GENG Bojie. A revision of the genera of bamboos from the world (Ⅵ) [J]. J Bamboo Res, 1984, 3(2): 2-21. [4] 杜凡, 薛嘉榕, 杨宇明, 等. 15年来云南竹子的开花现象及其类型研究[J].林业科学, 2000, 36(6): 57-68. DU Fan, XUE Jiarong, YANG Yuming, et al. Study on flowering phenomenon and its types of bamboo in Yunnan in past fifteen years [J]. Sci Silv Sin, 2000, 36(6): 57-68. [5] GUERREIRO C. Flowering cycles of woody bamboos native to southern South America [J]. J Plant Res, 2014, 127(2): 307-313. [6] 柴振林, 秦玉川, 华锡奇, 等.竹子开花原因研究进展[J].浙江林业科技, 2006, 26(2):53-57. CHAI Zhenlin, QIN Yuchuan, HUA Xiqi, et al. Advance of studies on bamboo flowering causes [J]. J Zhejiang For Sci Technol, 2006, 26(2): 53-57. [7] 谢寅峰, 张春霞, 丁雨龙.竹子开花人工诱导与逆转研究现状及前景[J].浙江林学院学报, 2000, 17(4):436-440. XIE Yinfeng, ZHANG Chunxia, DING Yulong. Current situation and prospects of study on artificial induction and reversion of bamboo flowering [J]. J Zhejiang For Coll, 2000, 17(4): 436-440. [8] CAMPBELL J J N. Bamboo flowering patterns: a global view with special reference to East Asia [J]. J Am Bamboo Soc, 1985, 6: 17-35. [9] ZULOAGA F O, MORRONE O, JUDZIEWICZ E J. Endemic herbaceous bamboo genera of Cuba (Poaceae: Bambusoideae: Olyreae) [J]. Ann Missouri Bot Gard, 1993, 80(4): 846-861. [10] JUDZIEWICZ E J, SEPSENWOL S. The world's smallest bamboo: Raddiella vanessiae (Poaceae: Bambusoideae: Olyreae), a new species from French Guiana [J]. J Bot Res Inst Texas, 2007, 1(1): 1-7. [11] McCLURE F A.竹子的生殖相(上)[J].洪效训, 译.竹子研究汇刊, 1982, 1(2):104-116. McCLURE F A. Reproductive phase of bamboos [J]. J Bamboo Res, 1982, 1(2): 104-116. [12] McCLURE F A.竹子的生殖相(下)[J].洪效训, 译.竹子研究汇刊, 1983, 2(2):119-135. McClure F A. Reproductive phase of bamboos [J]. J Bamboo Res, 1983, 2(2):119-135. [13] 耿伯介.试论竹类的花序及其演变[J].武汉植物学研究, 1986, 4(4):323-335. GENG Bojie. A preliminary study of the inflorescence type arising from bamboos and its variation [J]. J Wuhan Bot Res, 1986, 4(4): 323-335. [14] ABE Y, SHIBATA S. Flower and seed production in a series of flowerings from sporadic events before to after mass flowering of the dwarf bamboo Sasa veitchii var. hirsuta [J]. J For Res, 2014, 19(2): 268-275. [15] 李青丰, 常峰, 董天明.几种禾本科牧草开花结实特性的研究[J].内蒙古草业, 2000(1):41-43. LI Qingfeng, CHANG Feng, DONG Tianming. A preliminary study on flowering and seed production of 6 herbage grasses [J]. Inner Mongolia Pratac, 2000(1): 41-43. [16] 张新全, 杜逸, 郑德成, 等.几种禾草的开花习性[J].草业科学, 1994, 11(3):42-45. ZHANG Xinquan, DU Yi, ZHENG Decheng, et al. Flowering behavior of some grasses [J]. Pratacl Sci, 1994, 11(3): 42-45. [17] 邢新婷, 傅懋毅, 肖贤坦.麻竹开花生物学特性观察及控制授粉的初步研究[J].北京林业大学学报, 2006, 27(6):103-107. XING Xinting, FU Maoyi, XIAO Xiantan. Biological characteristics of flowering and controlled pollination of Dendrocalamus latiflorus Munro [J]. J Beijing For Univ, 2005, 27(6): 103-107. [18] 郑郁善, 高培军, 陈礼光, 等.绿竹开花生理生化特性研究[J].林业科学, 2003, 39(3):143-147. ZHENG Yushan, GAO Peijun, CHEN Liguang, et al. A study on the physiological and biochemical character of flowering for Dendrocalamopsis oldhami [J]. Sci Sliv Sin, 2003, 39(3): 143-147. [19] 李晓芬, 林新春, 时燕, 等.紫竹开花生物学特性观察及花粉生活力测定[J].林业科技开发, 2009, 23(6):64-67. LI Xiaofen, LIN Xinchun, SHI Yan, et al. Flowering biological characteristics and pollen viability of Phyllostachys nigra [J]. China For Sci Technol, 2009, 23(6): 64-67. [20] 林树燕, 丁雨龙, 张昊. 5种竹子花粉萌发率及开花特性[J].林业科学, 2008, 44(10):159-163. LIN Shuyan, DING Yulong, ZHANG Hao. Pollen germination percentage and the floral character of five bamboo species [J]. Sci Sliv Sin, 2008, 44(10): 159-163. [21] 林新春, 袁晓亮, 林绕, 等.雷竹开花生物学特性研究[J].福建林学院学报, 2010, 30(4):333-337. LIN Xinchun, YUAN Xiaoliang, LIN Rao, et al. Studies on floral biology of Phyllostachys violascens [J]. J Fujian Coll For, 2010, 30(4): 333-337. [22] WAIKHOM S D, LOUIS B, ROY P, et al. Scanning electron microscopy of pollen structure throws light on resolving Bambusa-Dendrocalamus complex: bamboo flowering evidence [J]. Plant Syst Evol, 2014, 300(6): 1261-1268. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2016.05.005
 点击查看大图
点击查看大图

图(2)
计量
- 文章访问数: 4293
- HTML全文浏览量: 1121
- PDF下载量: 440
- 被引次数: 0
