





-
淹水胁迫对植物生长的抑制作用除了低氧环境引起的根系活力下降、呼吸抑制以及矿质元素吸收受阻外,长时间淹水胁迫引起的叶绿素合成受阻与降解加速导致了叶光合色素含量下降、光能利用与转化活性改变,进而引起光合能力的大幅度下降[1]。光合作用是植物生存和繁衍的物质基础,在这个复杂的生理生化过程中,受到伤害的最原初部位是与光系统Ⅱ(PSⅡ)紧密联系的[2-5]。植物淹水后会导致PSⅡ光化学活性和电子传递速率降低[6],PSⅡ捕光色素蛋白复合物(LHCⅡa,LHCⅡb,LHCⅡc)各组分的变化,从而引起光合二氧化碳同化效率的降低[7]。另一方面,植物也可以以热的形式耗散过剩光能[8],PSⅡ反应中心的失活和周转[9]及Mehler反应[10]等减轻光抑制过程,从而保护光合机构免受破坏。叶绿素荧光参数最大光化学效率(Fv/Fm),PSⅡ实际光化学效率(Fv′/Fm′),光化学荧光猝灭系数(qP)和非光化学猝灭系数(qN),PSⅡ的实际光化学量子产量(Yyield),表观光合电子传递速率(RET)等的变化可反映逆境胁迫对PSⅡ的损伤程度[11-12],已经广泛应用于光抑制、水分、高温、低温等逆境生理研究[13-14]。竹子是集经济、生态和社会效益于一体的优良林种,是区域农村经济社会发展的重要资源和生态环境保护的重要屏障。水分、温度、光照等环境条件的变化直接影响着竹子的生长发育和分布。随着全球气候的变化,水资源不均匀分布造成近年来极端干旱和洪涝灾害频发,水分胁迫已经成为影响竹子生长发育的主要逆境因子之一,研究竹子对水分胁迫的适应能力越来越受到关注[15]。目前,国内外相关研究主要集中在短期干旱或水淹对竹子生长和生理生态的影响[16-21],而对于长期处于浸渍环境中的竹子生理生态响应及其机制研究甚少[22]。河竹Phyllostachys rivalis隶属禾本科Gramineae倭竹族Shibataeeae刚竹属Phyllostachys,主要分布于浙江、福建等地,生于溪涧边、山沟旁,性喜水湿,鞭根系统极为发达,竹鞭韧皮部密生一圈肉眼可见的气孔,具有耐淹植物的特征。我们前期的研究表明,河竹鞭根系统可以通过抗氧化系统平衡调节、生物量合理分配和异速生长调节等来适应长期淹水环境,维持生长和更新[23-24],而长期水淹胁迫下河竹叶绿素荧光变化特征、能量耗散过程及其与河竹耐受水淹的关系尚不清楚。为此,本研究以2年生河竹盆栽苗为试材,设置不同的水淹深度处理,测定分析不同水淹时间下叶片荧光参数和能量耗散的变化规律,探讨持续淹水对河竹光能的吸收和转化、能量的传递与分配、反应中心的活性、过剩能量的耗散以及光合作用的光抑制和光破坏等的影响,并从光合系统“内在性”揭示河竹对持续淹水的响应与适应机制。
-
试验地位于浙江省临安市太湖源观赏竹种园内。该地属中亚热带湿润季风气候区,年平均气温为15.4 ℃,极端低温-10.3 ℃,极端高温44.5 ℃,年日照时数为1 850~1 950 h,日均高于10 ℃活动积温为5 100 ℃,年平均无霜期为235 d,年降水量为1 250~1 600 mm,年平均空气相对湿度80%以上。
2012年2月在河竹种苗林中挖取2年生小丛状竹苗,竹苗地径(1.0 ± 0.2)cm,全高(1.0 ± 0.4)m,保留5~6盘枝,选择规格基本一致的竹苗移栽到加仑盆中(上端直径32.0 cm,下端直径23.0 cm,高度27.0 cm),以V(红壤):V(细沙)=3:1为培养基质,填充基质约15.0 kg·盆-1,栽植竹苗10株·盆-1。移栽后正常喷灌和清除竹笋、杂草等管理。
2013年4月15日选择生长状况一致的河竹盆栽苗进行淹水处理。设3个梯度,即对照(ck),处理Ⅰ和处理Ⅱ。对照实行正常浇水,使盆栽基质相对含水率保持85.0%±5.0%;处理Ⅰ淹水水位高于土壤表面5.0 cm;处理Ⅱ淹水水位高于土壤表面10.0 cm。试验盆栽苗置于长度4.3 m,宽度3.3 m和深度0.5 m的方形水泥池中进行淹水处理,试验期间保持设定水位。设置重复10个·处理-1,即盆栽苗10盆。
-
根据已有报道,自然消落带淹水时间平均3个月左右,最长时间可达6个月[8]。而前期试验表明,淹水360 d后,河竹仍能正常生长,且表现出一定的更新能力。为了解河竹叶片荧光参数和能量耗散等方面对持续淹水的响应,本研究设置淹水处理时间分别为30,90,180,270和360 d。在设定时间,随机选取3盆·处理-1河竹盆栽苗,在9:00-10:00,采用PAM-2500便携式脉冲调制叶绿素荧光仪(德国Walz公司)测定叶片叶绿素荧光参数。对河竹顶部倒数第3~4盘枝选择4~7片成熟叶片,先将测定植株叶片用黑色布袋子罩住,暗适应30 min,使得待测叶片所处光环境一致,全部使用仪器提供的测量光、光化光及饱和脉冲光测定叶片的初始荧光(Fo)和最大荧光(Fm)。作用光打开后测定光下最小荧光(Fo′)和光下最大荧光(Fm′),以荧光慢诱导模式测定光系统Ⅱ(PSⅡ)的Fv/Fm,Fv′/Fm′,qP,qN,Yyield和RET。PSⅡ吸收光能分配百分率参照Demmig Adams和Adams公式计算[25]:光化学反应的能量(P)= Fv′/Fm′×qP;非光化学反应耗散的能量(E)=(1-qP)×Fv′/Fm′;天线色素耗散的能量(D)=1-Fv′/Fm′。
-
采用SPSS 20.0统计软件进行单因素方差分析(One-way ANOVA),用Duncan方法进行多重比较,用Excel 2010绘制图表。
-
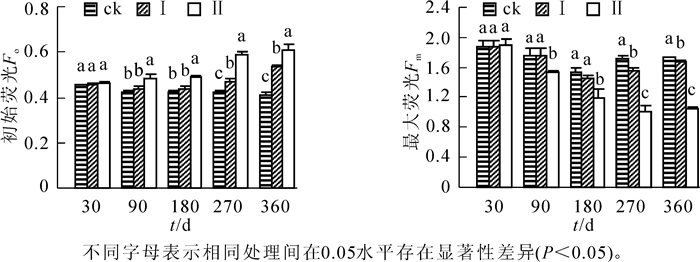
由图 1可知:随淹水时间的延长,淹水处理的河竹叶片Fo总体呈升高趋势,而Fm总体呈下降趋势。短期淹水处理(30 d)对河竹叶片Fo和Fm并无明显影响,但淹水时间进一步延长,处理间差异增大,水深效应也日趋明显,至淹水90 d和180 d时,处理Ⅱ的河竹叶片Fo显著高于处理Ⅰ和ck(P<0.05),而Fm显著低于处理Ⅰ和ck(P<0.05),且后两者Fo和Fm均无显著差异(P>0.05),其后至淹水处理结束,淹水处理的河竹叶片Fo持续升高,Fm总体上持续下降,且水位效应更加明显,处理间差异均达显著水平(P<0.05)。

图 1 持续淹水下河竹叶片初始荧光和最大荧光
Figure 1. Effects of long-term flooding on Fo and Fm in leaves of Phyllostachys rivalis
-
随着淹水时间的延长和淹水深度的增大,河竹叶片Fv/Fm呈下降趋势,Fv′/Fm′呈先升高后降低的趋势(图 2)。相对ck,短期淹水(30 d),河竹叶片Fv/Fm降低,但处理间差异并不显著(P>0.05),而Fv′/Fm′则升高,且处理Ⅱ显著高于ck(P<0.05);至淹水90 d和180 d时,淹水处理河竹叶片Fv/Fm明显下降,处理Ⅱ显著低于处理Ⅰ和ck(P<0.05),而后两者间无显著差异(P>0.05);其后至淹水处理结束,淹水处理河竹叶片Fv/Fm持续下降,处理间差异达显著水平(P<0.05),水位效应明显,但处理Ⅰ和处理Ⅱ仍有ck的89.4%和55.4%。河竹叶片Fv′/Fm′较Fv/Fm对淹水胁迫更敏感,短期淹水即会引起Fv′/Fm′的明显升高,淹水处理30 d时,处理Ⅱ就显著高于ck(P<0.05),处理90 d时,处理Ⅰ和处理Ⅱ均显著高于ck(P<0.05),但至处理180 d时各处理的Fv′/Fm′均明显下降,且处理间并无显著差异(P>0.05),其后淹水处理的Fv′/Fm′显著下降,水位效应较为明显。

图 2 持续淹水下河竹叶片psⅡ的最大光化学效率和psⅡ实际光化学效率
Figure 2. Effects of long-term flooding on Fv/Fm and Fv′Fm′ in leaves of Phyllostachys rivalis
-
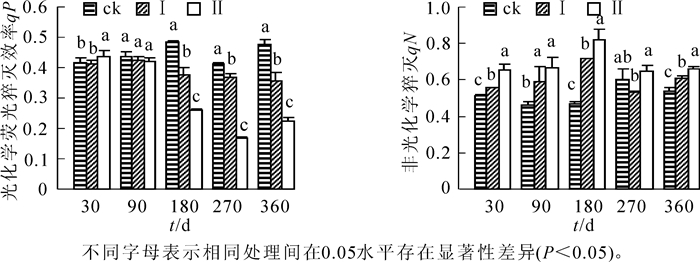
由图 3可知:随着淹水时间的延长,不同淹水处理下的河竹叶片qP总体呈下降趋势,qN呈升高趋势,不同处理间变化幅度不同。相对ck,淹水30 d时,处理Ⅱ的qP显著升高(P<0.05);淹水90 d时,河竹叶片qP开始降低,但处理Ⅰ和处理Ⅱ与ck差异不显著(P>0.05);淹水180 d时至处理结束,河竹叶片qP为ck>处理Ⅰ>处理Ⅱ,各处理间均有显著差异(P<0.05),水位效应明显。整个淹水过程中,河竹叶片qN基本上为ck<处理Ⅰ<处理Ⅱ,总体上淹水处理显著高于ck(P<0.05),水位效应也较为明显。

图 3 持续淹水下河竹叶片光化学荧光猝灭系数和非光化学猝灭系数
Figure 3. Effects of long-term flooding on qP and qN in leaves of Phyllostachys rivalis
-
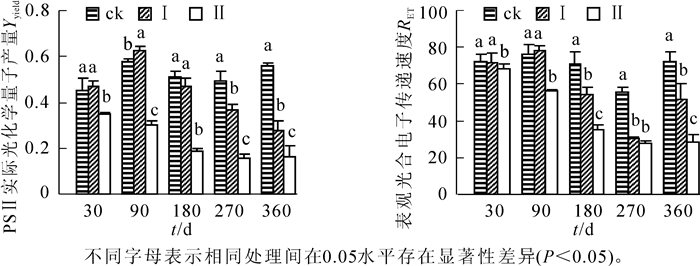
由图 4可知:不同淹水处理的河竹叶片Yyield和RET的变化不同。随着淹水时间的延长,处理Ⅰ的Yyield先升高后降低,RET先升高后降低再升高,在淹水90 d时均达最高值。在处理180 d后,各处理间河竹叶片Yyield和RET总体上差异显著(P<0.05);处理Ⅱ的Yyield和RET均随着淹水时间的延长而逐渐降低,各处理时间点上均显著低于ck(P<0.05)。至淹水180 d后,河竹叶片的Yyield和RET均随着淹水深度的增大而显著降低(P<0.05),存在明显的水位效应。

图 4 持续淹水下河竹叶片psⅡ实际化学量子产量和电子传递速率
Figure 4. Effects of long-term flooding on Yyield and RET in leaves of Phyllostachys rivalis
-
由表 1可知:随着淹水时间的延长,处理Ⅰ和处理Ⅱ的河竹叶片光化学反应能量(P)均呈先升高后下降的变化趋势。淹水30 d和90 d时,处理Ⅰ和处理Ⅱ的P较ck升高,且处理Ⅱ与ck差异显著(P<0.05);淹水180,270和360 d时,处理Ⅰ和处理Ⅱ的P均显著低于ck(P<0.05),且处理Ⅰ和处理Ⅱ间差异显著(P<0.05),水位效应明显。天线色素耗散能量(D)随着淹水时间的延长呈先降低后升高的变化趋势。淹水30 d和90 d时,处理Ⅰ和处理Ⅱ的D均显著低于ck(P<0.05);淹水180 d时,处理Ⅰ和处理Ⅱ的D仍低于ck,但未达显著差异水平(P>0.05);淹水270 d和360 d时,处理Ⅰ和处理Ⅱ的D均显著高于ck(P<0.05)。整个淹水处理过程中,河竹叶片天线色素耗散能量(D)的水位效应总体上并不明显。在淹水30,90和180 d时,处理Ⅰ和处理Ⅱ的非光化学反应耗散能量(E)较ck显著升高(P<0.05),但处理Ⅰ和处理Ⅱ之间差异并不显著(P>0.05);淹水270 d时,处理Ⅰ的E较ck显著降低(P<0.05);淹水360 d时,处理Ⅰ和处理Ⅱ的E较对照降低(P>0.05),河竹叶片非光化学反应耗散能量(E)的水位效应总体上也并不明显。
表 1 持续淹水对河竹叶片吸收光能分配的影响
Table 1. Effects of long-term flooding on characteristics fractions of absorbed light utilized in leaves of Phyllostachys rivalis
参数 处理 不同淹水时间河竹叶叶片吸收光能/% 30 90 180 270 360 d 光化学反应能量(P) ck 22.8 ± 0.7 b 24.6 ± 0.6 b 23.5 ± 1.1 a 26.3 ± 0.3 a 28.1 ± 3.4 a Ⅰ 23.8 ± 0.7 b 26.1 ± 0.2 a 20.0 ± 1.2 b 19.5 ± 0.8 b 15.1 ± 0.7 b Ⅱ 25.2 ± 0.6 a 26.4 ± 0.2 a 12.6 ± 1.0 c 7.6 ± 0.3 c 8.5 ± 0.7 c 天线色素耗散能量(D) ck 45.1 ± 1.3 a 43.7 ± 3.0 a 51.3 ± 2.1 a 36.5 ± 1.1 c 41.2 ± 5.7 b Ⅰ 42.6 ± 1.6 b 38.4 ± 0.9 b 46.8 ± 3.5 a 47.3 ± 2.0 b 57.3 ± 2.5 a Ⅱ 42.2 ± 0.5 b 37.0 ± 0.5 b 51.5 ± 3.3 a 55.1 ± 2.4 a 61.7 ± 0.9 a 非光化学反应耗散能量(E) ck 32.1 ± 0.7 b 31.8 ± 2.6 b 25.2 ± 1.0 b 37.2 ± 0.8 a 30.7 ± 2.3 a Ⅰ 33.7 ± 0.9 a 35.6 ± 0.7 a 33.3 ± 2.3 a 33.3 ± 1.2 b 27.5 ± 2.8 a Ⅱ 32.5 ± 0.2 ab 36.6 ± 0.3 a 35.9 ± 2.4 a 37.3 ± 2.1 a 29.8 ± 0.6 a 说明:同列不同字母表示在0.05水平存在显著性差异。 -
环境胁迫影响植物的光合作用过程,造成光化学转换效率和电子传递速率与能量分配之间产生矛盾,从而影响光合碳同化能力,PSⅡ反应中心生理功能的稳定性是植物抵抗逆境胁迫的能力体现[26-28]。叶绿素荧光参数从能量代谢与转换的角度反映光合机构受逆境胁迫伤害的程度[29-31],Fm,Fv/Fm降低表明植物叶片发生光抑制[10],而Fv/Fm下降的同时Fo上升,表明PSⅡ反应中心受到损伤[29]。本研究中,处理Ⅰ在淹水30 d至180 d时,Fo,Fm和Fv/Fm变化不显著,说明淹水深度5 cm持续淹水180 d,河竹叶片PSⅡ的活性一直维持在正常水平,具有良好的适应能力,至淹水270 d时,处理Ⅰ的Fo显著升高,Fm和Fv/Fm降低,说明此时河竹叶片PSⅡ反应中心的內禀光能转化效率和活性随淹水时间持续而降低,光合作用的原初反应受到抑制,不利于河竹叶片捕获光能的转化。而处理Ⅱ在淹水90 d时,Fo显著上升,Fm和Fv/Fm下降,说明淹水深度10 cm持续淹水90 d,河竹叶片PSⅡ开始受到损伤。淹水360 d,相对对照,处理Ⅰ和处理Ⅱ河竹叶片的Fv/Fm显著降低,但仍具有相对较高的水平,这与长期水淹后枫杨Pterocarya stenoptera幼苗[8]的研究结果一致。说明长期淹水影响河竹叶片PSⅡ反应中心的活性,但对PSⅡ的功能反应中心影响较小。
荧光猝灭是植物内光合量子效率调节的一个方面,分为光化学荧光猝灭(qP)和非光化学猝灭(qN)2类[32]。qP值的大小反映PSⅡ反应中心开放部分的比例及电子传递速率[33-34]。本研究中,淹水30 d时,处理Ⅰ维持相稳定对的qP,Yyield和RET值,有效地避免或减轻因PSⅡ吸收而引起的光抑制和光氧化,保护了光合机构正常运转。处理Ⅱ的qP值上升,说明初期水淹有利于河竹叶片PSⅡ反应中心电子传递能力的提高,促进捕获的光能更高效地用于光合作用。淹水180 d后,处理Ⅰ和处理Ⅱ的qP均显著下降,说明PSⅡ反应中心电子传递受阻,在一定程度上降低了河竹叶片PSⅡ的活性。非光化学猝灭(qN)常用来评价植物耗散过剩激发能的能力[4, 35]。本研究中,淹水处理Ⅰ和处理Ⅱ条件下河竹叶片的qN随着淹水深度及持续时间而增高,热耗散保护性作用增强。这可能是河竹适应淹水环境而形成的自我保护机制。
通过叶片的化学反应的能量(P),天线色素耗散的能量(D)和非光化学反应耗散的能量(E)及其占总吸收光能的比例,可以了解植物逆境环境下的光能利用能力[36]。本研究表明,淹水30 d和90 d时,处理Ⅰ和处理Ⅱ的P,E显著升高,D降低,说明淹水初期不同水位处理均提高了光合作用碳同化电子需求,E的增加可能会引起光合机构的可逆失活甚至破坏[10]。淹水360 d时,处理Ⅰ和处理Ⅱ的P明显减少,D显著升高,而E恢复对照水平,减轻了PSⅡ的激发压,从而能及时的使单线态叶绿素(1Chl)返回三线态叶绿素(3Chl)[37],降低了形成单线态氧(1O2)的机会,表明长期淹水后河竹叶片PSⅡ能力恢复,天线色素耗散的能量(D)上升,可减少PSⅡ和电子传递的过分还原,从而防止过剩光能对光合机构的破坏。持续淹水环境下,河竹吸收光强主要以天线色素耗散(D)为主要光能分配途径,淹水后期PSⅡ反应中心的非光化学反应耗散(E)的恢复起着重要作用,这种变化充分反映了河竹对淹水的适应能力。
-
综上所述,淹水环境下河竹能通过维持相对较高的RET,qP和P值,增强qN来调节自身能量代谢,以热耗散形式散失过多的光能,有效地避免或减轻光抑制和光氧化,河竹吸收光强主要以天线色素耗散(D)为主要光能分配途径,淹水后期PSⅡ反应中心的非光化学反应耗散(E)的恢复起着重要作用。这些可能是河竹适应淹水环境的自我保护机制。
Chlorophyll fluorescence and excitation energy dissipation of pot-grown Phyllostachys rivalis leaves after long-term flooding
-
摘要: 为揭示河竹Phyllostachys rivalis对持续淹水的生理生态响应与适应机制,为河竹在水陆交错带的应用提供理论依据,以2年生河竹盆栽苗为试材,设置不同深度的淹水处理[水位高出栽培基质5 cm(处理Ⅰ),10 cm(处理Ⅱ)和正常供水(ck)],测定持续淹水30,90,180,270和360 d时河竹叶片叶绿素荧光参数,分析叶片光能的吸收和转化、能量的传递与分配、反应中心的活性、过剩能量的耗散对持续淹水胁迫的响应。结果表明:① 持续淹水胁迫下,河竹叶片能通过维持相对较高的表观光合电子传递速率(RET),光化学荧光猝灭(qP)和光化学反应能量(P),增强非光化学猝灭(qN)来调节自身能量代谢,以热耗散形式散失过多的光能,有效地避免或减轻光抑制和光氧化,河竹吸收光强主要以天线色素耗散(D)为主要光能分配途径,淹水后期PSⅡ反应中心的非光化学反应耗散(E)的恢复起着重要作用,持续淹水一定程度上会损害河竹叶片光系统Ⅱ(PSⅡ),但对于PSⅡ的功能反应中心影响较小。② 不同淹水处理叶绿素荧光参数存在差异,持续淹水30,90 d时,处理Ⅰ的初始荧光(Fo),最大荧光(Fm),最大光化学效率(Fv/Fm),qP和RET等与ck差异不显著(P > 0.05),而处理Ⅱ在淹水90 d时与ck差异显著(P < 0.05),至淹水270,360 d时,各处理间差异均达显著水平(P < 0.05),持续淹水胁迫对叶绿素荧光参数及能量耗散的影响存在明显的水位效应。Abstract: To determine the physiological and biochemical responses and adaptive mechanisms of Phyllostachys rivalis to long-term soil flooding, two-year-old potted seedlings of Ph. rivalis were subjected to different flooding depths. The flooding treatment was set with water levels of 5 cm (TreatmentⅠ) and 10 cm (TreatmentⅡ) higher than the soil surface and normal water supply (ck). Then the chlorophyll fluorescence parameters in leaves were determined after continuous flooding for 30, 90, 180, 270, and 360 d. Responses to the continuous flooding stress for light energy absorption and transformation, energy transfer and distribution, reaction center activity, and excitation energy dissipation in leaves of three leaves per seedlings and three seedlings were measured and analyzed. A one-way analysis of variance with duncan's test was conducted at a significant level of 0.05. Results showed that the chlorophyll fluorescence parameters varied with different flooding levels. When flooding for 30 d and 90 d, minimal fluorescence (Fo), maximum fluorescence (Fm), photochemical maximum efficiency of PS Ⅱ (Fv/Fm), photochemical quenching coefficient (qP), and electron transport rate(RET) of Treatment Ⅰ were not significantly different (P > 0.05) compared with ck; however, Treatment Ⅱ when flooding for 90 d compared with ck was significantly increased for Fo while significantly decreased for Fm, Fv/Fm and RET (P < 0.05). With flooding treatments of 270 d and 360 d, Fm, Fv/Fm, qP and RET of Treatment Ⅰ and Ⅱ were both significant decreased while Fo were significant increased than that of ck (P < 0.05). The effect of flooding on chlorophyll fluorescence parameters and energy dissipation was also dependent on the water level. In leaves of Ph. rivalis, Fm, Fv/Fm, and qP were significantly decreased (P < 0.05) with continuous flooding stress; whereas, Fo and non-photochemical quenching coefficient (qN) increased gradually but not reached significant level (P > 0.05). The amount of absorbed light in photochemistry (P) and excess energy (E) increased first and then decreased (P < 0.05), but the energy of dissipation of the antenna heat dissipation (D) observed were opposite. Taken together, Ph. rivalis could maintain a relatively high RET, qP, and P in the early stages of flooding, and enhance qN to regulate their energy metabolism, dissipate excess light energy via heat dissipation, thereby alleviating the light photoinhibition and photooxidation; however, continuous long-term soil flooding could damage photosystem Ⅱ (PS Ⅱ) even though there was minimal adverse effect on the functional center of PS Ⅱ (P < 0.05). It can be inferred that Ph. rivalis can tolerate a short term flooding which facilitate its possible application in plantation restoration of riparion zone.
-
图 1 持续淹水下河竹叶片初始荧光和最大荧光
Figure 1 Effects of long-term flooding on Fo and Fm in leaves of Phyllostachys rivalis
图 2 持续淹水下河竹叶片psⅡ的最大光化学效率和psⅡ实际光化学效率
Figure 2 Effects of long-term flooding on Fv/Fm and Fv′Fm′ in leaves of Phyllostachys rivalis
图 3 持续淹水下河竹叶片光化学荧光猝灭系数和非光化学猝灭系数
Figure 3 Effects of long-term flooding on qP and qN in leaves of Phyllostachys rivalis
图 4 持续淹水下河竹叶片psⅡ实际化学量子产量和电子传递速率
Figure 4 Effects of long-term flooding on Yyield and RET in leaves of Phyllostachys rivalis
表 1 持续淹水对河竹叶片吸收光能分配的影响
Table 1. Effects of long-term flooding on characteristics fractions of absorbed light utilized in leaves of Phyllostachys rivalis
参数 处理 不同淹水时间河竹叶叶片吸收光能/% 30 90 180 270 360 d 光化学反应能量(P) ck 22.8 ± 0.7 b 24.6 ± 0.6 b 23.5 ± 1.1 a 26.3 ± 0.3 a 28.1 ± 3.4 a Ⅰ 23.8 ± 0.7 b 26.1 ± 0.2 a 20.0 ± 1.2 b 19.5 ± 0.8 b 15.1 ± 0.7 b Ⅱ 25.2 ± 0.6 a 26.4 ± 0.2 a 12.6 ± 1.0 c 7.6 ± 0.3 c 8.5 ± 0.7 c 天线色素耗散能量(D) ck 45.1 ± 1.3 a 43.7 ± 3.0 a 51.3 ± 2.1 a 36.5 ± 1.1 c 41.2 ± 5.7 b Ⅰ 42.6 ± 1.6 b 38.4 ± 0.9 b 46.8 ± 3.5 a 47.3 ± 2.0 b 57.3 ± 2.5 a Ⅱ 42.2 ± 0.5 b 37.0 ± 0.5 b 51.5 ± 3.3 a 55.1 ± 2.4 a 61.7 ± 0.9 a 非光化学反应耗散能量(E) ck 32.1 ± 0.7 b 31.8 ± 2.6 b 25.2 ± 1.0 b 37.2 ± 0.8 a 30.7 ± 2.3 a Ⅰ 33.7 ± 0.9 a 35.6 ± 0.7 a 33.3 ± 2.3 a 33.3 ± 1.2 b 27.5 ± 2.8 a Ⅱ 32.5 ± 0.2 ab 36.6 ± 0.3 a 35.9 ± 2.4 a 37.3 ± 2.1 a 29.8 ± 0.6 a 说明:同列不同字母表示在0.05水平存在显著性差异。  下载: 导出CSV
下载: 导出CSV
-
[1] DREENWAY H, ARMSTRONG W, COLMER T D. Conditions leading to high CO2 ( > 5 kPa) in waterlogged-flooded soils and possible effects on root growth and metabolism [J]. Ann Bot, 2006, 98(1): 9-32. [2] WEIS E, BERRY J A. Plants and high temperature stress [J]. Symp Soc Exp Biol, 1988, 42: 329-346. [3] HAVAUX M, TARDY F. Temperature-dependent adjustment of the thermal stability of photosystem Ⅱ in vivo: possible involvement of xanthophyll-cycle pigments [J]. Planta, 1996, 198(3): 324-333. [4] 史胜青, 袁玉欣, 杨敏生, 等.水分胁迫对4种苗木叶绿素荧光的光化学淬灭和非光化学淬灭的影响[J].林业科学, 2004, 40(1):168-172. SHI Shengqing, YUAN Yuxin, YANG Minsheng, et al. Effects of water stress on photochemical quenching and non-photochemical quenching of chlorophyll a fluorescence in four tree seedlings [J]. Sci Silv Sin, 2004, 40(1): 168-172. [5] 罗俊, 张木清, 吕建林, 等.水分胁迫对不同甘蔗品种叶绿素a荧光动力学的影响[J].福建农业大学学报, 2000, 29(1):18-22. LUO Jun, ZHANG Muqing, LÜ Jianglin, et al. Effects of water stress on the chlorophyll a fluorescence induction kinetics of sugarcane genotypes [J]. J Fujian Agric Univ, 2000, 29(1): 18-22. [6] 衣英华, 樊大勇, 谢宗强, 等.模拟淹水对枫杨和栓皮栎气孔交换、叶绿素荧光和水势的影响[J].植物生态学报, 2006, 30(6):960-968. YI Yinghua, FAN Dayong, XIE Zongqiang, et al. Effects of waterlogging on the gas exchange, chlorophyll fluorescence and water potential of Quercus variabilis and Pterocarya stenoptera [J]. J Plant Ecol, 2006, 30(6): 960-968. [7] 韦振泉, 林宏辉, 何军贤, 等.水分胁迫对小麦捕光色素蛋白复合物的影响[J].西北植物学报, 2000, 20(4):555-560. WEI Zhenquan, LIN Honghui, HE Junxian, et al. Effects of water stress on the light-harvesting complexes in wheat leaves [J]. Acta Bot Boreal-Occident Sini, 2000, 20(4): 555-560. [8] 贾中民, 魏虹, 田晓峰, 等.长期水淹对枫杨幼苗光合生理和叶绿素荧光特性的影响[J].西南大学学报(自然科学版), 2009, 31(5):124-129. JIA Zhongmin, WEI Hong, TIAN Xiaofeng, et al. Effects of long-term flooding on photosynthesis and chlorophyll fluorescence parameters of Pterocarya stenoptera seedlings [J]. J Southwest Univ Nat Sci Ed, 2009, 31(5): 124-129. [9] 陈贻竹, 李晓萍, 夏丽, 等.叶绿素荧光技术在植物环境胁迫研究中的应用[J].热带亚热带植物学报, 1995, 3(4):79-86. CHEN Yizhu, LI Xiaoping, XIA Li, et al. The application of chlorophyll fluorescence technique in the study of responses of plants to environmental stresses [J]. J Trop Subtrop Bot, 1995, 3(4): 79-86. [10] 胡文海, 肖宜安, 喻景权, 等.低夜温后不同光强对榕树叶片PSⅡ功能和光能分配的影响[J].植物研究, 2005, 25(2):159-162. HU Wenhai, XIAO Yian, YU Jingquan, et al. Effects of different light intensity after low night temperature stress on PSⅡ functions and absorbed light allocation in leaves of Ficus microcarpa [J]. Bull Bot Res, 2005, 25(2): 159-162. [11] 刘云峰, 秦洪文, 石雷, 等.水淹对水芹叶片结构和光系统Ⅱ光抑制的影响[J].植物学报, 2010, 45(4):426-434. LIU Yunfeng, QIN Hongwen, SHI Lei, et al. Effects of submergence on leaf anatomy and photoinhibition of photosystem Ⅱin Oenanthe javanica plants [J]. Chin Bull Bot, 2010, 45(4): 426-434. [12] 赵竑绯, 赵阳, 张驰, 等.模拟淹水对杞柳生长和光合特性的影响[J].生态学报, 2013, 33(3):898-906. ZHAO Hongfei, ZHAO Yang, ZHANG Chi, et al. Effect of flooding stress on growth and photosynthesis characteristics of Salix integra [J]. Acta Ecol Sin, 2013, 33(3): 898-906. [13] 李鹏民, 高辉远, STRASSER R J.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J].植物生理与分子生物学学报, 2005, 31(6):559-566. LI Pengmin, GAO Huiyuan, STRASSER R J. Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study [J]. J Plant Physiol Mol Biol, 2005, 31(6): 559-566. [14] 王树凤, 孙海菁, 陈益泰, 等.模拟干旱胁迫下弗吉尼亚栎苗木叶片相关生理参数的分析[J].南京林业大学学报(自然科学版), 2011, 35(6):6-10. WANG Shufeng, SUN Haijing, CHEN Yitai, et al. Analysis of physiological indexes of Quercus virginiana under drought stress [J]. J Nanjing For Univ Nat Sci Ed, 2011, 35(6): 6-10. [15] 胡俊靖, 陈卫军, 郭子武, 等.水分胁迫对竹子生理特性影响的研究进展[J].西南林业大学学报, 2015, 35(1): 91-95. HU Junjing, CHEN Weijun, GUO Ziwu, et al. Review of the water stress on the physiological characteristics of bamboo [J]. J Southwest For Univ, 2015, 35(1): 91-95. [16] 李在军, 蔡孔瑜, 陈桂芳, 等.干旱胁迫和复水对麻竹渗透调节物质及细胞膜透性的影响[J].四川林业科技, 2010, 31(3):55-59. LI Zaijun, CAI Kongyu, CHEN Guifang, et al. Effect of continuous drought stress and rewatering on osmo-regulation substances and cell membrane permeability in leaves of Dendrocalamus latiflorus [J]. J Sichuan For Sci Technol, 2010, 31(3): 55-59. [17] 应叶青, 郭璟, 魏建芬, 等.自然干旱胁迫及复水处理对红秆寒竹生理特性的影响[J].浙江林学院学报, 2010, 27(4): 513-517. YING Yeqing, GUO Jing, WEI Jianfen, et al. Physiological characteristics of Chimonobambusa marmoreal f. variegate with natural drought stress and rewetting [J]. J Zhejiang For Coll, 2010, 27(4): 513-517. [18] 应叶青, 郭璟, 魏建芬, 等.干旱胁迫对毛竹幼苗生理特性的影响[J].生态杂志, 2011, 30(2):262-266. YING Yeqing, GUO Jing, WEI Jianfen, et al. Effects of drought stress on physiological characteristics of Phyllostachys edulis seedlings [J]. Chin J Ecol, 2011, 30(2): 262-266. [19] 赵兰, 邢新婷, 江泽慧, 等. 4种地被观赏竹的抗旱性研究[J].林业科学研究, 2010, 23(2):221-226. ZHAO Lan, XING Xinting, JIANG Zehui, et al. Study on drought resistance of four dwarf ornamental bamboos [J]. For Res, 2010, 23(2): 221-226. [20] 顾大形, 陈双林.四季竹对土壤水分胁迫的生理适应[J].西北植物学报, 2012, 32(4): 751-758. GU Daxing, CHEN Shuanglin. Physiological adaptation of Oligostachyum lubricum under water stress [J]. Acta Bot Boreal-Occident Sin, 2012, 32(4): 751-758. [21] 张艳华, 刘国华, 王福升.淹水胁迫下5种竹子生理生化指标的变化[J].林业科技开发, 2009, 23(5):71-74. ZHANG Yanhua, LIU Guohua, WANG Fusheng. Variation of inner physiological and biochemical characteristics of five bamboos under flooding stress [J]. China For Sci Technol, 2009, 23(5): 71-74. [22] 刘玉芳, 陈双林, 李迎春, 等.河竹鞭根对长期淹水环境的生理响应[J].林业科学研究, 2014, 27(5):621-625. LIU Yufang, CHEN Shuanglin, LI Yingchun, et al. Physiological response of Phyllostchys rivalis rhizome roots to long-term water stress [J]. For Res, 2014, 27(5): 621-625. [23] 刘玉芳, 陈双林, 郭子武, 等.淹水对河竹鞭根系统生物量分配及异速生长模式的影响[J].林业科学研究, 2015, 28(4):502-507. LIU Yufang, CHEN Shuanglin, GUO Ziwu, et al. Effect of waterlogging on biomass allocation and allometric pattern of rhizome and root system of Phyllostachys rivalis [J]. For Res, 2015, 28(4): 502-507. [24] 刘玉芳, 陈双林, 李迎春, 等.淹水环境下河竹鞭根养分吸收和积累的适应性调节[J].生态学报, 2016, 36(10):2926-2933. LIU Yufang, CHEN Shuanglin, LI Yingchun, et al. Adaptive adjustment to nutrient absorption and accumulation of Phyllostachys rivalis rhizome-roots under waterlogged conditions [J]. Acta Ecol Sin, 2016, 36(10): 2926-2933. [25] DEMMING-ADAMS B, ADAMS Ⅲ W W, BARKER D H, et al. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation [J]. Physiol Plant, 1996, 98(2): 253-264. [26] 张会慧, 张秀丽, 王娟, 等.利用快相叶绿素荧光参数综合评价3种丁香的耐盐性[J].南京林业大学学报(自然科学版), 2013, 37(5):13-19. ZHANG Huihui, ZHANG Xiuli, WANG Juan, et al. A comprehensive evaluation of salt resistance in three clove varieties by the fast fluorescence transient parameters [J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(5): 13-19. [27] 王海珍, 陈加利, 韩路, 等.地下水位对胡杨Populus euphratica和灰胡杨Populus pruinosa叶绿素荧光光响应与光合色素含量的影响[J].中国沙漠, 2013, 33(4):1054-1063. WANG Haizhen, CHEN Jiali, HAN Lu, et al. Effects of groundwater levels on photosynthetic pigments and light response of chlorophyll fluorescence parameters of Populus euphratica and Populus pruinosa [J]. J Desert Res, 2013, 33(4): 1054-1063. [28] 吴雪霞, 陈建林, 查丁石.低温胁迫对茄子幼苗叶绿素荧光特性和能量耗散的影响[J].植物营养与肥料学报, 2009, 15(1):164-169. WU Xuexia, CHEN Jianlin, ZHA Dingshi. Effects of low temperature stress on chlorophyll fluorescence characteristics and excitation energy dissipation in eggplant seeding leaves [J]. Plant Nutr Fert Sci, 2009, 15(1): 164-169. [29] van KOOTEN O, SNEL J F. The use of chlorophyll fluorescence nomenclature in plant stress physiology [J]. Photosynth Res, 1990, 25(3): 147-150. [30] 师生波, 李天才, 李妙, 等.土壤干旱和强光对高山蒿草叶片PSⅡ反应中心非光化学猝灭的交互影响分析[J].植物生理学报, 2015, 51(10):1687-1686. SHI Shengbo, LI Tiancai, LI Miao, et al. Interaction effect analysis of soil drought and strong light on PSⅡ nonphotochemical quenching in Kobresia pygmaea leaves [J]. Plant Physiol J, 2015, 51(10): 1687-1686. [31] 王巧, 聂鑫, 刘秀梅, 等.遮光对松属3个树种幼树光合特性和荧光参数的影响[J].浙江农林大学学报, 2016, 33(4):643-651. WANG Qiao, NIE Xin, LIU Xiumei, et al. Photosynthetic characteristics and chlorophyll fluorescence of three Pinus tree species with shading [J]. J Zhejiang A & F Univ, 2016, 33(4): 643-651. [32] 徐凯, 郭延平, 张上隆, 等.草莓叶片光合作用对强光的响应及其机理[J].应用生态学报, 2005, 16(1):73-78. XU Kai, GUO Yanping, ZHANG Shanglong, et al. Response of strawberry leaves photosynthesis to strong light and its mechanism [J]. Chin J Appl Ecol, 2005, 16(1): 73-78. [33] 温国胜, 田海涛, 张明如, 等.叶绿素荧光分析技术在林木培育中的应用[J].应用生态学报, 2006, 17(10):1973-1977. WEN Guosheng, TIAN Haitao, ZHANG Mingru, et al. Application of chlorophyll fluorescence analysis in forest tree cultivation [J]. Chin J Appl Ecol, 2006, 17(10): 1973-1977. [34] 黄磊, 姜国斌, 朱玉, 等.高温对北高丛蓝莓叶片气孔交换及叶绿素荧光参数的影响[J].生态学杂志, 2016, 35(4):871-879. HUANG Lei, JIANG Guobin, ZHU Yu, et al. Effects of high temperature on leaf gas exchange and chlorophyll fluorescence parameters of the north highbush blueberry [J]. Chin J Ecol, 2016, 35(4): 871-879. [35] 宫丽丹, 魏丽萍, 倪书邦, 等.持续干旱对油棕幼苗叶绿素荧光动力学参数的影响[J].中国农业通报, 2016, 32(13):1-6. GONG Lidan, WEI Liping, NI Shubang, et al. Effect of persistent drought stress on chlorophyll fluorescence parameters in leaves of oil palm [J]. Chin Agric Sci Bull, 2016, 32(13): 1-6. [36] 周艳虹, 黄黎锋, 喻景权.持续低温弱光对黄瓜叶片气孔交换、叶绿素荧光猝灭和吸收光能分配的影响[J].植物生理与分子生物学学报, 2004, 30(2):153-160. ZHOU Yanhong, HUANG Lifeng, YU Jingquan. Effects of sustained chilling and low light on gas exchange, chlorophyll fluorescence quenching and absorbed light allocation in cucumber leaves [J]. J Plant Physiol Mol Biol, 2004, 30(2): 153-160. [37] ASADA K. The water-water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons [J]. Annu Rev Plant Physiol Plant Mol Boil, 1999, 50: 601-639. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.05.015
 点击查看大图
点击查看大图
计量
- 文章访问数: 3896
- HTML全文浏览量: 695
- PDF下载量: 378
- 被引次数: 0
