





-
开花是高等植物进入生殖生长的标志,是植物完成生活史的必要环节。在拟南芥Arabidopsis thaliana中的研究发现,开花过程受多种内源因素和外源环境因素的影响,由复杂基因网络严格调控[1]。光周期途径中植物开花受季节性昼长变化影响[2];赤霉素(GA)途径是在非诱导光周期途径的条件下,在开花过程中发挥作用[3];春化途径中植物开花要受到低温积累的影响[4];自主途径中通过感知植物自身内部发育状态而调控植物开花[5],多种途径的成花调节信号最终都集中于2个整合因子:开花促进因子FLOERING LOCUS T(FT)和SUPPRESSPR OF OVEREXPRESION OF CONSTANS 1(SOC1)[6-8]。在拟南芥中,过表达FT基因会明显促进SOC1基因的表达量;同时在ft突变体中,SOC1的表达量明显下降[9-11],但是对ft和soc1双突变体与ft单突变体植株的开花时间进行统计,结果显示没明显差异,可知除FT蛋白调控SOC1的表达外,还存在另外的途径影响SOC1基因的表达[10-13]。其他开花控制因子可以通过FT和SOC1参与开花途径的调控,例如开花抑制因子SHORT VEGETATIVE PHASE(SVP)可以结合SOC1和FT的调控序列,对FT和SOC1产生抑制作用[14],进而下调它们的表达量。近年,发现拟南芥中属于J蛋白家族一员的J3蛋白,可以与SVP蛋白相互作用调控SOC1和FT基因的表达量,而实现对成花信号的调节,在调控开花途径中发挥着十分重要的作用[15]。J蛋白(J-domain protein)是广泛存在于动植物中的一个庞大的分子伴侣家族,该类蛋白N末端包含1个约由70个氨基酸构成的保守序列——J结构域而被命名为J蛋白[16-17]。J蛋白的J结构域由4个α-螺旋组成,第2和第3个螺旋之间有一个极为保守的由组氨酸、脯氨酸、天冬氨酸组成的HPD三肽,这是J结构域的重要特征[18];邻近J结构域的是G/F结构域(富含甘氨酸和苯丙氨酸);之后是锌指结构域。根据结构域的不同J蛋白可分为4类[19]:Ⅰ类J蛋白包含所有常见的结构;Ⅱ类J蛋白缺少锌指结构;Ⅲ类J蛋白只有J结构域;Ⅳ类J蛋白也被称作J-like蛋白,这类蛋白的J结构域没有HPD模块[20-21]。目前发现拟南芥中共有120个J蛋白,其中Ⅰ类J蛋白8个,Ⅱ类J蛋白16个,Ⅲ类J蛋白92个,Ⅳ类J蛋白4个[22]。在植物中,有关J蛋白的研究主要集中在拟南芥中,现有的结果表明,J蛋白可以参与多种生物学过程,包括叶绿体的发育[23-25]、植物的抗逆性[26-27]、信号转导[28-29]和成花途径[15]等。在拟南芥开花调控中,J3蛋白通过与SVP等其他开花关键因子相互作用而影响植物成花过程,SVP的突变体svp-41表现为早花现象,J3的突变j3-1表现为晚花现象,同时在j3-1和svp-41的双突变体中,拟南芥表现出早花现象,与svp-41的单突变体的表型相似,这一结果表明J3基因通过与SVP的相互作用实现对开花时间的调控[30-31],进一步研究发现在细胞核中J3基因通过与营养生长短期(SVP)蛋白直接相互作用,衰减SVP蛋白与SOC1和FT调控序列的结合从而上调SOC1和FT基因的表达量[15]。竹类植物开花周期较长并具有时间上的不确定性,而且很多竹子开花后会成片死亡,雷竹Phyllostachys violascens是一种优良的笋用竹种,近年来,雷竹普遍开花,开花后部分雷竹会死亡,导致雷竹笋产量大幅度下降,带来严重的经济损失。有关竹类植物开花的研究已经很多,但是竹子成花机制和花期调控模式至今尚未完全清楚[32]。目前,竹子中还未见有关J3基因生物学功能的研究,本研究以雷竹为研究对象,通过同源克隆技术获得1个J3同源基因,经过亚细胞定位、表达模式和功能分析,以期能为深入研究J3同源基因在竹类植物开花调控机制中的作用提供科学依据。
-
雷竹材料来源于浙江农林大学智能温室大棚,选取位于同一竹鞭上的笋、花芽、花,开花和不开花雷竹的幼叶、成叶、茎秆,标记分装后放入液氮中速冻,保存在-80 ℃超低温冰箱。
-
用全RNA提取试剂盒(鼎国)分别提取采集的雷竹材料的RNA,以提取的RNA(少于0.5 g·L-1)为模板,用Prime ScriptTM RT Master Mix(Perfect Real Time)试剂盒(Clontech),反转录获得雷竹各组织的cDNA,检测合格后,于-20 ℃保存备用。
-
通过本地Blast,从毛竹Phyllostachys edulis基因组文库中找到一个候选基因序列号(bphylf027h21),根据该候选基因的基因序列设计引物(J3,表 1),雷竹cDNA为模板,进行聚合酶链式反应(PCR)扩增,琼脂糖凝胶电泳检测PCR扩增条带,用胶回收试剂盒(上海生工生物公司)回收纯化电泳条带,将获得的目的片段与pMD19(simple)-T载体置于连接仪上16 ℃连接1.0~3.0 h,之后转化大肠埃希菌Escherichia coli DH5α感受态细胞(热转化法),37 ℃恒温培养箱培养6.0~8.0 h,挑取白色圆形单菌落,菌落PCR鉴定出阳性单克隆,送测序,得到碱基序列。
表 1 本研究所用的引物
Table 1. PCR primers used in this study
引物名称 序列(5'→3') 用途 J3-F CTCTCCGTCTCACTCCGCTC 基因开放阅读框(ORF)扩增 J3-R GTCAAATTGCCTTTATAGAA PvJ3-F ATGTTCGGGCGCGCGCCG ORF扩增 PvJ3-R TTACTGTTGTGCGCACTGCAC PvJ3-1-F CCGGAATTCTGGAGAGAGCATTAACG 荧光定量PCR PvJ3-1-R CCGGAATTCCTTCTGCTGGAGGAC PeNTB-F TCTTGTTTGACACCGAAGAGGAG 荧光定量PCR内参引物 PeNTB-R AATAGCTGTCCCTGGAGGAGTTT PvJ3-2-F TCCCCCGGGATGTTCGGGCGCGCG 亚细胞定位 PvJ3-2-R CGCGGATCCCTGTTGTGCGCACTGCACTC PvJ3-3-F tccATCGATATGTTCGGGCGCGCG 过表达载体构建 PvJ3-3-R cgcGGATcCttaCTGTTGTGCGCACTGCACTC 用Vector NTI预测基因序列的开放阅读框(ORF)区,据此设计引物(PvJ3,表 1),重复上述实验过程,最终测序得到雷竹PvJ3基因序列。运用TMHMM和ProtScale分析PvJ3蛋白的结构特征和理化性质;Smart在线分析其蛋白的结构域;通过Clustal X2比对雷竹PvJ3与其他物种中的J3同源基因的氨基酸序列的相似性;并用Mega 5.0构建系统发育树,选择邻近法(NJ),bootstrap回复次数为1 000次。
-
用雷竹各组织部位的cDNA为模板,通过实时定量聚合酶链式反应(qRT-PCR),分析雷竹PvJ3基因的表达模式。根据实验要求设计目的基因的定量引物(PvJ3-1,表 1),毛竹的PeNTB基因作为内参基因[33]。各种雷竹组织做3次重复,避光条件下操作,参照SYBR Premix Ex TaqTMⅡ(Takara)试剂盒要求加样。PCR扩增程序(两步法):预变性95 ℃,5 min;95 ℃,30 s;60 ℃,30 s,循环数为40。反应结束后,分析确定数据的可靠性,选择合适的数据,用2-△△CT方法[34]对数据进行分析。
-
根据PvJ3基因ORF区序列(不含终止密码子)和pCaM35S-sGFP载体序列上的酶切位点,在目的基因ORF区的上下游分别引入酶切位点SmaI和BamHI,设计引物(PvJ3-2,表 1),提取pMD19-PvJ3质粒做为模板,进行PCR反应,胶回收纯化得到带有酶切位点的基因片段与pMD19(simple)-T载体连接转化后,阳性单克隆送测序。将测序正确的单克隆进行扩大培养,用质粒提取试剂盒(上海生工生物公司)提取质粒,用限制性内切酶SmaI和BamHI分别对pMD19-PvJ3和pCaM35S-sGFP载体质粒进行双酶切实验,之后用T4 DNA连接酶在16 ℃条件下过夜连接胶回收的酶切产物,连接产物转化大肠埃希菌DH5α感受态细胞,经测序正确,瞬时表达载体PvJ3-GFP构建成功。通过基因枪轰击洋葱Allium cepa表皮,25 ℃下避光暗培养1~2 d后,用荧光显微镜观察绿色荧光蛋白信号,金粉的准备和样品的制备参照文献[35]。
-
根据PvJ3基因ORF区序列和pC1301载体(含有35 S启动子)的序列设计引物(PvJ3-3,表 1),在目的基因ORF区的上下游同样分别引入酶切位点SmaI和BamHI,构建过表达载体pC1301-PvJ3,实验方法参照1.5节。
-
通过热转化法将构建成功的pC1301-PvJ3过表达载体转入农杆菌Agrobacterium tumefaciens GV3101感受态细胞中,提取阳性单克隆的质粒,重新转化大肠埃希菌DH5α感受态细胞,再次测序检验转入重组质粒是否正确。用已正确转入pC1301-PvJ3重组载体的农杆菌配制侵染液,侵染进入花期的野生型拟南芥花序[36],侵染3次,间隔约1周·次-1,侵染后的拟南芥即T0代植株,将消毒处理后的T0代种子均匀撒在加入潮霉素的1/2 MS(Murashige and Skoog)固体培养基上,潮霉素质量浓度为100.00 mg·L-1,按照V(潮霉素溶液):V(1/2MS培养基)=1.50:1 000.00比例加入培养基中,筛选抗性植株,以采用十六烷基三甲基溴化铵(CTAB)法[37]提取的抗性植株的基因组DNA为模板,PCR鉴定出T1代阳性植株,并对植株表型进行初步统计。
-
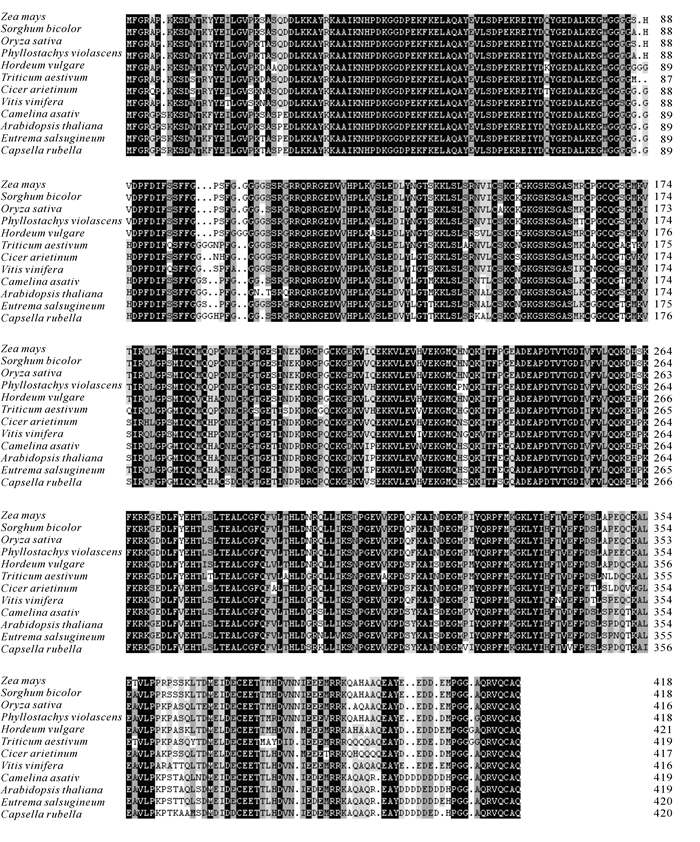
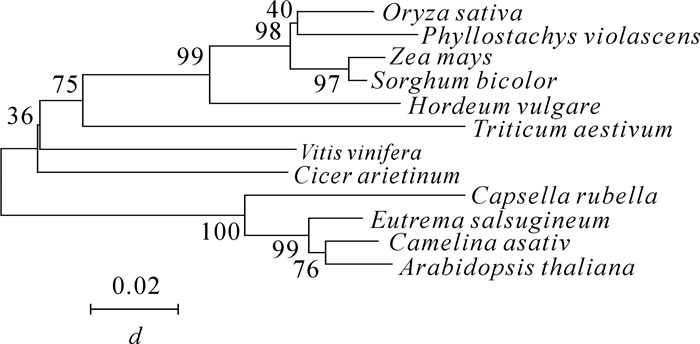
根据在毛竹转录组数据库中挑选的基因设计引物,利用同源克隆的方法,从雷竹中分离得到1个J3同源基因,命名为PvJ3,分析PvJ3基因的碱基序列,可知,其ORF区由1 260 bp个碱基组成,能编码419个氨基酸。经蛋白结构域和氨基酸序列分析(图 1~2),可知PvJ3蛋白的N端有J蛋白家族典型的J结构域,同时具有锌指结构域,C末端有法尼基化信号CAQQ序列。用ClustalX2软件对不同物种的J3同源蛋白的氨基酸进行序列比对,结果发现:J3蛋白在各类物种中的相似性很高,与拟南芥J3蛋白的一致性可达80.90%(图 2)。采用临近法构建系统进化树,结果显示:雷竹J3蛋白与单子叶植物高粱Sorghum bicolor和玉米Zea mays的亲缘关系最近,水稻Oryza sativa,大麦Hordeum vulgare和小麦Triticum aestivum次之,与双子叶拟南芥和亚麻荠Camelina sativa的亲缘关系最远(图 3)。

图 1 PvJ3蛋白结构示意图
Figure 1. Structure diagram of PvJ3

图 2 PvJ3与其同源基因的氨基酸序列比对
Figure 2. Alignment of the PvJ3 amino acid sequences from different plant species

图 3 不同物种J3蛋白的系统进化树分析
Figure 3. Phylogenetic tree analysis of J3 in different plant species
-
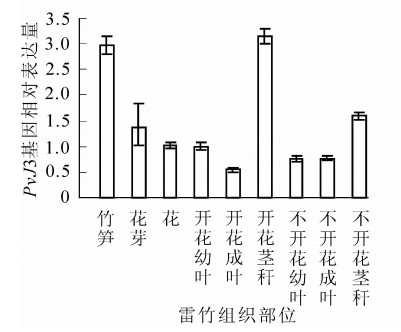
通过qRT-PCR技术,对同一竹鞭上开花和不开花雷竹中PvJ3基因在不同组织部位的表达水平进行分析。结果表明:PvJ3基因在同一竹鞭的笋、花芽、花,开花与不开花的茎秆、成叶、幼叶中均有表达。在开花雷竹的茎秆中PvJ3基因的表达量最高,茎、花芽、花、幼叶和成叶中的表达依次降低。在不开花雷竹的茎秆中PvJ3基因的表达量同样也是最高,幼叶、成叶中表达量基本一样(图 4)。

图 4 PvJ3在开花雷竹与不开花雷竹的不同部位表达水平
Figure 4. Expression of PvJ3 in different tissues of Phyllostachys violascens
-
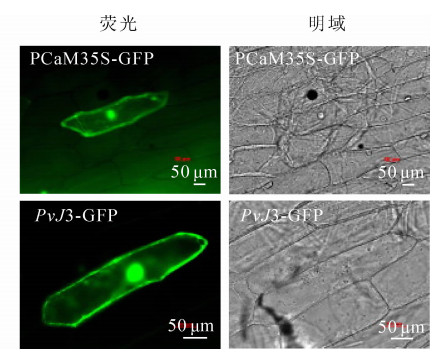
用基因枪轰击洋葱表皮细胞,通过荧光显微镜观察瞬时表达载体PvJ3-GFP绿色荧光蛋白的信号,确定PvJ3蛋白的定位情况(图 5)。pCaM35S-GFP对照在整个洋葱细胞里均有荧光信号,而PvJ3-GFP在细胞质核细胞核里有荧光信号,因此推断PvJ3蛋白主要定位在细胞质和细胞核中。

图 5 PvJ3蛋白亚细胞定位结果
Figure 5. Subcellular localization of PvJ3 fusion protein
-
构建过表达载体pC1301-PvJ3转化野生型拟南芥收取T0代种子,潮霉素筛选抗性植株,以抗性植株基因组DNA为模板,经PCR检测得到T1代拟南芥共37株。观察统计T1代植株中长势较好的30株转基因植株的表型,发现PvJ3基因的过表达植株开花时间明显早于野生型(图 6B)。统计结果显示:转基因植株开花天数比野生型植株显著提早(P=6.51E-15<0.01),平均提早3.50 d(图 6C),开花时莲座叶数目比野生型植株显著减少(P=9.24E-09<0.01),平均减少4.43叶(图 6D),且表现出多主茎现象(图 6A)。

图 6 PvJ3过表达拟南芥T1植株表型
Figure 6. Phenotypes of the PvJ3 transgenic Arabidopsis thaliana in T1 generation
-
高等植物成花过程存在多种诱导途径,大量基因参与这些途径的调控,成花关键基因的研究对于开花过程及花发育调控有着重要意义[38]。拟南芥中研究发现,SOC1和FT是感知不同开花途径信号的整合因子,SVP是直接调控FT和SOC1的转录调控因子[8, 39],J3蛋白通过与SVP直接相互作用而衰减SVP与SOC1和FT调控序列的结合从而上调SOC1和FT的表达量,参与拟南芥的成花过程,这一发现使得拟南芥成花调控网络更加完善[15]。本研究成功从雷竹中克隆得到J3同源基因并命名为PvJ3,雷竹PvJ3所编码的氨基酸序列与拟南芥J3蛋白氨基酸序列一致性达80.90%,对其蛋白结构域分析可知,PvJ3拥有相对保守的J结构域以及锌指结构域,可以说明PvJ3属于J家族中的一员。
为了研究雷竹PvJ3蛋白的功能,本研究通过qPCR技术和亚细胞定位分析该蛋白在雷竹中的表达模式和表达部位。对雷竹PvJ3基因进行了不同组织部位特异性表达分析,结果显示雷竹PvJ3基因在同一竹鞭的不同组织部位都有表达,已发现双子叶植物拟南芥J3基因同样在根、茎、叶、花芽、花和果荚等不同组织中表达[29],说明J3基因在单双子叶植物中均表现出全株表达的现象。在相对表达量上,PvJ3基因在开花雷竹茎秆中表达量最高,在花芽和花中的表达量降低,这与已报道的拟南芥中J3基因在花中表达量最高[15]的结果存在差异。PvJ3蛋白亚细胞定位结果显示该蛋白定位在细胞质和细胞核,与拟南芥中J3蛋白的亚细胞定位结果相一致[15],拟南芥中和J3蛋白具有互作关系的SVP,SOC1和FT蛋白都是核蛋白,说明雷竹PvJ3可能在细胞核中与这些开花整合因子相互作用,从而对开花途径进行调控。
将PvJ3基因导入野生型拟南芥植株验证其功能,T1代转基因植株的表型统计结果表明:转基因拟南芥植株开花时间明显早于野生型植株,开花时莲座叶数量明显减少(图 6 C,图 6D),而拟南芥J3基因的过表达植株和野生型植株的开花时间并没有明显差别[15],这说明雷竹PvJ3基因可以显著影响花期,可能是一个开花促进因子。值得注意的是,转雷竹PvJ3基因拟南芥植株同时表现出多主茎现象(图 6A),这一现象在拟南芥J3过表达植株中并没有被观测到[15]。以上结果表明,雷竹PvJ3基因不仅在成花调控中的功能和拟南芥J3基因存在一定差异,同时还可能参与茎的发育等其他生长发育过程,在基因功能上比拟南芥同源基因要复杂的多。
目前,J3基因功能的有关研究主要以模式植物拟南芥为研究对象,单子叶植物中还未见报道。通过对比J3基因在雷竹和拟南芥中的表达模式不难发现:雷竹PvJ3基因和拟南芥J3基因都是全株表达,同时编码蛋白都定位在细胞质和细胞核,这表明J3基因在不同植物中的表达部位不存在明显差异,并且可能是一个核内作用蛋白。在相对表达量上,雷竹PvJ3基因和拟南芥J3基因存在一定差异,同一竹鞭上开花和不开花雷竹中PvJ3基因在茎秆中表达量最高,而拟南芥中J3基因表达量在花中最高,这可能与雷竹PvJ3基因参与茎的发育有关。转基因实验的结果表明:雷竹PvJ3基因不仅可以显著提早拟南芥开花时间,同时还促使拟南芥出现多主茎现象,这表明雷竹PvJ3基因很可能参与除了开花调节以外的其他生长发育过程的调控。由于拟南芥和竹子之间的差异很大,雷竹PvJ3基因在竹子中是否可以促进植物提早开花,同时还参与哪些生物学过程,需要后续研究进一步验证。
Cloning and functional analysis of the PvJ3 gene from Phyllostachys violascens
-
摘要: 拟南芥Arabidopsis thaliana中J3基因参与花期的调节,调控开花时间。为了探究竹类植物中J3基因的功能,根据植物中J同源基因的高度保守性,采用同源克隆技术从雷竹Phyllostachys vilascens中克隆出1个J3基因,命名为PvJ3。该基因的开放阅读框(ORF)区长为1 260 bp,编码419个氨基酸。通过同源比对和系统进化树分析,可知雷竹PvJ3蛋白和其他物种中J3同源蛋白的氨基酸序列同源性很高。通过实时定量聚合酶链式反应(qRT-PCR),发现PvJ3基因在同一竹鞭的笋、花芽、花,开花与不开花雷竹的茎秆、成叶、幼叶中都有表达,且开花雷竹茎秆中表达量最高,表明PvJ3基因在雷竹各组织部位都有表达。亚细胞定位分析结果显示,PvJ3蛋白定位在细胞质和细胞核。通过转化拟南芥对其功能进行了初步分析,表型统计结果显示,PvJ3转基因拟南芥与野生型相比,开花时间明显提早,而且出现多主茎现象,表明PvJ3可以促使植物提前开花并且参与植物茎秆的发育。Abstract: The J3 gene mediates the integration of flowering signals in Arabidopsis thaliana. To explore the functions of the J3 gene in bamboo plants, a J3 homolog named PvJ3 was isolated from Phyllostachys violascens, and a preliminary analysis of its function was conducted with Ph. violascens. Analyses included a sequencing alignment and phylogenetic analysis, real-time quantitative PCR, and protein subcellular localization. Results showed that the phenotype of overexpression of PvJ3 in Ph. violascens predated flowering and had more main stems. The full-length ORF of the PvJ3 in Ph. violascens was 1 260 bp which encoded 419 amino acids. The sequencing alignment and phylogenetic analysis of PvJ3 showed a high homology with other species. Real-time quantitative PCR results showed that the PvJ3 was expressed in all organs both in flowering and non-flowering bamboo plants and was highest in the stem of flowering bamboo plants. Protein subcellular localization showed that PvJ3 was located in the cytoplasm and cell nucleus. These results suggest that PvJ3 may promote flowering, may participate in stem development, and may provide a scientific basis for the regulation of bamboo flowering.
-
Key words:
- forest tree breeding /
- Phyllostachys violascens /
- PvJ3 /
- subcellular localization /
- functional analysis
-
图 2 PvJ3与其同源基因的氨基酸序列比对
Figure 2 Alignment of the PvJ3 amino acid sequences from different plant species
图 3 不同物种J3蛋白的系统进化树分析
Figure 3 Phylogenetic tree analysis of J3 in different plant species
图 4 PvJ3在开花雷竹与不开花雷竹的不同部位表达水平
Figure 4 Expression of PvJ3 in different tissues of Phyllostachys violascens
图 6 PvJ3过表达拟南芥T1植株表型
Figure 6 Phenotypes of the PvJ3 transgenic Arabidopsis thaliana in T1 generation
表 1 本研究所用的引物
Table 1. PCR primers used in this study
引物名称 序列(5'→3') 用途 J3-F CTCTCCGTCTCACTCCGCTC 基因开放阅读框(ORF)扩增 J3-R GTCAAATTGCCTTTATAGAA PvJ3-F ATGTTCGGGCGCGCGCCG ORF扩增 PvJ3-R TTACTGTTGTGCGCACTGCAC PvJ3-1-F CCGGAATTCTGGAGAGAGCATTAACG 荧光定量PCR PvJ3-1-R CCGGAATTCCTTCTGCTGGAGGAC PeNTB-F TCTTGTTTGACACCGAAGAGGAG 荧光定量PCR内参引物 PeNTB-R AATAGCTGTCCCTGGAGGAGTTT PvJ3-2-F TCCCCCGGGATGTTCGGGCGCGCG 亚细胞定位 PvJ3-2-R CGCGGATCCCTGTTGTGCGCACTGCACTC PvJ3-3-F tccATCGATATGTTCGGGCGCGCG 过表达载体构建 PvJ3-3-R cgcGGATcCttaCTGTTGTGCGCACTGCACTC  下载: 导出CSV
下载: 导出CSV
-
[1] QUESADA V, DEAN C, SIMPSON G G, et al. Regulated RNA processing in the control of Arabidopsis flowering[J]. Int J Dev Biol, 2005, 49(5/6):773-780. [2] SIMPSON G G, DEAN C. Arabidopsis, the Rosetta stone of flowering time?[J]. Science, 2002, 296(5566):285-289. [3] LIU Chang, CHEN Hongyan, HONG Linger, et al. Direct interaction of AGL24 and SOC1 integrates flowering signals in Arabidopsis[J]. Development, 2008, 135(8):1481-1491. [4] DENNIS E S, PEACOCK W J. Epigenetic regulation of flowering[J]. Curr Opin Plant Biol, 2007, 10(5):520-527. [5] BOSS P K, BASTOW R M, MYLNE J S, et al. Multiple pathways in the decision to flower:enabling, promoting, and resetting[J]. Plant Cell, 2004, 16(suppl):S18-S31. [6] WANG Cunxi, TIAN Qing, HOU Zhenglin, et al. The Arabidopsis thaliana AT PRP39-1 gene, encoding a tetratricopeptide repeat protein with similarity to the yeast pre mRNA processing protein PRP39, affects flowering time[J]. Plant Cell Rep, 2007, 26(8):1357-1366. [7] MOURADOV A, CREMER F, COUPLAND G. Control of flowering time interacting pathways as a basis for diversity[J]. Plant Cell, 2002, 14(suppl):S111-S130. [8] PENG Zhenhua, LU Ying, LI Lubin, et al. The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla)[J]. Nat Gen, 2013, 45(4):456-461. [9] TORTI S, FORNARA F. AGL24 acts in concert with SOC1 and FUL during Arabidopsis floral transition[J]. Plant Sign Behav, 2012, 7(10):1251-1254. [10] NAVARRO C, CRUZ-ORÓE, PRAT S. Conserved function of FLOWERING LOCUS T (FT) homologues as signals for storage organ differentiation[J]. Curr Opin Plant Biol, 2015, 23(1):45-53. [11] YAMASHINO T, YAMAWAKI S, HAGUI E, et al. Clock-controlled and FLOWERING LOCUS T (FT)-dependent photoperiodic pathway in Lotus japonicus Ⅰ:verification of the flowering-associated function of an FT homolog[J]. Biosci Biotechnol Biochem, 2010, 77(4):747-753. [12] NAKANO Y, HIGUCHI Y, YOSHIDA Y, et al. Environmental responses of the FT/TFL1 gene family and their involvement in flower induction in Fragaria×ananassa[J]. J Plant Physiol, 2015, 177(24):60-66. [13] LEE J, LEE I. Regulation and function of SOC1, a flowering pathway integrator[J]. J Exp Bot, 2010, 61(9):2247-2254. [14] LEE J H, YOO S J, PARK S H, et al. Role of SVP in the control of flowering time by ambient temperature in Arabidopsis[J]. Genes Dev, 2007, 21(4):397-402. [15] SHEN Lisha, KANG Y G G, LIU Lu, et al. The J-domain protein J3 mediates the integration of flowering signals in Arabidopsis[J]. Plant Cell, 2011, 23(2):499-514. [16] RAJAN V, D'SILVA P. Arabidopsis thaliana J-class heat shock proteins:cellular stress sensors[J]. Funct Int Genom, 2009, 9(4):433-446. [17] MIERNYK J A. The J-domain proteins of Arabidopsis thaliana:an unexpectedly large and diverse family of chaperones[J]. Cell Stress Chap, 2001, 6(3):209-218. [18] QIAN Yanqiu, PATEL D, HARTL F U, et al. Nucleic magnetic resonance solution structure of the human Hsp40(HDJ-1) J-domain[J]. J Mol Biol, 1996, 260(2):224-235. [19] CRAIG E A, HUANG P, ARON R, et al. The diverse roles of J-proteins, the obligate Hsp70 co-chaperone[J]. Rev Physiol Biochem Pharmacol, 2006, 156(1):1-21. [20] MARTINEZ-YAMOUT M, LEGGE G B, ZHANG Ouwen, et al. Solution structure of the cysteine-rich domain of the Escherichia coli chaperone protein DnaJ[J]. J Mol Biol, 2000, 300(4):805-818. [21] MIERNYK J A. The J-domain proteins of Arabidopsis thaliana:an unexpectedly large and diverse family of chaperones[J]. Cell Stress Chap, 2001, 6(3):209-218. [22] SALAS-MUÑOZ S, RODRÍGUEZ-HERNÁNDEZ A A, ORTEGA-AMARO M A, et al. Arabidopsis AtDjA3 null mutant shows increased sensitivity to Abscisic acid, salt, and osmotic stress in germination and post-germination stages[J]. Plant Sci, 2016, 7(2):220. doi:10.3389/fpls.2016.00220. [23] VITHA S, FROEHLICH J E, KOKSHAROVA O, et al. ARC6 is a J-domain plastid division protein and an evolutionary descendant of the cyanobacterial cell division protein Ftn2[J]. Plant Cell, 2003, 15(8):1918-1933. [24] CHEN Kunming, HOLMSTROM M, RAKSAJIT W, et al. Small chloroplast-targeted DnaJ proteins are involved in optimization of photosynthetic reactions in Arabidopsis thaliana[J]. BMC Plant Biol, 2010, 10(1):43. doi:10.1186/1471-2229-10-43. [25] SHIMADA H, MOCHIZUKI M, OGURA K, et al. Arabidopsis cotyledon-specific chloroplast biogenesis factor CYO1 is a protein disulfide isomerase[J]. Plant Cell, 2007, 19(10):3157-3169. [26] SEKI M, NARUSAKA M, ABE H, et al. Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray[J]. Plant Cell, 2001, 13(1):61-72. [27] CHAI Tuanyao, ZHANG Yuxiu, ZHAO Wenming. Cloning of cDNA and expression analysis of DnaJ-like gene under heavy metal stress in bean[J]. Progr Nat Sci, 2000, 10(2):135-140. [28] SEDBROOK J C, CHEN Rujin, MASSON P H. ARG1(altered response to gravity) encodes a DnaJ-like protein that potentially interacts with the cytoskeleton[J]. Proc Natl Acad Sci USA, 1999, 96(3):1140-1145. [29] GUAN Changhui, ROAEN E S, BOONSIRICHAI K, et al. The ARG1-LIKE2 gene of Arabidopsis functions in a gravity signal transduction pathway that is genetically distinct from the PGM pathway[J]. Plant Physiol, 2003, 133(1):100-112. [30] THEIβEN G, KIM J T, SAEDLER H. Classification and phylogeny of the MADS-box multigene family suggest defined roles of MADS-box gene subfamilies in the morphological evolution of eukaryotes[J]. J Mol Evol, 1996, 43(5):484-516. [31] WIGGE P A, KIM M C, JAEGER K E, et al. Integration of spatial and temporal information during floral induction in Arabidopsis[J]. Science, 2005, 309(5737):1056-1059. [32] de CARVALHO A L, NELSON B W, BIANCHINI M C, et al. Bamboo-dominated forests of the southwest Amazon:detection, spatial extent, life cycle length and flowering waves[J]. PLoS One, 2013, 8(1):e54852. doi:10.1371/journal.pone/0054852. [33] 齐飞艳, 胡陶, 彭镇华, 等.毛竹实时荧光定量PCR内参基因的筛选及成花基因PheTFL1表达分析[J].西北植物学报, 2013, 33(1):48-52. QI Feiyan, HU Tao, PENG Zhenhua, et al. Screening of reference genes used in qRT-PCR and expression analysis of PheTFL1 gene in moso bambo[J]. Acta Bot Boreal-Occident Sin, 2013, 33(1):48-52. [34] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△Cr method[J]. Methods, 2001, 25(4):402-408. [35] 王小利, 吴佳海, 刘晓霞, 等.高羊茅春化基因FaVRN1亚细胞定位与差异表达分析[J].基因组学与应用生物学, 2011, 30(2):152-158. WANG Xiaoli, WU Jiahai, LIU Xiaoxia, et al. Subcellular localization and differential expression analysis of vernalizational gene FaVRN1 in tall fescue[J]. Genomics Appl Biol, 2011, 30(2):152-158. [36] CLOUGH S J, BENT A F. Floral dip:a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana[J]. Plant J Cell Mol Biol, 1998, 16(6):735-743. [37] 高志民, 范少辉, 高健, 等.基于CTAB法提取毛竹基因组DNA的探讨[J].林业科学研究, 2006, 19(6):725-728. GAO Zhimin, FAN Shaohui, GAO Jian, et al. Extract genomic DNA form Phyllostachys edulis by CTAB-based method[J]. For Res, 2006, 19(6):725-728. [38] 张素芝, 左建儒.拟南芥开花时间调控的研究进展[J].生物化学与生物物理进展, 2006, 33(4):301-309. ZHANG Suzhi, ZUO Jianru. Advance in the flowering time control of Arabidopsis[J]. Prog Biochem Biophys, 2006, 33(4):301-309. [39] LI Dan, LIU Chang, SHEN Lisha, et al. A repressor complex governs the integration of flowering signals in Arabidopsis[J]. Dev Cell, 2008, 15(1):110-120. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2018.02.014
 点击查看大图
点击查看大图

计量
- 文章访问数: 3159
- HTML全文浏览量: 623
- PDF下载量: 339
- 被引次数: 0
