




-
长江流域发源于唐古拉山脉,自西向东流经青藏高原、横断山区、云贵高原、四川盆地、江南丘陵和长江中下游平原,注入东海[1]。地域辽阔,横跨中国东部、中部和西部地区,包括19个省(直辖市和自治区)。长江流域地形复杂,地势西高东低呈阶梯状分布。气候类型复杂,除青藏高原为高原山地气候、西南热带季风气候外,主要为亚热带季风气候[2]。夏季高温多雨,冬季温和少雨,四季分明,年降水量在1 000 mm以上,这为生物提供了良好的栖息环境,成为生物多样性极为丰富的地区。长江流域种子植物约占全国种子植物的50%,珍稀濒危植物和国家重点保护植物共占全国珍稀濒危保护植物的39.7%。
兰科Orchidaceae植物因花期长、花朵造型奇异和色彩绚丽,具有极高的观赏价值[3];入药具有清热解毒、滋阴润肺等功效,药用价值也较高[4]。兰科植物对环境的依赖性比较强[5],易受到人类活动的影响。全世界所有的野生兰科植物均被列为《野生动植物濒危物种国际贸易公约》(CITES)的保护范围,为野生植物保护的旗舰类群[6]。目前,对兰科植物分布格局的研究主要有垂直分析和水平分析2种方法。在垂直尺度上,将区域性的山地海拔高度分段,分析兰科植物的分布格局以及区系特征,如藏东南[7]、滇西北[8]、西藏南迦巴瓦[9]、西双版纳[10]等。通过计算兰科植物属和种的丰富度,分析兰科植物丰富度的分布中心和变化趋势,进而研究水平意义上的分布格局,如滇西北[11]和全国兰科植物[12]的分布格局。也有学者采用地理信息系统(GIS)空间分析方法分析地区多物种的分布范围[13]和空间分布特征[14],揭示物种空间分布规律,达到维护当地生物多样性的稳定[15-16]和预防外来物种入侵的目的[17-18]。
总的来看,关于长江流域兰科植物的空间分布格局研究较少,且根据濒危程度进行长时序分析还不多见。基于此,本研究采用核密度、空间自相关和热点分析等GIS空间分析方法,分析了长江流域代表性濒危兰科植物的空间分布格局,为更有效地保护兰科植物提供理论依据。
-
全球生物多样性信息资讯机构(GBIF)是由世界各国政府资助的国际资讯机构。该网站收集了兰科植物的分布地区、物种名称、物种出现时间、物种经纬度坐标等信息。根据《中国物种红色名录》[19]的物种濒危等级,以及1981—2019年在长江流域分布的物种,选取了130个(10种)兰科植物空间分布点(表1),所选取的代表性10种兰科植物均有观赏和药用价值。
表 1 代表性兰科植物的分布
Table 1. Distribution of representative orchids
序号 属名 物种名 生境 生活型 濒危等级 1 舌唇兰属 Platanthera 小舌唇兰 Platanthera minor 山坡林下或草地 地生 近危 2 头蕊兰属 Cephalanthera 金兰 Cephalanthera falcata 山坡林下 地生 近危 3 兰属 Cymbidium 建兰 Cymbidium ensifolium 山坡林下 地生 易危 4 兰属 蕙兰 Cymbidium faberi 向南山坡或黄山松 Pinus taiwanensis 林下 地生 易危 5 虾脊兰属 Calanthe 反瓣虾脊兰 Calanthe reflexa 常绿阔叶林下、山谷溪边 地生 无危 6 虾脊兰属 钩距虾脊兰 Calanthe graciliflora 山坡林下或石壁上 地生 易危 7 兜被兰属 Neottianthe 二叶兜被兰 Neottianthe cucullata 针叶林下或高山草甸上 地生 近危 8 无柱兰属 Amitostigma 无柱兰 Amitostigma gracile 岩石上或沟边阴湿草地上 地生 近危 9 独蒜兰属 Pleione 独蒜兰 Pleione bulbocodioides 苔藓覆被的岩石上 附生 易危 10 风兰属 Neofinetia 风兰 Neofinetia falcata 山中林地树干上 附生 濒危 -
将兰科植物按照《中国物种红色名录》[19]标准,根据极危、濒危、易危、近危、无危的濒危等级,分别赋值为5、4、3、2、1,濒危程度越严重,赋值越大。对代表性兰科植物根据全球气候变暖速度划分为2个时段进行研究:1981—1997年为变暖加速期,1998—2019年为变暖暂缓期[20-22]。采用ArcGIS软件的核密度方法定性分析2个时间段内兰科植物聚集分布特征。采用空间自相关分析对2个时间段的兰科植物濒危等级进行全局莫兰指数(Moran’s I)计算。采用Open Geoda软件对代表性兰科植物计算局部莫兰指数,以标准化的濒危属性值作为横轴,相邻濒危属性值的加权平均值作为纵轴,分别绘制1981—1997年和1998—2019年的莫兰指数散点图。使用Open Geoda软件的热点分析值(
$G_i^{*} $ ),分析兰科植物濒危等级的热点分析,得出1981—1997年(置信度95%)和1998—2019年(置信度99%)的热点分布表[23],并将分析结果进行可视化显示。 -
核密度函数值可反映兰科植物在长江流域的空间聚集和分散特征,函数值越高,说明兰科植物聚集程度越高,且值随中心辐射距离的增大而逐渐变小[24]。计算表达式如下:
$$ f_{n}(y)=\frac{b}{n h} \sum_{b=1}^{n} k\left(\frac{y-y_{b}}{h}\right)。 $$ (1) 式(1)中:
$ f_{n}(y)$ 为兰科植物n个空间分布点的核密度测算值,n=130;k为核密度常数;h为核密度测算带宽的平滑参数;$ \left(y-y_{b}\right)$ 为兰科植物估计值y与分布点b样本值yb之间的空间距离。参考相关研究成果的基础上[25],得出兰科植物的密度空间分布特征。 -
全局莫兰指数表示地理对象属性值在全部空间范围内的聚集程度[26],是对属性在整个空间分布区域的特征刻画。计算表达式如下:
$$ {I}=\frac{m \displaystyle \sum_{i=1}^{m} \displaystyle \sum_{j=1}^{m} {\boldsymbol{w}}_{{\boldsymbol{i j}}}\left(x_{i}-\bar{x}\right)\left(x_{j}-\bar{x}\right)}{\displaystyle \sum_{i=1}^{m} \displaystyle \sum_{j=1}^{m} {\boldsymbol{w}}_{{\boldsymbol{i j}}} \displaystyle \sum_{i=1}^{m}\left(x_{i}-\bar{x}\right)^{2}} 。 $$ (2) 式(2)中:I为濒危兰科植物的全局莫兰指数值;m为空间单元数量;xi、xj分别为第i个空间单元和第j个空间单元兰科植物的濒危等级值;
$\bar x $ 为濒危等级均值;wij为空间单元i、j的空间权重矩阵。标准化Z值检验全局莫兰指数自相关水平,当Z>1.96或Z<−1.96 (α=0.05)时,说明濒危兰科植物在空间上存在显著的空间自相关水平。
-
局部空间自相关可以检测局部区域是否存在变量聚集的现象[26],进一步明确相邻兰科植物生存状况的分布关系,弥补全局空间自相关不能确定具体聚集区域的不足。热点分析可以进一步分析兰科植物局部空间自相关关系。经过反复试验,莫兰指数散点图采用“Queen’s”原则,判断是否邻接。计算表达式如下:
$$ G_{i}^{*}=\frac{\displaystyle \sum_{j=1}^{m} {\boldsymbol{w}}_{{\boldsymbol{i j}}} x_{j}}{\displaystyle \sum_{j=1}^{m} x_{j}} 。$$ (3) 式(3)中:
$ G_{i}^{*}$ 为兰科植物濒危等级的热点分析值;${\boldsymbol{w}}_{{\boldsymbol{i j}}}$ 为空间单元i、j的空间权重矩阵;xj为第j个空间单元兰科植物的濒危等级值;m为空间单元数量。 -
通过核密度分析结果可知:1981—2019年,兰科植物的空间分布整体上呈“多核破碎化”,但1998年后,破碎化程度有所减弱。在四川分布的代表性兰科植物北移,数量减少。318国道的建设经过康定县和泸定县,这些地区早期发现有大量兰科植物分布,如今兰科植物减少,这在一定程度上说明兰科植物受到国道建设的影响(表2),国道建设加剧了人类活动,缩减了兰科植物的生存空间。
表 2 1981—2019年四川代表性兰科植物的空间分布
Table 2. Spatial distribution of representative Orchidaceae plants in Sichuan from 1981 to 2019
年份 地区 经纬度 代表性兰科植物 年份 地区 经纬度 代表性兰科植物 1994 平武县 32°26′24″N,104°33′00″E 钩距虾脊兰 1981 泸定县 29°54′50″N,102°14′02″E 反瓣虾脊兰 1983 雷波县 28°15′43″N,103°34′15″E 金兰 1984 泸定县 29°54′50″N,102°14′02″E 反瓣虾脊兰 1984 泸定县 29°54′50″N,102°14′02″E 金兰 1997 泸定县 29°30′00″N,101°54′00″E 二叶兜被兰 1984 泸定县 29°54′50″N,102°14′02″E 金兰 1997 康定县 30°06′00″N,101°48′00″E 二叶兜被兰 1981 泸定县 29°54′50″N,102°14′02″E 建兰 1997 康定县 30°08′45″N,101°51′36″E 二叶兜被兰 1981 泸定县 29°54′50″N,102°14′02″E 建兰 1997 泸定县 29°34′30″N,101°59′56″E 二叶兜被兰 1986 九寨沟县 33°15′36″N,104°13′48″E 蕙兰 2019 成都市青羊区 30°40′55″N,104°00′36″E 蕙兰 1982 泸定县 29°54′36″N,102°13′48″E 反瓣虾脊兰 2005 翁达自然保护区 31°54′00″N,100°54′00″E 二叶兜被兰 1983 九寨沟县 33°15′36″N,104°13′48″E 反瓣虾脊兰 2007 阿坝藏族羌族自治区金川县 31°42′00″N,102°00′00″E 二叶兜被兰 1981 泸定县 29°54′50″N,102°14′02″E 反瓣虾脊兰 2005 翁达自然保护区 31°58′55″N,100°57′50″E 二叶兜被兰 1984 北川县 31°53′34″N,104°26′09″E 反瓣虾脊兰 2005 翁达自然保护区 31°58′55″N,100°57′50″E 二叶兜被兰 1984 北川县 31°53′34″N,104°26′09″E 反瓣虾脊兰 从兰科植物分布密度来看,中密度分布在湖北中部、湖南中部、重庆中部、贵州东部和北部、四川中部和北部、云南北部、甘肃南部;高密度分布在湖南西部、湖北西南部、广西东北部、重庆东南部和贵州东北部。中密度分布省份(直辖市和自治区)数量未减少,且分布面积增加了1 969 m2 (2.9%);高密度分布省份(直辖市和自治区)数量减少,少了广西和贵州,且分布面积减少了222 m2 (0.32%)。高密度栖息地数量和面积减少,兰科植物生存受到威胁。研究发现:南岭地区存在村民采挖和贩卖观赏兰花的现象,并且南岭地区修建了大量水电站[27],改变了兰科植物的生长环境,这对环境要求严格的兰科植物来说,无疑产生了重要影响(表3)。贵州兰科植物主要分布在铜仁地区的松桃、桐梓、印江、石阡,该地区矿产资源丰富,大量的矿产开发会铲除覆盖地表植被,破坏兰科植物生长环境,再加上村民采挖兰属和虾脊兰属植物,导致兰科植物在贵州的分布发生明显变化(表3)。
表 3 1981—2019年贵州和广西代表性兰科植物的空间分布
Table 3. Spatial distribution of representative Orchidaceae plants in Guizhou and Guangxi from 1981 to 2019
年份 省份 地区 经纬度 代表性兰科植物 1986 贵州 松桃苗族自治县 28°09′00″N,109°12′00″E 钩距虾脊兰 1996 贵州 桐梓县 28°07′48″N,106°49′12″E 金兰 1988 贵州 印江土家族
苗族自治县27°59′24″N,108°24′00″E 小舌唇兰 1988 贵州 松桃苗族自治县 28°00′00″N,109°12′00″E 蕙兰 1988 贵州 石阡县 27°30′36″N,108°13′48″E 反瓣虾脊兰 1988 贵州 施秉县 27°01′48″N,108°11′24″E 反瓣虾脊兰 1981 广西 金秀县老山伐木场十六公里阴冲 24°10′30″N,110°14′06″E 钩距虾脊兰 1991 广西 兴安县猫儿山老山界 25°36′03″N,110°35′06″E 金兰 1984 广西 资源县猫儿山万亩林场 26°00′18″N,110°35′56″E 金兰 1992 广西 资源县塘洞猫儿山大竹坪 26°00′18″N,110°35′56″E 金兰 1982 广西 兴安县猫儿山梯子岭 25°36′03″N,110°35′06″E 金兰 1982 广西 金秀县 24°10′30″N,110°14′06″E 小舌唇兰 1994 广西 环江县川山乡大沙坡 25°06′25″N,108°18′54″E 独蒜兰 2016 广西 百色市那坡县 23°20′45″N,105°54′46″E 建兰 1998 广西 龙胜各族自治县花坪红毛界下界 25°44′31″N,110°02′24″E 反瓣虾脊兰 1998 广西 龙胜各族自治县花坪红毛界下界 25°44′31″N,110°02′24″E 反瓣虾脊兰 1998 广西 金秀老山银杉保护区 24°10′30″N,110°14′06″E 钩距虾脊兰 1998 广西 金秀老山银杉保护区 24°10′30″N,110°14′06″E 钩距虾脊兰 说明:银杉Cathaya argyrophylla 气温是影响植物生长的自然因素之一,气温与植物的光合作用、呼吸作用和代谢作用密不可分[28]。经研究发现:1981—1997年和1998—2019年兰科植物中、高密度区存在较明显的变化,这与1998年之后全球变暖速度进入“停滞”相吻合[21-22],说明长江流域兰科植物分布位置和数量变化也可能与全球变暖有关。
-
全局莫兰指数可以表示要素的空间自相关性,当Z>1.96时,说明莫兰指数值具有明显的聚类特征。从表4看出:1981—1997年濒危兰科植物全局莫兰指数为0.241,1998—2019年为0.805,通过了显著性水平α=0.05的检验(Z>1.96)。说明这2个时段的濒危兰科植物在空间上呈正相关,具有明显的空间集聚特征。1998—2019年的莫兰指数大于1981—1997年,且更趋向于1,表明1998—2019年濒危兰科植物的空间自相关性水平显著增强,生存空间急剧缩减,抱团分布的现象更加明显。
表 4 1981—2019年兰科植物濒危等级的全局莫兰指数
Table 4. Global Moran’ s I index of the endangered species of representative Orchidaceae plants from 1981 to 2019
时段 莫兰指数 Z 域值(α=0.05) 1981—1997年 0.241 3.727 1.96 1998—2019年 0.805 4.450 1.96 -
图1表明:2个时段的局部莫兰指数均大于0,与全局莫兰指数结果相符。趋势线主要分布在第1和第3象限,说明兰科植物的濒危等级值在2个时段都存在高—高值聚集和低—低值聚集。图1A中,落入第1象限的兰科植物濒危等级值有22个,占总点数的25.0%,落入第3象限的有48个,占总点数的54.6%。图1B中,落入第1象限的兰科植物濒危等级值有21个,占总点数的50.0%,落入第3象限的有16个,占总点数的38.1%。可见,兰科植物在1981—1997年表现出明显的低—低聚集,1998—2019年表现出明显的高—高聚集,说明兰科植物局部空间异质性增强,且处于高风险区的兰科植物数量明显增加。在莫兰散点图趋势线以下,图1A数据点的数量为55个,占总点数的62.5%,图1B数据点的数量为18个,占总点数的42.9%。1998—2019年与1981—1997年相比,数据点的数量明显减少。兰科植物濒危等级值的“核”效应明显减弱,“多核破碎化”的态势也在弱化,这与核密度分析结果相吻合。
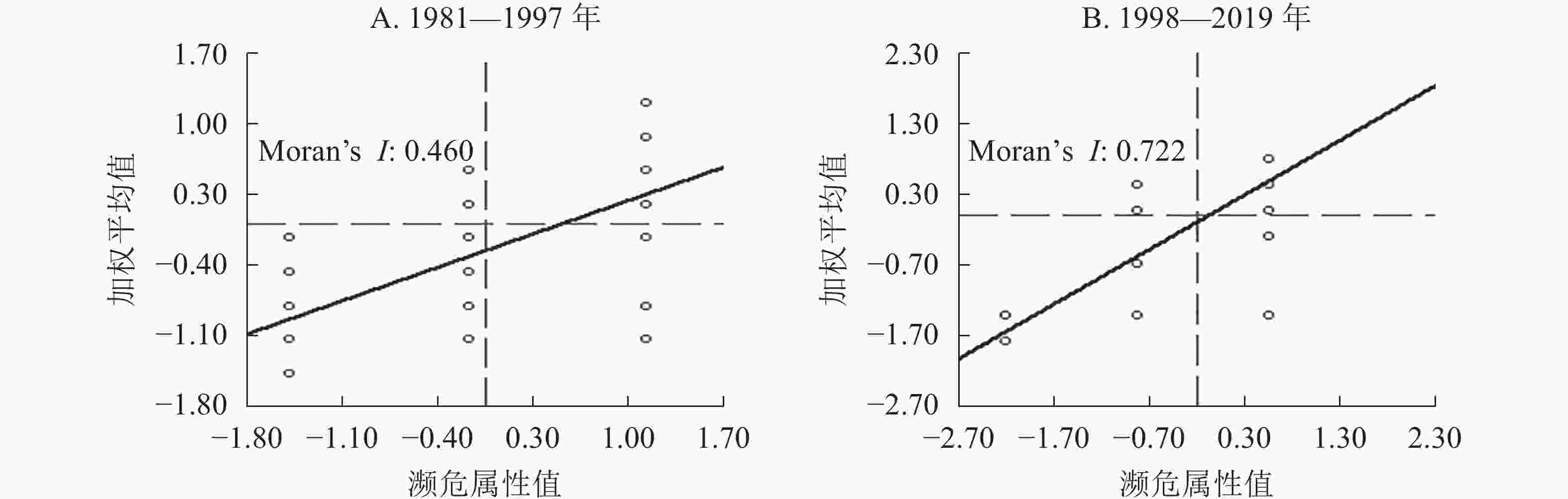
图 1 兰科植物的莫兰指数散点图
Figure 1. Moran scatter diagram of typical orchids
-
热点分析是根据统计学方法,识别具有统计显著性的热点和冷点。由表5可以看出:1981—1997年兰科植物处在高—高聚集模式下的省份只有贵州和湖北,说明这2个省份兰科植物的生存状况面临威胁。1998—2019年兰科植物在湖北的热点值更高,濒危程度进一步加重。由此可知:1998—2019年与1981—1997年相比,湖北空间分布的代表性兰科植物濒危程度更集中。代表性兰科植物濒危热点分布地区发生变化,即高危险区发生变化。其中需要重点保护的兰科植物是蕙兰、小舌唇兰、钩距虾脊兰、反瓣虾脊兰和金兰。湖北应该成为长江流域兰科植物监控、管理、保护的重点省份。虽然蕙兰在江浙地区分布整体处于冷点区域,但由于人为采挖,蕙兰的生存状况在江浙一带也须引起重视。
表 5 1981—2019兰科植物濒危等级热点值分布
Table 5. Distribution of endangered hot spot values of Orchidaceae plants from 1981 to 2019
省份 1981—1997年 1998—2019年 Z P Gi_ConfInvl
字段Z P Gi_ConfInvl
字段浙江 −0.87 0.38 0 0.10 0.92 0 云南 1.13 0.26 0 −0.32 0.75 0 西藏 −0.63 0.53 0 1.30 0.19 0 四川 −0.20 0.84 0 −0.23 0.82 0 陕西 0.27 0.79 0 0.11 0.91 0 青海 0.52 0.61 0 −0.75 0.46 0 江西 −0.48 0.63 0 0.11 0.91 0 湖南 1.55 0.12 0 0.46 0.64 0 湖北 1.75 0.08 1 1.96 0.05 2 河南 0.70 0.48 0 1.04 0.30 0 贵州 1.66 0.10 1 −0.74 0.46 0 广西 0.96 0.34 0 0.45 0.65 0 甘肃 −0.80 0.42 0 −0.69 0.49 0 福建 −1.60 0.11 0 −0.71 0.48 0 安徽 −0.87 0.38 0 0.10 0.92 0 上海 −1.11 0.27 0 −0.35 0.72 0 重庆 0.29 0.77 0 0.17 0.86 0 江苏 −1.11 0.27 0 −0.35 0.72 0 广东 0.13 0.90 0 0.45 0.65 0 说明:Z>1.65,P<0.10,Gi_ConfInvl字段为1,说明热点 具有置信度为90%的统计显著性;Z>1.96, P<0.05,Gi_ConfInvl 字段为2,说明热点具有置信 度为95%的统计显著性 -
温度和降水的变化可能是影响兰科植物生长的自然因素。从空间格局来看,1998—2019年与1981—1997年相比,长江流域代表性兰科植物多核破碎化趋势有所减弱,濒危值和空间位置表现出高度的空间自相关,兰科植物更趋向于集中分布。从长江流域关注重点地区来说,四川、贵州和广西是代表性兰科植物密度分布发生变化的省份,湖北是现阶段濒危等级值较高的省份,即热点省份。可能是因为适宜兰科植物生长的区域受到气候变化和人为因素的双重影响。在1998年以后,全球变暖进入停滞化阶段[29],20世纪90年代中后期长江流域气温产生明显变化[30],增长速度有所减缓[20-22]。气候变化给兰科植物的生长带来了较大影响。LIU等[31]研究认为:在气候变化的大背景下,兰科植物15%的种类和25%的属存在数量减少或局部灭绝的高风险,这与本研究提出的全球变暖会影响兰科植物生存发展的结论相符。
1998—2019年兰科植物的密度高值区减少,分布在广西、贵州的兰科植物消失。兰科植物的密度中值区由四川中部向四川北部迁移。濒危等级值和空间位置的正相关水平显著增强,聚集特征更加明显。高危险区数量由贵州和湖北2个省份减少为湖北1个省份。高危险区濒危水平增长,热点值更高,兰科植物的生存处境更加严峻,湖北将成为重点关注的地区。
修建基础设施虽然带给人们极大便利,但也影响了兰科植物的生境,导致兰科植物数量减少、分布范围改变。道路修建会破坏地表植被,增加车流量,排放有害气体,从而影响兰科植物的生长。水利设施的修建改变了小气候,也影响了兰科植物的生长。此外,国家对兰科植物的保护较弱,监管部门的责任不到位,导致采挖和线上线下销售现象严重。人们对森林资源的过度采伐,使附生兰和地生兰生存的场所受到影响,这也极不利于兰科植物的生长和繁殖。
Spatial distribution of endangered orchids in the Yangtze River Watershed
-
摘要:
目的 兰科Orchidaceae植物已经成为旗舰保护类群,科学规范识别长江流域兰科植物的迁移规律,探究濒危兰科植物的空间分布,确定重点保护物种和热点地区,对于更好地研究和保护珍稀濒危兰科植物具有重要意义。 方法 以长江流域1981—2019年的代表性兰科植物为研究对象,基于野外考察和全球生物多样性信息资讯机构(GBIF)数据库选取兜被兰属Neottianthe、独蒜兰属Pleione等130个(10种)物种分布点为代表,根据植物地理学和地理信息系统(GIS)空间分析理论,采取核密度、莫兰指数和热点分布等方法,对长江流域濒危兰科植物的空间分布格局进行了研究。 结果 ①1998—2019年长江流域代表性兰科植物的密度高值区减少,分布在广西、贵州的兰科植物消失。密度中值区发生变化,兰科植物由四川中部向四川北部迁移;②长江流域代表性兰科植物的濒危等级值和空间位置呈显著正相关水平,1998—2019年比1981—1997年相关性更强;③高危险区发生变化,1981—1997年兰科植物高危险区主要分布在贵州和湖北,而1998—2019年主要分布在湖北。 结论 全球变暖、基础设施建设、非法采挖销售和过度砍伐森林都有可能成为兰科植物分布变化和数量减少的原因。湖北将成为以后需要重点关注的地区。图1表5参31 Abstract:Objective This study, with an investigation of the scientific and standard identification of the migration of Orchidaceae, a protected and flagship group, in the Yangtze River Watershed, is aimed at an exploration of the spatial distribution of endangered orchids and the determination of key protected species and hot spots so as to serve the better research and protection of rare and endangered orchids. Method Taking the representative Orchidaceae plants in the Yangtze River Watershed from 1981 to 2019 as the research object, 130 species distribution points of 10 species including Neottianthe and Pleione were selected based on the field investigation and Global Biodiversity Information Facility (GBIF) database. Then, with the guidance of the theory of phytogeography and the conduct of a GIS spatial analysis, the spacial distribution of endangered orchids was investigated employing kernel density, Moran index and hot spot distribution. Result (1) The period between 1998 and 2019 witnessed the disappearance of Orchidaceae plants in Guangxi and Guizhou, a decrease in high-density areas of them in the Yangtze River Watershed, changes in the median density areas as well as their migration from central Sichuan to north Sichuan. (2) There was a significant positive correlation between the level of endangerment of Orchidaceae plants and their spatial location and the correlation between 1998 and 2019 was stronger than that of 1981−1997. (3) There was a shift of high risk areas: from 1981 to 1997, the high risk areas of Orchidaceae were mainly distributed in Guizhou and Hubei whereas from 1998 to 2019, they were mainly distributed in Hubei Province. Conclusion There have been changes in the distribution of Orchidaceae plants and a reduction in quantity which mainly attribute to global warming, infrastructure construction, illegal harvesting and excessive deforestation and in the future Hubei would become the focus of attention. [Ch, 1 fig. 5 tab. 31 ref.] -
表 1 代表性兰科植物的分布
Table 1. Distribution of representative orchids
序号 属名 物种名 生境 生活型 濒危等级 1 舌唇兰属 Platanthera 小舌唇兰 Platanthera minor 山坡林下或草地 地生 近危 2 头蕊兰属 Cephalanthera 金兰 Cephalanthera falcata 山坡林下 地生 近危 3 兰属 Cymbidium 建兰 Cymbidium ensifolium 山坡林下 地生 易危 4 兰属 蕙兰 Cymbidium faberi 向南山坡或黄山松 Pinus taiwanensis 林下 地生 易危 5 虾脊兰属 Calanthe 反瓣虾脊兰 Calanthe reflexa 常绿阔叶林下、山谷溪边 地生 无危 6 虾脊兰属 钩距虾脊兰 Calanthe graciliflora 山坡林下或石壁上 地生 易危 7 兜被兰属 Neottianthe 二叶兜被兰 Neottianthe cucullata 针叶林下或高山草甸上 地生 近危 8 无柱兰属 Amitostigma 无柱兰 Amitostigma gracile 岩石上或沟边阴湿草地上 地生 近危 9 独蒜兰属 Pleione 独蒜兰 Pleione bulbocodioides 苔藓覆被的岩石上 附生 易危 10 风兰属 Neofinetia 风兰 Neofinetia falcata 山中林地树干上 附生 濒危  下载: 导出CSV
下载: 导出CSV
表 2 1981—2019年四川代表性兰科植物的空间分布
Table 2. Spatial distribution of representative Orchidaceae plants in Sichuan from 1981 to 2019
年份 地区 经纬度 代表性兰科植物 年份 地区 经纬度 代表性兰科植物 1994 平武县 32°26′24″N,104°33′00″E 钩距虾脊兰 1981 泸定县 29°54′50″N,102°14′02″E 反瓣虾脊兰 1983 雷波县 28°15′43″N,103°34′15″E 金兰 1984 泸定县 29°54′50″N,102°14′02″E 反瓣虾脊兰 1984 泸定县 29°54′50″N,102°14′02″E 金兰 1997 泸定县 29°30′00″N,101°54′00″E 二叶兜被兰 1984 泸定县 29°54′50″N,102°14′02″E 金兰 1997 康定县 30°06′00″N,101°48′00″E 二叶兜被兰 1981 泸定县 29°54′50″N,102°14′02″E 建兰 1997 康定县 30°08′45″N,101°51′36″E 二叶兜被兰 1981 泸定县 29°54′50″N,102°14′02″E 建兰 1997 泸定县 29°34′30″N,101°59′56″E 二叶兜被兰 1986 九寨沟县 33°15′36″N,104°13′48″E 蕙兰 2019 成都市青羊区 30°40′55″N,104°00′36″E 蕙兰 1982 泸定县 29°54′36″N,102°13′48″E 反瓣虾脊兰 2005 翁达自然保护区 31°54′00″N,100°54′00″E 二叶兜被兰 1983 九寨沟县 33°15′36″N,104°13′48″E 反瓣虾脊兰 2007 阿坝藏族羌族自治区金川县 31°42′00″N,102°00′00″E 二叶兜被兰 1981 泸定县 29°54′50″N,102°14′02″E 反瓣虾脊兰 2005 翁达自然保护区 31°58′55″N,100°57′50″E 二叶兜被兰 1984 北川县 31°53′34″N,104°26′09″E 反瓣虾脊兰 2005 翁达自然保护区 31°58′55″N,100°57′50″E 二叶兜被兰 1984 北川县 31°53′34″N,104°26′09″E 反瓣虾脊兰
下载: 导出CSV
表 3 1981—2019年贵州和广西代表性兰科植物的空间分布
Table 3. Spatial distribution of representative Orchidaceae plants in Guizhou and Guangxi from 1981 to 2019
年份 省份 地区 经纬度 代表性兰科植物 1986 贵州 松桃苗族自治县 28°09′00″N,109°12′00″E 钩距虾脊兰 1996 贵州 桐梓县 28°07′48″N,106°49′12″E 金兰 1988 贵州 印江土家族
苗族自治县27°59′24″N,108°24′00″E 小舌唇兰 1988 贵州 松桃苗族自治县 28°00′00″N,109°12′00″E 蕙兰 1988 贵州 石阡县 27°30′36″N,108°13′48″E 反瓣虾脊兰 1988 贵州 施秉县 27°01′48″N,108°11′24″E 反瓣虾脊兰 1981 广西 金秀县老山伐木场十六公里阴冲 24°10′30″N,110°14′06″E 钩距虾脊兰 1991 广西 兴安县猫儿山老山界 25°36′03″N,110°35′06″E 金兰 1984 广西 资源县猫儿山万亩林场 26°00′18″N,110°35′56″E 金兰 1992 广西 资源县塘洞猫儿山大竹坪 26°00′18″N,110°35′56″E 金兰 1982 广西 兴安县猫儿山梯子岭 25°36′03″N,110°35′06″E 金兰 1982 广西 金秀县 24°10′30″N,110°14′06″E 小舌唇兰 1994 广西 环江县川山乡大沙坡 25°06′25″N,108°18′54″E 独蒜兰 2016 广西 百色市那坡县 23°20′45″N,105°54′46″E 建兰 1998 广西 龙胜各族自治县花坪红毛界下界 25°44′31″N,110°02′24″E 反瓣虾脊兰 1998 广西 龙胜各族自治县花坪红毛界下界 25°44′31″N,110°02′24″E 反瓣虾脊兰 1998 广西 金秀老山银杉保护区 24°10′30″N,110°14′06″E 钩距虾脊兰 1998 广西 金秀老山银杉保护区 24°10′30″N,110°14′06″E 钩距虾脊兰 说明:银杉Cathaya argyrophylla
下载: 导出CSV
表 4 1981—2019年兰科植物濒危等级的全局莫兰指数
Table 4. Global Moran’ s I index of the endangered species of representative Orchidaceae plants from 1981 to 2019
时段 莫兰指数 Z 域值(α=0.05) 1981—1997年 0.241 3.727 1.96 1998—2019年 0.805 4.450 1.96
下载: 导出CSV
表 5 1981—2019兰科植物濒危等级热点值分布
Table 5. Distribution of endangered hot spot values of Orchidaceae plants from 1981 to 2019
省份 1981—1997年 1998—2019年 Z P Gi_ConfInvl
字段Z P Gi_ConfInvl
字段浙江 −0.87 0.38 0 0.10 0.92 0 云南 1.13 0.26 0 −0.32 0.75 0 西藏 −0.63 0.53 0 1.30 0.19 0 四川 −0.20 0.84 0 −0.23 0.82 0 陕西 0.27 0.79 0 0.11 0.91 0 青海 0.52 0.61 0 −0.75 0.46 0 江西 −0.48 0.63 0 0.11 0.91 0 湖南 1.55 0.12 0 0.46 0.64 0 湖北 1.75 0.08 1 1.96 0.05 2 河南 0.70 0.48 0 1.04 0.30 0 贵州 1.66 0.10 1 −0.74 0.46 0 广西 0.96 0.34 0 0.45 0.65 0 甘肃 −0.80 0.42 0 −0.69 0.49 0 福建 −1.60 0.11 0 −0.71 0.48 0 安徽 −0.87 0.38 0 0.10 0.92 0 上海 −1.11 0.27 0 −0.35 0.72 0 重庆 0.29 0.77 0 0.17 0.86 0 江苏 −1.11 0.27 0 −0.35 0.72 0 广东 0.13 0.90 0 0.45 0.65 0 说明:Z>1.65,P<0.10,Gi_ConfInvl字段为1,说明热点 具有置信度为90%的统计显著性;Z>1.96, P<0.05,Gi_ConfInvl 字段为2,说明热点具有置信 度为95%的统计显著性
下载: 导出CSV
-
[1] 袁喆, 喻志强, 冯兆洋, 等. 长江流域陆地生态系统NDVI时空变化特征及其对水热条件的响应[J]. 长江科学院院报, 2019, 36(11): 7 − 15. YUAN Zhe, YU Zhiqiang, FENG Zhaoyang, et al. Spatiotemporal variations of NDVI in terrestrial ecosystem in Yangtze River Basin and response to hydrothermal condition [J]. J Yangtze River Sci Res Inst, 2019, 36(11): 7 − 15. [2] 贾怡童, 林爱文, 朱弘纪. 长江流域不同气候分区生长季植被总初级生产力对极端气候变化的时空响应研究[J]. 国土与自然资源研究, 2020, 42(1): 38 − 42. JIA Yitong, LIN Aiwen, ZHU Hongji. Study on spatio-temporal response of total primary productivity of vegetation to extreme climate change in different climatic growing seasons in the Yangtze River Basin [J]. Territ Nat Resour Study, 2020, 42(1): 38 − 42. [3] 王重阳, 赵联军, 孟世勇. 王朗国家级自然保护区滑坡体兰科植物分布格局及其保护策略[J]. 生物多样性, 2022, 30(2): 21 − 30. WANG Chongyang, ZHAO Lianjun, MENG Shiyong. Spatial distribution pattern and protection strategy for orchids in landslide mass of the Wanglang National Nature Reserve [J]. Biodiversity Sci, 2022, 30(2): 21 − 30. [4] 胡会强, 余泽平, 王国兵, 等. 江西兰科药用植物资源调查[J]. 中国实验方剂学杂志, 2019, 25(21): 148 − 154. HU Huiqiang, YU Zeping, WANG Guobing, et al. Resources of Orchidaceae medicinal plants in Jiangxi Province [J]. Chin J Exp Tradit Medical Formulae, 2019, 25(21): 148 − 154. [5] 张殷波, 杜昊东, 金效华, 等. 中国野生兰科植物物种多样性与地理分布[J]. 科学通报, 2015, 60(2): 179 − 188. ZHANG Yinbo, DU Haodong, JIN Xiaohua, et al. Species diversity and geographic distribution of wild Orchidaceae in China [J]. Chin Sci Bull, 2015, 60(2): 179 − 188. [6] 罗毅波, 贾建生, 王春玲. 中国兰科植物保育的现状和展望[J]. 生物多样性, 2003, 11(1): 70 − 77. LUO Yibo, JIA Jiansheng, WANG Chunling. A general review of the conservation status of Chinese orchids [J]. Biodiversity Sci, 2003, 11(1): 70 − 77. [7] 王喜龙, 土艳丽, 文雪梅, 等. 藏东南兰科植物多样性及其沿海拔梯度的分布格局[J]. 中南林业科技大学学报, 2018, 38(12): 45 − 51. WANG Xilong, TU Yanli, WEN Xuemei, et al. Diversity and altitudinal distribution patterns of orchids in Southeastern of Tibet [J]. J Cent South Univ For Technol, 2018, 38(12): 45 − 51. [8] 刘洋, 杜凡, 李瑞年, 等. 滇西北兰科植物海拔分布格局[J]. 西南林业大学学报, 2012, 32(3): 40 − 46. LIU Yang, DU Fan, LI Ruinian, et al. Altitudinal distribution pattern of orchid plants in northwest of Yunnan Province [J]. J Southwest For Univ, 2012, 32(3): 40 − 46. [9] 弓莉, 罗建, 林玲. 南迦巴瓦兰科植物多样性及垂直分布格局[J]. 高原农业, 2020, 4(5): 499 − 505. GONG Li, LUO Jian, LIN Ling. Species diversity and vertical distribution pattern of Orchidaceae in Namcha Barwa, Tibet [J]. J Plateau Agric, 2020, 4(5): 499 − 505. [10] 杨正斌, 余东莉, 刘强. 西双版纳兰科植物海拔分布格局[J]. 林业调查规划, 2014, 39(3): 71 − 75, 101. YANG Zhengbin, YU Dongli, LIU Qiang. Altitudinal distribution of orchids in Xishuangbanna [J]. For Inventory Plann, 2014, 39(3): 71 − 75, 101. [11] 戍祖芳, 冯建孟. 滇西北地区兰科植物多样性的分布格局及其解释[J]. 楚雄师范学院学报, 2016, 31(6): 39 − 45. SHU Zufang, FENG Jianmeng. Geographical pattern of Orchidaceae diversity in northwest Yunnan and its explanation [J]. J Chuxiong Norm Univ, 2016, 31(6): 39 − 45. [12] 张晓龙. 中国野生兰科植物地理分布格局研究[D]. 太原: 山西大学, 2014. ZHANG Xiaolong. Study on Geographic Distribution Pattern of the Wild Orchidaceae Plants in China [D]. Taiyuan: Shanxi University, 2014. [13] JOBSON B, KERRI W, LARA J, et al. Home range and habitat selection of captive-bred and rehabilitated cape vultures Gyps coprotheres in southern Africa [J]. Oryx, 2020, 55(4): 607 − 612. [14] HANNAH B, NORMAN S L, PATRICIA A F. GIS Investigation of the relationship of sex and season on the population distribution of common bottle nose dolphins (Tursiops truncatus) in Charleston, South Carolina [J]. Int J Geogr Inf Sci, 2020, 34(8): 1552 − 1566. [15] 陈龙, 刘春兰, 马明睿, 等. 太行山生物多样性保护优先区(北京区域)急需保护物种的空间分布、热点识别及保护成效评价[J]. 生态与农村环境学报, 2019, 35(4): 451 − 458. CHEN Long, LIU Chunnan, MA Mingrui, et al. Spatial distribution, hot spots identification and protection effectiveness evaluation of urgently protected species in priority area of Taihang Mountains biodiversity conservation (Beijing region) [J]. J Ecol Rural Environ, 2019, 35(4): 451 − 458. [16] 王芳, 袁兴中, 熊森, 等. 重庆澎溪河湿地自然保护区生物多样性空间格局及热点区[J]. 应用生态学报, 2020, 31(31): 1682 − 1690. WANG Fang, YUAN Xingzhong, XIONG Sen, et al. Spatial patterns of biodiversity and hotspots in Chongqing Pengxi River Wetland Nature Reserve, China [J]. Chin J Appl Ecol, 2020, 31(31): 1682 − 1690. [17] 杨文涛, 吕春彤, 陈浩. 地理环境条件约束的入侵物种虚拟负样本生成方法[J]. 干旱区资源与环境, 2020, 34(9): 179 − 187. YANG Wentao, LÜ Chuntong, CHEN Hao. Generating pseudo-absence samples of invasive species under the constraint of geographical environment [J]. J Arid Land Resour Environ, 2020, 34(9): 179 − 187. [18] 徐海根, 雷军成. 外来入侵植物假高粱在我国的潜在分布区分析[J]. 植物保护, 2011, 37(3): 87 − 92. XU Haigen, LEI Juncheng. Prediction of the potential distribution of the alien invasive plant Sorghum halepense in China [J]. Plant Prot, 2011, 37(3): 87 − 92. [19] 汪松, 解焱. 中国物种红色名录(第1卷) [M]. 北京: 高等教育出版社, 2004: 300 − 468. WANG Song, XIE Yan. Red List of Chinese Species (Volume 1) [M]. Beijing: Higher Education Press, 2004: 300 − 468. [20] 张冲, 赵景波. 厄尔尼诺/拉尼娜事件对长江流域气候的影响研究[J]. 水土保持通报, 2011, 31(3): 1 − 6, 11 − 12. ZHANG Chong, ZHAO Jingbo. Effects of El Niño-Southern Oscillation events on climate in Yangtze River Basin [J]. Bull Soil Water Conserv, 2011, 31(3): 1 − 6, 11 − 12. [21] 张凤英, 张增信, 田佳西, 等. 长江流域森林NPP模拟及其对气候变化的响应[J]. 南京林业大学学报(自然科学版), 2021, 45(1): 175 − 181. ZHANG Fengying, ZHANG Zengxin, TIAN Jiaxi, et al. Forest NPP simulation in the Yangtze River Basin and its response to climate change [J]. J Nanjing For Univ Nat Sci Ed, 2021, 45(1): 175 − 181. [22] 张晓娅, 杨世伦. 流域气候变化和人类活动对长江径流量影响的辨识(1956—2011)[J]. 长江流域资源与环境, 2014, 23(12): 1729 − 1739. ZHANG Xiaoya, YANG Shilun. Climatic and anthropogenic impacts on water discharge in the Yangtze River over the last 56 years (1956−2011) [J]. Resour Environ Yangtze Basin, 2014, 23(12): 1729 − 1739. [23] 任平, 洪步庭, 周介铭. 基于空间自相关模型的农村居民点时空演变格局与特征研究[J]. 长江流域资源与环境, 2015, 24(12): 1993 − 2002. REN Ping, HONG Buting, ZHOU Jieming. Research of spatiotemporal pattern and characteristics for the evolution of rural settlements based on spatial autocorrelation model [J]. Resour Environ Yangtze Basin, 2015, 24(12): 1993 − 2002. [24] 蔡雪娇, 吴志峰, 程炯. 基于核密度估算的路网格局与景观破碎化分析[J]. 生态学杂志, 2012, 31(1): 158 − 164. CAI Xuejiao, WU Zhifeng, CHENG Jiong. Analysis of road network pattern and landscape fragmentation based on kernel density estimation [J]. Chin J Ecol, 2012, 31(1): 158 − 164. [25] 刘锐, 胡伟平, 王红亮, 等. 基于核密度估计的广佛都市区路网演变分析[J]. 地理科学, 2011, 31(1): 81 − 86. LIU Rui, HU Weiping, WANG Hongliang, et al. The road network evolution of Guangzhou-Foshan metropolitan area based on kernel density estimation [J]. J Geogr Sci, 2011, 31(1): 81 − 86. [26] 许章华, 刘健, 余坤勇, 等. 福建省马尾松毛虫害空间自相关分析[J]. 安全与环境学报, 2013, 13(6): 167 − 171. XU Zhanghua, LIU Jian, YU Kunyong, et al. Analysis of the spatial autocorrelation of Dendrolimus punctatus Walker in Fujian [J]. J Saf Environ, 2013, 13(6): 167 − 171. [27] 田怀珍, 陈林, 邢福武. 广东南岭国家级自然保护区兰科植物物种多样性及其保护[J]. 生物多样性, 2013, 21(2): 224 − 234. TIAN Huaizhen, CHEN Lin, XING Fuwu. Species diversity and conservation of orchids in Nanling National Nature Reserve, Guangdong [J]. Biodiversity Sci, 2013, 21(2): 224 − 234. [28] 黎磊, 陈家宽. 气候变化对野生植物的影响及保护对策[J]. 生物多样性, 2014, 22(5): 549 − 563. LI Lei, CHEN Jiakuan. Influence of climate change on wild plants and the conservation strategies [J]. Biodiversity Sci, 2014, 22(5): 549 − 563. [29] 于志磊, 秦天玲, 章数语, 等. 近年来长江流域植被指数变化规律及气候因素影响研究[J]. 中国水利水电科学研究院学报, 2016, 14(5): 362 − 366, 373. YU Zhilei, QIN Tianling, ZHANG Shuyu, et al. Analysis of vegetation dynamic variations and response to climatic factor in Yangtze River Basin in recent decades [J]. J China Inst Water Resour Hydropower Res, 2016, 14(5): 362 − 366, 373. [30] 丁斌, 顾显跃, 缪启龙. 长江流域近50年来的气温变化特征[J]. 长江流域资源与环境, 2006, 15(4): 531 − 536. DING Bin, GU Xianyue, MIU Qilong. Characteristics in the variation of temperature over the Yangtze River Valley over last 50 years [J]. Resour Environ Yangtze Basin, 2006, 15(4): 531 − 536. [31] LIU Hong, FENG Changlin, LUO Yibo, et al. Potential challenges of climate change to orchid conservation in a wild orchid hotspot in southwestern China [J]. Bot Rev, 2010, 76(2): 174 − 192. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210551
 点击查看大图
点击查看大图
计量
- 文章访问数: 1849
- HTML全文浏览量: 646
- PDF下载量: 50
- 被引次数: 0









