




-
桉树 Eucalyptus生长迅速,作为一种重要工业用材树种,在满足日益增长的全球木材供应需求方面发挥了重要作用[1]。在以往桉树育种策略中,培育速生桉树无性系能短时间内满足木材需求,前提需在无性系区域试验基础上,选出适宜本区域的优良无性系以推广应用[2]。材积是桉树重要的经济性状,对木材产量有显著影响。然而,推广的无性系在不同生境生长难免出现经济性状不良的问题,因此桉树育种策略需兼顾培育适应范围广的优良无性系。区域试验为选育适应性范围广的品种提供依据,且能反映植物生长状况及性状、品种稳定性的问题。
区域试验也存在着基因型与环境互作 (genotype × environment interaction, G×E) 影响品种在不同环境下的排序。G×E指基因型的相对表现在不同环境下缺乏稳定性[3],即基因型与环境互作效应越强,稳定性越低,是影响品种稳定性的主要原因。基因型与环境互作研究的重要性包括3点:其一,可认识不同性状在不同生长环境中的遗传规律,如赤桉 E. camaldulensis的保存率受土壤盐浓度的影响[4],桉树无性系的胸径受株行距的正向影响[5],翅茎西番莲 Passiflora alata的生长性状受海拔的影响[6]等;其二,可为特定生境选择经济性状表现最优的基因型,如在特定的恶劣环境中,选育出抗寒[7]、抗盐抗旱[8]、抗风[9]的桉树品种,使跨区域 (或气候) 种植成为可能;其三,可为多种生境选择稳定性好的优良基因型,如在不同土壤特性、气候类型的环境中选择较优的桉树品种[10−12]。通过基因型与环境互作的研究,可选择适应范围广、品种产量稳定的优良无性系。
几乎所有重要的林木品种的基因型和环境互作都有报道,包括杨属 Populus[13]、松属Pinus [14]、柳属 Salix[15]等。对桉树无性系与环境互作的研究,目前大多数使用的分析方法有稳定性分析、主效可加互作可乘 (additive main multiplicative interaction, AMMI) 分析、主成分分析以及近期流行的基因型和基因型与环境互作双标图[genotype main effects (G) and genotype × environment interaction (G×E), GGE]分析方法等[10]。其中GGE的优势是在AMMI分析基因型与环境互作效应的基础上,又更多考虑基因型的作用[16]。此外,对遗传和表型参数的估计,包括各性状方差分量、无性系重复力、遗传增益等,对育种策略的成功至关重要[17]。如仅从一个环境中估算遗传和表型参数,未考虑较大的G×E影响,会导致结果不准确,因此,从多个环境中获得的参数具有参考价值[18]。目前,仍缺乏对桉树基因型与环境互作效应的研究,已有的研究也仅在一个地点开展[19−21],因此评估栽培桉树不同基因与环境互作的效应具有重要意义。本研究通过研究不同地点中桉树无性系的生长性状,筛选各地点中的适生品种;对单地点、多地点建立线性混合模型以计算遗传参数;最后以材积平均值和最佳线性无偏预测(best linear unbiased prediction, BLUP)值进行GGE分析,选出优良无性系及其理想种植地点,为桉树优良品种的推广提供借鉴。
-
参试无性系共21个,4.5年生,详见表1,以种植面积较大的无性系DH32-29作为对照。
表 1 参试的无性系
Table 1. Clones tested
树种和杂交组合(♀×♂) 无性系 赤桉 SX95 粗皮桉×巨桉 EC199 粗皮桉×尾叶桉 EC195 巨桉×粗皮桉 EC183 巨桉×巨桉 EC184 巨桉×尾叶桉 EC182 尾叶桉×赤桉 EC189 尾叶桉×粗皮桉 EC186、EC187、EC190、
EC191、EC192、EC193、
EC194、EC196、EC200尾叶桉×巨桉 EC181、DH32-29(ck) 细叶桉×粗皮桉 EC198 细叶桉×巨桉 EC188 细叶桉×尾叶桉 EC180 说明:赤桉 E. camaldulensis,粗皮桉 E. pellita,巨桉 E. grandis,尾叶桉 E. urophylla。 -
采用完全随机区组设计,每个小区4行,每行5株,株行距为2 m×3 m,样地设置4个重复,1个重复内的各个无性系重复20次。于2016年6月种植,2020年12月调查树高、胸径、保存率。试验地分布在广西、广东及海南,详见表2。
表 2 各试验地基本情况
Table 2. Basic information of each test area
试验地 编码 纬度
(N)经度
(E)土壤
类型年均气
温/℃年均降
水量/mm广西南宁 E1 22°82′ 108°37′ 赤红壤 23.5 1 350 广西钦州 E2 21°98′ 108°66′ 砖红壤 24.0 1 600 广东雷州 E3 20°92′ 110°10′ 沙壤 23.2 1 864 广西扶绥 E4 22°64′ 107°91′ 赤红壤 24.0 1 250 海南定安 E5 19°68′ 110°36′ 砖红壤 26.0 1 639 说明:气候信息来源参考中国气象数据网 。 -
树高 (H, m) 和胸径 (D, cm) 分别使用瑞典Vertex Ⅳ超声波林木测高仪和卷尺测量,计算单株材积 (V, m3·株−1) [22],公式为:V=D2H/30 000。各无性系保存率为活立木株数占总株数的百分比。表型材积变异系数(Cv)公式为:Cv=σ/μ×100%,其中σ表示无性系材积标准差,μ表示无性系材积平均值。
-
单地点方差分析使用线性混合模型 (linear mixed model),模型[23]为:${{Y}}_{{ijk}}{=}{\mu}{+}{{B}}_{{i}}{+}{{C}}_{{j}}{+}{{B}}_{{C}{ij}}{+}{{ \varepsilon }}_{{ijk}}$。其中Yijk为单点试验第i重复第j无性系第k单株的观测值,μ为群体的总体平均值,Bi为第i重复效应,Cj为第j无性系效应,BCij为第j无性系和第i重复的互作效应,εijk为剩余项。其中以无性系、无性系与重复的互作效应为随机效应,获得单地点各无性系的BLUP值用于GGE分析。
多地点方差分析同样建立线性混合模型,计算各性状的方差分量和方差效应量,模型[23]为:${{Y}}_{{hij}}{=}{\mu}{+} {{L}}_{{h}}{+}{{B}{(}{L}{)}}_{{i}{(}{h}{)}}{+}{{C}}_{{j}}{+}{{I}}_{{{\rm{LC}}hj}}{+}{{ \varepsilon }}_{{hij}}$。其中,Yhij为单点试验第h地点内第i重复第j无性系的小区平均值,μ为群体的总体平均值,Lh为第h地点效应,B(L)i(h)为第h地点内第i重复效应,Cj为第j无性系效应,ILChj为第j无性系和第h地点的互作效应,εijk为剩余项。此外各性状的均方 (MS) 是由第Ⅰ类平方和与自由度 (DF) 的比值计算。偏效应量 (ES,η2) 基于第Ⅰ类平方和计算,公式为:${{\eta}}_{\left({A}\right)}^{{2}}{=}\dfrac{{{S}}_{{{\rm{S}}}{A}}}{{{S}}_{{{\rm{S}}}{A}}{+}{{S}}_{{{\rm{S}}}{{\rm{E}}}}}$。其中,A表示某个变异因子 (或因素)。SSA表示某个变异因子的平方和,SSE表示误差平方和。
基于线性混合模型,单地点以无性系、无性系与重复互作效应作为随机因子;多地点以无性系、无性系与地点互作效应作为随机因子,其他变异因子作为固定效应。获得的各性状方差分量用于计算遗传参数。无性系重复力(Rc)的计算公式[21]为:
$$ {{R}}_{{{\rm{c}}}}=\dfrac{{{V}}_{{{\rm{g}}}}}{{{V}}_{{{\rm{g}}}}+\dfrac{{{V}}_{{{\rm{ge}}}}}{{L}}+\dfrac{{{V}}_{{{\rm{e}}}}}{{N_L}}} 。 $$ 其中,Vg为无性系基因型方差,Vge为无性系与重复 (或地点)交互作用方差,Ve为残差,L为重复 (或地点) 个数,NL为重复 (或地点) 内各无性系个数,其中每个重复 (或地点) 内各无性系的个数不同,NL的值需使用调和平均数。
-
为了鉴别各试点中表现最好的无性系、分析各试点对无性系评价的相似性、筛选理想选材环境和无性系,分别基于材积平均值和BLUP值进行GGE模型分析,该模型公式[24−25]为:
$$ {{Y}}_{{ij}}-{{\overline{Y}}}_{{i}{j}}={{ \lambda }}_{{1}}{\xi}_{{i}{1}}{\eta}_{{j}{1}}+{{ \lambda }}_{{2}}{\xi}_{{i}{2}}{\xi}_{{j}{2}}+{{ \varepsilon }}_{{ij}} 。 $$ 其中,Yij为无性系i在环境j下的预测产量;$\overline Y_{ij} $为所有无性系i在环境j下的总体平均值;λ1和λ2为主成分单值分解,ξi1和ξi2是无性系i在主成分上的特征向量,ηj1和ηj2为环境j在主成分上的特征向量;εij是随机误差。
使用R 4.1.1、Rstudio v.2022.12.0+353、SPSS 26、Excel 2019进行数据分析。
-
各无性系的材积、材积变异系数及保存率的数据统计见表3,单地点方差分析表明各无性系的材积差异极显著 (P<0.01, 未列表)。5个试验地相比,南宁的平均材积 (0.101 0 m3·株−1) 较其他地点大,定安的平均材积 (0.064 6 m3·株−1) 较小,说明定安对于参试无性系的适应性相对差。在各地中选择前3个优良无性系 (除对照),其中南宁优良无性系为EC186、EC184和EC189;钦州为EC184、EC183和EC191;雷州为EC192、EC186和EC188;扶绥为EC188、EC184和EC183;定安为EC180、EC187和EC190。可见EC184在南宁、钦州和扶绥3地表现优良。各无性系 (多地点分析) 中,EC186、EC188、EC184的平均材积大于DH32-29。EC182、EC183、EC184和EC189的保存率均大于80.0%,但EC182与EC189的材积表现略低。此外,EC183、E184、EC189材积的变异系数 (38.51%~39.82%) 相对较低,表明这些无性系稳定性相对较好;EC186的材积变异系数 (45.61%) 则较高。
表 3 不同参试无性系生长性状
Table 3. Growth traits of different clones tested
无性系 材积/(m3·株−1) 平均材积/
(m3·株−1)变异系
数/%保存率/
%广西南宁 广西钦州 广东雷州 广西扶绥 海南定安 EC180 0.071 2 hi 0.046 7 fg 0.101 9 bcde 0.084 6 def 0.081 3 a 0.085 9 47.20 68.5 EC181 0.075 0 ghi 0.053 9 defg 0.077 9 g 0.062 2 hi 0.047 7 g 0.065 0 43.95 74.3 EC182 0.118 4 abcde 0.028 9 gh 0.058 5 h 0.050 5 ij 0.072 2 abc 0.064 2 45.55 83.1 EC183 0.116 3 bcdef 0.116 8 a 0.096 7 def 0.111 9 b 0.065 7 cde 0.098 9 38.52 83.8 EC184 0.137 8 ab 0.121 2 a 0.097 7 cdef 0.113 2 b 0.069 7 bc 0.102 1 38.85 80.7 EC186 0.151 9 a 0.084 9 bc 0.114 9 ab 0.096 7 cd 0.077 1 ab 0.104 8 45.61 66.7 EC187 0.112 9 bcdef 0.079 7 bcde 0.077 7 g 0.087 0 cdef 0.079 7 ab 0.085 1 33.42 53.7 EC188 0.104 0 bcdefgh 0.066 5 cdef 0.112 6 bc 0.126 2 a 0.064 4 cdef 0.103 2 45.22 57.7 EC189 0.134 8 abc 0.072 2 bcdef 0.084 3 fg 0.081 5 efg 0.068 5 bcd 0.082 3 38.49 84.0 EC190 0.094 6 defghi 0.051 7 efg 0.093 5 defg 0.060 3 hi 0.079 4 ab 0.075 6 46.78 78.5 EC191 0.128 5 abcd 0.099 1 ab 0.086 6 efg 0.083 7 ef 0.056 5 efg 0.085 4 44.51 76.6 EC192 0.113 2 bcdef 0.084 6 bc 0.127 6 a 0.090 9 cde 0.061 5 cdef 0.100 0 49.00 63.0 EC193 0.110 5 bcdefg 0.081 6 bcd 0.085 0 fg 0.083 1 ef 0.056 1 efg 0.080 5 50.97 56.7 EC194 0.079 7 fghi 0.046 7 fg 0.097 4 cdef 0.075 1 fg 0.065 4 cde 0.078 2 37.41 73.2 EC195 0.079 6 fghi 0.072 7 bcdef 0.099 3 cdef 0.048 5 jk 0.070 5 abc 0.073 9 46.21 78.0 EC196 0.100 0 cdefgh 0.077 5 bcde 0.096 4 def 0.097 1 c 0.071 4 abc 0.090 1 42.97 69.9 EC198 0.083 7 efghi 0.057 4 cdef 0.092 9 defg 0.069 3 gh 0.061 4 cdef 0.075 8 50.21 66.7 EC199 0.058 5 ij 0.074 8 bcdef 0.052 9 hi 0.077 8 fg 0.057 3 defg 0.063 3 51.25 73.3 EC200 0.090 2 efghi 0.079 0 bcde 0.108 9 bcd 0.081 6 efg 0.053 1 fg 0.084 2 40.71 56.4 SX95 0.037 0 j 0.014 2 h 0.041 5 i 0.038 0 k 0.032 7 h 0.036 6 51.61 76.2 DH32-29(ck) 0.134 7 − 0.091 7 − − 0.100 3 32.98 94.0 平均 0.101 0 0.070 5 0.070 5 0.081 0 0.064 6 说明:同列不同字母表示在0.05水平下差异显著。−表示数据未检测。 -
由表4可见:对于树高,地点×无性系的交互作用占总表型贡献率 (即方差效应量) 的14.3%,地点的效应量为22.0%,无性系的效应量为17.0%;对于胸径,地点×无性系的效应量为12.2%,地点的效应量为6.8%,无性系的效应量为21.2%;对于材积,地点×无性系的效应量为13.9%,地点的效应量为8.7%,无性系的效应量为20.7%。可见树高受到地点影响较其他性状大,胸径受地点影响较小。
表 4 地点与无性系的方差汇总
Table 4. Summary of variances of sites and clones
变异因子 自由度 树高 胸径 材积 SS MS ES/% SS MS ES/% SS MS ES/% 修正模型 110 10 992.9 99.9** 7 491.9 68.1** 2.132 0.019** 截距 1 927 809.3 927 809.3** 567 606.6 567 606.6** 25.965 25.965** 地点 4 4 411.0 1 102.8** 22.0 1 116.6 281.9** 6.8 0.378 0.095** 8.7 无性系 19 3 201.2 168.5** 17.0 4 104.9 215.5** 21.2 1.037 0.055** 20.7 地点×无性系 76 2 617.3 34.4** 14.3 2 108.6 27.8** 12.2 0.644 0.008** 13.9 重复 (地点) 11 763.5 69.4** 4.7 161.8 14.7** 1.1 0.072 0.007** 1.8 误差 3 759 15 632.8 4.2 15 244.1 4.1 3.975 0.001 总计 3 870 954 435.0 590 342.6 32.071 修正后总计 3 869 26 625.7 22 736.0 6.107 说明:SS、MS、ES分别表示第1类平方和、均方、方差效应量。**与*分别表示在0.01、0.05水平下F检验差异显著。 -
基于线性混合模型,获得各性状 (除保存率) 的方差结果和遗传参数值,结果如表5。树高、胸径、材积的无性系重复力分别为0.711 4~0.918 6、0.678 1~0.934 6、0.640 3~0.920 7,说明各性状无性系之间的生长变异主要受遗传控制。
表 5 不同地点无性系各性状的方差结果和遗传参数
Table 5. Variance results and genetic parameters of each character in clones from different locations
性状 方差结果/遗传参数 地点 广西南宁 广西钦州 广东雷州 广西扶绥 海南定安 5地点结合 树高 重复 − 13.409* 143.413** 101.418** 8.696** 无性系 34.142** 80.532** 57.043** 101.538** 16.978** 重复×无性系 − 9.511** 17.439** 8.793** 5.235** 误差 5.392 3.894 4.780 4.038 1.310 Ve 5.389 3 3.914 5 4.784 7 3.858 7 1.320 1 4.407 2 Vg 1.762 5 3.474 3 0.886 3 1.663 1 0.383 6 0.753 3 Vge − 0.561 1 0.835 2 0.331 8 0.432 1 0.936 0 Rc 0.830 5 0.881 1 0.751 1 0.918 6 0.711 4 0.781 6 胸径 重复 − 7.150 30.401** 15.180** 6.115** 无性系 37.191** 56.974** 117.873** 88.210** 13.528** 重复×无性系 − 6.480 8.302** 5.599** 4.505** 误差 7.348 4.787 5.248 3.127 1.150 Ve 7.345 1 4.863 6 5.450 3 3.173 9 1.1842 4.147 0 Vg 1.852 8 2.942 2 1.827 2 1.420 3 0.280 6 1.033 3 Vge − 0.107 1 0.131 5 0.164 7 0.362 2 0.672 5 Rc 0.790 7 0.888 3 0.934 6 0.937 8 0.678 1 0.868 5 材积 重复 − 0.001 0.014** 0.006** 0.002** 无性系 0.013** 0.014** 0.028** 0.028** 0.003** 重复×无性系 − 0.001 0.002** 0.002** 0.001** 误差 0.002 0.001 0.001 0.001 0.000 Ve 0.00177 8 0.00103 2 0.001 365 0.000 876 0.000 296 0.001 058 Vg 0.00073 8 0.00064 0 0.000 441 0.000 469 6.26E−05 0.000 247 Vge − 3.10E−05 5.51E−05 5.83E−05 9.80E−05 0.000 219 Rc 0.861 5 0.886 6 0.920 7 0.941 3 0.640 3 0.833 3 说明:广西南宁无性系、误差自由度分别为19、304,其他地的重复、无性系、重复×无性系、误差的自由度分别为广西钦州 (2、19、25、325);广东雷州 (3、19、54、1149);广西扶绥 (3、19、54、1174);海南定安 (3、19、43、632)。Ve表示残差,Vg表示无性系基因型方差,Vge表示重复 (或地点) 与基因型互作方差,Rc表示无性系重复力。**与*分别表示在0.01、0.05水平下F检验差异显著。−表示缺少重复数据。 广西南宁试验地材积无性系重复力较高,为0.861 5,胸径较低,为0.7907 ,该地残差较其他地大。广西钦州试验地各性状的无性系重复力几乎相等 (趋于0.89),基因型方差分量占较大比例,说明各无性系内单株之间生长变异受到遗传控制较高,不同重复、重复×无性系之间的胸径性状差异不显著,表明不同重复的种植环境对胸径生长无影响。广东雷州试验地树高性状无性系重复力 (0.751 1) 较低,可能其对环境更加敏感;而该地区无性系内胸径、材积性状重复力都较高。广西扶绥试验地各性状无性系重复力大于0.90,可能受环境因素的影响较小,而且方差结果表明该试验地各性状的无性系效应量较其他变异来源高。海南定安试验地各性状重复×无性系方差所占的分量比基因型大,导致无性系重复力相对较低,同时该地区的G×E大小 (通过重复×无性系的方差分量占无性系与重复×无性系两者之和的百分比来衡量) 较其他地点高。
5个地点联合分析中,无性系与地点的互作方差比单一地点重复互作方差要高,说明地点很大程度影响无性系的生长且该混合模型更有合理性。此时性状之间无性系重复力由大到小为胸径、材积、树高。
-
对材积原始数据进行GGE双标图分析见图1,PC1与PC2共同解释总G+G×E效应的78.96%,图1A中多边形由离原点最远的无性系连接而成,通过原点到多边形各边的垂线将双标图分成若干扇区,每个扇形区域中顶角的品种是该扇区中表现最好的无性系,靠近原点的品种对环境不敏感。根据落点的扇区可将广西南宁、海南定安划为一组,其中EC186在该环境材积最高;广西钦州、扶绥划为一组,其中EC184在该环境材积最高;广东雷州划为一组,其中EC183在该环境材积最高。
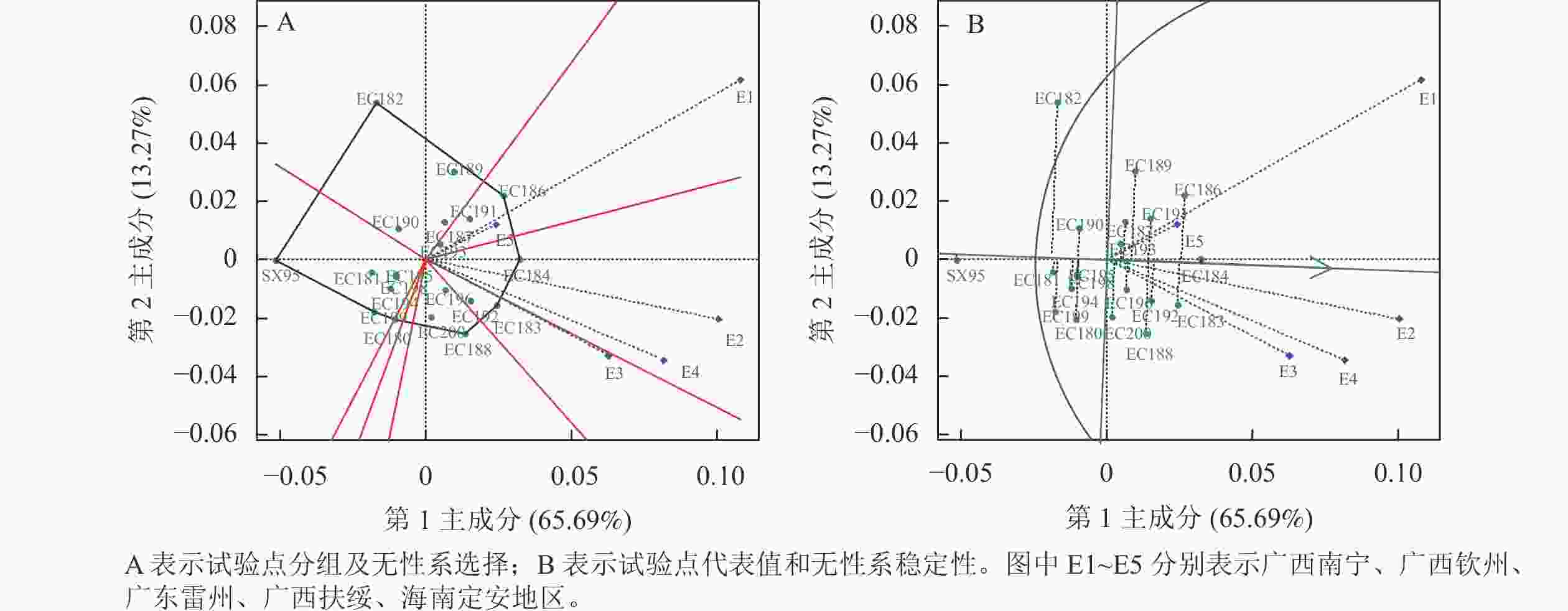
图 1 基于材积平均值的GGE分析图
Figure 1. GGE analysis diagram based on average wood volume
图1A中两地线段间夹角的余弦值是它们的相关系数,夹角小于90°表示正相关,说明两地对品种排序相似[26]。夹角较小说明试验点是重复设置的,去掉1个试验点不影响对品种的评价。各地点之间存在正相关,其中广西南宁和海南定安夹角最小,说明这2个环境对无性系的选择具有一致性。广西南宁和钦州线段较长,说明广西南宁和钦州有较强的无性系区分能力。然而,海南定安线段较短,说明海南定安的不同无性系之间的生长差异小。
图1B中带箭头的直线为平均环境轴,直线上圆圈表示平均环境值,试验地线段和平均环境轴的角度越小,代表性越强[27]。其中广西钦州与平均环境轴角度小,且线段较长,表明广西钦州具有较强的代表性和区分力,有区分力却没代表性的地区可以淘汰稳定性差的品种,既有区分力又有代表性的地区才能选择高产稳产的品种[28]。即在广西钦州选择的高产稳产品种较为可靠;广西南宁适于无性系的快速生长,同时也淘汰不稳定品种。
图1B中环境平均轴所指方向是无性系平均值的走向,无箭头的绿直线表示该性状的总平均值[27]。材积由大到小依次列出EC184、EC186、EC183、EC192、EC191、EC188、EC189、EC196、EC193、EC200 (只列出大于材积总平均值的无性系)。无性系与平均环境轴之间的垂线越长,表示无性系越不稳定,其中EC184具有较强的稳定性,即该无性系在各环境下生长表现差异不大;其次EC182的稳定性最低,说明该无性系在各环境下生长表现差异大,由表3数据:EC182在广西南宁生长较好,广西钦州生长较差。综合选择EC184为高材积量、高稳定性的无性系。
对材积的BLUP值进行GGE双标图分析可知 (因结果基本相似图未列出):主成分共同解释G+G×E效应的80.68%。选出无性系EC184在广西南宁、广西钦州、广西扶绥、海南定安环境下材积的表现最优,EC188在广东雷州环境下表现最好。除定安地区外,其他地区有较强的无性系区分能力。广西钦州 (E2) 与前文分析相同,为理想的材积试验环境。无性系材积由大到小依次为EC184、EC183、EC186、EC192、EC188、EC191、EC196、EC189、EC187、EC193、EC200 (只列出大于BLUP值的总平均值的无性系)。尽管EC184材积量最大,但BLUP-GGE模型得出EC183具有较强的材积稳定性,与前面分析有差异,因此综合选择EC183为高材积量、高稳定性的无性系。
-
无性系生长不仅受到遗传控制,而且受许多环境因子的影响,如降水量、土壤类型、气温、风力等。本研究表明:南宁地区的无性系生长表现最好。ROCHA等[29]研究表明:潮湿环境更有利于桉树无性系材积增长,然而广西南宁的年均降水量相比广西钦州、广东雷州、海南定安较低,意味着这3个点地的无性系本应该有较好的材积生长量,但是无性系材积都低于广西南宁,推断有其他因素影响这3个地区的无性系生长。本研究无性系品种以尾叶桉和巨桉为母本居多,不排除因品种因素导致广西南宁在较低年均降水量下有良好生长表现的可能[29]。另外,由于气候条件相差不大,还可能是土壤类型所致。广西南宁的土壤类型为赤红壤,广西钦州和海南定安的土壤为浅海沉积物发育的砖红壤,一般认为赤红壤比浅海沉积物砖红壤更有高导水性、持水和供水的能力[30]。其次,海南定安材积量较其他地小,可能与其年均气温较高、地理位置靠近赤道有关,这与de ARAUJO等[24]认为桉树无性系接近赤道的生产力在下降的结果有一定关联。此外,广西钦州、广东雷州、海南定安沿海,常年受到台风的影响,也可能导致无性系生长受抑制。由此可见,无性系的生长是遗传与环境相互作用的结果。
-
无性系生长过程中,各性状在不同环境下表现程度有所不同,有的性状受环境影响较大,不同性状的重复力也有所差异。本研究表明:树高易受到环境影响,重复力较低。方差效应量是衡量各变异因子大小的指标,树高性状中地点的方差效应量 (22.0%) 较无性系、无性系与地点的交互作用大,说明树高主要受地点影响,而胸径、材积主要受遗传的影响,与FREITAS等[31]、dos SANTOS等[32]的研究结果一致。本研究无性系各性状的重复力 (0.640 3~0.934 6) 处于高水平,说明桉树无性系生长性状受较高程度的遗传控制。多点结合的各性状重复力中,树高重复力较低,与朱显亮等[33]得出尾细桉无性系树高重复力高于胸径的结果不一致,可能是其研究仅涉及单个试验地,同时本研究单地点试验中广西南宁与海南定安的树高重复力较胸径高,说明单地点重复之间影响树高的环境因子较小,导致树高重复力高于胸径。可见重复力大小与环境因子有一定关联,影响树高的环境因子有待进一步研究。
-
本研究表明:GGE模型得出EC184为高产稳产的无性系,BLUP-GGE模型则为EC183,2个模型一致得出广西钦州为理想选材环境。倘若没有应用基因型与环境互作相关分析模型,以传统手段从多个地点综合选出优于DH32-29的优良无性系,那么结果可能与预期不一致。本研究优于DH32-29的无性系包括EC184、EC186、EC188,但EC186的变异系数大,EC188则保存率低。周家维等[7]在贵州地区中筛选出6个优良尾细桉优于DH32-29,李光友等[34]在桂北地区筛选出优于DH32-29的无性系均为尾巨桉杂交所得,并认为尾巨桉杂交值得关注,但都局限于单一地点,单一地区选择的优良无性系并不能说明该无性系在其他环境具有代表性。
以往研究表明:桉树生长性状受显著的G×E影响[35],本研究亦如此,不同地点的无性系重复之间是由环境造成,每个重复甚至单株之间所生长的空间环境独立,然而本研究广西钦州胸径的重复×无性系相互作用不显著,其他地区却显著,表明同一地点内往往会表现微环境、块状或梯度型的空间差异,而广西钦州的胸径则没有表现出环境差异。为了提高无性系遗传效应估算的准确性,有学者利用各个树体的行列数据进行空间分析[36],前提是种植布局的行列需整齐。对于G×E大小,本研究通过方差分量的比值来权衡,得出海南定安各性状的G×E最大,各性状中树高G×E较大。同样地,OSORIO等[37]通过两两地点的B型遗传相关 (实际上是方差分量的比值) 表明巨桉的树高G×E较材积、木材密度大。
GGE分析中,基于BLUP数据的GGE模型比原始数据具有更高的变异解释能力,因此BLUP-GGE模型更为可靠,这与程玲等[38]的研究一致。为了解决试验地环境异质性问题并使分析结果更可靠,程玲等[38]基于平均值、BLUP值与GGE双标图联合分析火炬树Rhus typhina的基因型与环境互作效应,发现原始平均值与BLUP值对应的理想选材环境不同,但是本研究平均值和BLUP值对应的理想选材环境都是广西钦州,可能是前者多考虑了空间分析造成选材环境有所差异。其次,基于平均值、BLUP值的GGE模型分别筛选出EC184、EC183为高材积量、高稳定性品种,2个无性系都是以巨桉为母本,说明巨桉在生长方面具有较强的基因稳定性以及适应能力,李光友等[34]也认为巨桉为母本具有良好的表现。
-
本研究综合5个地点的数据,桉树各无性系的生长表现存在显著差异。5个地点联合方差分析及遗传参数中,胸径的地点效应量 (6.8%) 较其他性状低,胸径的无性系重复力较其他性状高,说明胸径性状较其他更稳定,受到遗传控制较高。
基于材积平均值的GGE分析,广西钦州为理想选材环境,EC184为材积较高且稳定的无性系;基于材积BLUP值的GGE分析,广西钦州同样为材积理想选材环境,EC183为材积较高且稳定的无性系。所以GGE模型筛选出EC183和EC184为高材积和高稳定性的无性系。
Genotype × environment interaction on growth traits of Eucalyptus clones and selection of elite clones
-
摘要:
目的 对20个桉树 Eucalyptus无性系,包括1个赤桉E. camaldulensis无性系和19个杂交桉无性系,分别在广西南宁、广西钦州、广东雷州、广西扶绥和海南定安的立地条件下进行测试,探讨基因型、环境及其互作对无性系选育的影响,旨在为桉树的无性系筛选适生环境并推广提供借鉴。 方法 以各地点所测试的4.5年生桉树无性系为研究对象,基于各无性系的树高、胸径、单株材积、保存率的调查数据,比较各地点各无性系的生长表现。对单地点、多地点建立线性混合模型,计算各性状的方差分量和无性系重复力,同时计算单地点的最佳线性无偏预测(BLUP)值和多地点各性状的方差效应量。最后通过材积平均值和BLUP值进行基因型和基因型与环境互作双标图(GGE)分析。 结果 单地点分析中,各地点的最优无性系生长排序不同,无性系各性状受到环境的影响也不同。多地点分析中,桉树无性系基因型、环境以及基因型与环境互作存在显著差异,各性状的无性系重复力为0.781 6~0.868 5,其中胸径的无性系重复力最高且表现最稳定,树高易受环境影响。基于单株材积性状,筛选出EC186、EC188、EC184为优良无性系。基于材积平均值和BLUP值进行GGE模型分析得出EC183和EC184为高材积、高稳定性的无性系。 结论 20个无性系中,EC183和EC184为高材积、高稳定性的无性系。5个地点相比,广西南宁地区适合这批无性系的快速生长,广西钦州为这批无性系理想的选材环境。图1表5参38 -
关键词:
- 桉树无性系 /
- BLUP-GGE联合分析 /
- 重复力 /
- 基因型与环境互作
Abstract:Objective 20 Eucalyptus clones, including 1 Eucalyptus camaldulensis and 19 Eucalyptus hybrid clones, are tested at Nanning, Qinzhou, Fusui in Guangxi, Leizhou in Guangdong and Dingan in Hainan, respectively. And, the influence of genotype and environment interaction on Eucalyptus clones were mainly discussed. The results can provide a basement for screening suitable environment to the tested clones and reference for the promotion of excellent clones of Eucalyptus. Method The 4.5-year-old Eucalyptus clones tested at different sites were selected as the research objects. Based on the survey data of tree height, diameter at breast height (DBH), individual volume and survival rate of each clone, the growth performance of each clone at different sites was compared. A linear mixing model was established for single site and multi-site to calculate the variance components and clonal repeatability of each trait. The best linear unbiased predictive (BLUP) value of single site and the variance effect size of each trait at multi-site were also calculated. Finally, the average volume and BLUP values were used for genotype main effects (G) and genotype × environment interaction (G×E) analysis model (GGE). Result In single site analysis, the order of optimal clones in each site were different, and the traits in different sites were also affected by different environments. In the multi-site analysis, there were significant differences in genotype, environment and G×E interaction among Eucalyptus clones. The clone repeatability of each trait ranged from 0.781 6 to 0.868 5, among which the DBH trait had the highest and most stable repeatability, the tree height trait was most easily affected by environment. Based on the individual volume trait, EC186, EC188 and EC184 were selected as superior clones. Based on the average volume and BLUP value, the GGE model analysis showed that EC183 and EC184 were clones with high volume and high stability. Conclusion Among the 20 clones, EC183 and EC184 are clone with high individual volume and high stability. Compared with the five regions, Nanning in Guangxi is suitable for the rapid growth of these clones, Qinzhou in Guangxi is the ideal environment for the material selection of these clones. [Ch, 1 fig. 5 tab. 38 ref.] -
Key words:
- Eucalyptus clone /
- BLUP-GGE analysis /
- repeatability /
- genotype × environment interaction
-
表 1 参试的无性系
Table 1. Clones tested
树种和杂交组合(♀×♂) 无性系 赤桉 SX95 粗皮桉×巨桉 EC199 粗皮桉×尾叶桉 EC195 巨桉×粗皮桉 EC183 巨桉×巨桉 EC184 巨桉×尾叶桉 EC182 尾叶桉×赤桉 EC189 尾叶桉×粗皮桉 EC186、EC187、EC190、
EC191、EC192、EC193、
EC194、EC196、EC200尾叶桉×巨桉 EC181、DH32-29(ck) 细叶桉×粗皮桉 EC198 细叶桉×巨桉 EC188 细叶桉×尾叶桉 EC180 说明:赤桉 E. camaldulensis,粗皮桉 E. pellita,巨桉 E. grandis,尾叶桉 E. urophylla。  下载: 导出CSV
下载: 导出CSV
表 2 各试验地基本情况
Table 2. Basic information of each test area
试验地 编码 纬度
(N)经度
(E)土壤
类型年均气
温/℃年均降
水量/mm广西南宁 E1 22°82′ 108°37′ 赤红壤 23.5 1 350 广西钦州 E2 21°98′ 108°66′ 砖红壤 24.0 1 600 广东雷州 E3 20°92′ 110°10′ 沙壤 23.2 1 864 广西扶绥 E4 22°64′ 107°91′ 赤红壤 24.0 1 250 海南定安 E5 19°68′ 110°36′ 砖红壤 26.0 1 639 说明:气候信息来源参考中国气象数据网 ( http://data.cma.cn )。
下载: 导出CSV
表 3 不同参试无性系生长性状
Table 3. Growth traits of different clones tested
无性系 材积/(m3·株−1) 平均材积/
(m3·株−1)变异系
数/%保存率/
%广西南宁 广西钦州 广东雷州 广西扶绥 海南定安 EC180 0.071 2 hi 0.046 7 fg 0.101 9 bcde 0.084 6 def 0.081 3 a 0.085 9 47.20 68.5 EC181 0.075 0 ghi 0.053 9 defg 0.077 9 g 0.062 2 hi 0.047 7 g 0.065 0 43.95 74.3 EC182 0.118 4 abcde 0.028 9 gh 0.058 5 h 0.050 5 ij 0.072 2 abc 0.064 2 45.55 83.1 EC183 0.116 3 bcdef 0.116 8 a 0.096 7 def 0.111 9 b 0.065 7 cde 0.098 9 38.52 83.8 EC184 0.137 8 ab 0.121 2 a 0.097 7 cdef 0.113 2 b 0.069 7 bc 0.102 1 38.85 80.7 EC186 0.151 9 a 0.084 9 bc 0.114 9 ab 0.096 7 cd 0.077 1 ab 0.104 8 45.61 66.7 EC187 0.112 9 bcdef 0.079 7 bcde 0.077 7 g 0.087 0 cdef 0.079 7 ab 0.085 1 33.42 53.7 EC188 0.104 0 bcdefgh 0.066 5 cdef 0.112 6 bc 0.126 2 a 0.064 4 cdef 0.103 2 45.22 57.7 EC189 0.134 8 abc 0.072 2 bcdef 0.084 3 fg 0.081 5 efg 0.068 5 bcd 0.082 3 38.49 84.0 EC190 0.094 6 defghi 0.051 7 efg 0.093 5 defg 0.060 3 hi 0.079 4 ab 0.075 6 46.78 78.5 EC191 0.128 5 abcd 0.099 1 ab 0.086 6 efg 0.083 7 ef 0.056 5 efg 0.085 4 44.51 76.6 EC192 0.113 2 bcdef 0.084 6 bc 0.127 6 a 0.090 9 cde 0.061 5 cdef 0.100 0 49.00 63.0 EC193 0.110 5 bcdefg 0.081 6 bcd 0.085 0 fg 0.083 1 ef 0.056 1 efg 0.080 5 50.97 56.7 EC194 0.079 7 fghi 0.046 7 fg 0.097 4 cdef 0.075 1 fg 0.065 4 cde 0.078 2 37.41 73.2 EC195 0.079 6 fghi 0.072 7 bcdef 0.099 3 cdef 0.048 5 jk 0.070 5 abc 0.073 9 46.21 78.0 EC196 0.100 0 cdefgh 0.077 5 bcde 0.096 4 def 0.097 1 c 0.071 4 abc 0.090 1 42.97 69.9 EC198 0.083 7 efghi 0.057 4 cdef 0.092 9 defg 0.069 3 gh 0.061 4 cdef 0.075 8 50.21 66.7 EC199 0.058 5 ij 0.074 8 bcdef 0.052 9 hi 0.077 8 fg 0.057 3 defg 0.063 3 51.25 73.3 EC200 0.090 2 efghi 0.079 0 bcde 0.108 9 bcd 0.081 6 efg 0.053 1 fg 0.084 2 40.71 56.4 SX95 0.037 0 j 0.014 2 h 0.041 5 i 0.038 0 k 0.032 7 h 0.036 6 51.61 76.2 DH32-29(ck) 0.134 7 − 0.091 7 − − 0.100 3 32.98 94.0 平均 0.101 0 0.070 5 0.070 5 0.081 0 0.064 6 说明:同列不同字母表示在0.05水平下差异显著。−表示数据未检测。
下载: 导出CSV
表 4 地点与无性系的方差汇总
Table 4. Summary of variances of sites and clones
变异因子 自由度 树高 胸径 材积 SS MS ES/% SS MS ES/% SS MS ES/% 修正模型 110 10 992.9 99.9** 7 491.9 68.1** 2.132 0.019** 截距 1 927 809.3 927 809.3** 567 606.6 567 606.6** 25.965 25.965** 地点 4 4 411.0 1 102.8** 22.0 1 116.6 281.9** 6.8 0.378 0.095** 8.7 无性系 19 3 201.2 168.5** 17.0 4 104.9 215.5** 21.2 1.037 0.055** 20.7 地点×无性系 76 2 617.3 34.4** 14.3 2 108.6 27.8** 12.2 0.644 0.008** 13.9 重复 (地点) 11 763.5 69.4** 4.7 161.8 14.7** 1.1 0.072 0.007** 1.8 误差 3 759 15 632.8 4.2 15 244.1 4.1 3.975 0.001 总计 3 870 954 435.0 590 342.6 32.071 修正后总计 3 869 26 625.7 22 736.0 6.107 说明:SS、MS、ES分别表示第1类平方和、均方、方差效应量。**与*分别表示在0.01、0.05水平下F检验差异显著。
下载: 导出CSV
表 5 不同地点无性系各性状的方差结果和遗传参数
Table 5. Variance results and genetic parameters of each character in clones from different locations
性状 方差结果/遗传参数 地点 广西南宁 广西钦州 广东雷州 广西扶绥 海南定安 5地点结合 树高 重复 − 13.409* 143.413** 101.418** 8.696** 无性系 34.142** 80.532** 57.043** 101.538** 16.978** 重复×无性系 − 9.511** 17.439** 8.793** 5.235** 误差 5.392 3.894 4.780 4.038 1.310 Ve 5.389 3 3.914 5 4.784 7 3.858 7 1.320 1 4.407 2 Vg 1.762 5 3.474 3 0.886 3 1.663 1 0.383 6 0.753 3 Vge − 0.561 1 0.835 2 0.331 8 0.432 1 0.936 0 Rc 0.830 5 0.881 1 0.751 1 0.918 6 0.711 4 0.781 6 胸径 重复 − 7.150 30.401** 15.180** 6.115** 无性系 37.191** 56.974** 117.873** 88.210** 13.528** 重复×无性系 − 6.480 8.302** 5.599** 4.505** 误差 7.348 4.787 5.248 3.127 1.150 Ve 7.345 1 4.863 6 5.450 3 3.173 9 1.1842 4.147 0 Vg 1.852 8 2.942 2 1.827 2 1.420 3 0.280 6 1.033 3 Vge − 0.107 1 0.131 5 0.164 7 0.362 2 0.672 5 Rc 0.790 7 0.888 3 0.934 6 0.937 8 0.678 1 0.868 5 材积 重复 − 0.001 0.014** 0.006** 0.002** 无性系 0.013** 0.014** 0.028** 0.028** 0.003** 重复×无性系 − 0.001 0.002** 0.002** 0.001** 误差 0.002 0.001 0.001 0.001 0.000 Ve 0.00177 8 0.00103 2 0.001 365 0.000 876 0.000 296 0.001 058 Vg 0.00073 8 0.00064 0 0.000 441 0.000 469 6.26E−05 0.000 247 Vge − 3.10E−05 5.51E−05 5.83E−05 9.80E−05 0.000 219 Rc 0.861 5 0.886 6 0.920 7 0.941 3 0.640 3 0.833 3 说明:广西南宁无性系、误差自由度分别为19、304,其他地的重复、无性系、重复×无性系、误差的自由度分别为广西钦州 (2、19、25、325);广东雷州 (3、19、54、1149);广西扶绥 (3、19、54、1174);海南定安 (3、19、43、632)。Ve表示残差,Vg表示无性系基因型方差,Vge表示重复 (或地点) 与基因型互作方差,Rc表示无性系重复力。**与*分别表示在0.01、0.05水平下F检验差异显著。−表示缺少重复数据。
下载: 导出CSV
-
[1] ALFRED K, ZAITON S, NORZANALIA S. A review on the potential socio-economic impact of Eucalyptus plantation on local community [J]. Malays Forester, 2020, 83(2): 322 − 339. [2] 徐建民, 白嘉雨, 陆钊华. 华南地区桉树可持续遗传改良与育种策略[J]. 林业科学研究, 2001, 14(6): 587 − 594. XU Jianmin, BAI Jiayu, LU Zhaohua. Some sustainable strategies of improvement and breeding for Eucalyptus tree species in southern China [J]. Forest Research, 2001, 14(6): 587 − 594. [3] 林元震. 林木基因型与环境互作的研究方法及其应用[J]. 林业科学, 2019, 55(5): 142 − 151. LIN Yuanzhen. Research methodologies for genotype by environment interactions in forest trees and their applications [J]. Scientia Silvae Sinicae, 2019, 55(5): 142 − 151. [4] CHA-UM S, KIRDMANEE C. Response of Eucalyptus camaldulensis Dehnh. to different salt affected soils [J]. Acta Horticulturae, 2012, 937: 1057 − 1064. [5] ANDRÉ J L, OLIVEIRA R S, SETTE C R, et al. Wood volume of Eucalyptus clones established under different spacings in the Brazilian Cerrado [J]. Forest Science, 2021, 67(4): 478 − 489. [6] CHAVARRÍA-PEREZ L M, GIORDANI W, DAS GRAÇAS DIAS K O, et al. Improving yield and fruit quality traits in sweet passion fruit: evidence for genotype by environment interaction and cross-compatibility in selected genotypes [J/OL]. PLoS One, 2020, 15(5): e0232818[2022-11-15]. doi: 10.1371/journal.pone.0232818. [7] 周家维, 董飞, 袁昌选, 等. 不同桉树无性系在黔东南的速生性和耐寒性分析[J]. 种子, 2018, 37(9): 84 − 88. ZHOU Jiawei, DONG Fei, YUAN Changxuan, et al. Analysis of fast growth and cold tolerance of different Eucalyptus clone in Southeast Guizhou [J]. Seed, 2018, 37(9): 84 − 88. [8] MURAKAMI A. Development of salt tolerant Eucalyptus globulus selection [J]. Japan TAPPI Journal, 2006, 60(1): 69 − 74. [9] 朱英娟, 刘丽婷, 张水花, 等. 25个桉树无性系生长量及抗风性研究[J]. 广东农业科学, 2016, 43(7): 37 − 44. ZHU Yingjuan, LIU Liting, ZHANG Shuihua, et al. Study on increment and wind-resistance of Eucalyptus clones [J]. Guangdong Agricultural Sciences, 2016, 43(7): 37 − 44. [10] FURLAN R A, TAMBARUSSI E V, MORAES C B. Genetic parameters of Eucalyptus spp. clones in northeastern Brazil [J]. Floresta, 2020, 50(2): 1267 − 1278. [11] SANTOS G A, NUNES A C P, RESENDE M D V, et al. Genetic control and genotype-by-environment interaction of wood weight in Eucalyptus clones in the state of Rio Grande do Sul, Brazil [J]. Revista Árvore, 2016, 40(5): 867 − 876. [12] 崔之益, 徐大平, 杨曾奖, 等. 桉树无性系在华南6种立地条件下的适生性评价[J]. 华南农业大学学报, 2017, 38(3): 79 − 86. CUI Zhiyi, XU Daping, YANG Zengjiang, et al. Adaptability evaluation of Eucalyptus clones at six stand conditions in southern China [J]. Journal South China Agricultural University, 2017, 38(3): 79 − 86. [13] ALVAREZ J A, CORTIZO S C, GYENGE J E. Yield stability and phenotypic plasticity of Populus spp. clones growing in environmental gradients: I-yield stability under field conditions [J/OL]. Forest Ecology and Management, 2020, 463: 117995[2022-11-15]. doi: 10.1016/j.foreco.2020.117995. [14] SHALIZI M N, ISIK F. Genetic parameter estimates and GxE interaction in a large cloned population of Pinus taeda L. [J]. Tree Genet Genomes, 2019, 15(3): 1 − 13. [15] BRAGA R C, PALUDETO J G Z, SOUZA B M, et al. Genetic parameters and genotype × environment interaction in Pinus taeda clonal tests [J/OL]. Forest Ecology and Management, 2020, 474: 118342[2022-11-15]. doi: 10.1016/j.foreco.2020.118342. [16] 王大为, 王延波, 常程, 等. 多主成分玉米区域试验数据的GGE双标图分析[J]. 山西农业大学学报 (自然科学版), 2021, 41(1): 30 − 39. WANG Dawei, WANG Yanbo, CHANG Cheng, et al. GGE biplot analysis of maize multi-environment trial data with multi principal component [J]. Journal Shanxi Agricultural University (Natural Science Edition), 2021, 41(1): 30 − 39. [17] BIANCHI M C, BRUZI A T, SOARES I O, et al. Heritability and the genotype × environment interaction in soybean [J/OL]. Agrosystems Geosciences & Environment, 2020, 3(1): e20020 [2022-11-15]. doi: 10.1002/AGG2.20020. [18] SOARES I O, REZENDE P M, BRUZI A T, et al. Adaptability of soybean cultivars in different crop years [J]. Genetics and Molecular Research, 2015, 14(3): 8995 − 9003. [19] 王家妍, 莫雅芳, 申礼凤, 等. 桂南地区23个桉树无性系遗传变异分析和选择[J]. 西南农业学报, 2019, 32(9): 2174 − 2179. WANG Jiayan, MO Yafang, SHEN Lifeng, et al. Genetic variation analysis and selection of 23 Eucalyptus clones in Southern Guangxi [J]. Southwest China Journal of Agricultural Sciences, 2019, 32(9): 2174 − 2179. [20] de MORAES C B, de FREITAS T C M, PIERONI G B, et al. Genetic parameters of Eucalyptus clones for early selection in frost occurrence region [J]. Scencei Forestalis, 2014, 42(102): 219 − 227. [21] KARTIKANINGTYAS D, NIRSATMANTO A, SUNARTI S, et al. Trends of genetic parameters and stand volume productivity of selected clones of Eucalyptus pellita observed in clonal trials in Wonogiri, Central Java [J/OL]. IOP Conference Series: Earth and Environmental Science, 2020, 522(1): 012005[2022-11-15]. doi: 10.1088/1755-1315/522/1/012005. [22] 李宝琦, 徐建民, 李光友, 等. 桉树大径材无性系中期选择[J]. 安徽农业科学, 2009, 37(34): 17170 − 17174, 17225. LI Baoqi, XU Jianmin, LI Guangyou, et al. Research on the medium-term selection of the clone of Eucalyptus timber with large-diameter [J]. Journal of Anhui Agricultural Sciences, 2009, 37(34): 17170 − 17174, 17225. [23] 陈兴彬, 肖复明, 余林, 等. 基于混合线性模型估算杉木生长性状遗传参数[J]. 森林与环境学报, 2018, 38(4): 419 − 424. CHEN Xingbin, XIAO Fuming, YU Lin, et al. Estimation of genetic parameters of Cunninghamia lanceolata growth traits based on mixed linear model [J]. Journal of Forest Environment, 2018, 38(4): 419 − 424. [24] de ARAUJO M J, de PAULA R C, CAMPOE O C, et al. Adaptability and stability of eucalypt clones at different ages across environmental gradients in Brazil [J/OL]. Forest Ecology and Management, 2019, 454: 117631[2022-11-15]. doi: 10.1016/j.foreco.2019.117631. [25] dos SANTOS A, CECCON G, TEODORO P E, et al. Adaptability and stability of erect cowpea genotypes via REML/BLUP and GGE biplot [J]. Bragantia, 2016, 75(3): 299 − 306. [26] 陈朝阳, 魏建伟, 陈淑萍, 等. 黄淮海夏玉米品种籽粒产量基因型与环境互作分析[J]. 分子植物育种, 2019, 17(8): 2749 − 2760. CHEN Chaoyang, WEI Jianwei, CHEN Shuping, et al. Genotype by environment interaction effect on grain yield of Huanghuaihai summer maize cultivars [J]. Molecular Plant Breeding, 2019, 17(8): 2749 − 2760. [27] 王兵伟, 覃嘉明, 郑加兴, 等. 一年两季鲜食糯玉米区域试验的GGE双标图分析[J]. 南方农业学报, 2017, 48(11): 1961 − 1968. WANG Bingwei, QIN Jiaming, ZHENG Jiaxing, et al. GGE biplot analysis on regional trial of fresh-eaten waxy maize with two seasons per year [J]. Journal of Southern Agriculture, 2017, 48(11): 1961 − 1968. [28] 严威凯. 双标图分析在农作物品种多点试验中的应用[J]. 作物学报, 2010, 36(11): 1805 − 1819. YAN Weikai. Optimal use of biplots in analysis of multi-location variety test data [J]. Acta Agronomica Sinica, 2010, 36(11): 1805 − 1819. [29] ROCHA S M G, VIDAURRE G B, PEZZOPANE J E M, et al. Influence of climatic variations on production, biomass and density of wood in Eucalyptus clones of different species [J/OL]. Forest Ecology and Management, 2020, 473: 118290[2022-11-15]. doi: 10.1016/j.foreco.2020.118290. [30] 钟继洪, 郭庆荣, 谭军, 等. 桉林-砖红壤水分性能特征研究[J]. 土壤与环境, 2002, 11(2): 136 − 139. ZHONG Jihong, GUO Qingrong, TAN Jun, et al. Latosol water properties characteristics under Eucalyplus in Leizhou Peninsula [J]. Soil and Environmental Sciences, 2002, 11(2): 136 − 139. [31] FREITAS T P, OLIVEIRA J T D S, PAES J B, et al. Environmental effect on growth and characteristics of Eucalyptus wood [J/OL]. Florestae Ambiente, 2019, 26(4): e20160302[2022-11-15]. doi: 10.1590/2179-8087.030216. [32] dos SANTOS O P, CARVALHO I R, SZARESKI V J, et al. Multivariate approach in Eucalyptus breeding and its effect on genotype × environment interactions [J/OL]. Genetics and Molecular Research, 2018, 17(3): gmr18025[2022-11-15]. doi: 10.4238/gmr18025. [33] 朱显亮, 兰俊, 王建忠, 等. 中大径材尾细桉杂种无性系选择研究[J]. 南京林业大学学报 (自然科学版), 2020, 44(2): 43 − 50. ZHU Xianliang, LAN Jun, WANG Jianzhong, et al. Clonal selection of middle/large diameter timber of Eucalyptus urophylla × E. tereticornis hybrid clones [J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2020, 44(2): 43 − 50. [34] 李光友, 徐建民, 李昌荣, 等. 杂交桉家系在桂北生长及优良性评价[J]. 中南林业科技大学学报, 2021, 41(2): 8 − 15. LI Guangyou, XU Jianmin, LI Changrong, et al. Study on the grouped and genetic analysis of Eucalyptus hybrids in northern Guangxi [J]. Journal of Central South University of Forestry &Technology, 2021, 41(2): 8 − 15. [35] WU Shijun, XU Jianmin, LI Guangyou, et al. Genotypic variation in wood properties and growth traits of Eucalyptus hybrid clones in southern China [J]. New Forest, 2011, 42(1): 35 − 50. [36] 郑聪慧, 张鸿景, 王玉忠, 等. 基于BLUP和GGE双标图的华北落叶松家系区域试验分析[J]. 林业科学, 2019, 55(8): 73 − 83. ZHENG Conghui, ZHANG Hongjing, WANG Yuzhong, et al. An analysis of a regional trial of Larix principis-rupprechtii families based on BLUP and GGE biplot [J]. Scientia Silvae Sinicae, 2019, 55(8): 73 − 83. [37] OSORIO L F, WHITE T L, HUBER D A. Age trends of heritabilities and genotype-by-environment interactions for growth traits and wood density from clonal trials of Eucalyptus grandis Hill ex Maiden [J]. Silvae Genetica, 2001, 50(3/4): 108 − 116. [38] 程玲, 张心菲, 张鑫鑫, 等. 基于BLUP和GGE双标图的林木多地点试验分析[J]. 西北农林科技大学学报 (自然科学版), 2018, 46(3): 87 − 93. CHENG Ling, ZHANG Xinfei, ZHANG Xinxin, et al. Forestry multi-environment trial analysis based on BLUP and GGE biplot [J]. Journal of Northwest A&F University (Natural Science Edition), 2018, 46(3): 87 − 93. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220734
 点击查看大图
点击查看大图
计量
- 文章访问数: 1933
- HTML全文浏览量: 419
- PDF下载量: 70
- 被引次数: 0
