

 下载:
下载:


/%(</td>
<td class="table_top_border">时间(B) /h</td>
<td class="table_top_border">料液比(C)</td>
<td class="table_top_border">总黄酮得率/%</td>
</tr>
</thead>
<tbody>
<tr align="center">
<td class="table_top_border2">1</td>
<td class="table_top_border2">30</td>
<td class="table_top_border2">0.5</td>
<td class="table_top_border2">1:80</td>
<td class="table_top_border2">0.499</td>
</tr>
<tr align="center">
<td>2</td>
<td>30</td>
<td>1.0</td>
<td>1:40</td>
<td>0.594</td>
</tr>
<tr align="center">
<td>3</td>
<td>30</td>
<td>1.5</td>
<td>1:60</td>
<td>0.411</td>
</tr>
<tr align="center">
<td>4</td>
<td>70</td>
<td>0.5</td>
<td>1:40</td>
<td>0.936</td>
</tr>
<tr align="center">
<td>5</td>
<td>70</td>
<td>1.0</td>
<td>1:60</td>
<td>0.733</td>
</tr>
<tr align="center">
<td>6</td>
<td>70</td>
<td>1.5</td>
<td>1:80</td>
<td>0.476</td>
</tr>
<tr align="center">
<td>7</td>
<td>95</td>
<td>0.5</td>
<td>1:60</td>
<td>0.877</td>
</tr>
<tr align="center">
<td>8</td>
<td>95</td>
<td>1.0</td>
<td>1:80</td>
<td>0.625</td>
</tr>
<tr align="center">
<td>9</td>
<td>95</td>
<td>1.5</td>
<td>1:40</td>
<td>0.501</td>
</tr>
<tr align="center">
<td><i>K</i>1</td>
<td>1.504 0</td>
<td>2.312 0</td>
<td>2.031 0</td>
<td>
</td></tr>
<tr align="center">
<td><i>K</i>2</td>
<td>2.145 0</td>
<td>1.952 0</td>
<td>2.021 0</td>
<td>
</td></tr>
<tr align="center">
<td><i>K</i>3</td>
<td>2.003 0</td>
<td>1.388 0</td>
<td>1.600 0</td>
<td>
</td></tr>
<tr align="center">
<td><i>R</i>j</td>
<td>0.214 0</td>
<td>0.308 0</td>
<td>0.144 0</td>
<td>
</td></tr>
</tbody>
<tfoot>
<tr>
<td class="table_top_border" colspan="5">说明: <i>K</i>为第"<i>j</i>"列因素3水平所对应的试验指标和; <i>R<sub>j</sub></i>为第"<i>j</i>"列因素的极差。</td>
</tr>
</tfoot>
</table></div></foreignObject></svg>)
-
藜麦Chenopodium quinoa又称南美藜、藜谷、奎奴亚藜等,是1年生的藜科Chenopodiaceae草本作物,在安第斯山脉种植已有5 000多年的历史,被印加人称为“谷物之母”和“安第斯山的真金”[1-2]。藜麦蛋白质含量高,具有近乎完美的氨基酸组成,富含不饱和脂肪酸、类黄酮、维生素E等多种有益化合物,是联合国粮农组织认定的唯一的完美营养食品,被誉为“未来的超级谷物”“营养黄金”“有机谷类之王”等[3-4]。长期食用藜麦,对心脏病、高血压、高血糖、高血脂等有很好防治作用。藜麦喜热带、亚热带干湿气候,生长温度为2.0~35.0 ℃,生长适温14.0~18.0 ℃,在营养生长阶段可耐轻度霜冻(-1.0~0 ℃),在种子结实之后可耐-6.0 ℃低温,对盐碱、干旱、霜冻、病虫害等的抗性能力都很强,植株在自然肥力低的情况下仍能生长良好[5]。由于藜麦的营养价值在提供粮食和营养安全等方面的突出作用,联合国大会宣布2013为“国际藜麦年”[6]。黄酮类化合物是一种生理活性活泼的物质,具有降低血管脆性及异常的通透性、降血压、降血脂及胆固醇、抗病毒、抗炎、抗癌防癌、抗氧化等药理作用,尤以对心脑血管疾病的治疗作用而备受重视[7-8]。研究者已用不同方法在银杏Ginkgo biloba[9],甘薯Ipomoea batatas[8],花生Arachis hypogaea[10],大蒜Allium sativum[11],荞麦Fagopyrum esculentum[12],小白菜Brassica rapa chinensis[13],金银花Lonicera japonica[14]以及豆科Leguminosae植物[15-16]中等进行了黄酮提取和含量测定。藜麦谷粒中比较丰富的类黄酮物质异黄酮和维生素E组合可明显促进糖、脂代谢和胰岛素分泌,对糖尿病的治疗作用明显[17-19]。然而,有关藜麦黄酮类化合物的研究国内外报道甚少[20]。本研究着重讨论藜麦叶片黄酮类物质的提取和测定方法,进而分析基因型间的差异,为藜麦叶片黄酮类物质的开发利用和高黄酮高氧化性资源的筛选提供依据。
-
藜麦盆栽40 d后,摘取新鲜叶片,于阴凉通风处晾干后,置于80 ℃鼓风烘箱中干燥12 h,取出后用粉碎机充分粉碎,过0.5 mm孔筛筛选,装入干燥器皿中备用。
-
主要试剂:芦丁对照品购自国药集团化学试剂有限公司;亚硝酸钠、硝酸铝、氢氧化钠、乙醇等试剂均为国产分析纯。
主要仪器:DHG 9123A电热恒温鼓风干燥箱(上海精宏实验设备有限公司)、TP?鄄214电子天平(丹佛仪器有限公司)、XMTD?鄄6000恒温水浴锅(上海申胜生物技术有限公司)、TDL?鄄40B台式离心机(上海安亭科学仪器厂)、752PC紫外可见分光光度计(上海光谱仪器有限公司)。
-
以藜麦品种PI814932为材料,精确称取其全粉1.00 g,置于圆底烧瓶中。采用水浴回流加热法进行提取试验。选择影响藜麦叶片黄酮得率的主要因素如乙醇体积分数、料液比、浸提时间等3个因素为考察因子,按照L9(33)正交表安排不同提取条件试验(表 1),以确定最佳提取条件。
表 1 L9(33)正交表和实验结果
Table 1. Orthogonal table L9(33) and experimental results
试验编号 乙醇体积分数(A)/%( 时间(B) /h 料液比(C) 总黄酮得率/% 1 30 0.5 1:80 0.499 2 30 1.0 1:40 0.594 3 30 1.5 1:60 0.411 4 70 0.5 1:40 0.936 5 70 1.0 1:60 0.733 6 70 1.5 1:80 0.476 7 95 0.5 1:60 0.877 8 95 1.0 1:80 0.625 9 95 1.5 1:40 0.501 K1 1.504 0 2.312 0 2.031 0 K2 2.145 0 1.952 0 2.021 0 K3 2.003 0 1.388 0 1.600 0 Rj 0.214 0 0.308 0 0.144 0 说明: K为第"j"列因素3水平所对应的试验指标和; Rj为第"j"列因素的极差。 -
鉴于目前以芦丁为标样的比色法是测定黄酮得率最常用的方法[21],本研究仍沿用该法来测定藜麦总黄酮得率。准确称取芦丁标准试剂5.000 mg,用体积分数为60%乙醇完全溶解后定容至50.0 mL,摇匀的质量浓度为0.1 g·L-1的芦丁标准溶液。分别吸取芦丁标准溶液0.0,1.0,2.0,3.0,4.0,5.0 mL于6只10.0 mL刻度试管中,用体积分数为60%乙醇补至5.0 mL,加入质量分数为5%的亚硝酸钠溶液0.3 mL,摇匀,放置6 min后加入质量分数为10%的硝酸铝溶液0.3 mL,放置6 min,再加入1.0 mol·L-1氢氧化钠溶液4.0 mL,混匀,再加体积分数为60%的乙醇0.4 mL,室温放置15 min后于波长501 nm处测定其吸光度,以体积分数为60%乙醇溶液为空白对照,建立测定标准曲线和线性回归方程。测定时,以不同的样品液代替芦丁标准溶液,其他步骤与制作芦丁标准方程相同。计算公式如下:总黄酮得率(%)=(C×V1×V2×10-3)/(W/V0)×100。其中:C为测定样液的质量浓度(g·L-1);V0为测定吸光度所用样液的体积(mL);V1为测定时稀释体积(mL);V2为样液定容后体积(mL);W为样品质量(g)。
-
精确称取藜麦品种‘TEMUCO Quinoa TRADITIONAL’共6份相同样品,按上述最佳提取和测定方法进行重复性试验,验证可靠性。选上述藜麦品种‘TEMUCO Quinoa TRADITIONAL’,设定不同加入量,6次重复,加入已知量芦丁样液,计算回收率,黄酮提取和测定方法同上。
-
选择‘TEMUCO Quinoa TRADITIONAL’‘QuinoaB.Rain Sow’‘Temuco’‘1591 Quinoa Cherry’‘Tomico Quinoa’‘CQ?鄄TEMVCC’‘PI814932’‘PI596293’‘Tumuco(7)hybrids’‘PsI596498’10个不同基因型藜麦品种,统一播种和管理,采用上述同样方法进行制样。然后采用确定的最优提取工艺,对所选藜麦品种进行提取,分析藜麦叶片黄酮得率的基因型差异。
-
芦丁在0~50.0 mg·L-1质量浓度范围内,以吸光值(y)为纵坐标,芦丁标准品质量浓度(x)为横坐标,绘制标准曲线,得到回归方程:y=13.771x-0.013 6(R2=0.998 0)。该结果表明:芦丁在该质量浓度范围内吸光度值与质量浓度之间存在良好的线性关系。
-
选用30%,70%和95% 3个有代表性的乙醇体积分数作为提取液。因乙醇沸点在80 ℃左右,采用水浴热回流法(80~90 ℃)提取对它影响很小,因此,温度因素可不考虑在内。其他提取条件为提取时间和料液比。利用3因子3水平正交试验研究各因素对工艺的影响,同时为明确影响提取得率的显著性因子,对正交表中的黄酮得率作方差分析。结果分别见表 1和表 2。
表 2 藜麦总黄酮得率的方差分析
Table 2. Variation analysis of extraction rate of flavonoids in quinoa
方差来源 平方和(S) 自由度(f) 均方(s/f) F值 临界值 乙醇体积分数(A) 0.075 5 2 0.037 8 6.51 F0.05(2,2)=19 时间(B) 0.144 6 2 0.072 3 12.47 F0.1(2,2)=9 料液比(C) 0.040 3 2 0.020 2 3.47 F0.2(2,2)=4 误差 0.011 6 2 0.005 8 总和 0.272 0 8 由表 2可知:FB>F0.1,因素B对黄酮提取量的影响显著,F0.2<FA<F0.1,因素A对黄酮提取量有一定的影响,但不显著。由表 1可知,RB>RA>RC,据此,可确定各因素对黄酮提取量影响主次顺序为B>A>C。
-
由表 1可知:提取时间为0.5 h时黄酮得率最高,其后逐渐降低。因此,提取时间以0.5 h为佳。延长浸提时间,可能由于黄酮降解,从而降低了提取量[8]。
-
由表 2可知:乙醇休积分数对指标具有一定的影响。根据表 1和因素A的数据,乙醇体积分数为30%~70%时,黄酮得率剧增;而乙醇体积分数为70%~95%时,黄酮得率明显下降,显现一个抛物线变化的趋势。因此,确定乙醇最佳体积分数为70%。
-
由表 2可知:料液比对指标影响不显著。当对不同料液比进行试验时,发现料液比在1:40时,黄酮?鄄得率略高于料液比为1:60,其后随着料液比的增加黄酮?鄄得率逐渐降低。因此,从节约材料与药品综合比较后,确定最适料液比为1:40。
综上所述,藜麦总黄酮得率的最优提取条件为:乙醇体积分数70%,浸泡时间0.5 h,料液比1:40。
-
根据已获得的芦丁标准曲线与线性回归方程,计算出每毫升提取液中的总黄酮含量,再根据公式计算总黄酮得率。为验证比色法测定黄酮得率的可靠性,进行了6次重复性试验和回收率试验。实验结果显示:在波长510 nm条件下,各重复实验组的吸光度值分别为0.638,0.637,0.635,0.637,0.634,0.639。结果证明了比色法测定的可靠性;从回收率结果(表 3)可以看出,本测定方法各处理的回收率均接近100%,重现性也较高。综合上述,本法适用于测定藜麦黄酮类化合物的得率。
表 3 藜麦黄酮分析的回收率实验结果
Table 3. Experimental results of recovery in extraction rate of flavonoids in quinoa
试验编号 样品中含量/mg 加入芦丁量/mg 测得量/mg 回收率/% 平均回收率/% 相对标准偏差/% 1 7.134 0.100 7.441 102.86 100.25 4.22 2 14.268 0.100 13.415 93.37 3 7.039 0.100 7.441 104.23 4 14.078 0.100 14.265 100.61 5 7.998 0.100 8.368 103.33 6 15.996 0.100 15.630 97.10 -
表 4列出了10个藜麦品种的黄酮得率。由表 4可知:藜麦黄酮得率在基因型间存在很大差异,变幅为0.215%~0.933%,平均为0.619%,变异系数达34.44%。其中,以品种‘PI814932’的总黄酮得率最高,达0.933%。其次为‘QuinoaB. Rain Sow’和‘Temuco’,分别列第2位和3位,但两者基因型黄酮得率差异不大。紧随其后的分别是品种‘1591 Quinoa Cherry’和‘TEMUCO Quinoa TRADITIONAL’。品种‘Tomico Quinoa’和‘CQ?鄄TEMVCC’的基因型黄酮得率相近,分别排列第8位和第9位。而叶片表面覆有紫色粉层的‘PI596498’品种的总黄酮得率最低,为0.215%,藜麦总黄酮得率可能与其叶片表面的紫色粉层有关。
表 4 藜麦黄酮得率的基因型差异
Table 4. Genotype variation of extraction yields of flavonoids in quinoa
品种 叶表情况 叶形状 总黄酮得率/% TEMUCO Quinoa TRADITIONAL 绿色 提琴形锯齿状 0.635 QuinoaB.Rain Sow 绿色 提琴形锯齿状 0.818 Temuco 绿色 提琴形锯齿状 0.805 1591 Quinoa Cherry 绿色 提琴形锯齿状 0.751 Tomico Quinoa 绿色 提琴形锯齿状 0.472 CQ-TEMVCC 绿色 提琴形锯齿状 0.477 PI814932 绿色 提琴形锯齿状 0.933 PI596293 绿色 提琴形锯齿状 0.512 Tumuco(7)hybrids 绿色 提琴形锯齿状 0.572 PI596498 绿色(表面覆有紫色粉层) 卵形略有锯齿 0.215 平均值 0.619 变异系数/% 34.44 -
黄酮类化合物因其独特的保健功能而越来越得到人们的重视[8]。藜麦被认为是最适宜人类的完美“全营养食品”,是最具潜力的农作物之一。藜麦种子及新芽中的花青素、总多酚有较高的抗氧化性,可以作为传统食物的替代品,具有很高的营养价值[22]。藜麦的叶子可当蔬菜吃,与菠菜Spinacia oleracea叶子相似,既可用来凉拌生食,又是新鲜蔬菜的色拉的理想原料。本研究首次对藜麦的不同基因型品种的叶片黄酮得率进行了测定比较。参照国内外提取黄酮的多种方法[23-24],紫外可见分光光度法操作■简单方便,标准对照品易得,结果可靠,是黄酮类化合物的定量分子最常用的方法之一[25]。本着溶剂无毒性、易回收、对黄酮溶解力强和工艺简单的原则,确定乙醇为最佳提取剂。本研究表明,藜麦黄酮类化合物的提取率与乙醇体积分数、提取时间和料液比有关。经采用3因素3水平正交试验,其最佳提取工艺是乙醇体积分数70%,提取时间0.5 h,料液比1:40,且一次提取即可使得率达85%以上,这为藜麦黄酮类化合物的开发利用提供了理论依据和工艺参数。
黄酮得率的比色法测定比较简单可靠,但易受脂类物质的干扰[8]。从重复性试验和回收率试验中可看出,比色法适于测定藜麦总黄酮得率,且具有良好的重现性和稳定性。说明藜麦叶片中含脂量不足以影响藜麦黄酮的提取和测定。研究表明:‘PI814932’品种的黄酮提取量最高,达0.933%,该含量高于赪桐Clerodendrum japonicum根[24], 鲜青蒿Artemisia carvifolia[26], 甜菜Beta vulgaris[27]等的黄酮含量。不同品种间藜麦叶片黄酮得率差异系数达34.44%,说明通过适当的育种改良途径和方法,可以筛选到高黄酮高抗氧化性藜麦品种。藜麦黄酮提取量还可能受叶片粉质层的成分、叶片中叶绿素、花青苷、类胡萝卜素含量[28]以及内酯含量大小的遗传和环境因子[29]等因素影响。本项目将对不同生长发育时期的藜麦叶片黄酮含量变化,以及叶片和种子黄酮含量的相关性作进一步研究,更好地促进藜麦作为一种保健的多功能多方面可利用的食物源的大力开发。
Flavonoid extraction and flavonoid content with genotypic variation from Chenopodium quinoa leaves
-
摘要: 为了优化藜麦Chenopodium quinoa叶片黄酮的乙醇提取工艺和分析基因型间的差异, 为藜麦黄酮的开发和高黄酮的品种筛选提供理论依据, 采用3因子3水平正交试验设计, 探讨了乙醇体积分数、料液比和浸提时间等因素对藜麦叶片黄酮提取率的影响; 并采用最佳提取条件, 对10个不同基因型品种藜麦的叶片黄酮得率进行了比较分析。结果表明:藜麦叶片黄酮最佳提取条件为体积分数70%乙醇, 1:40料液比, 80℃水浴下回流浸提0.5 h。在优化条件下, 1次提取工艺得率达85%以上。各因素对叶片黄酮提取率的影响程度依次为:浸提时间>乙醇体积分数>料液比。比较结果发现:藜麦叶片黄酮得率存在明显基因型差异, 其中品种PI814932的叶片黄酮类物质得率最高, 达0.933%。所测藜麦品种的叶片黄酮平均得率为0.619%, 变异系数为34.44%。研究表明:乙醇回流法适于提取藜麦总黄酮类化合物。Abstract: The suitable technology of extracting flavonoids from Chenopodium quinoa leaves using the ethanol reflux extraction method, as well as the analysis of flavonoid content with genotypic variation, were studied in the paper. The research aims to provide theoretical basis for flavonids exploitation from C. quinoa and high-flavonoid variety screening. The effect of each factor such as ethanol concentration, extracting time and ratio of sample with ethanol solution on flavonoid extraction from C. quinoa was investigated using the orthogonal design L9(33) test. Moreover, flavonoids from C. quinoa leaves of ten different genotypes were extracted by the optimum extraction method and were subsequently analyzed for averages and coefficient of variation (CV). Results showed that optimum extracting conditions consisted of an extracting reagent of 70% ethanol, an extracting temperature of water-based heating of 80℃, a ratio of the leaf power to reagent of 1:40, and an extraction time of 0.5 h. For the extraction rate of falvonoids from C. quinoa leaves extracting time> ethanol concentration> ratio of sample to the ethanol solution. In one extraction cycle with optimum extracting conditions, the flavonoid extraction rate reached more than 85%. Additionally, extraction yields of flavonoids from different genotypes different with cultivar 'PI814932' having the highest flavonoid extraction rate (0.933%). The average extraction rate of flavonids from leaves of the ten tested varieties was 0.619%, and the CV was up to 34.44%. Hence, the ethanol reflux extraction method was suitable for flavonoid extraction from C. quinoa leaves.
-
Key words:
- botany /
- Chenopodium quinoa /
- flavonoid /
- extraction conditions /
- genotype variation
-
密码子是识别和传递生物体遗传信息、联系蛋白质与DNA之间的重要桥梁,在生物体遗传和变异中起着至关重要的作用[1]。编码同一氨基酸的不同密码子被称为同义密码子。由于基因突变和自然选择的影响,某些同义密码子在蛋白质翻译过程中往往被高频使用,被称为密码子的使用偏好性[2−3]。物种的生物学功能与密码子偏好性密切相关,密码子偏好性不仅可以影响生物编码基因的蛋白质合成速率和翻译速率[4],还会影响蛋白质结构、折叠程度和mRNA的合成[5]。研究表明:同一物种或亲缘关系相近的物种,具有相似的密码子偏好使用模式[6],通过分析物种的密码子偏好性可以衡量物种之间的基因表达量,进而探究物种之间亲属关系[7]。通过密码子偏好性的研究,能够更好地阐明物种进化过程中基因的表达规律[8],为利用基因工程技术改良物种目标基因提供参考依据[9]。
梁山慈竹Dendrocalamus farinosus属竹亚科Bambusoideae牡竹属Dendrocalamus,又名大叶竹和瓦灰竹,是中国西南地区重要的经济竹种[10],生长速度快,适应性强,竹笋效益高,属于优良的笋竹两用竹种,与硬头黄竹Bambusa rigida都属于竹编和制浆造纸的优质原料[11]。针对梁山慈竹叶绿体基因组密码子使用偏好性的研究鲜见报道。为了更好地挖掘和利用梁山慈竹的潜在经济价值,本研究以梁山慈竹叶绿体基因组序列为研究对象,分析其密码子偏好性使用模式,探究并总结其相关表达基因的密码子偏好性,以期分析影响梁山慈竹叶绿体基因组密码子偏好性的主要因素,并筛选出最优密码子,为后续梁山慈竹叶绿体基因工程改造等研究提供理论基础。
1. 材料与方法
1.1 叶绿体基因组序列的获取
根据GenBank登录号MZ681865.156在美国国家生物技术信息中心(NCBI)数据库中搜索并下载梁山慈竹叶绿体基因组序列,共有85条编码序列(CDS)。序列重复或小于300 bp会对密码子偏好性指标的测定产生影响[12]。对基因序列进行筛选,剔除序列长度小于300 bp且重复的序列,获取起始密码子为ATG,终止密码子为TAG、TGA和TAA的序列,最终获得51条CDS序列作为后续分析的样本序列。
1.2 密码子组成分析
运用CodonW1.4.2 (
http://sourceforge.net/projects/codonw )和EMBOSS (http://imed.med.ucm.es/EMBOSS/ )计算有效密码子数(ENC)、适应指数(CAI)、密码子偏性指数(CBI)、最优密码子频率(FOP)以及密码子第3位核苷酸A、T、C、G的含量(分别记为A3、T3、C3、G3)。利用ENC判断密码子偏好性程度,ENC>35说明密码子偏好性比较弱;反之,说明偏好性强[13]。通过CUSP软件分析并获得密码子鸟嘌呤(G)和胞嘧啶(C)所占的比率(GC比率)及GC平均比率(GCall),使用SPSS 25.0软件对梁山慈竹密码子各位置的GC比率与ENC进行相关分析。1.3 相对同义密码子使用度分析
运用CodonW 1.4.2对同义密码子相对使用度(RSCU)进行分析,即该密码子的实际使用频率与其理论使用频率的比值[14]。当RSCU大于1时,同义密码子中偏好使用该密码子,被称为高频密码子;当RSCU等于1时,密码子无偏好性;当RSCU小于1时,密码子使用偏好性较弱[15]。
1.4 中性绘图分析
中性绘图分析是对影响密码子使用偏好性的关键因素进行分析,X轴为GC3,Y轴为GC1和GC2的平均值,绘制二维散点图对GC3和GC12 (各基因 GC1和GC2的平均值)的相关性进行分析(GC1、GC2、GC3分别代表第1、2、3位密码子的GC比例)。若回归系数接近1,代表GC3和GC12显著相关,碱基组成没有差异,说明突变是决定密码子偏好性的主要因素;若回归系数接近0,则代表自然选择是主要因素。
1.5 ENC-plot绘图分析
ENC-plot绘图分析表现密码子的使用偏好性受到突变和自然选择的影响程度。使用Python 3.7进行ENC-plot绘图分析,构建散点图,横纵坐标分别为GC3、ENC,并绘制ENC的标准曲线。基因位点靠近或在标准曲线上,表明突变是决定密码子偏好性的主要因素,若基因位点和标准曲线距离很大,则说明偏好性主要由自然选择决定。
1.6 PR2-plot偏倚分析
PR2-plot分析表明基因中密码子的第3位碱基的构成情况。计算密码子碱基中第3位上4种碱基A、T、C、G比例,G3/(G3+C3)为X轴,A3/(A3+T3)为Y轴,绘制PR2-plot散点图,中心点为碱基比例A=T、C=G时的值,代表处于此区域的密码子并无使用偏好性[16]。
1.7 最优密码子分析
将51条基因升序排列后的ENC前后两端10%的基因建立高、低表达基因库。通过CodonW软件计算2个表达库中密码子的RSCU和ΔRSCU,同时满足高频密码子(RSCU>1)和高表达密码子(ΔRSCU≥0.08)的为最优密码子[17]。
1.8 梁山慈竹和其他几种生物密码子偏好性比较分析
在Codon Usage Database (
http://www.kazusa.or.jp/codon/ )下载异源表达宿主和植物代表类群,包括巨龙竹D. farinosus、粉麻竹D. sinicus、小叶龙竹D. pulverulentus、硬头黄竹、大肠埃希菌Escherichia coli、烟草Nicotiana tabacum、拟南芥Arabidopsis thaliana和酿酒酵母Saccharomyces cerevisiae等物种基因组密码子的使用频率,与梁山慈竹基因组密码子使用频率比值进行比较分析,当梁山慈竹密码子使用频率比其他生物的比值≥2.0或≤0.5时,说明该物种与梁山慈竹的同义密码子的使用偏好性差异较大,当比值不在上述范围内时,表明这2个物种对该密码子的偏好性较接近。1.9 对应分析
将叶绿体基因如表1所示进行功能分类,使用CodinW软件,选择对应分析计算样本中各个基因的RSCU,将分析结果分布在59维向量空间中,分析指标间的对应性。
表 1 梁山慈竹叶绿体基因结构分析Table 1 Structural analysis of the choroplast genome of D. farinosus基因分类 基因分组 基因名称 光合系统基因 光系统Ⅰ基因 psaA、psaB、psbA、psbC、psbD、psbB 光系统Ⅱ基因 petA、petB、petD 细胞色素b/f复合体基因 atpA、atpB、atpE、atpF、atpI 三磷酸腺苷合成酶基因 ndhA、ndhB、ndhC、ndhD、ndhE、ndhF、ndhG、ndhH、ndhI、ndhJ、ndhK 遗传系统基因 烟酰胺腺票吟二核甘酸氧化还原酶基因 rbcL 二磷酸核酮糖羧化酶大亚基基因 rpoA、rpoB、rpoC1、rpoC2 RNA聚合酶亚基基因 rps2、rps3、rps4、rps7、rps8、rps11、rps12、rps14、rps18 核糖体蛋白小亚基基因 rpl2、rpl14、rpl16、rpl20、rpl22 其他基因 成熟酶K基因 matK 膜蛋白基因 cemA 细胞色素合成基因 ccsA 酪蛋白分解蛋白酶基因 clpP 未知功能基因 假定叶绿体阅读框 ycf2、ycf3、infA 2. 结果与分析
2.1 密码子的碱基组成分析
分析梁山慈竹叶绿体基因组CDS序列的碱基组成:梁山慈竹的4种碱基所对应的同义密码子的第3位碱基比例 (T3s、A3s、C3s、G3s)分别为45.28%、42.07%、18.13%、17.96%,T3s和A3s远高于G3s和C3s,表明梁山慈竹叶绿体基因组密码子的第3位碱基以A/U结尾为主。梁山慈竹的ENC为50.40,CAI为16.6%,第3位同义密码子的GC比率 (GC3S)为28.1%,表明其叶绿体基因组密码子偏好性较弱。
梁山慈竹叶绿体基因组密码子的GC平均比率为39.48%,且GC1 (47.69%)>GC2 (39.70%)>GC3 (31.05%)。ENC为 39.04~61.00,均值为49.51,GC比率在基因密码子上并没有均匀分布(表2)。ENC和密码子3个位置GC比率的相关分析(表3)结果发现:ENC与GC3比率显著相关,与GC1、GC2不显著相关,说明密码子使用偏好性形成过程中GC3的影响作用大于GC1、GC2。
表 2 梁山慈竹叶绿体基因组各基因密码子相关参数统计Table 2 Statistics of codon related parameters of various genes in the chloroplast genome of D. farinosus基因 GC比率/% ENC CAI FOP 基因 GC比率/% ENC CAI FOP GC GC1 GC2 GC3 GC GC1 GC2 GC3 rps12 41.87 52.00 47.20 26.40 44.85 0.140 0.341 rps18 33.53 34.50 39.77 26.32 39.04 0.147 0.333 psbA 42.56 49.72 42.94 35.03 41.33 0.313 0.532 rpl20 36.11 38.33 40.83 29.17 50.97 0.112 0.298 matK 34.44 40.82 32.42 30.08 49.49 0.166 0.329 clpP 43.01 52.53 38.25 38.25 52.37 0.175 0.337 psbD 44.44 53.39 43.50 36.44 48.99 0.242 0.456 psbB 44.01 54.42 45.97 31.63 50.73 0.190 0.380 psbC 44.66 53.59 44.73 35.63 48.91 0.183 0.386 petB 41.06 48.93 41.20 33.05 47.31 0.191 0.333 rpoB 39.19 49.81 38.01 29.74 49.69 0.153 0.353 petD 40.37 50.93 39.13 31.06 49.46 0.161 0.305 rpoC1 39.87 49.93 38.07 31.63 52.77 0.156 0.347 rpoA 37.06 46.18 35.59 29.41 49.94 0.151 0.311 rpoC2 38.95 49.01 36.64 31.18 52.29 0.154 0.333 rps11 43.52 50.69 56.25 23.61 44.33 0.174 0.396 rps2 38.40 40.51 40.93 33.76 52.55 0.168 0.338 infA 40.35 43.86 35.96 41.23 61.00 0.181 0.409 atpI 38.84 47.58 36.29 32.66 50.55 0.163 0.353 rps8 36.50 41.61 41.61 26.28 46.62 0.122 0.374 atpF 38.27 47.62 35.45 31.75 53.17 0.147 0.353 rpl14 38.71 54.84 37.10 24.19 51.90 0.181 0.392 atpA 42.06 56.01 39.96 30.12 49.96 0.182 0.385 rpl16 44.76 52.14 53.57 28.57 39.41 0.115 0.354 rps14 39.42 39.42 46.15 32.69 41.73 0.135 0.384 rps3 33.47 43.75 31.67 25.00 48.03 0.193 0.402 psaB 41.81 48.71 43.13 33.61 49.34 0.172 0.350 rpl22 37.56 41.33 36.67 34.67 47.48 0.188 0.415 psaA 43.68 51.80 43.28 35.95 52.07 0.198 0.373 rpl2 44.56 51.77 48.58 33.33 53.33 0.143 0.361 ycf3 39.69 47.40 38.15 33.53 55.45 0.156 0.343 ndhB 38.16 42.07 39.33 33.07 46.71 0.156 0.348 rps4 37.13 47.52 37.13 26.73 49.59 0.169 0.386 rps7 39.49 49.68 45.22 23.57 48.31 0.164 0.373 ndhJ 39.38 49.38 36.88 31.88 51.48 0.176 0.356 ndhF 34.19 37.84 38.92 25.81 46.19 0.144 0.321 ndhK 38.60 41.70 43.72 30.36 51.91 0.159 0.329 ccsA 33.64 33.74 41.10 26.07 45.60 0.152 0.307 ndhC 39.67 50.41 36.36 32.33 48.75 0.177 0.345 ndhD 36.19 40.72 36.93 30.94 48.98 0.133 0.314 atpE 42.51 52.17 39.13 36.23 59.51 0.167 0.405 ndhE 33.33 41.18 32.35 26.47 59.06 0.144 0.316 atpB 42.62 53.91 41.68 32.26 47.43 0.192 0.381 ndhG 34.46 44.07 32.77 26.55 45.77 0.125 0.250 rbcL 44.14 57.11 43.93 31.38 50.19 0.271 0.454 ndhI 34.99 37.57 38.67 28.73 52.09 0.171 0.345 ycf4 41.22 48.39 39.78 35.48 47.14 0.162 0.385 ndhA 33.98 42.42 36.36 23.14 44.35 0.140 0.321 cemA 33.62 41.99 27.71 31.17 55.91 0.176 0.342 ndhH 37.82 50.76 34.77 27.92 49.95 0.155 0.322 petA 40.29 53.58 35.2 32.09 51.12 0.155 0.331 表 3 梁山慈竹叶绿体基因组中各基因参数的相关性分析Table 3 Correlation analysis of various gene parameters in the chloroplast genome of D. farinosus参数 GC1 GC2 GC3 ENC CAI CBI FOP GC3s GC GC1 1 GC2 0.300* 1 GC3 0.265 −0.009 1 ENC 0.142 −0.425** 0.389** 1 CAI 0.409** 0.076 0.370** 0.012 1 CBI 0.438** 0.272 0.322* −0.092 0.774** 1 FOP 0.402** 0.312* 0.341* −0.064 0.797** 0.965** 1 GC3s 0.271 −0.029 0.946** 0.445** 0.330* 0.330* 0.370** 1 GC 0.814** 0.673** 0.525** 0.010 0.407** 0.512** 0.518** 0.499** 1 说明: *表示显著相关 (P<0.05);**表示极显著相关 (P<0.01)。 2.2 相对同义密码子使用度分析
梁山慈竹叶绿体基因组中共包含18110个密码子(表4),总计编码20个氨基酸,密码子数为12~705个,其中密码子UGA共有12个,密码子含量最多的是编码谷氨酸的GAA,共有705个。梁山慈竹叶绿体基因组蛋白编码序列RSCU分析表明:氨基酸含量较高的有亮氨酸(Leu)和精氨酸(Arg),均为6个密码子编码,编码精氨酸的是UUA、UUG、CUU、CUC、CUA和CUG;编码亮氨酸的有AGA、AGG、CGU、CGC、CGA和CGG;除此之外,蛋氨酸(Met)和色氨酸(Trp)均只有1个密码子编码,分别是AUG和UGG,其余氨基酸密码子编码个数分别为2~4个。
表 4 梁山慈竹叶绿体基因组蛋白编码序列RSCU分析Table 4 RSCU of protein coding region in the chloroplast of D. farinosus氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU Phe UUU* 644 1.29 Tyr UAU* 532 1.59 Ser UCU* 343 1.58 Cys UGU* 151 1.53 Phe UUC 351 0.71 Tyr UAC 137 0.41 Ser UCC* 260 1.19 Cys UGC 47 0.47 Leu UUA* 634 1.94 TER UAA* 28 1.56 Ser UCA* 222 1.02 Arg AGA* 322 1.75 Leu UUG* 362 1.11 TER UAG 14 0.78 Ser UCG 119 0.55 Arg AGG 119 0.64 Leu CUU* 420 1.29 TER UGA 12 0.67 Ser AGU* 273 1.25 Arg CGU* 261 1.41 Leu CUC 138 0.42 Trp UGG* 328 1.00 Ser AGC 89 0.41 Arg CGC 95 0.51 Leu CUA 295 0.90 Gln CAA* 477 1.53 Thr ACU* 403 1.68 Arg CGA* 234 1.27 Leu CUG 107 0.33 Gln CAG 148 0.47 Thr ACC 181 0.75 Arg CGG 76 0.41 Ile AUU* 740 1.48 Glu GAA* 705 1.46 Thr ACA* 259 1.08 Gly GGU* 421 1.24 Ile AUC 295 0.59 Glu GAG 263 0.54 Thr ACG 116 0.48 Gly GGC 145 0.43 Ile AUA 461 0.92 Lys AAA* 647 1.44 Ala GCU* 493 1.73 Gly GGA* 538 1.58 Met AUG* 416 1.00 Lys AAG 253 0.56 Ala GCC 172 0.60 Gly GGG 259 0.76 Val GUU* 382 1.47 Asp GAU* 522 1.54 Ala GCA* 343 1.20 Pro CCU* 286 1.48 Val GUC 126 0.49 Asp GAC 155 0.46 Ala GCG 135 0.47 Pro CCC* 196 1.01 Val GUA* 390 1.50 His CAU* 311 1.47 Asn AAU* 528 1.48 Pro CCA* 209 1.08 Val GUG 139 0.54 His CAC 112 0.53 Asn AAC 187 0.52 Pro CCG 84 0.43 说明:*表示RSCU大于1的高频密码子。 梁山慈竹叶绿体基因组RSCU大于1的密码子数目为34个(分别为UUU、UUA、UUG、CUU、AUU、AUG、GUU、GUA、UCU、UCC、UCA、AGU、ACU、ACA、GCU、GCA、AAU、UAU、UAA、UGG、CAA、GAA、AAA、GAU、CAU、UGU、AGA、CGU、CGA、GGU、GGA、CCU、CCC和CCA),即筛选出了34个高频密码子,其中以A、U、C、G结尾的密码子分别有13、16、2和1个,这说明密码子偏好以A和U结尾,RSCU较高的3个密码子分别为UUU (1.94)、CUA (1.73)和UCU (1.75)。
2.3 中性绘图分析
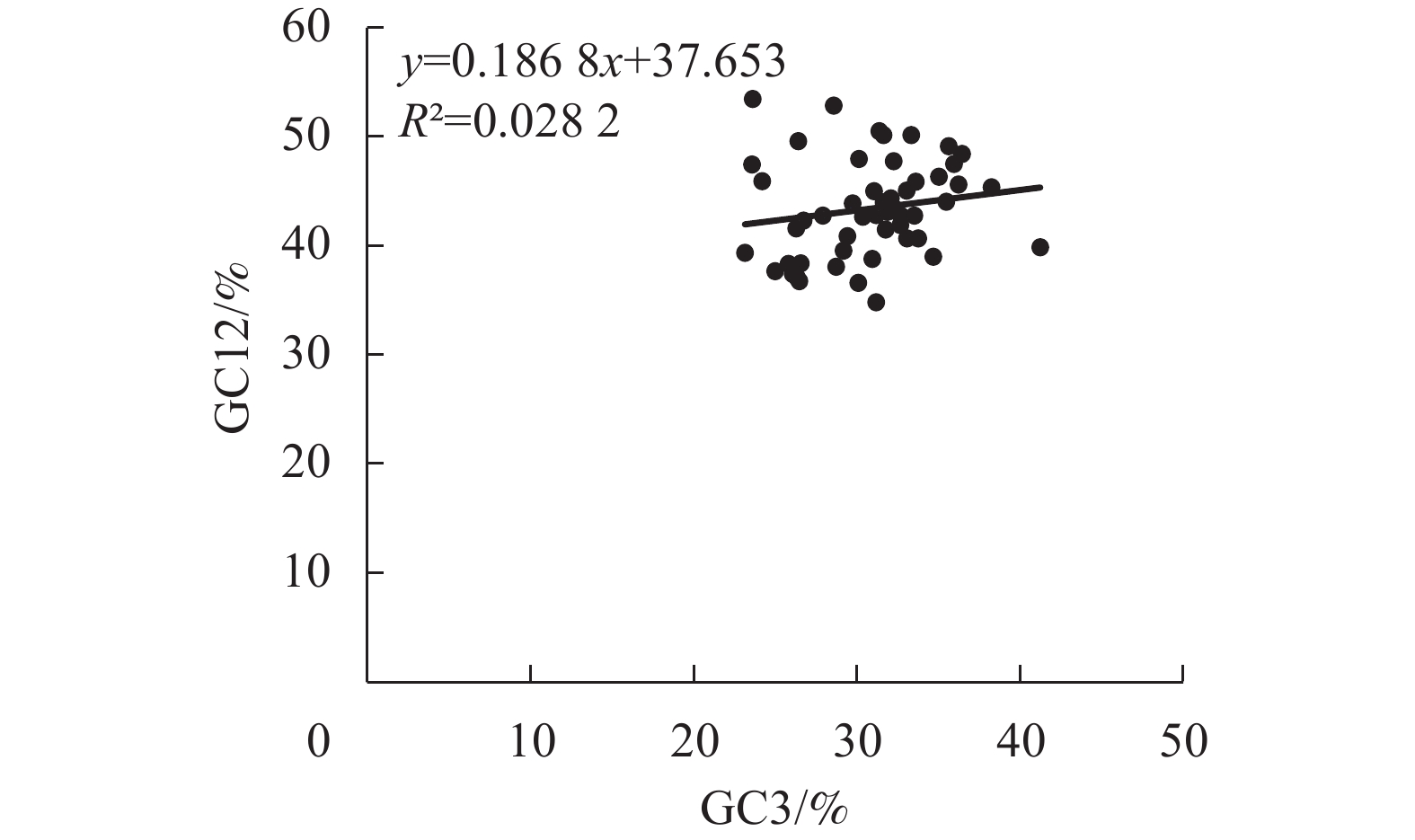
中性绘图分析量化自然选择和突变压力之间的关系,阐明3个密码子位置之间的联系。结果表明:横坐标GC3的数值为23.14%~41.23%,纵坐标GC12的数值为39.04%~61.00% (图1)。梁山慈竹的Pearson相关系数为0.17,呈正相关关系,数据拟合后的回归系数为0.1868,决定系数(R2)较小,为0.0282,GC12和GC3的相关性不显著,说明其叶绿体基因组密码子偏好性受自然选择影响较大。
2.4 ENC-plot绘图分析
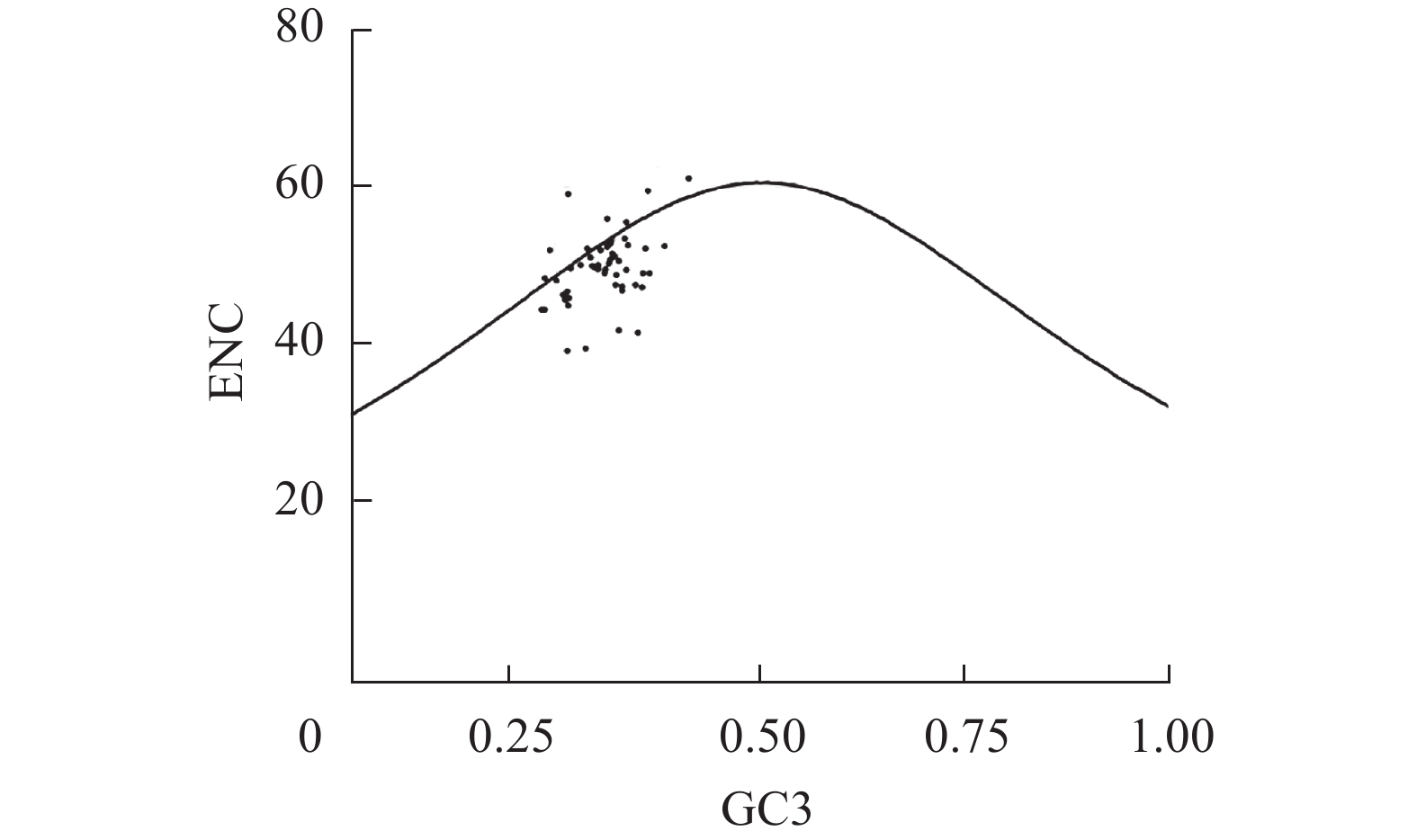
图2显示:ENC分布并不紧密,少量分布在标准曲线附近,还有个别分布在标准曲线上侧,位点的ENC均大于35,与预期ENC值有差距。说明梁山慈竹密码子偏好性较弱且自然选择和突变都对其偏好性有影响。由于落在标准曲线下方的基因点数量比较多,所以梁山慈竹基因组密码子使用偏好性主要受自然选择的影响。
2.5 PR2-plot绘图分析
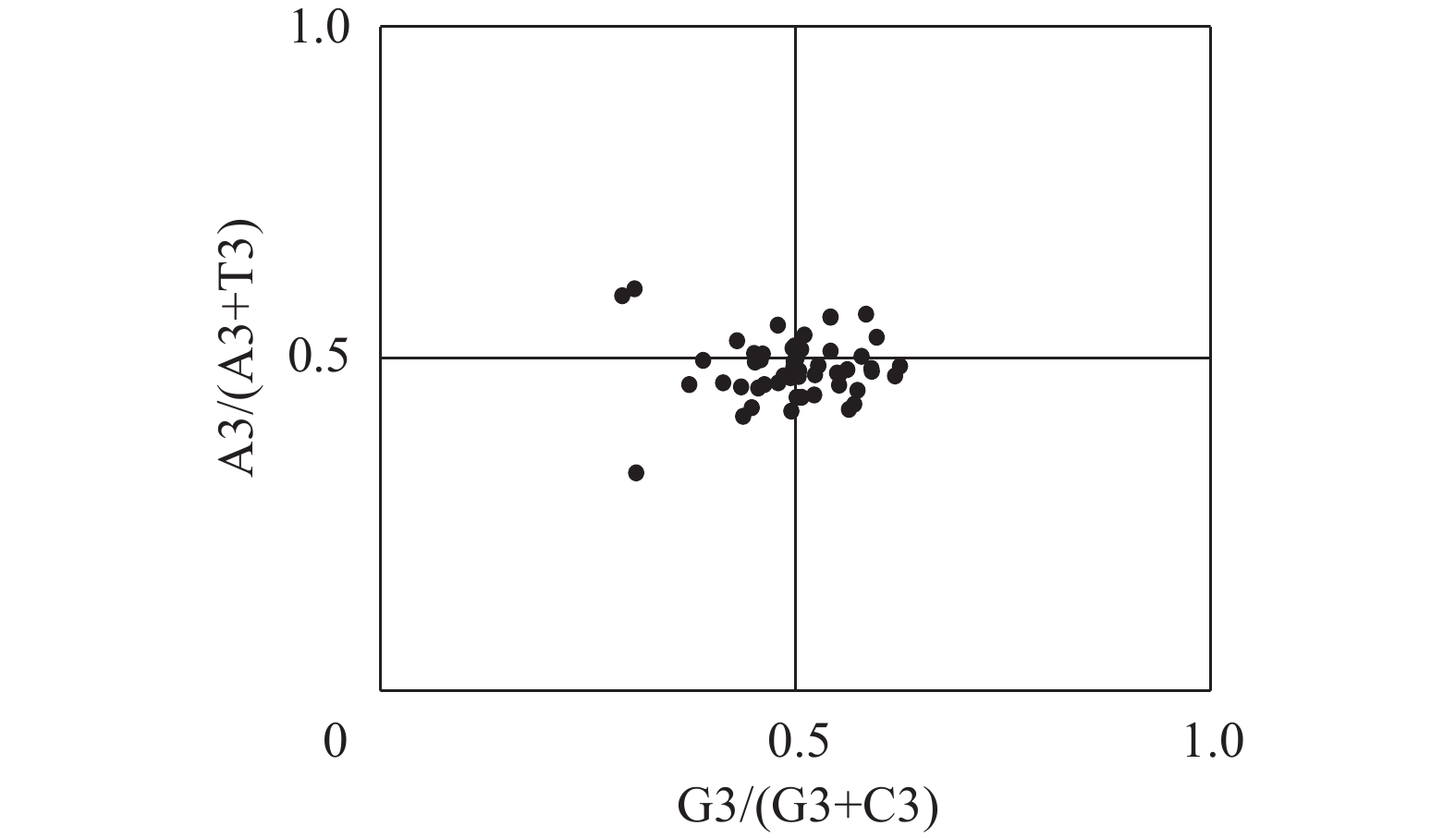
图3显示:基因位点在平面图4个区域内分布并不均匀,在A3/(A3+T3)<0.5和G3/(G3+C3)>0.5区域范围内分布最多。表明第3位碱基使用频率为:T>A、G>C,梁山慈竹叶绿体基因组密码子的第3位碱基在选择上具有偏好性,同时说明其密码子使用偏好性主要受自然选择的影响。
2.6 最优密码子的确定
对梁山慈竹的ENC进行升序排列,前10%为高表达基因,即rps18、rpl16、psbA、rps14、rps11,后10%为低表达基因,即 ycf3、cemA、ndhE、atpE、infA。梁山慈竹的RSCU和ΔRSCU表明(表5):梁山慈竹叶绿体基因组有32个高频密码子,筛选出GCA、GCU等25个高表达密码子,最终确定18个密码子作为梁山慈竹叶绿体基因组的最优密码子,分别为UAA、GCA、GCU、UUC、GGU、AAA、CUU、UUA、CCA、CCU、CAA、AGA、CGU、AGU、UCC、ACU、GUA、GUU。其中16个以A/U结尾,2个以C结尾。
表 5 梁山慈竹叶绿体基因组各氨基酸的RSCU分析及最优密码子分析Table 5 RSCU analysis and optimal codon analysis of amino acids in chloroplast genome of D. farinosus氨基酸 密码子 基因组
RSCU高表达
RSCU低表达
RSCUΔRSCU 氨基酸 密码子 基因组
RSCU高表达
RSCU低表达
RSCUΔRSCU Ter UAA*** 1.560 0 1.800 0 1.200 0 0.600 0 Met AUG 1.000 0 1.000 0 1.000 0 0 UAG 0.780 0 0.600 0 1.200 0 −0.600 0 Asn AAC* 0.520 0 0.893 6 0.625 0 0.268 6 UGA 0.670 0 0.600 0 0.600 0 0 AAU 1.480 0 1.106 4 1.375 0 −0.268 6 Ala GCA** 1.200 0 1.200 0 0.734 7 0.465 3 Pro CCA** 1.080 0 0.800 0 0.500 0 0.300 0 GCC 0.600 0 0.457 1 0.653 1 −0.196 0 CCC 1.010 0 0.800 0 1.166 7 −0.366 7 GCG 0.470 0 0.228 6 0.734 7 −0.506 1 CCG 0.430 0 0.444 4 1.000 0 −0.555 6 GCU* 1.730 0 2.114 3 1.877 6 0.236 7 CCU*** 1.480 0 1.955 6 1.333 3 0.622 3 Cys UGC** 0.470 0 0.400 0 0 0.400 0 Gln CAA* 1.530 0 1.500 0 1.368 4 0.131 6 UGU 1.530 0 1.600 0 2.000 0 −0.400 0 CAG 0.470 0 0.500 0 0.631 6 −0.131 6 Asp GAC* 0.460 0 0.500 0 0.411 8 0.088 2 Arg AGA* 1.750 0 1.723 4 1.534 9 0.188 5 GAU 1.540 0 1.500 0 1.588 2 -0.088 2 AGG 0.640 0 0.319 1 0.837 2 −0.518 1 Glu GAA 1.460 0 1.189 2 1.578 9 −0.389 7 CGA 1.270 0 1.276 6 1.395 3 −0.118 7 GAG** 0.540 0 0.810 8 0.421 1 0.389 7 CGC 0.510 0 0.319 1 0.837 2 −0.518 1 Phe UUC** 1.290 0 1.041 7 0.650 0 0.391 7 CGG 0.410 0 0.319 1 0.279 1 0.040 0 UUU 0.710 0 0.958 3 1.350 0 −0.391 7 CGU*** 1.410 0 2.042 6 1.116 3 0.926 3 Gly GGA 1.580 0 1.253 7 1.818 2 −0.564 5 Ser AGC 0.410 0 0.384 6 0.470 6 −0.086 0 GGC 0.430 0 0.417 9 0.484 8 −0.066 9 AGU** 1.250 0 1.846 2 1.411 8 0.434 4 GGG 0.760 0 0.119 4 0.363 6 −0.244 2 UCA 1.020 0 0.615 4 1.058 8 −0.443 4 GGU*** 1.240 0 2.209 0 1.333 3 0.875 7 UCC*** 1.190 0 1.769 2 0.941 2 0.828 0 His CAC** 0.530 0 0.941 2 0.571 4 0.369 8 UCG 0.550 0 0.153 8 0.705 9 −0.552 1 CAU 1.470 0 1.058 8 1.428 6 −0.369 8 UCU 1.580 0 1.230 8 1.411 8 −0.181 0 Ile AUA 0.920 0 0.850 7 0.949 4 −0.098 7 Thr ACA 1.080 0 1.181 8 1.176 5 0.005 3 AUC* 0.590 0 0.626 9 0.531 6 0.095 3 ACC 0.500 0 0.818 2 1.058 8 −0.240 6 AUU 1.480 0 1.522 4 1.519 0 0.003 4 ACG 0.480 0 0.363 6 0.588 2 −0.224 6 Lys AAA** 1.440 0 1.471 7 1.155 6 0.316 1 ACU** 1.680 0 1.636 4 1.176 5 0.459 9 AAG 0.560 0 0.528 3 0.844 4 −0.316 1 Val GUA*** 1.500 0 1.767 4 1.257 1 0.510 3 Leu CUA 0.900 0 0.833 3 1.295 5 −0.462 2 GUC 0.490 0 0 1.028 6 −1.028 6 CUC 0.420 0 0 0.545 5 −0.545 5 GUG 0.540 0 0.372 1 0.342 9 0.029 2 CUG 0.330 0 0.250 0 0.477 3 −0.227 3 GUU** 1.470 0 1.860 5 1.371 4 0.489 1 CUU* 1.290 0 1.333 3 1.227 3 0.106 0 Trp UGG 1.000 0 1.000 0 1.000 0 0 UUA*** 1.940 0 2.166 7 1.022 7 1.144 0 Tyr UAC** 0.410 0 0.521 7 0.166 7 0.355 0 UUG 1.110 0 1.416 7 1.431 8 −0.015 1 UAU 1.590 0 1.478 3 1.833 3 −0.355 0 说明: 高频密码子(RSCU>1.00)带下划线;*. ΔRSCU≥0.08;**. ΔRSCU≥0.3;***. ΔRSCU≥0.5; 加粗的密码子表示最优密码子。 2.7 梁山慈竹和其他几种生物密码子偏好性比较分析
将梁山慈竹基因组密码子使用频率与巨龙竹、粉麻竹、小叶龙竹、硬头黄竹、大肠埃希菌、烟草、拟南芥和酿酒酵母等物种的基因组密码子使用频率进行比较(图4)。结果显示:梁山慈竹与巨龙竹、粉麻竹、小叶龙竹和硬头黄竹的密码子使用频率为0.5~2.0,说明它们的密码子使用偏好性相似,推测具有亲缘关系的禾本科Gramineae牡竹属植物叶绿体基因组密码子偏好性相似;在大肠埃希菌、烟草、拟南芥和酿酒酵母的密码子使用比值中筛选≥2.0或≤0.5的密码子,分别有28和15、15、14个,表明梁山慈竹与这些物种在同义密码子的偏好性上有一定差异。
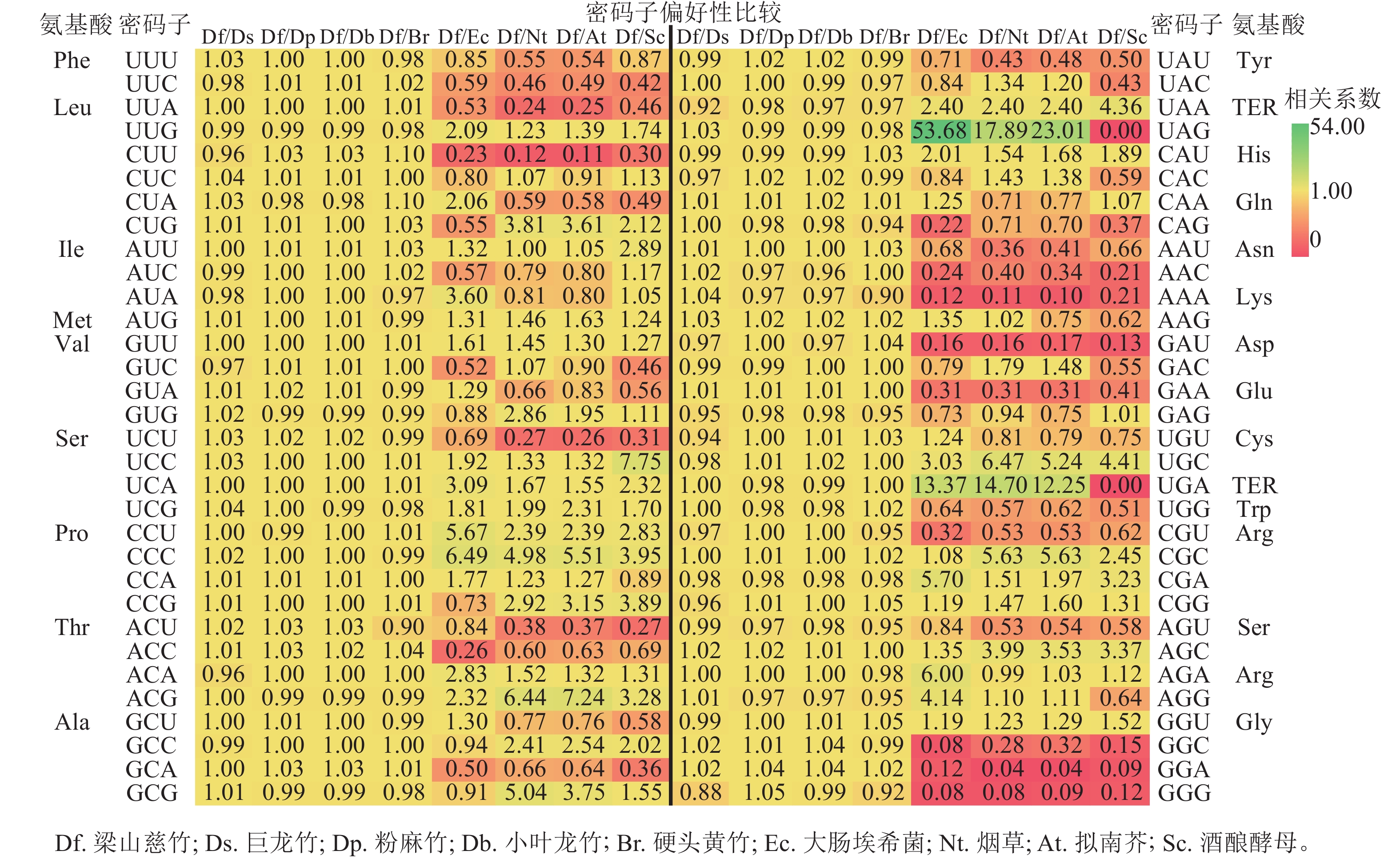 图 4 梁山慈竹与其他物种密码子偏好性比较Figure 4 Comparison of codon preference between D. farinosus and other species
图 4 梁山慈竹与其他物种密码子偏好性比较Figure 4 Comparison of codon preference between D. farinosus and other species2.8 对应分析
将梁山慈竹的51个叶绿体基因的基因功能分为光合系统基因、遗传系统基因、其他基因和未知功能基因四大类,在计算RSCU的基础上将各个基因分布到59维的向量空间。对应分析结果(图5)显示:前4个向量轴分别存在18.3%、16.8%、15.6%和15.4%的差异,前4向量轴累计差异为66.1%,4个轴对密码子均有不同程度的影响;第1轴的值大于其他轴,说明第1轴对梁山慈竹叶绿体基因组密码子偏好性的影响较大。对第1轴与CAI、CBI、FOP、ENC和GC3s等指数进行进一步的相关分析发现:梁山慈竹基因在第1轴上的坐标值与CAI (r=−0.001 7,P<0.01)、CBI (r=0.099 0,P<0.01)、FOP (r=0.083 0,P<0.01)、ENC (r=0.112 0,P<0.01)、GC3s (r=−0.145 0,P<0.01)间具有极显著的相关关系,其中CAI和GC3s第1轴具有负相关关系,表明基因组密码子的偏好性不止受单一因素的影响,自然选择、基因突变均有可能影响梁山慈竹基因组密码子使用偏好性[18]。
3. 讨论
本研究对梁山慈竹叶绿体基因组密码子进行使用偏好性分析,筛选出51条CDS序列,分析表明:GC1>GC2>GC3,密码子在3个位置上的分布并不均匀,密码子偏好使用以A或U结尾的碱基,且梁山慈竹叶绿体基因组的ENC均值为49.51,表明其叶绿体基因组密码子使用偏好性较弱。这与乳油木Vitellaria paradoxa[19]和二乔玉兰Magnolia soulangeana[20]等植物叶绿体基因组密码子偏好性相似。
对梁山慈竹叶绿体基因组密码子进行中性绘图、ENC-plot分析、PR2-plot分析和对应分析。在中性绘图分析中,回归系数为0.412 8,说明密码子偏好性更多受到自然选择的影响;在ENC-plot分析中,多数基因离标准曲线距离较远,实际ENC和预期ENC有差距,表明该部分基因的密码子偏好性主要受自然选择的影响;在PR2-plot绘图分析中,大部分基因位于平面图的右下方,即T>A、G>C,表明其密码子的使用更多受自然选择的影响。综上所述,影响梁山慈竹叶绿体基因组密码子偏好性的主要原因是自然选择。该研究结果与巨桉Eucalyptus grandi[21]、灰毛浆果楝Cipadessa cinerascens、酸枣Ziziphus jujuba var. spinosa[22]和云南油杉Keteleeria evelyniana[23]等叶绿体基因组密码子偏好性研究结果基本一致;但在对4种蔷薇科 Rosaceae果树[24]和银白杨Populus alba[25]的研究中发现:突变是影响密码子偏好性的主要因素。这说明密码子的使用偏好性受自然选择或基因突变因素影响。基于RSCU的对应分析表明:梁山慈竹的密码子使用变异原因除了突变和自然选择之外,还有其他的因素,这其中光合系统基因和遗传系统基因分布相对集中,各类基因密码子使用偏好性较为接近。该结论与木薯Manihot esculenta[26]和高山松Pinus densata[27]的研究结果一致。密码子使用频率比较结果显示:梁山慈竹与禾本科牡竹属的植物密码子偏好性相似,在基因选择外源系统表达时,可以选择密码子偏好性差异相对较小的酿酒酵母,在选择大肠埃希菌、烟草和拟南芥作为外源表达宿主时,需要根据密码子使用偏好性进行碱基优化,从而使基因在宿主体内更好地表达。
最优密码子分析表明:梁山慈竹叶绿体基因组有GCU、GAU以及GGU等18个最优密码子,最优密码子大部分以A或U结尾。该结果与抽筒竹Gelidocalamus tessellatus[28]和毛竹Phyllostachys edulis[29]叶绿体基因组最优密码子分析结果一致,这可能与亲缘关系相近,但不同物种之间叶绿体基因组进化过程中的相对保守性有关系[21]。通过筛选获取梁山慈竹偏好使用密码子,可进一步对目标基因进行密码子优化,提高梁山慈竹的竹笋产量和造纸纤维含量,以及利用新一代精准基因编辑工具CRISPR/Cas9优化梁山慈竹密码子,从而改造梁山慈竹基因组编辑的Cas9基因,提高该基因在梁山慈竹中的表达水平[30]。
4. 结论
本研究通过分析梁山慈竹叶绿体基因组的CDS序列,对梁山慈竹的叶绿体基因组进行生物信息学分析,筛选出梁山慈竹叶绿体基因组有GCU、GAU以及GGU等18个最优密码子。研究结果表明:影响梁山慈竹密码子偏好性的主要因素是自然选择。研究结果为后续在分子层面上利用基因工程开发梁山慈竹优良资源提供参考。
-
表 1 L9(33)正交表和实验结果
Table 1. Orthogonal table L9(33) and experimental results
试验编号 乙醇体积分数(A)/%( 时间(B) /h 料液比(C) 总黄酮得率/% 1 30 0.5 1:80 0.499 2 30 1.0 1:40 0.594 3 30 1.5 1:60 0.411 4 70 0.5 1:40 0.936 5 70 1.0 1:60 0.733 6 70 1.5 1:80 0.476 7 95 0.5 1:60 0.877 8 95 1.0 1:80 0.625 9 95 1.5 1:40 0.501 K1 1.504 0 2.312 0 2.031 0 K2 2.145 0 1.952 0 2.021 0 K3 2.003 0 1.388 0 1.600 0 Rj 0.214 0 0.308 0 0.144 0 说明: K为第"j"列因素3水平所对应的试验指标和; Rj为第"j"列因素的极差。  下载: 导出CSV
下载: 导出CSV
表 2 藜麦总黄酮得率的方差分析
Table 2. Variation analysis of extraction rate of flavonoids in quinoa
方差来源 平方和(S) 自由度(f) 均方(s/f) F值 临界值 乙醇体积分数(A) 0.075 5 2 0.037 8 6.51 F0.05(2,2)=19 时间(B) 0.144 6 2 0.072 3 12.47 F0.1(2,2)=9 料液比(C) 0.040 3 2 0.020 2 3.47 F0.2(2,2)=4 误差 0.011 6 2 0.005 8 总和 0.272 0 8
下载: 导出CSV
表 3 藜麦黄酮分析的回收率实验结果
Table 3. Experimental results of recovery in extraction rate of flavonoids in quinoa
试验编号 样品中含量/mg 加入芦丁量/mg 测得量/mg 回收率/% 平均回收率/% 相对标准偏差/% 1 7.134 0.100 7.441 102.86 100.25 4.22 2 14.268 0.100 13.415 93.37 3 7.039 0.100 7.441 104.23 4 14.078 0.100 14.265 100.61 5 7.998 0.100 8.368 103.33 6 15.996 0.100 15.630 97.10
下载: 导出CSV
表 4 藜麦黄酮得率的基因型差异
Table 4. Genotype variation of extraction yields of flavonoids in quinoa
品种 叶表情况 叶形状 总黄酮得率/% TEMUCO Quinoa TRADITIONAL 绿色 提琴形锯齿状 0.635 QuinoaB.Rain Sow 绿色 提琴形锯齿状 0.818 Temuco 绿色 提琴形锯齿状 0.805 1591 Quinoa Cherry 绿色 提琴形锯齿状 0.751 Tomico Quinoa 绿色 提琴形锯齿状 0.472 CQ-TEMVCC 绿色 提琴形锯齿状 0.477 PI814932 绿色 提琴形锯齿状 0.933 PI596293 绿色 提琴形锯齿状 0.512 Tumuco(7)hybrids 绿色 提琴形锯齿状 0.572 PI596498 绿色(表面覆有紫色粉层) 卵形略有锯齿 0.215 平均值 0.619 变异系数/% 34.44
下载: 导出CSV
-
[1] 朱剑宏.南美藜的化学组成和营养价值[J].成都大学学报:自然科学版, 2002, 21(2):24-28. ZHU Jianhong. The chemical composition and nutrition of quinoa[J]. J Chengdu Univ Nat Sci, 2002, 21(2):24-28. [2] VEGA-GÁLVEZ A, MIRANDA M, VERGARA J, et al. Nutrition facts and functional potential of quinoa (Chenopodium quinoa Willd.), an ancient Andean grain:a review[J]. J Sci Food Agric, 2010, 90(15):2541-2547. [3] OSHODI A A, OGUNGBENLE H N, OLADIMEJI M O. Chemical composition, nutritionally valuable minerals and functional properties of benniseed (Sesamun radiatum), pearl millet (Pennisetum typhoides) and quinoa (Chenopodium quinoa) flours[J]. Int J Food Sci Nutr, 1999, 50(5):325-331. [4] COMAI S, BERTAZZO A, BAILONI L, et al. The content of proteic and nonproteic (free and protein-bound) tryptophan in quinoa and cereal flours[J]. Food Chem, 2007, 100(4):1350-1355. [5] JACOBSEN S E, MUJICA A, JENSEN C R. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors[J]. Food Rev Int, 2003, 19(1/2):99-109. [6] 王晨静, 赵习武, 陆国权, 等.多功能藜麦的研究进展[J].浙江农林大学学报, 2014, 31(2):296-301. WANG Chenjing, ZHAO Xiwu, LU Guoquan, et al. A review of characteristics and utilization of Chenopodium quinoa[J]. J Zhejiang A & F Univ, 2014, 31(2):296-301. [7] 刘步东, 李登昌, 丁宁.勿忘我花中总黄酮含量测定[J].中兽医医药杂志, 2013(1):41-42. LIU Budong, LI Dengchang, DING Ning. Content determination of total flavonoid in Myosotis sylvatica flower[J]. J Trad Chin Vet Med, 2013(1):41-42. [8] 陆国权, 任韵, 唐忠厚, 等.甘薯黄酮类物质的提取及其基因型差异研究[J].浙江大学学报:农业与生命科学版, 2005, 31(5):541-544. LU Guoquan, REN Yun, TANG Zhonghou, et al. Flavonoid extraction and flavonoid content genotypic variation in sweetpotato storage roots[J]. J Zhejiang Univ Agric & Life Sci, 2005, 31(5):541-544. [9] 林建原, 季丽红.响应面优化银杏叶中黄酮的提取工艺[J].中国食品学报, 2013, 13(2):83-90. LIN Jianyuan, JI Lihong. Optimization of flavonoids from Ginkgo biloba using response surface analysis[J]. J Chin Inst Food Sci Technol, 2013, 13(2):83-90. [10] 张斌, 孙兰萍, 马龙, 等.大孔树脂分离纯化花生壳总黄酮的研究[J].中国粮油学报, 2010, 25(2):126-130. ZHANG Bin, SUN Lanping, MA Long, et al. Separation and purification of flavonids from peanut hull by macroporous resins[J]. J Chin Cerea Oils Assoc, 2010, 25(2):126-130. [11] 高淑云, 徐婷婷.响应面优选大蒜中黄酮提取工艺研究[J].中国调味品, 2013, 38(3):44-49. GAO Shuyun, XU Tingting. Optimization of extraction technique of flavonoids from garlic by response surface methodology[J]. China Condiment, 2013, 38(3):44-49. [12] 王静霞, 黄艳菲, 赵小燕, 等.荞麦和商品苦荞茶中总黄酮的含量测定[J].食品工业科技, 2013, 34(2):58-60. WANG Jingxia, HUANG Yanfei, ZHAO Xiaoyan, et al. Determination of total flavonoids in buckwheat and buckwheat goods[J]. Sci Technol Food Ind, 2013, 34(2):58-60. [13] 李长新, 张鲁刚, 孙希禄, 等.紫色小白菜花青素的提取工艺优化[J].西北农林科技大学学报:自然科学版, 2011, 39(9):200-206. LI Changxin, ZHANG Lugang, SUN Xilu, et al. Optimization of extracting process of anthocyanidin in purple pakchoi[J]. J Northwest A & F Univ Nat Sci Ed, 2011, 39(9):200-206. [14] 贾霖, 黄国清, 肖军霞.金银花中黄酮类化合物的提取工艺[J].食品研究与开发, 2013, 34(9):41-43. JIA Lin, HUANG Guoqing, XIAO Junxia. Study on extraction of flavonoids from honeysuckle[J]. Food Res Develop, 2013, 34(9):41-43. [15] 李琼.超声波辅助法提取芸豆种子黄酮类化合物研究[J].食品研究与开发, 2012, 33(2):40-42. LI Qiong. Study on the extraction of bioflavonoid compound from seeds of kidney beans by the super-sonic wave auxiliary method[J]. Food Res Develop, 2012, 33(2):40-42. [16] 任顺成, 王鹏, 王国良, 等.常用食用豆类中黄酮类化合物含量的测定[J].中国粮油学报, 2009, 24(7):132-137. REN Shuncheng, WANG Peng, WANG Guoliang, et al. Determination of flavonoid contents in common edible legumes[J]. J Chin Oils Assoc, 2009, 24(7):132-137. [17] de SIMONE F, DINI A, PIZZA C, et al. Two flavonol glycosides from Chenopodium quinoa[J]. Phytochemistry, 1990, 29(11):3690-3692. [18] OGUNGBENLE H N. Nutrional evaluation and functional properties of quinoa (Chenopodium quinoa) flour[J]. Int J Food Sci Nutr, 2003, 54(2):153-158. [19] HILAL M, PARRADO M F, ROSA M, et al. Epidermal lignin deposition in quinoa cotyledons in response to UV-B radiations[J]. Photochem Photobiol, 2004, 79(2):205-210. [20] JAMES A, LILIAN E. Quinoa (Chenopodium quinoa Willd.):composition, chemistry, nutritional and functional properties[J]. Adv Food Nutr Res, 2009, 58:1-31. [21] 杨美华, 卢兖伟, 匡岩巍, 等.柱色谱-紫外分光光度法测定广金钱草中总黄酮的含量[J].中草药, 2004, 35(6):688-690. YANG Meihua, LU Yanwei, KUANG Yanwei, et al. Quantitative determination of total flavonoids in Desmodium styracifolium by Column chromatogram-UV spectrophotometry[J]. Chin Trad Herb Drug, 2004, 35(6):688-690. [22] PAŚKO P, BARTOŃ H, ZAGRODZKI P, et al. Antnocyanins, total polyphenols and antioxidant activity in amaranth and quinoa seeds and sprouts during their growth[J]. Food Chem, 2009, 115(3):994-998. [23] 陈君, 周光明, 杨远高, 等.离子液体-超声辅助萃取/高效液相色谱法测定白花杜鹃叶中的黄酮[J].分析测试学报, 2013, 32(3):341-345. CHEN Jun, ZHOU Guangming, YANG Yuangao, et al. Simultaneous determination of flavonoids in Rhododendron mucronatum leaves by lonic liquid-based ultrasonic-assisted extraction combined with high performance liquid chromatography[J]. J Instrum Anal, 2013, 32(3):341-345. [24] 秦祖杰, 梁洁, 孙正伊.赪桐根中总黄酮的提取工艺优选[J].中国实验方剂学杂志, 2013, 19(5):60-62. QIN Zujie, LIANG Jie, SUN Zhengyi. Optimization of extraction technology of total flavones from roots of Clerodendrum japonicum[J]. Chin J Exp Trad Med Form, 2013, 19(5):60-62. [25] 刘璐, 付明哲, 王侠, 等. 5种棘豆总黄酮含量的测定比较[J].草叶科学, 2011, 28(4):683-686. LIU Lu, FU Mingzhe, WANG Xia, et al. Determination of total flavonoids on 5 species of Oxytropis[J]. Pratacul Sci, 2011, 28(4):683-686. [26] 陈金娥, 赵金玲, 张海容.青蒿中黄酮、多酚和VC含量测定及抗氧化性研究[J].食品研究与开发, 2013, 34(4):1-3. CHEN Jin'e, ZHAO Jinling, ZHANG Hairong. Determination of flavonoids, polyphenol and vitamin C and its antioxidant properties of Atemisia annual L.[J]. Food Res Dev, 2013, 34(4):1-3. [27] 陈全斌, 谭冬明, 李佳, 等.甜菜植株不同部位总黄酮含量的测定[J].林业科技, 2007, 32(4):68-70. CHEN Quanbin, TANG Dongming, LI Jia, et al. Determination of total flavonoids in the different parts of Rubus suavissimus[J]. For Sci & Technol, 2007, 32(4):68-70. [28] 程水源, 王燕, 李俊凯, 等.银杏叶片色素含量与黄酮含量关系的研究[J].林业科学, 2001, 37(5):31-34. CHEN Shuiyuan, WANG Yan, LI Junkai, et al. Study on the realationship between the flavonoids and pigments in Ginkgo biloba leaf[J]. Sci Silv Sin, 2001, 37(5):31-34. [29] 谢宝东, 王华田.银杏不同家系和无性系叶片黄酮与内酯含量变异[J].林业科技开发, 2008, 22(2):33-37. XIE Baodong, WANG Huatian. Genetic variation of flavonoid and terpene content in Ginkgo biloba leaves and growth among various families and clones[J]. China For Sci Technol, 2002, 22(2):33-37. 期刊类型引用(1)
1. 何刀山,阳小强,秦雅林,何海燕,谢维,罗治国,李鹏. ‘湘辣14号’叶绿体基因组密码子偏好性分析. 中国果菜. 2025(01): 47-54+79 .  百度学术
百度学术其他类型引用(0)
-

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.04.007
