




-
蕙兰Cymbidium faberi是中国传统名花之一,栽培历史悠久。近年来,由于自然环境改变和人类活动干扰等原因,蕙兰的自然种群数量在不断减少,已被列为《国家重点保护野生植物名录》二级保护植物。对野生蕙兰的历史分布格局及影响因素进行研究,有助于了解蕙兰自然种群的演变规律及生长环境特点,引导相应保育措施的制定,遏制或减缓野生蕙兰的濒危进程,为蕙兰种质资源的保育工作提供科学依据。
目前,对野生蕙兰的研究集中在根部内生真菌的生态学特性及共生作用[1−2]、遗传多样性[3−4]、基因功能的解析及验证[5−6]、野外种质资源调查[7]及潜在分布[8]等方面。其中,野生蕙兰的资源分布研究时段集中于1949年以后,而对历史数据研究较少;研究角度多以资源的性状等本体属性为主,空间信息属性常被忽略;研究区域集中于单一区域[9],系统性构建全国野生蕙兰种质资源分布的研究相对匮乏。地理信息技术的发展,为历史数据的量化、野生植物资源信息化提供了可能[10]。地理信息系统在预测物种分布、可视化物种扩散迁移路径模拟[11−12]及物种生态适宜性评价[13−14]等研究中发挥了重要作用。关于植物时空分布长期演化的历史,前人多借助孢粉化石[15−16]。对近几百年内植物种群的动态变化,前人则多借鉴方志文献[17−18]。自1368年以来,诗文、方志、游记出现大量关于野生蕙兰的记载,为蕙兰自然种群的历史分布研究提供可靠的数据来源。基于此,本研究在梳理野生蕙兰空间地理信息分布、构建野生蕙兰历史地理信息数据库的基础上,总结其时空分布特征、分布格局及迁移方向,探讨蕙兰的自然种群分布区域与自然环境变化的关系,旨在为中国野生蕙兰种群的保育提供科学依据。
-
本研究所涉及的野生蕙兰历史分布数据主要来源于各历史时期的方志、植物志以及林史资料、专著、中国数字植物标本馆、中国植物图像库(
http://ppbc.iplant.cn/ )等。①方志:梳理1368—1912年的《八闽通志》(2017年福建人民出版社出版)和《浙江通志》(1991年上海古籍出版社出版)等110余本方志;②植物志:《中国植物志》(2016年科学出版社出版)和《农业野生植物资源调查与保护系列丛书》(2015年中国农业出版社出版) 等10余本植物志;③专著:《兰蕙纵横》(2018年中国林业出版社出版)和《夏兰》(2004年杭州出版社出版)等20余本专著。本研究以南京林业大学白雪锋等提出的从方志中提取历史空间信息的方法为参考依据,以“应采尽采”为原则,共搜集到980处野生蕙兰资源分布区域,数据空间坐标定位方法主要分为直接定位法和间接定位法2种。 -
从中国植物数字标本馆、中国植物图像库以及相关调查资料中,直接对野生蕙兰采集点位置的经纬度进行记录;又如《中国养兰集成》[19]第8章第1节中“兰花的引种”中详述记录了野生蕙兰具体的采集位置。可采用直接定位采集的野生蕙兰资源集中于1949年以后,数量为560处,占总数的57.1%。
-
间接定位法是指无法通过现有研究成果进行空间定位,需要通过文献史料中的信息进行定位的方法。通过对各方志的山川卷、物产卷等卷中“蕙”“九节兰”“夏兰”“蝉兰”等异名进行查阅,抽取地理位置的相关描述。主要可分为方位关系、拓扑关系和距离关系3种类型。①方位关系。方位指地理空间中的东西南北(基本方位)、上下左右(相对方位)等概念。相关文献史料中常用山水等地名作为参考点,描述野生蕙兰的所处位置。如《广东通志·卷十》山川“合兰洲”中记录道:“三门海中与龙穴相对,上多兰蕙,有二石并峙潮至此则合”;《陕西通志·卷三》山川“石瓮谷”中记录道:“东绣岭左谷中尽植兰蕙亦谓之芝兰谷”。②拓扑关系。拓扑关系是指风景资源之间的空间结构关系,包括拓扑邻接、拓扑关联和拓扑包含等类型。相关史料中多以“$在 \cdots \cdots 之 $内”“$与 \cdots \cdots 相 $邻”等形式出现,如《贵州通志·卷十五》记录道:“贵阳府境山谷中亦有蕙”;《兰蕙同心录》载“赤蕙永春梅产富阳砂石山中”。③距离关系。距离关系包含定性和定量2种方法。定性距离通常描述为“$在 \cdots \cdots 附 $近”“$在 \cdots \cdots 之 $间”。定量距离则描述为“城东八里”“西约百步”等。定性距离可参考方位关系和拓扑关系的原则,结合史料文本进行定位。而定量距离则可换算后直接定位。1644—1912年每里合576 m。《广东通志·卷十三》载:“阳山县香山在城东十里一名香冈山上,多蕙草”;《广西通志·卷十四》山川卷“东香山”中载:“东香山在州西北四十里,山势高险,芳兰异蕙四时有焉”。
-
本研究中,1970—2020年气候数据来源于WorldClim (
https://worldclim.org ),1901—1970年的数据来源于CRU TS ,1901年之前的气候数据来源于相关文献[20−22]。 -
土壤数据(包括土壤类型、土壤酸碱度等)来源于中国科学院南京土壤研究所(
www.issas.ac.cn )。 -
本研究数字高程模型(DEM)数据来源于地理空间数据云(
www.gscloud.cn ),空间分辨率为30 m。 -
本研究中的河网数据是基于空间分辨率为30 m的DEM数据,借助ArcGIS中的“Spatial Analyst”工具对其进行河网提取,数据集包括中国所有河网和流域面积大于100 km²的子流域。
-
基于ArcGIS 10.8.2平台,借助核密度分析[23]及最近邻指数(R)[24],分析不同时期野生蕙兰的分布区域,当R<1时,说明要素呈聚集性分布,当R>1时,说明要素呈均匀分布;借助标准差椭圆[25],探究自1368年以来野生蕙兰的分布重心与迁移方向,椭圆的长短半轴差距越大,则野生蕙兰分布格局的方向性越明显。
-
基于SPSS Statistics数据编辑器,运用逻辑(Logistic)回归方法,建立野生蕙兰的历史分布模型,探究野生蕙兰分布与自然环境之间的关系。方程如下[24]:Logit(P)=β0 +β1x1+β2x2+β3x3$+\cdots + $βmXm。其中:P为野生蕙兰存在的概率,β0为回归截距,代表直线的起点;βm为回归系数,表示在其他所有自变量保持不变的情况下,自变量xm对P的影响程度;xm表示影响野生蕙兰历史分布的自然因子;m表示自然因子数量(表1)。
表 1 野生蕙兰历史分布模型自变量
Table 1. Historical distribution model independent variables of C. faberi
自然因子 描述 数据来源及处理工具 自然因子 描述 数据来源及处理工具 海拔 地形分析因子 DEM数据 气温 年均气温 BIO1及CRU TStemp数据集 坡度 地表起伏形态 坡度工具 降水量 年均降水量 BIO12及CRU TSpre数据集 坡向 坡向 坡向工具 水源距离 分布点至水系的距离 河网数据 土壤类型 分布区域的土壤类型 土壤数据 -
以不同时期方志撰写的年代为划分依据,以每个时期的结束时间作为时间断面,将中国野生蕙兰时空演变特征划分为1368—1644、1644—1912、1912—1949、1949—1978年以及1978年至今共5个时期。
-
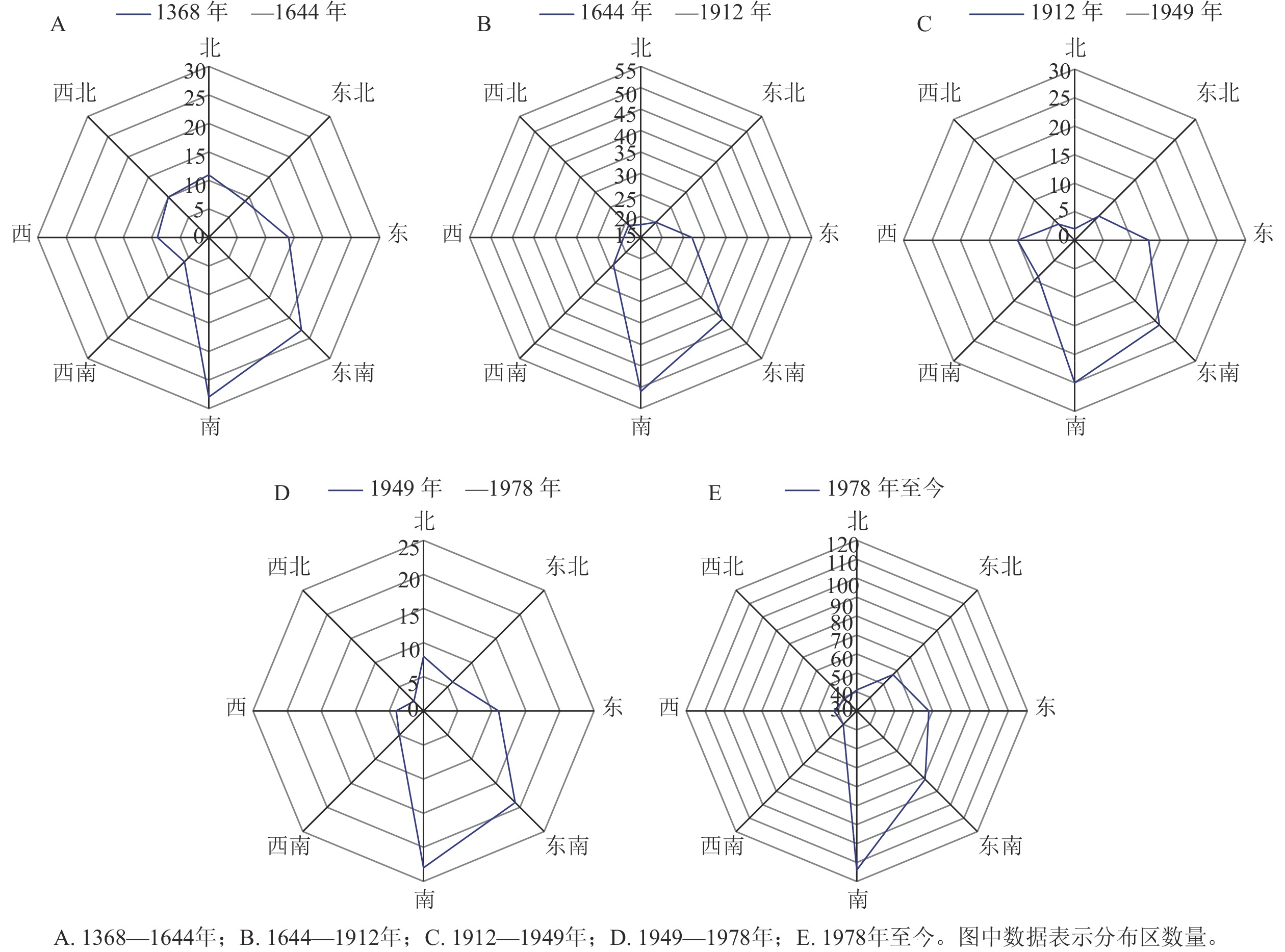
总体上看,野生蕙兰集中分布于南方地区,特别是东南沿海区域及西南地区。1368—1644年所载野生蕙兰分布区域110处,R为0.80,表现为聚集分布,主要的聚集区域为江南、福建、广东和四川、云南交界处,其中江南为高密度区。1644—1912年分布区域多达220处,R为0.58,江南、广东、福建地区聚集增强,四川、云南交界处一带的聚集程度减弱;华北中部、东北南部有少量的野生蕙兰分布点。1912—1949年所载野生蕙兰分布区域共90处,R为0.72,表现为聚集分布;聚集程度较1644—1912年减弱,但江南地区仍为高密度聚集区,而福建、广东地区的聚集程度减弱,香港有少量分布。1949—1978年所载野生蕙兰的分布区域仅78处,R为0.82,江南地区仍为野生蕙兰的主要聚集区,四川、重庆、贵州一带的聚集程度持续增强。1978至今所载野生蕙兰分布区域共482处,R为0.55,出现明显的多点聚集分布态势,江南、华中、云贵高原等地为主要聚集区,北京、天津、河北和黄河中游一带有少量分布。
-
从野生蕙兰分布中心的偏移情况来看(表2):1368—1912年,分布中心由湖南省岳阳市逐渐向江西省九江市迁移,椭圆短半轴变长,向心力逐渐减弱。1912—1949年,分布中心逐渐向江西省宜春市北部偏移,椭圆长短半轴的差值增大,扁率变大,说明1912—1949年野生蕙兰向西南方向偏移的趋势更为显著;1949—1978年,野生蕙兰的分布中心点逐渐由江西省宜春市北部向湖北省荆州市南部迁移;1978年至今,分布中心点迁移变化不大。总体来看,从1368年至今,野生蕙兰分布中心整体上呈现向高纬度地区迁移的趋势,分布的方向性由强变弱,分布范围进一步扩大,离散度先降低后增高,空间聚合度呈下降趋势。
表 2 1368年以来野生蕙兰资源分布标准差椭圆分析
Table 2. Analysis for the standard deviational ellipse of the distribution sites of wild C. faberi resources since the year of 1368
年份 椭圆周长/km 椭圆面积/km² 椭圆中心点(N) 椭圆中心点(E) 长半轴/km 短半轴/km 椭圆的方向角度/(°) 1368—1644 72.088 167 208.792 730 28.584 810° 113.503 226° 16.941 399 3.924 427 91.145 131 1644—1912 41.965 962 120.646 298 29.519 570° 116.191 821° 8.548 608 4.492 692 79.365 494 1912—1949 46.740 223 112.516 143 28.872 057° 114.902 188° 10.549 990 3.395 456 80.062 387 1949—1978 42.521 332 116.530 018 29.564 576° 112.446 761° 8.944 940 4.147 208 88.409 640 1978年至今 39.360 687 105.286 125 29.364 727° 112.674 798° 8.056 393 4.160 246 78.474 936 -
1368—1644年总体干冷,但江南、福建、江西、广东、云南洱海及贵州却温暖湿润[26−27],是这一时期野生蕙兰资源分布的聚集地区;较为干旱的西北地区(如甘肃南部)及川北地区分布较少。结合不同时期气候因子作用下野生蕙兰的分布情况来看(表3):1644—1912年江南、福建、江西、广东等地区气温较低,但降水充沛[27−28],野生蕙兰分布区域较多;四川、云南地区年平均气温较低,降水少,聚集程度减弱。1912—1949年,除四川、陕西、安徽外,整体气温上升,江南地区仍为重要聚集区域;云南、贵州地区平均气温相较于1644—1912年上升了0.3~0.6 ℃,聚集程度较1644—1912年增强。总体上看,1912—1949年气候温暖湿润,西南地区相较于1644—1912年有回温现象,降水量有所增加,野生蕙兰资源分布中心又转向西南方向偏移。1949—1978年,江南、四川、重庆地区的年均气温为10.1~17.9 ℃,年降水量约687.2~1 480.6 mm,野生蕙兰的聚集程度增强。陕南地区的降水量增加,分布数量有所增多。1978年至今,中国气候明显变暖。江南、广东、福建等地区的年降水量为1 300.0~1 950.0 mm;华中地区年降水量增多至836.9~1 766.8 mm。野生蕙兰呈现多点聚集分布,并逐渐向纬度更高的华中、陕南地区扩展。
表 3 不同时期气候因子与野生蕙兰分布情况
Table 3. Distribution sites of wild C. faberi under the effect of climatic factors in various periods
分布区 1644—1912年 1912—1949年 气温/℃ 降水量/mm 分布区数量/个 气温/℃ 降水量/mm 分布区数量/个 江苏 13.3~15.5 850.0~983.6 34 13.0~16.2 807.0~901.0 9 浙江 13.6~16.7 750.0~983.6 62 13.6~19.0 901.8~980.0 31 安徽 13.7~16.1 750.0~980.2 19 12.9~15.3 807.7~980.3 7 上海 14.9~15.8 810.0~820.0 5 11.2~13.4 807.4~901.8 1 福建 15.3~21.0 800.0~850.0 21 13.0~19.8 901.8~980.4 4 广东 17.8~23.2 750.0~900.0 8 16.2~22.8 901.8~980.4 5 广西 16.3~22.8 600.0~897.0 3 16.2~22.8 567.7~925.6 1 云南 7.1~23.2 295.0~950.0 10 7.8~24.8 308.2~980.4 15 四川 −1.6~18.3 51.0~450.0 9 −0.8~18.5 80.3~901.8 6 贵州 14.6~18.1 413.0~802.0 7 14.8~18.4 697.4~920.6 2 湖南 15.2~17.4 600.0~950.0 7 15.4~17.3 901.8~980.4 2 湖北 11.3~16.2 250.0~983.6 5 10.4~16.3 697.4~980.4 2 江西 15.3~19.1 750.0~930.0 6 15.2~18.9 901.8~980.4 2 重庆 12.1~18.2 250.0~700.0 0 16.2~19.6 807.5~901.8 0 陕西 6.3~13.3 169.0~450.0 5 5.7~13.0 567.7~901.8 1 甘肃 −4.8~11.8 51.0~169.0 3 −4.2~9.4 80.2~567.8 0 台湾 11.1~20.3 897.0~983.0 2 9.4~19.6 901.8~980.4 0 山东 10.4~14.1 450.0~950.0 2 5.7~13.0 807.4~901.8 0 河南 11.1~15.3 350.0.0~897.0 4 9.4~16.2 697.4~901.8 1 河北 2.6~13.7 543.0~820.0 2 2.4~13.0 807.5~890.5 0 山西 4.4~12.5 100.0~450.0 1 2.4~9.4 697.4~901.8 0 北京 1.3~11.4 450.0~700.0 2 2.4~13.0 901.8~980.4 0 吉林 0.3~6.2 680.0~950.0 1 −0.8~5.7 807.5~980.4 0 香港 21.8~24.3 802.0~897.0 2 16.2~19.6 901.8~980.4 0 西藏 −4.1~19.5 51.0~450.0 0 −9.4~19.4 50.0~550.0 0 海南 19.1~24.3 295.7~413.6 0 19.6~24.8 450.0~650.0 1 表 3 (续)
Table 3. Continued
分布区 1949—1978年 1978年至今 气温/℃ 降水量/mm 分布区数量/个 气温/℃ 降水量/mm 分布区数量/个 江苏 13.0~16.0 687.5~1 294.5 2 14.1~16.7 982.5~1 320.0 15 浙江 13.0~19.3 1 294.5~1 670.0 16 14.6~18.3 1 294.5~1 701.0 71 安徽 13.0~16.0 687.5~1 572.0 5 14.1~18.3 690.2~1 639.2 39 上海 15.9~17.2 982.3~1 294.5 0 15.9~16.1 983.5~1 310.4 1 福建 16.0~20.3 982.3~1 572.1 6 15.7~21.4 1 024.2~1 653.0 11 广东 16.0~24.5 1 294.5~2 070.9 2 18.3~24.7 1 350.0~2 600.0 21 广西 16.3~24.5 982.3~1 971.0 2 18.3~24.6 723.9~2 983.8 12 云南 1.84~24.5 687.5~1 572.1 3 2.8~24.6 781.8~2 118.2 17 四川 −1.5~19.3 444.6~1 294.5 13 −3.4~18.9 601.2~1 294.5 14 贵州 9.1~19.4 687.5~1 082.3 3 9.9~18.4 687.5~1 449.7 84 湖南 13.0~19.3 982.3~1 374.2 0 14.2~20.7 1 150.0~2 100.0 25 湖北 9.1~16.7 687.5~1 380.6 1 9.9~18.4 860.5~2 100.0 84 江西 16.0~20.3 1 294.5~1 970.9 5 15.6~19.8 1 143.2~1 935.0 19 重庆 13.0~19.3 982.3~1 572.1 5 14.2~19.4 1 000.0~1 572.1 8 陕西 6.2~13.0 444.6~982.3 5 7.1~14.2 460.2~904.5 15 甘肃 −4.8~9.1 187.8~687.5 1 −6.5~9.9 136.6~734.9 6 台湾 9.1~20.6 1 294.5~2 800.5 0 9.4~23.5 1 294.5~2 760.0 2 山东 9.1~16.0 687.5~982.3 0 9.5~16.5 565.5~982.3 7 河南 13.0~16.3 450.6~1 094.5 8 13.5~17.3 500.4~1 100.5 22 河北 −1.5~14.1 444.6~687.5 0 −0.3~14.2 503.4~687.5 1 山西 1.8~10.9 444.6~687.5 0 2.8~14.2 567.2~864.9 2 北京 5.3~13.0 444.6~687.5 1 6.1~14.2 374.6~687.5 3 吉林 −1.5~9.1 350.8~982.3 0 −0.3~9.9 380.5~703.2 0 香港 16.0~24.5 1 294.5~2 070.9 0 18.3~24.7 1 350.0~2 600.0 1 西藏 −10.5~19.3 85.0~687.5 0 −6.5~23.5 76.2~700.2 1 海南 19.3~24.5 1 572.0~1970.0 0 18.3~27.1 1 320.0~2 600.0 1 -
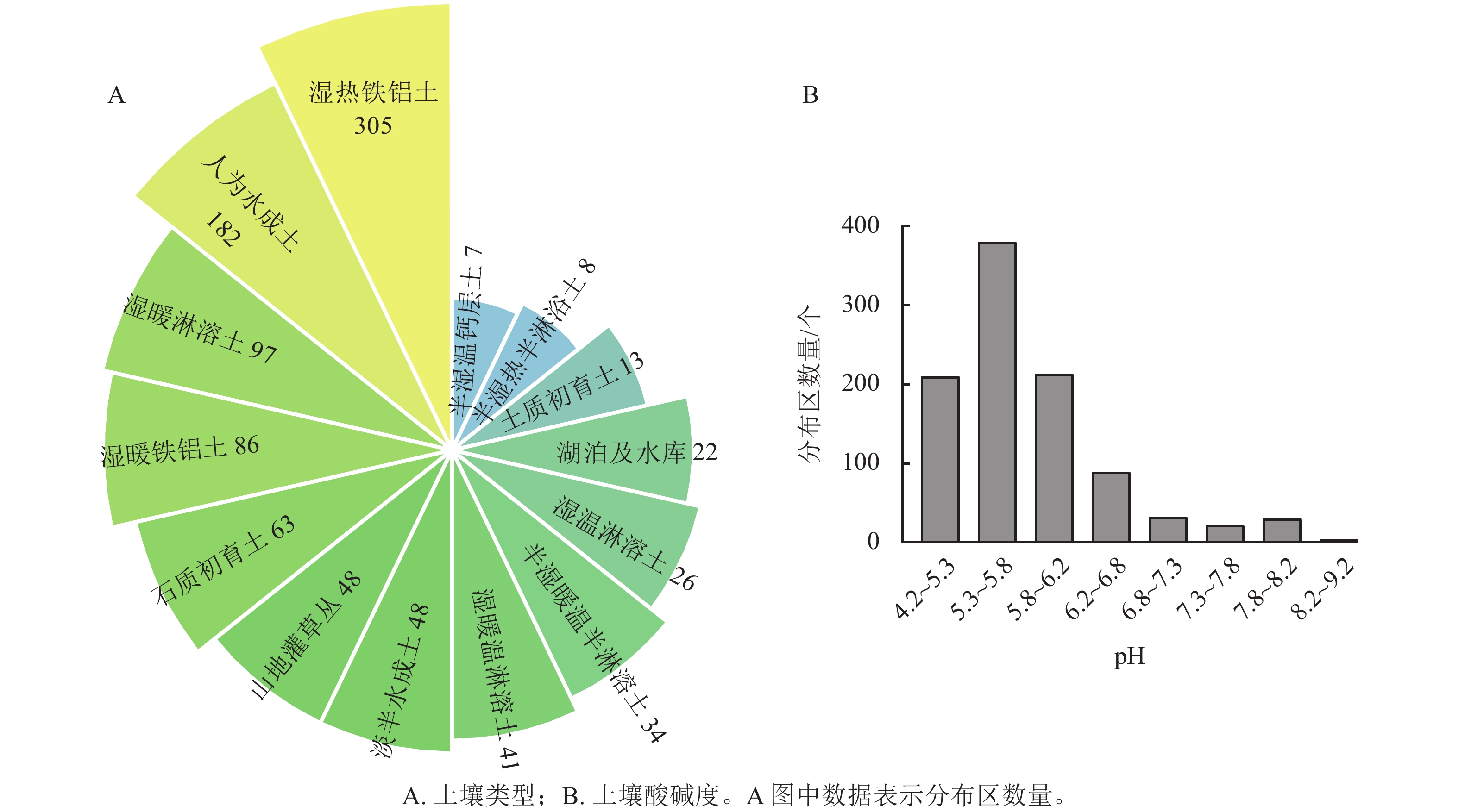
野生蕙兰分布区域的土壤类型主要为温热铁铝土和人为水成土。温热铁铝土含有氧化铁和氧化铝,具有富铁作用;人为水成土肥力较高、微生物酶活性较强。野生蕙兰分布区域的土壤pH大多数为5.3~6.2(图1A~B)。
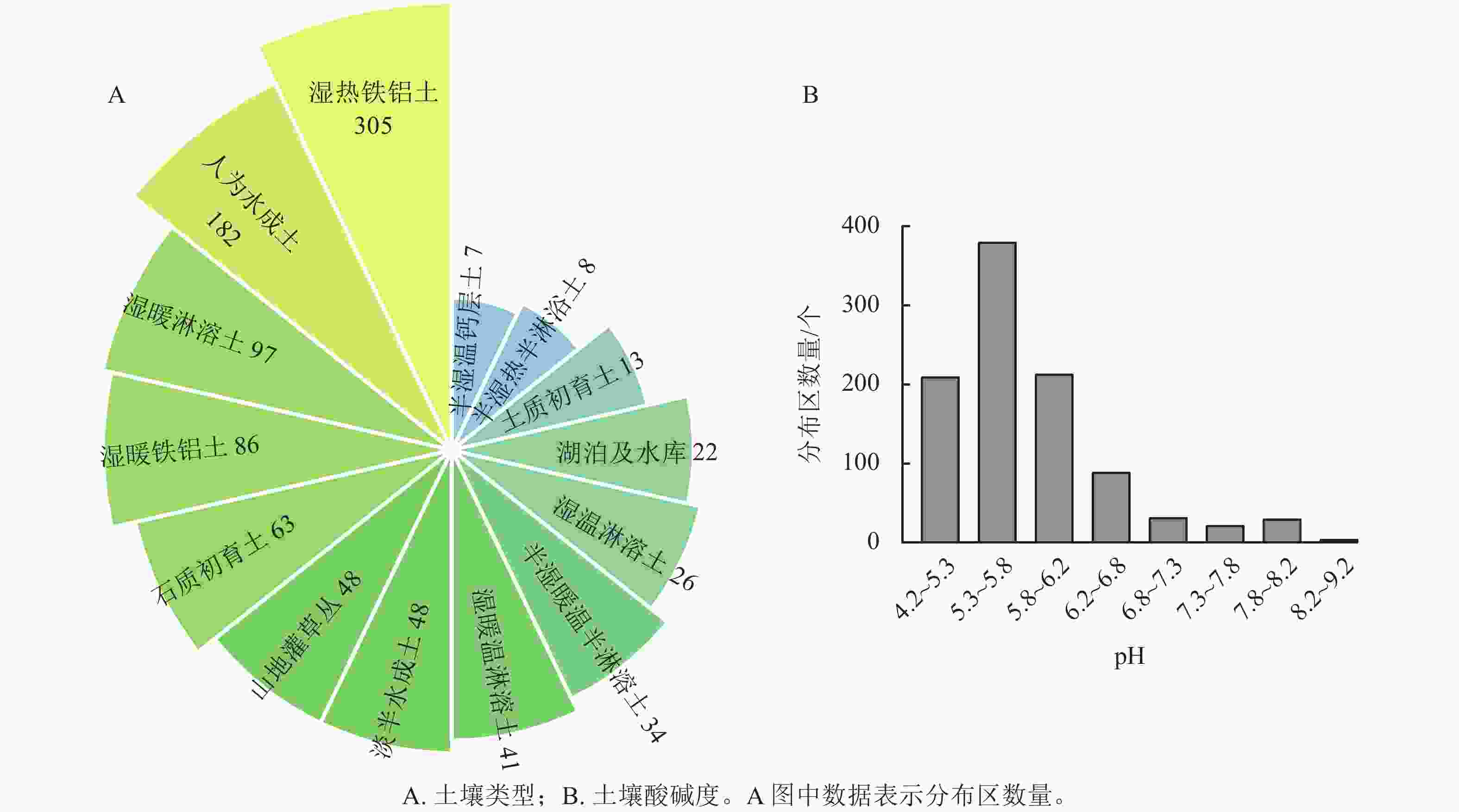
图 1 野生蕙兰在不同土壤中的分布区数量
Figure 1. Numbers for wild C. faberi in different soil types
-
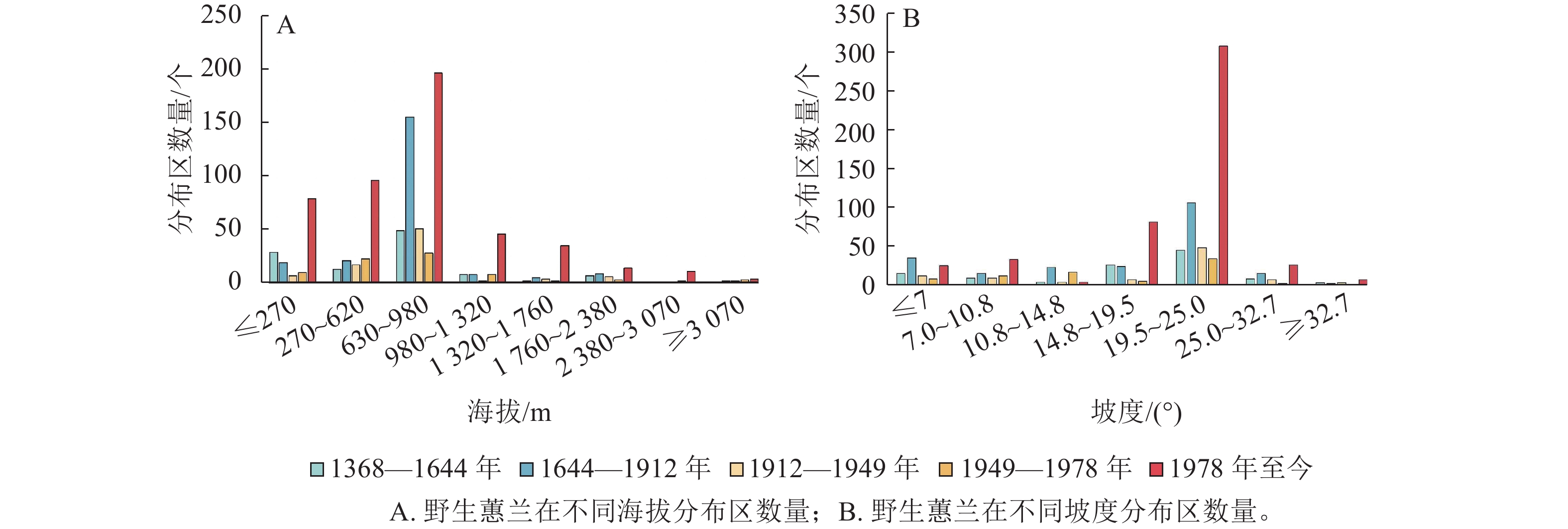
海拔、坡度和坡向等影响着野生蕙兰的分布。各时期的野生蕙兰主要分布在海拔620~980 m的区域 (图2A)。叠加统计分析后发现:野生蕙兰大多数生长在19.5°~25.0°的斜坡上(图2B)。野生蕙兰的分布以南坡居多,北坡最少(图3)。
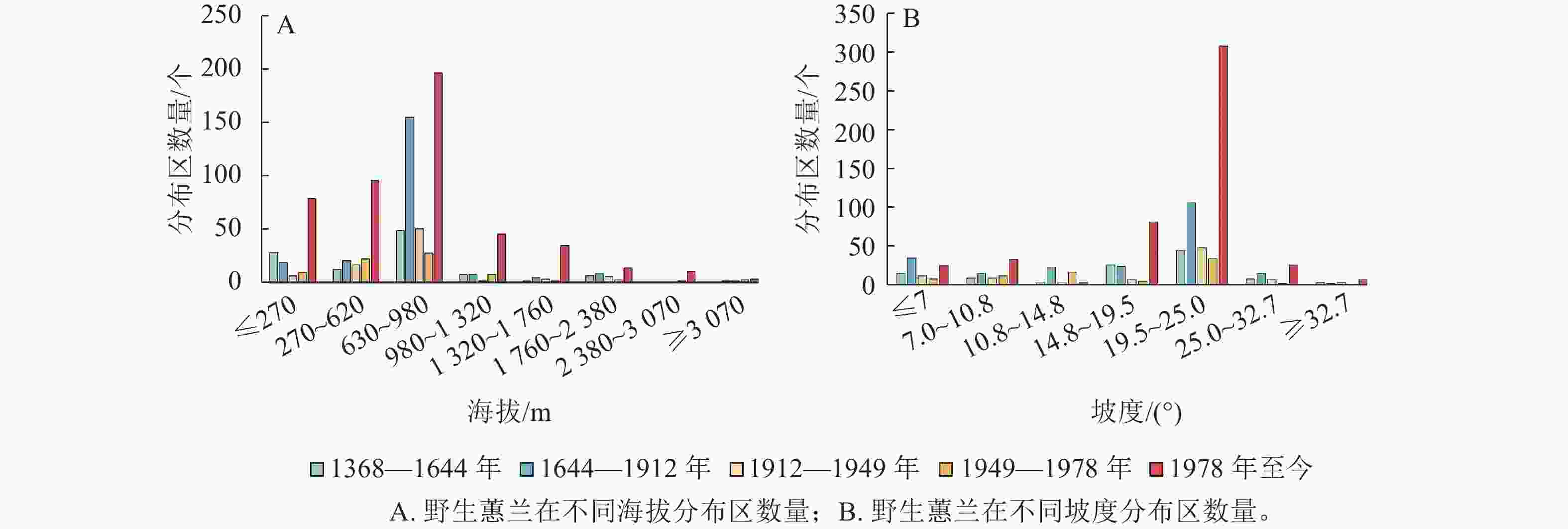
图 2 野生蕙兰在不同海拔和坡度下的分布区数量
Figure 2. Numbers of wild C. faberi distribution sites in the different altitudes and slopes
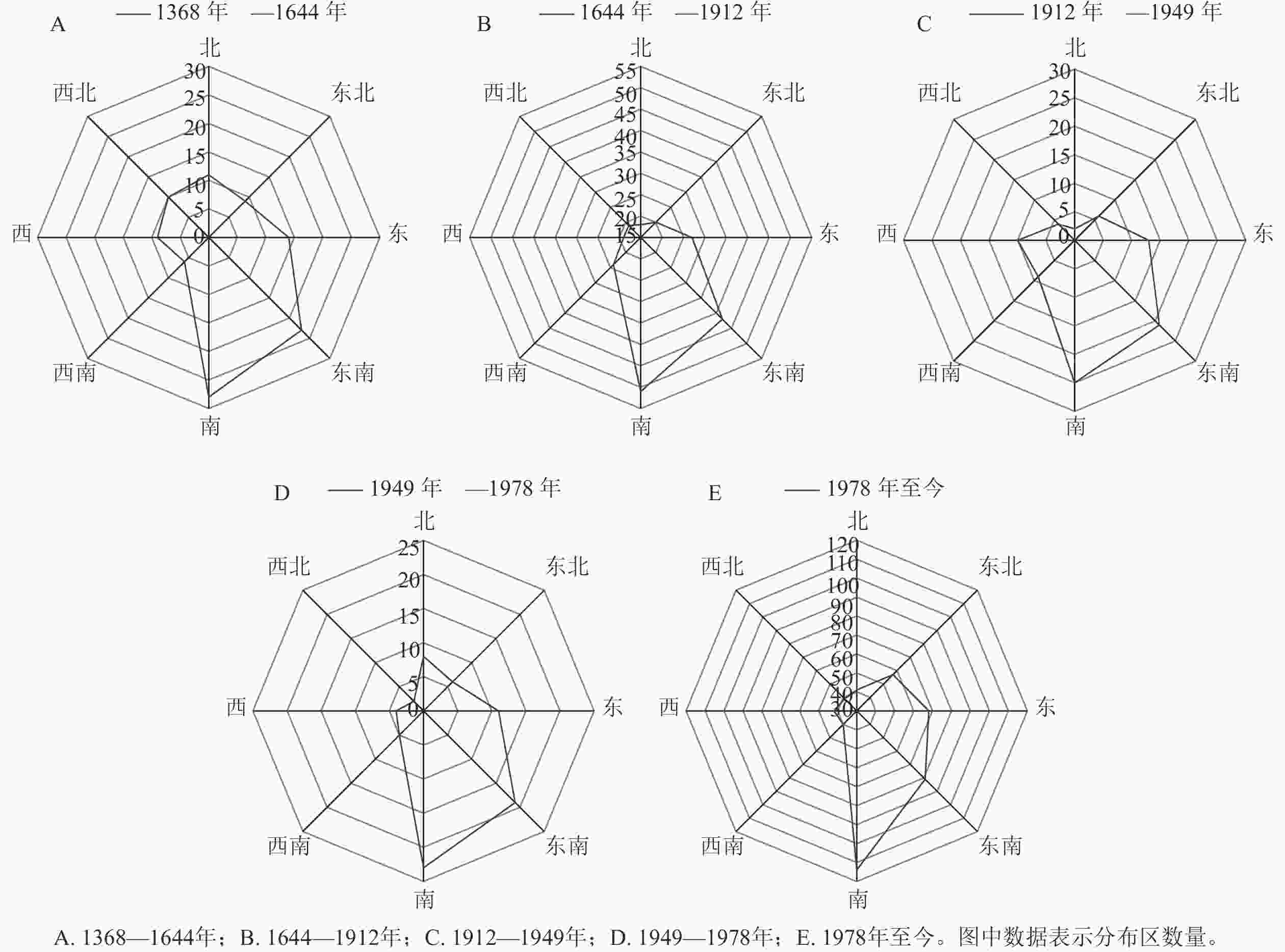
图 3 野生蕙兰在不同坡向的分布区数量
Figure 3. Numbers of wild C. faberi distribution sites in the different slope aspects
-
利用ArcGIS建立水系缓冲区,并结合野生蕙兰的分布情况(表4)进行分析可知:自1368年以来,野生蕙兰在距水系1 000~2 000 m的缓冲区分布相对集中。江南地区作为5个历史时期的野生蕙兰分布聚集点之一,北侧河网相对密集,野生蕙兰距离水源距离适中,分布相对集中。
表 4 野生蕙兰历史分布区域与水系缓冲区关系
Table 4. Relationship between the historical distribution sites of wild C. faberi and the water buffer area
离水距离/m 不同时期分布区数量/个 1368—1644年 1644—1912年 1912—1949年 1949—1978年 1978年至今 <500 18 42 10 7 92 500~1 000 21 32 18 14 103 1 000~2 000 44 87 24 29 153 >2 000 27 59 38 28 134 -
对1368年以来的野生蕙兰分布区数量与自然因素各因子以及离水距离数据建立二元Logistic回归模型,探究其影响因子(表5)。
表 5 各时期野生蕙兰分布模型
Table 5. Distribution model for wild C. faberi in various periods
自然因素 1644—1912年 自然因素 1912—1949年 βm 标准误 卡方值 显著性 exp(βm) βm 标准误 卡方值 显著性 exp(βm) 海拔 0.000 0.001 0.217 0.041 1.000 海拔 0.001 0.001 1.029 0.031 1.001 坡度 −0.038 0.075 0.263 0.018 0.962 坡度 0.297 0.144 1.226 0.040 1.346 坡向 0.006 0.002 2.914 0.009 1.006 坡向 0.001 0.002 0.084 0.007 1.001 土壤类型 0.014 0.010 1.978 0.061 1.015 土壤类型 −0.010 0.014 0.505 0.002 0.990 气温 0.001 0.001 0.531 0.006 1.001 气温 0.032 0.000 6.924 0.009 1.032 降水量 0.247 0.115 4.592 0.032 1.003 降水量 0.001 0.002 0.435 0.003 1.031 离水距离 0.000 0.001 0.217 0.081 1.000 离水距离 0.004 0.003 1.438 0.231 1.004 自然因素 1949—1978年 自然因素 1978年至今 βm 标准误 卡方值 显著性 exp(βm) βm 标准误 卡方值 显著性 exp(βm) 海拔 −0.001 0.001 0.999 0.018 0.999 海拔 0.001 0.001 2.962 0.045 1.001 坡度 0.261 0.198 0.741 0.187 1.298 坡度 −0.008 0.069 0.013 0.910 0.992 坡向 −0.004 0.004 0.672 0.035 0.996 坡向 0.004 0.003 2.158 0.042 1.004 土壤类型 0.014 0.019 0.564 0.003 1.014 土壤类型 −0.008 0.006 1.871 0.071 0.992 气温 0.038 0.241 0.024 0.046 1.038 气温 0.347 0.145 5.749 0.016 1.414 降水量 −0.001 0.002 0.435 0.003 0.999 降水量 −0.001 0.001 2.756 0.047 0.999 离水距离 0.000 0.000 0.715 0.013 1.000 离水距离 0.000 0.000 0.022 0.042 1.000 说明:βm为回归系数;exp(βm)为优势比。 7个自变量中,海拔、坡向、土壤类型、气温、降水量、离水距离均通过检验(P<0.05),表示野生蕙兰的分布区随以上自然因子的改变而存在差异。坡度等级在模型中的影响力较小,除1644—1912年之外,地势起伏变化对野生蕙兰的分布无显著影响。进一步分析模型中自变量的回归系数βm和优势比exp(βm)可知:降水量和气温是影响野生蕙兰分布变化的最大诱因。
-
1368—1912年,因“劝奖垦荒”和“改土归流”政策激励,贵州西部、四川南部、云南南部的山地边缘成为垦荒的重点区域。四川、云南、贵州人口的激增加大了对垦殖的刚性需求[29],导致四川、云南、贵州等地的森林资源被严重破坏,该区域野生蕙兰因生境破坏,分布锐减。1912—1949年,因垦殖破坏和工业布局西迁,四川、云南、贵州3省的森林覆盖率分别由40.0%、52.7%和21.0%下降至20.0%、40.0%和9.0%[30],野生蕙兰适生区进一步减少。1949—1978年,由于土地开垦范围的进一步扩大,使四川省和云南省的森林覆盖率分别减少了8.0%和13.0%[30],野生蕙兰的适生区面积进一步缩小。1978年至今,由于经济的发展和人们对生态环境保护的增强,四川、云南、贵州地区的森林资源逐渐恢复,至2020年,四川、云南、贵州森林覆盖率分别达40.0%、65.0%和60.0%,野生蕙兰的分布区数量增多。
-
前人的研究结果表明[31−32]:借助方志分析植物历史分布变迁是一种有效的手段,但受到古代交通条件和科学技术的限制,可能会存在少数记载盲区。1949年后,特别是1978年后中国经济和科技的高速发展,林史资料更加丰富,物种信息库更加完善,野生蕙兰的记载也更加科学,保障了数据采集的科学性。
-
气温和降水量对各时期野生蕙兰的分布均造成影响,但不同时期的影响程度不同。本研究发现:1368—1912年,气温无显著变化,但野生蕙兰的时空分布具有一定差异。1368—1644年,野生蕙兰在陕西、河南及华北诸地的分布极少,而1645—1912年野生蕙兰分布点在以上区域明显增多,并逐渐向华北中部迁移,这种时空分布差异可能是由降水量差异导致的。前人研究表明:最干月降水量会限制其分布。极端的干旱不仅会抑制兰科Orchidaceae植物根的活性,还会减少其开花的数量,降低传粉效率[8, 33]。1470—1644年整体干旱,特别是陕西、河南及华北诸地长期处于低降水状态,平均1.8 a发生1次旱灾[27, 34−36],而1644—1912年较为湿润,东亚季风增强,年际降水量增多,华北诸地雨泽无缺[37−38]。由于蕙兰适宜生长在湿润的地区[19],因此降水量的增多能够进一步促进其生长繁殖。此外,本研究发现:1368—1912年,野生蕙兰的分布均呈现出“东南多而西北少”的分布格局,中国东南地区降水多,西北地区降水少,更加佐证了降水量影响了1368—1912年野生蕙兰分布的结论。前人研究表明:全球变暖对野生蕙兰适生区的扩张具有一定的积极作用[8]。本研究同样发现:气温升高促进了野生蕙兰分布区数量的增多,并且是导致1912年后野生蕙兰分布变化的主要原因。1912—1949年整体湿润,中国西南地区和华南地区平均气温升高,云南平均气温较1644—1912年上升了约0.6 ℃,该地区野生蕙兰分布区数量增多;而四川、陕西和安徽等地的平均气温较1644—1912年分别下降了约0.4、0.3和0.8 ℃,使这些地区野生蕙兰分布区数量减少。1978年至今,华北、西南、湖南和湖北的平均气温持续升高,陕西、云南、贵州、湖北等的气温相较于1978年前上升,这些区域野生蕙兰分布区明显增加,并呈多点聚集的趋势;陕西、湖北等地成为新的野生蕙兰聚集区。野生蕙兰的分布中心逐渐向高纬度地区迁移,进一步说明气温能直接影响野生蕙兰的分布。
前人研究发现:兰科植物在贵州、广西交界山区,湖北西部和重庆东部山地,秦岭—伏牛山一带最为丰富,地形地貌和土壤类型是造成其在小区域分布差异的重要原因[39−41]。本研究发现:野生蕙兰在小尺度区域上的分布同样受到地形地貌和土壤类型的影响,如四川、云南、贵州地理位置相近,但野生蕙兰分布数量却不同。四川省地形地貌复杂,海拔较低;云南省海拔整体较高,地貌丰富,土壤类型多以微酸性的红壤为主;贵州省西高东低,土壤类型以红壤、黄壤为主。另外,本研究发现:野生蕙兰适宜生长在pH 5.3~6.2的温热铁铝土区,及海拔620~980 m的南坡或东南坡,云南、贵州两地合适的土壤类型和地形地貌与野生蕙兰的生态习性相耦合,为其生长繁育提供有利条件,因此云南、贵州两省的野生蕙兰资源相比于四川省更加集中。
-
前人研究表明:过度砍伐森林和采集销售都有可能成为兰科植物分布变化和数量减少的原因[42]。本研究同样发现:人为导致的生境恶化以及过度采集是野生蕙兰自然数量减少的原因之一。首先,以西南地区为例,1363—1978年,该地区经历了过度的森林砍伐、农业垦殖和工业布局西迁,导致野生蕙兰的生境被严重破坏,分布区数量减少。1978年至今,随着经济的发展和生态环境保护力度的加强,西南地区的天然林面积增加了58%[43],野生蕙兰的适宜生境增加,分布区数量增多。其次,以南岭地区为例,集市对野生蕙兰的采集销售加剧了该地区的人为采挖,导致该地区野生蕙兰的生存环境恶化[42]。21世纪后,互联网的发展为野生蕙兰的售卖提供了更加便捷的平台,使其生存数量减少[44]。
-
为了对野生蕙兰及其遗传多样性进行有效保育,应有针对性地对其分布集中且适宜生长的区域进行重点保护。结合本研究结果来看,仅有28.4%的野生蕙兰分布区在现有的保护区范围内,且仅有广西雅长国家级自然保护区是以兰科植物为保护对象的保护区。应进一步加强对江南、广东、福建、云南、贵州、陕西、湖北等典型区域的野生蕙兰适生区营建或扩建,及时追踪以上区域内野生蕙兰种群的变化动态;同时,随着全球气候的变暖,野生蕙兰逐渐向纬度更高的区域迁移,可以加强对华北地区,特别是该地区年平均气温为15~25 ℃,海拔为620~980 m、坡度为19.9°~25.0°的南坡或东南坡野生蕙兰的资源调查及监测,及时制定长期保护规划,调整其保护区范围。同时,应提高公众的保护意识,倡导利用当地适宜的气候条件建立可持续的兰科植物栽培企业。
Spatiotemporal distribution and influencing factors of wild Cymbidium faberi from the perspective of historical geographic information system
-
摘要:
目的 明晰野生蕙兰Cymbidium faberi的分布与演变,有助于探究中国蕙兰自然种群的演替规律,为当代蕙兰自然种群的保育工作提供科学依据。 方法 基于ArcGIS平台,构建野生蕙兰历史地理信息数据库,对1368年以来野生蕙兰的时空分布及其影响因素进行研究。 结果 ①自1368年以来,野生蕙兰主要分布于中国秦岭—淮河以南的区域,分布中心由28.585°N,113.503°E逐渐向29.365°N,112.675°E迁移。1368—1644年主要聚集于江南、广东、福建和四川、云南交界处。1644—1912年在四川、云南一带聚集程度减弱; 1949—1978年四川、重庆地区聚集程度增强; 1978年至今,野生蕙兰呈多点聚集,湖北、陕西等地成为新的聚集区;②气温、降水量、土壤类型、地形地貌、水源缓冲距离等自然因素直接作用于野生蕙兰的分布,其主要分布于年平均气温为15~25 ℃,pH为5.3~6.2的温热铁铝土区及海拔为620~980 m、坡度为19.9°~25.0°的南坡或东南坡,且离水距离为1 000~2 000 m内的区域。③农业垦殖以及工业发展等人为活动间接影响野生蕙兰的分布与迁移。 结论 野生蕙兰主要聚集于中国南方地区,并趋于向高纬度地区迁移,其分布受气温和降水量影响显著,人为活动间接导致了其分布数量的减少,应对江南、广东、福建、云南、贵州、陕西、湖北等典型区域的野生蕙兰适生区进行营建或扩建,以加强对野生蕙兰的保护。图3表5参44 Abstract:Objective This study aims to clarify the distribution and evolution of wild Cymbidium faberi and explore the succession law of its natural population in China, so as to provide scientific basis for the conservation of its natural population. Method Based on the ArcGIS platform, a historical geographic information database of wild C. faberi was constructed to analyze its spatiotemporal distribution and influencing factors since 1368. Result (1) Since 1368, wild C. faberi has been mainly distributed in the south of Qinling Mountains-Huaihe River in China, with the distribution center gradually migrating from 28.585°N and 113.503°E to 29.365°N and 112.675°E. From 1368 to 1644, it was mainly concentrated in Jiangnan, Guangdong, Fujian, Jiangxi, Sichuan and Yunnan regions. From 1644 to 1912, the degree of aggregation in Sichuan and Yunnan weakened. From 1949 to 1978, the degree of aggregation in Sichuan and Chongqing regions increased. Since 1978, wild C. faberi has gathered at multiple sites, with Hubei and Shaanxi becoming new gathering areas. (2) Natural factors such as temperature, precipitation, soil type, topography and distance from water sources directly affected the distribution of wild C. faberi. It was mainly distributed in the warm bauxite areas with an annual average temperature of 15−25 ℃ and pH 5.3−6.2, as well as on the southern or southeastern slopes with an altitude of 620−980 m and a slope of 19.9°−25.0°, and within a distance of 1 000−2 000 m from water sources. (3) The distribution and migration of wild C. faberi was indirectly affected by activities such as agricultural cultivation and industrial development. Conclusion Wild C. faberi mainly gathers in southern China and tends to migrate to higher latitude areas. Its distribution is significantly affected by temperature and precipitation, and human activities indirectly lead to a decrease in its distribution. Therefore, it is necessary to build or expand suitable areas of wild C. faberi in typical areas such as Jiangnan, Guangdong, Fujian, Jiangxi, Yunnan, Guizhou, Shaanxi and Hubei to strengthen the protection. [Ch, 3 fig. 5 tab. 44 ref.] -
图 2 野生蕙兰在不同海拔和坡度下的分布区数量
Figure 2 Numbers of wild C. faberi distribution sites in the different altitudes and slopes
图 3 野生蕙兰在不同坡向的分布区数量
Figure 3 Numbers of wild C. faberi distribution sites in the different slope aspects
表 1 野生蕙兰历史分布模型自变量
Table 1. Historical distribution model independent variables of C. faberi
自然因子 描述 数据来源及处理工具 自然因子 描述 数据来源及处理工具 海拔 地形分析因子 DEM数据 气温 年均气温 BIO1及CRU TStemp数据集 坡度 地表起伏形态 坡度工具 降水量 年均降水量 BIO12及CRU TSpre数据集 坡向 坡向 坡向工具 水源距离 分布点至水系的距离 河网数据 土壤类型 分布区域的土壤类型 土壤数据  下载: 导出CSV
下载: 导出CSV
表 2 1368年以来野生蕙兰资源分布标准差椭圆分析
Table 2. Analysis for the standard deviational ellipse of the distribution sites of wild C. faberi resources since the year of 1368
年份 椭圆周长/km 椭圆面积/km² 椭圆中心点(N) 椭圆中心点(E) 长半轴/km 短半轴/km 椭圆的方向角度/(°) 1368—1644 72.088 167 208.792 730 28.584 810° 113.503 226° 16.941 399 3.924 427 91.145 131 1644—1912 41.965 962 120.646 298 29.519 570° 116.191 821° 8.548 608 4.492 692 79.365 494 1912—1949 46.740 223 112.516 143 28.872 057° 114.902 188° 10.549 990 3.395 456 80.062 387 1949—1978 42.521 332 116.530 018 29.564 576° 112.446 761° 8.944 940 4.147 208 88.409 640 1978年至今 39.360 687 105.286 125 29.364 727° 112.674 798° 8.056 393 4.160 246 78.474 936
下载: 导出CSV
表 3 不同时期气候因子与野生蕙兰分布情况
Table 3. Distribution sites of wild C. faberi under the effect of climatic factors in various periods
分布区 1644—1912年 1912—1949年 气温/℃ 降水量/mm 分布区数量/个 气温/℃ 降水量/mm 分布区数量/个 江苏 13.3~15.5 850.0~983.6 34 13.0~16.2 807.0~901.0 9 浙江 13.6~16.7 750.0~983.6 62 13.6~19.0 901.8~980.0 31 安徽 13.7~16.1 750.0~980.2 19 12.9~15.3 807.7~980.3 7 上海 14.9~15.8 810.0~820.0 5 11.2~13.4 807.4~901.8 1 福建 15.3~21.0 800.0~850.0 21 13.0~19.8 901.8~980.4 4 广东 17.8~23.2 750.0~900.0 8 16.2~22.8 901.8~980.4 5 广西 16.3~22.8 600.0~897.0 3 16.2~22.8 567.7~925.6 1 云南 7.1~23.2 295.0~950.0 10 7.8~24.8 308.2~980.4 15 四川 −1.6~18.3 51.0~450.0 9 −0.8~18.5 80.3~901.8 6 贵州 14.6~18.1 413.0~802.0 7 14.8~18.4 697.4~920.6 2 湖南 15.2~17.4 600.0~950.0 7 15.4~17.3 901.8~980.4 2 湖北 11.3~16.2 250.0~983.6 5 10.4~16.3 697.4~980.4 2 江西 15.3~19.1 750.0~930.0 6 15.2~18.9 901.8~980.4 2 重庆 12.1~18.2 250.0~700.0 0 16.2~19.6 807.5~901.8 0 陕西 6.3~13.3 169.0~450.0 5 5.7~13.0 567.7~901.8 1 甘肃 −4.8~11.8 51.0~169.0 3 −4.2~9.4 80.2~567.8 0 台湾 11.1~20.3 897.0~983.0 2 9.4~19.6 901.8~980.4 0 山东 10.4~14.1 450.0~950.0 2 5.7~13.0 807.4~901.8 0 河南 11.1~15.3 350.0.0~897.0 4 9.4~16.2 697.4~901.8 1 河北 2.6~13.7 543.0~820.0 2 2.4~13.0 807.5~890.5 0 山西 4.4~12.5 100.0~450.0 1 2.4~9.4 697.4~901.8 0 北京 1.3~11.4 450.0~700.0 2 2.4~13.0 901.8~980.4 0 吉林 0.3~6.2 680.0~950.0 1 −0.8~5.7 807.5~980.4 0 香港 21.8~24.3 802.0~897.0 2 16.2~19.6 901.8~980.4 0 西藏 −4.1~19.5 51.0~450.0 0 −9.4~19.4 50.0~550.0 0 海南 19.1~24.3 295.7~413.6 0 19.6~24.8 450.0~650.0 1
下载: 导出CSV
3 (续)
3. Continued
分布区 1949—1978年 1978年至今 气温/℃ 降水量/mm 分布区数量/个 气温/℃ 降水量/mm 分布区数量/个 江苏 13.0~16.0 687.5~1 294.5 2 14.1~16.7 982.5~1 320.0 15 浙江 13.0~19.3 1 294.5~1 670.0 16 14.6~18.3 1 294.5~1 701.0 71 安徽 13.0~16.0 687.5~1 572.0 5 14.1~18.3 690.2~1 639.2 39 上海 15.9~17.2 982.3~1 294.5 0 15.9~16.1 983.5~1 310.4 1 福建 16.0~20.3 982.3~1 572.1 6 15.7~21.4 1 024.2~1 653.0 11 广东 16.0~24.5 1 294.5~2 070.9 2 18.3~24.7 1 350.0~2 600.0 21 广西 16.3~24.5 982.3~1 971.0 2 18.3~24.6 723.9~2 983.8 12 云南 1.84~24.5 687.5~1 572.1 3 2.8~24.6 781.8~2 118.2 17 四川 −1.5~19.3 444.6~1 294.5 13 −3.4~18.9 601.2~1 294.5 14 贵州 9.1~19.4 687.5~1 082.3 3 9.9~18.4 687.5~1 449.7 84 湖南 13.0~19.3 982.3~1 374.2 0 14.2~20.7 1 150.0~2 100.0 25 湖北 9.1~16.7 687.5~1 380.6 1 9.9~18.4 860.5~2 100.0 84 江西 16.0~20.3 1 294.5~1 970.9 5 15.6~19.8 1 143.2~1 935.0 19 重庆 13.0~19.3 982.3~1 572.1 5 14.2~19.4 1 000.0~1 572.1 8 陕西 6.2~13.0 444.6~982.3 5 7.1~14.2 460.2~904.5 15 甘肃 −4.8~9.1 187.8~687.5 1 −6.5~9.9 136.6~734.9 6 台湾 9.1~20.6 1 294.5~2 800.5 0 9.4~23.5 1 294.5~2 760.0 2 山东 9.1~16.0 687.5~982.3 0 9.5~16.5 565.5~982.3 7 河南 13.0~16.3 450.6~1 094.5 8 13.5~17.3 500.4~1 100.5 22 河北 −1.5~14.1 444.6~687.5 0 −0.3~14.2 503.4~687.5 1 山西 1.8~10.9 444.6~687.5 0 2.8~14.2 567.2~864.9 2 北京 5.3~13.0 444.6~687.5 1 6.1~14.2 374.6~687.5 3 吉林 −1.5~9.1 350.8~982.3 0 −0.3~9.9 380.5~703.2 0 香港 16.0~24.5 1 294.5~2 070.9 0 18.3~24.7 1 350.0~2 600.0 1 西藏 −10.5~19.3 85.0~687.5 0 −6.5~23.5 76.2~700.2 1 海南 19.3~24.5 1 572.0~1970.0 0 18.3~27.1 1 320.0~2 600.0 1
下载: 导出CSV
表 4 野生蕙兰历史分布区域与水系缓冲区关系
Table 4. Relationship between the historical distribution sites of wild C. faberi and the water buffer area
离水距离/m 不同时期分布区数量/个 1368—1644年 1644—1912年 1912—1949年 1949—1978年 1978年至今 <500 18 42 10 7 92 500~1 000 21 32 18 14 103 1 000~2 000 44 87 24 29 153 >2 000 27 59 38 28 134
下载: 导出CSV
表 5 各时期野生蕙兰分布模型
Table 5. Distribution model for wild C. faberi in various periods
自然因素 1644—1912年 自然因素 1912—1949年 βm 标准误 卡方值 显著性 exp(βm) βm 标准误 卡方值 显著性 exp(βm) 海拔 0.000 0.001 0.217 0.041 1.000 海拔 0.001 0.001 1.029 0.031 1.001 坡度 −0.038 0.075 0.263 0.018 0.962 坡度 0.297 0.144 1.226 0.040 1.346 坡向 0.006 0.002 2.914 0.009 1.006 坡向 0.001 0.002 0.084 0.007 1.001 土壤类型 0.014 0.010 1.978 0.061 1.015 土壤类型 −0.010 0.014 0.505 0.002 0.990 气温 0.001 0.001 0.531 0.006 1.001 气温 0.032 0.000 6.924 0.009 1.032 降水量 0.247 0.115 4.592 0.032 1.003 降水量 0.001 0.002 0.435 0.003 1.031 离水距离 0.000 0.001 0.217 0.081 1.000 离水距离 0.004 0.003 1.438 0.231 1.004 自然因素 1949—1978年 自然因素 1978年至今 βm 标准误 卡方值 显著性 exp(βm) βm 标准误 卡方值 显著性 exp(βm) 海拔 −0.001 0.001 0.999 0.018 0.999 海拔 0.001 0.001 2.962 0.045 1.001 坡度 0.261 0.198 0.741 0.187 1.298 坡度 −0.008 0.069 0.013 0.910 0.992 坡向 −0.004 0.004 0.672 0.035 0.996 坡向 0.004 0.003 2.158 0.042 1.004 土壤类型 0.014 0.019 0.564 0.003 1.014 土壤类型 −0.008 0.006 1.871 0.071 0.992 气温 0.038 0.241 0.024 0.046 1.038 气温 0.347 0.145 5.749 0.016 1.414 降水量 −0.001 0.002 0.435 0.003 0.999 降水量 −0.001 0.001 2.756 0.047 0.999 离水距离 0.000 0.000 0.715 0.013 1.000 离水距离 0.000 0.000 0.022 0.042 1.000 说明:βm为回归系数;exp(βm)为优势比。
下载: 导出CSV
-
[1] YU Yun, CUI Yahua, HSIANG T, et al. Isolation and identification of endophytes from roots of Cymbidium goeringii and Cymbidium faberi (Orchidaceae) [J]. Nova Hedwigia, 2015, 101(1/2): 57 − 64. [2] 蒋楚楚, 辛静静, 夏树全, 等. 蕙兰‘红香妃’内生真菌分离鉴定及体外抑菌效应[J]. 浙江农林大学学报, 2023, 40(4): 783 − 791. JIANG Chuchu, XIN Jingjing, XIA Shuquan, et al. Isolation and identification of endophytic fungi from Cymbidium faberi ‘Hongxiangfei’ and their bacteriostatic effect in vitro [J]. Journal of Zhejiang A&F University, 2023, 40(4): 783 − 791. [3] 袁媛, 曹彬, 张咏琪, 等. 基于SRAP标记的国兰种质资源遗传多样性分析[J]. 热带作物学报, 2020, 41(5): 929 − 938. YUAN Yuan, CAO Bin, ZHANG Yongqi, et al. Genetic diversity analysis of Cymbidium germplasms based on SRAP markers [J]. Chinese Journal of Tropical Crops, 2020, 41(5): 929 − 938. [4] LIANG Hongyan, WANG Xiaoguo, CHEN Wei, et al. Spatial genetic structure of terrestrial orchid Cymbidium faberi in the Qinling Mountains revealed by microsatellite loci [J/OL]. Plant Systematics and Evolution, 2021, 307(1): 5[2023-01-13]. doi: 10.1007/s00606-020-01735-y. [5] FEI Yue, LIU Zhixiong. Isolation and characterization of the PISTILLATA ortholog gene from Cymbidium faberi Rolfe [J/OL]. Agronomy, 2019, 9(8): 425[2023-01-13]. doi: 10.3390/agronomy9080425. [6] XU Yanqin, ZHOU Junjiang, LIU Qingqing, et al. Construction and characterization of a high-quality cDNA library of Cymbidium faberi suitable for yeast one- and two-hybrid assays [J/OL]. BMC Biotechology, 2020, 20(1): 4[2-23-01-13]. doi: 10.1186/s12896-020-0599-2. [7] 徐婉, 林雅君, 赵荘, 等. 兰属植物资源与育种研究进展[J]. 园艺学报, 2022, 49(12): 2722 − 2742. XU Wan, LIN Yajun, ZHAO Zhuang, et al. Advances in genetic resources and breeding research of Cymbidium [J]. Acta Horticulturae Sinica, 2022, 49(12): 2722 − 2742. [8] 梁红艳, 姜效雷, 孔玉华, 等. 气候变暖背景下春兰和蕙兰的适生区分布预测[J]. 生态学报, 2018, 38(23): 8345 − 8353. LIANG Hongyan, JIANG Xiaolei, KONG Yuhua, et al. Prediction of the potential geographical distribution of Cymbidium goeringii and C. faberi under the background of global warming [J]. Acta Ecologica Sinica, 2018, 38(23): 8345 − 8353. [9] 费越, 黄承石, 贺心茹, 等. 湖北京山县野生蕙兰群落结构与种子植物区系分析[J]. 云南农业大学学报(自然科学), 2019, 34(6): 1055 − 1060. FEI Yue, HUANG Chengshi, HE Xinru, et al. Study on the community structure of wild Cymbidium faberi and the spermatophyte flora in Jingshan County, Hubei [J]. Journal of Yunnan Agricultural University (Natural Science), 2019, 34(6): 1055 − 1060. [10] 杨立. 基于物种历史分布数据与生态位模型的濒危物种分布模拟[D]. 北京: 北京林业大学, 2018. YANG Li. Reconstructing Distribution Change of Endangered Species Based on Species Historical Distribution Data and Ecological Niche Modeling [D]. Beijing: Beijing Forestry University, 2018. [11] 王志威. 药用植物云南土沉香潜在适生区及其关键气候影响因子[J]. 生态学杂志, 2022, 41(10): 1991 − 1997. WANG Zhiwei. The potential suitable areas and key climatic impacting factors of medicinal plant Excoecaria acerifolia Didr [J]. Chinese Journal of Ecology, 2022, 41(10): 1991 − 1997. [12] XIE Chunping, HUANG Boyang, JIM C Y, et al. Predicting suitable habitat for the endangered plant Cephalotaxus oliveri Mast. in China [J]. Environmental Conservation, 2022, 50(1): 50 − 57. [13] 王鑫, 任亦钊, 黄琴, 等. 基于GIS和Maxent模型的赤水河地区濒危植物桫椤生境适宜性评价[J]. 生态学报, 2021, 41(15): 6123 − 6133. WANG Xin, REN Yizhao, HUANG Qin, et al. Habitat suitability assessment of endangered plant Alsophila spinulosa in Chishui river area based on GIS and Maxent model [J]. Acta Ecologica Sinica, 2021, 41(15): 6123 − 6133. [14] TANG Shuangli, SONG Yaobin, ZENG Bo, et al. Potential distribution of the extremely endangered species Ostrya rehderiana (Betulaceae) in China under future climate change [J]. Environmental Science and Pollution Research, 2022, 29(5): 7782 − 7792. [15] 唐丽雅, 韩凯, 马明志, 等. 农作物传播研究: 以陕西汉中龙岗寺遗址新石器时代炭化植物遗存为例[J]. 第四纪研究, 2020, 40(2): 512 − 524. TANG Liya, HAN Kai, MA Mingzhi, et al. The dispersals of erops: s study on the remains of catonize plants in the Neolihic age at Longgangsisite in Hanzhong, Shaanxi Province [J]. Quaternary Sciences, 2020, 40(2): 512 − 524. [16] BIPPUS A C, FLORES J R, HYVONEN J, et al. The role of paleontological data in bryophyte systematics [J]. Journal of Experimental Botany, 2022, 73(13): 4273 − 4290. [17] 刘爽, 惠富平. 明清时期苜蓿的地域分布及其影响因素[J]. 草业学报, 2021, 30(2): 178 − 189. LIU Shuang, HUI Fuping. Distribution of alfalfa in the Ming and Qing Dynasties and the underlying driving factors [J]. Acta Prataculturae Sinica, 2021, 30(2): 178 − 189. [18] FENG Gang, MAO Lingfeng, BENITO B M, et al. Historical anthropogenic footprints in the distribution of threatened plants in China [J/OL]. Biological Conservation, 2017, 210[2023-01-13]. doi: 10.1016/j.biocon.2016.05.038. [19] 赵令妹. 中国养兰集成[M]. 北京: 中国林业出版社, 2007: 30, 383−385. ZHAO Lingmei. Chinese Orchid Raising Integration [M]. Beijing: China Forestry Publishing House, 2007: 30, 383 − 385. [20] 竺可桢. 中国近五千年来气候变迁的初步研究[J]. 考古学报, 1972(1): 15 − 38. ZHU Kezhen. Climatic change in China over the past 5000 years [J]. Acta Archaeologica Sinica, 1972(1): 15 − 38. [21] 张德二. 中国三千年气象记录总集[M]. 南京: 凤凰出版社, 2004: 1643 − 2297. ZHANG De’er. A Compendium of Chinese Meteorological Records of the Last 3000 Years [M]. Nanjing: Phoenix Publishing House, 2004: 1643 − 2297. [22] 文焕然. 历史时期中国气候变化[M]. 济南: 山东科学技术出版社, 2019: 243 − 283. WEN Huanran. Climate Changes in China in Historical Periods [M]. Ji’nan: Technology Press, 2019: 243 − 283. [23] 白雪锋, 许浩, 周燚. 基于路网中心性的近代南京城市寺庙选址特征[J]. 中国园林, 2023, 39(4): 71 − 76. BAI Xuefeng, XU Hao, ZHOU Yi. Characteristics of temple location in modern Nanjing based on the centrality of road network [J]. Chinese Landscape Architecture, 2023, 39(4): 71 − 76. [24] 白雪锋, 许浩. 近代南京钟山风景区历史景观资源时空分布与演变研究[J]. 中国园林, 2022, 38(7): 139 − 144. BAI Xuefeng, XU Hao. Study on temporal and spatial distribution and evolution of historical landscape resources in Zhongshan Scenic Area in modern Nanjing [J]. Chinese Landscape Architecture, 2022, 38(7): 139 − 144. [25] ZHANG Xinghang, ZHANG Baiping, YAO Yonghui, et al. Dynamics and climatic drivers of evergreen vegetation in the Qinling-Daba mountains of China [J/OL]. Ecological Indicators, 2022, 136: 108625[2023-01-13]. doi: 10.1016/j.ecolind.2022.108625. [26] CUI Jianxin, CHANG Hong, BURR G S, et al. Climatic change and the rise of the Manchu from northeast China during AD 1600−1650 [J]. Climatic Change, 2019, 156(3): 405 − 423. [27] 葛全胜. 中国历朝气候变化[M]. 北京: 科学出版社, 2010: 494 − 523. GE Quansheng. Climate Change in Chinese Dynasties [M]. Beijing: Science Press, 2010: 494 − 523. [28] XIAO Lingbo, FANG Xiuqi, ZHENG Jingyun et al. Famine, migration and war: comparison of climate change impacts and social responses in North China between the late Ming and late Qing dynasties [J]. Holocene, 2015, 25(6): 900 − 910. [29] WU J. Population growth in the Jiangnan area in the Qing Dynasty: an analysis [J]. Chinese Journal of Population Science, 1989, 1(2): 235 − 249. [30] 徐波. 近400年来中国西部社会变迁与生态环境[M]. 北京: 中国社会科学出版社, 2014: 280, 410 − 420. XU Bo. Social Changes and Ecological Environment in Western China in the Past 400 Years [M]. Beijing: China Social Sciences Press, 2014: 280, 410 − 420. [31] 李娜, 包平. 基于数字人文的明至民国山西物产分布研究[J]. 中国科技史杂志, 2022, 43(2): 204 − 217. LI Na, BAO Ping. The Chinese journal for the history of science and technology [J]. The Chinese Journal for the History of Science, 2022, 43(2): 204 − 217. [32] 刘京晶, 斯金平. 黄精本草考证与启迪[J]. 中国中药杂志, 2018, 43(3): 631 − 636. LIU Jingjing, SI Jinping. Herbal textual research on Chinese medicine Huangjing (Polygonati rhizoma) and some enlightenments [J]. China Journal of Chinese Materia Medica, 2018, 43(3): 631 − 636. [33] LIU Hong, FENG Changlin, CHEN Baoshan, et al. Overcoming extreme weather challenges: successful but variable assisted colonization of wild orchids in southwestern China [J]. Biological Conservation, 2012, 150(1): 68 − 75. [34] CHEN Feng, MARTIN H, ZHAO Xiaoen, et al. Abnormally low precipitation-induced ecological imbalance contributed to the fall of the Ming Dynasty: new evidence from tree rings [J/OL]. Climatic Change, 2022, 173(1): 13[2023-01-13]. doi: 10.1007/s10584-022-03406-y. [35] HAN Jianfu, YANG Yuda. The socioeconomic effects of extreme drought events in northern China on the Ming dynasty in the late fifteenth century [J/OL]. Climatic Change, 2021, 164[2023-01-11]. doi: 10.1007/s10584-021-02972-x. [36] BI Shouben, BI Shengjie, CHEN Changchun et al. Analysis of the spatiotemporal characteristics of drought disasters in north China during the Ming and Qing dynasties [J]. Geomatics,Natural Hazards and Risk, 2020, 11(1): 2509 − 2539. [37] 孟万忠. 近六百年华北地区霜雪灾害与寒冷气候事件研究[M]. 北京: 气象出版社, 2022: 375 − 380. MENG Wanzhong. Study of Frost and Snow Disasters and Cold Weather Events in North China in the Last 600 Years [M]. Beijing: China Meteorological Press, 2022: 375 − 380. [38] HAO Zhixin, YU Yingzhuo, GE Quansheng, et al. Reconstruction of high-resolution climate data over China from rainfall and snowfall records in the Qing Dynasty [J/OL]. Wiley Interdisciplinary Reviews: Climate Change, 2018, 9(3): e517[2023-01-13]. doi: 10.1002/wcc.517. [39] 周康, 张哲, 宋希强, 等. 海南主要陆域自然保护地兰科植物多样性与生境的关联分析[J]. 广西植物, 2022, 42(8): 1337 − 1356. ZHOU Kang, ZHANG Zhe, SONG Xiqiang, et al. Association analysis of orchid diversity and habitat in main land nature reserves in Hainan [J]. Guihaia, 2022, 42(8): 1337 − 1356. [40] 刘飞虎, 黄浪, 刘环, 等. 罗霄山脉兰科植物区系及其生态地理学特征[J]. 植物科学学报, 2020, 38(4): 467 − 475. LIU Feihu, HUANG Lang, LIU Huan, et al. Flora of Orchidaceae in Luoxiao Mountains and its ecogeographical characteristics [J]. Plant Science Journal, 2020, 38(4): 467 − 475. [41] 张殷波, 杜昊东, 金效华, 等. 中国野生兰科植物物种多样性与地理分布[J]. 科学通报, 2015, 60(2): 179 − 188. ZHANG Yinbo, DU Haodong, JIN Xiaohua, et al. Species diversity and geographic distribution of wild Orchidaceae in China [J]. Chinese Science Bulletin, 2015, 60(2): 179 − 188. [42] 张一林, 李功权, 刘颖. 长江流域濒危兰科植物的空间分布[J]. 浙江农林大学学报, 2022, 39(4): 750 − 757. ZHANG Yilin, LI Gongquan, LIU Ying. Spatial distribution of endangered orchids in the Yangtze River Watershed [J]. Journal of Zhejiang A&F University, 2022, 39(4): 750 − 757. [43] 陈定超, 李卫朋, 蔡晓雨, 等. 四川省森林资源动态变化及其影响因素分析[J]. 地球环境学报, 2021, 12(4): 425 − 435. CHEN Dingchao, LI Weipeng, CAI Xiaoyu, et al. Analysis on the dynamic changes and their influencing factors of forest resources in Sichuan Province [J]. Journal of Earth Environment, 2021, 12(4): 425 − 435. [44] 黄婧, 孙美, 余文峰, 等. 我国网络平台本土兰科植物贸易状况[J]. 生物多样性, 2022, 30(4): 78 − 87. HUANG Jing, SUN Mei, YU Wenfeng, et al. Trade of native orchids on Chinese online platforms [J]. Biodiversity Science, 2022, 30(4): 78 − 87. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220766
 点击查看大图
点击查看大图
计量
- 文章访问数: 2256
- HTML全文浏览量: 638
- PDF下载量: 29
- 被引次数: 0
