





-
陆地生态系统碳循环是全球碳循环的重要部分,土壤是陆地生态系统最大的碳库。有研究表明:陆地生态系统中土壤有机碳储量比大气中高2~3倍[1]。土壤有机碳是土壤碳库的重要存在形式,也是维持陆地生态系统结构和功能的重要基础[2]。但是由于自然因素或者人为破坏造成的陆地生态系统退化已经严重影响了土壤碳库的稳定和固持能力,甚至成为仅次于化石燃料燃烧的人为二氧化碳排放源[3]。植被恢复普遍被认为是提升退化土壤有机碳固持能力的有效措施[4],然而土壤碳库恢复往往滞后于植被,且不同植被恢复模式之间差异较大[5]。因此,研究不同植被恢复模式下土壤有机碳储量特征对正确认识退化区域土壤固碳潜力和可持续恢复具有指导意义。
在植被恢复过程中土壤有机碳的积累存在很大的不确定性,与造林前相比,植被恢复后土壤有机碳增加、减少或者无显著变化的报道均有存在[6]。这主要是由于造林打破了原生态系统土壤有机碳输入与输出的动态平衡,可能需要几十年的时间才能达到新的平衡,因此土壤有机碳主要取决于造林前储存碳的分解[7]。此外,人工造林与植被自然恢复作为退化生态系统非常重要的植被恢复模式,两者的土壤固碳能力大小也存在争议。WEI等[5]研究表明:人工造林的土壤固碳能力显著高于自然植被恢复,而JIN等[8]认为:自然植被恢复比人工造林更利于土壤固碳。这是因为2种植被恢复模式在经营管理、物种组成、微气候、根系周转及分泌物、凋落物等方面都存在很大差异[9],从而导致土壤碳汇效应差异,因此,分析不同人工恢复模式与自然恢复模式下的土壤碳汇效应对探讨其在调节碳平衡中的作用和贡献具有重要意义。
水利部监测结果表明:南方红壤丘陵考察区共有水土流失面积13.12万km2,约占土地总面积的15%,已经成为仅次于黄土高原的第二大土壤侵蚀区[10]。目前,已有学者对南方红壤侵蚀区的土壤碳库特征做了初步研究,如李燕燕等[11]研究了红壤丘陵区不同植被恢复模式对土壤有机碳周转的影响,结果表明:马尾松Pinus massoniana林土壤有机碳来源于森林碳的比例高于木荷Schima superba林,但木荷林中土壤有机碳分解更慢;XIAO等[12]分析了赣南退化红壤区不同植被恢复年限土壤有机碳密度的动态变化特征,发现与裸地相比,经过4、14、24和34 a恢复,土壤有机碳固存能力分别为2.97、2.01、12.91和19.79 Mg·hm−2;辜翔等[13]采用时空代替法揭示了湘中红壤丘陵区植被恢复对土壤有机碳、碳密度的影响,结果表明:土壤有机碳随植被恢复的变化受到植被因子和土壤因子的共同影响。然而,红壤侵蚀区长期植被恢复后,不同植被恢复模式土壤有机碳质量分数、密度变化及其影响因素还缺乏较为系统的研究。此外,吕茂奎等[14]研究表明:红壤侵蚀地植被恢复 7~11 a,土壤非保护性有机碳质量分数显著增加,其分配比例也明显升高;恢复至 27 a后分配比例保持在较稳定水平,土壤有机碳的输入与微生物分解过程逐渐达到相对平衡状态。本研究以江西退化红壤森林植被恢复国家长期科研基地经过27 a人工植被恢复重建的6种林分为研究对象,以未加人工干扰的自然恢复地为对照,分析不同植被恢复模式土壤有机碳质量分数、密度的变化特征,探讨植物生物量及土壤性质变化对土壤有机碳储量的影响,为退化红壤区生态修复及碳库管理提供科学支撑。
-
江西退化红壤森林植被恢复国家长期科研基地是典型红壤侵蚀退化丘陵区,地处江西省泰和县石山乡,26°54′~ 26°55′N,114°48′~ 114°49′E,为典型的亚热带湿润季风气候,水热资源丰富,年平均气温为18.6 ℃,年降雨量为1 726.0 mm,年平均日照时数为1 756.4 h。研究区土壤母质为第四纪红土,表层石砾含量较多,属于典型的红壤低丘岗。由于长期的乱砍滥伐、过度放牧、割灌铲草,导致土壤侵蚀严重、土地贫瘠,仅有一些干旱和半干旱草(狗牙根属Cynodon)零星分布,土壤有机质质量分数极低(6.5 g·kg−1)[5]。1991年选用湿地松P. elliottii、马尾松、晚松P. serotina、火炬松P. taeda、枫香Liquidambar formosana、樟树Cinnamomum camphora、木荷、油桐Vernicia fordii等树种,在该区采取不同配置模式开展荒山造林植被恢复试验研究,共有84个小班连片分布。目前,试验区已郁闭成林。
-
在研究区内选取6种典型人工植被恢复类型(马尾松纯林、湿地松纯林、木荷纯林、湿地松-木荷混交林、马尾松补植木荷林和湿地松补植木荷林),每种植被恢复类型设置5个20 m×20 m的标准样地。此外,在基地外选择3个未加人工干扰的自然恢复样地作为对照(表1)。于2018年7月在每个样地内沿对角线均匀挖取3个土壤剖面,共99个剖面。已有研究表明:红壤侵蚀地不同植被恢复措施对于40 cm以下土壤有机碳质量分数影响不大[15]。因此,本研究以40 cm为采样深度,按0~10、10~20、20~30、30~40 cm分层, 从下而上采集土壤样品, 同时用环刀法测定土壤容重、含水率。
表 1 7种植被恢复模式的基本情况
Table 1. General characteristics of seven vegetation restoration patterns
植被恢复模式 面积/hm2 优势种 恢复年限/a 胸径/cm 树高/m 马尾松纯林(PM) 17.16 马尾松 27 11.40±0.75 7.39±0.26 湿地松纯林(PE) 17.74 湿地松 27 17.32±2.05 8.98±0.74 木荷纯林(SS) 11.61 木荷 27 11.63±0.32 9.02±0.76 湿地松-木荷混交林(MES) 5.46 湿地松、木荷 27 13.64±0.47 9.33±0.63 马尾松补植木荷林(RMS) 19.07 马尾松、木荷 27 14.08±0.36 8.39±0.33 湿地松补植木荷林(RES) 14.81 湿地松、木荷 27 14.18±0.44 7.91±0.15 自然恢复样地(ck) 狗牙根、金樱子、狗尾草 27 说明:狗牙根Cynodon dactylon;金樱子Rosa laevigata;狗尾草Setaria viridis。马尾松补植木荷林初植为马尾松,2010年补植木荷;湿地松补植木荷林初植为湿地松,2006年补植木荷。胸径和树高数值为平均值±标准差。 对每个样地中胸径>5 cm的林木进行每木检尺,记录胸径、树高等,其次在每个样地沿对角线均匀布置5个1 m×1 m样方,收集样方内的全部凋落物,称湿质量后,将约300 g带回实验室。野外采集的样品带回实验室处理后用于测定相关指标。有机碳采用重铬酸钾容量法-外加热法(H2SO4-K2Cr2O7),土壤容重和孔隙度的测定采用环刀法,土壤含水率采用烘干法,pH采用电极法,全氮、全磷采用高氯酸-硫酸消煮法,全自动间断化学分析仪(Smart Chem 200 Alliance Corp.)上机测定[16]。
-
土壤有机碳密度及有机碳储量参考CHEN等[17]的计算方法。土壤有机碳密度(DSOC)计算公式如下:DSOCi= CiρiHi(1−F)/10。其中:DSOCi为第i层土壤有机碳密度 (Mg·hm−2);Ci为第i层土壤有机碳质量分数(g·kg−1);ρi为第i层土壤容重 (g·cm−3),Hi为剖面深度(cm);F为粒径>2 mm的砾石质量百分比(%);10−1为单位转换系数。乔木生物量按照目前中国执行的立木生物量模型及碳计量参数标准[18−20]进行估算。
样品数据录入后,采用SPSS 27.0对不同植被恢复类型和不同土层深度的有机碳质量分数和有机碳密度差异进行单因素方差分析(one-way ANOVA),采用最小显著性差异(LSD) 法进行差异显著性检验(P<0.05);采用皮尔逊(Pearson)相关分析探究土壤有机碳密度的影响因素,使用Origin 2022和ArcGIS 10.8.1绘图。
-
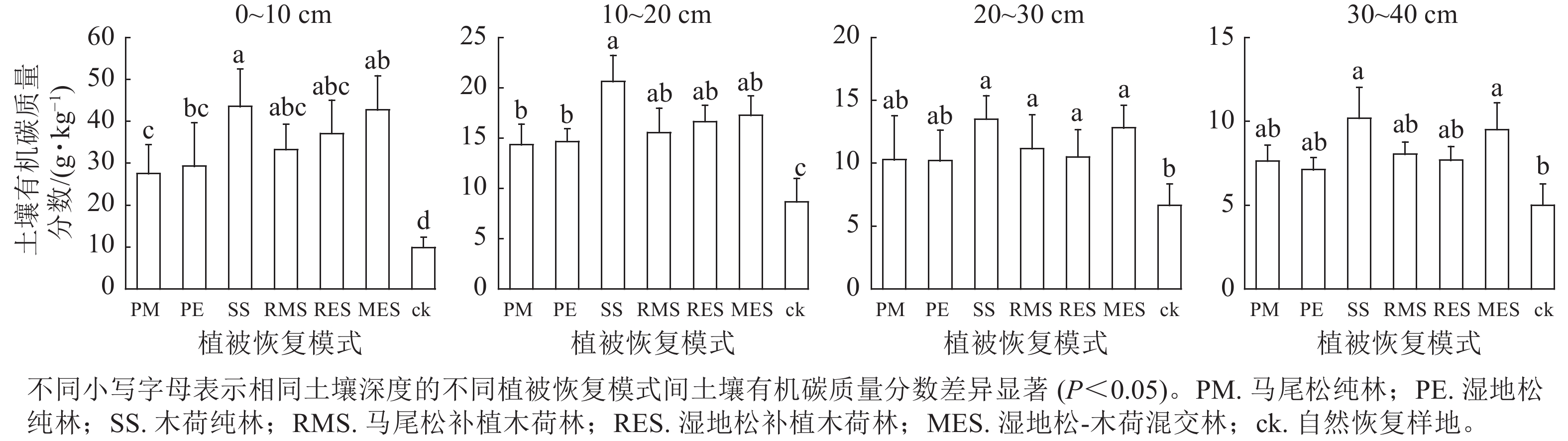
由图1可以看出:在0~20 cm土层中,退化红壤区6种人工植被恢复模式的土壤有机碳相较自然恢复样地均有显著增加(P<0.05);在20~30 cm土层中,除了马尾松纯林和湿地松纯林外,其余4种人工植被恢复模式土壤有机碳质量分数均显著高于对照(P<0.05);在30~40 cm土层中,仅有木荷纯林和湿地松-木荷混交林的土壤有机碳质量分数显著高于对照(P<0.05)。比较6种人工植被恢复模式发现,土壤有机碳在0~10 cm土层中木荷纯林显著高于湿地松纯林和马尾松纯林,湿地松-木荷混交林显著高于马尾松纯林(P<0.05);在10~20 cm土层中仅有木荷纯林显著高于湿地松纯林和马尾松纯林(P<0.05);在20~30和30~40 cm土层中6种人工植被恢复模式之间无显著差异。
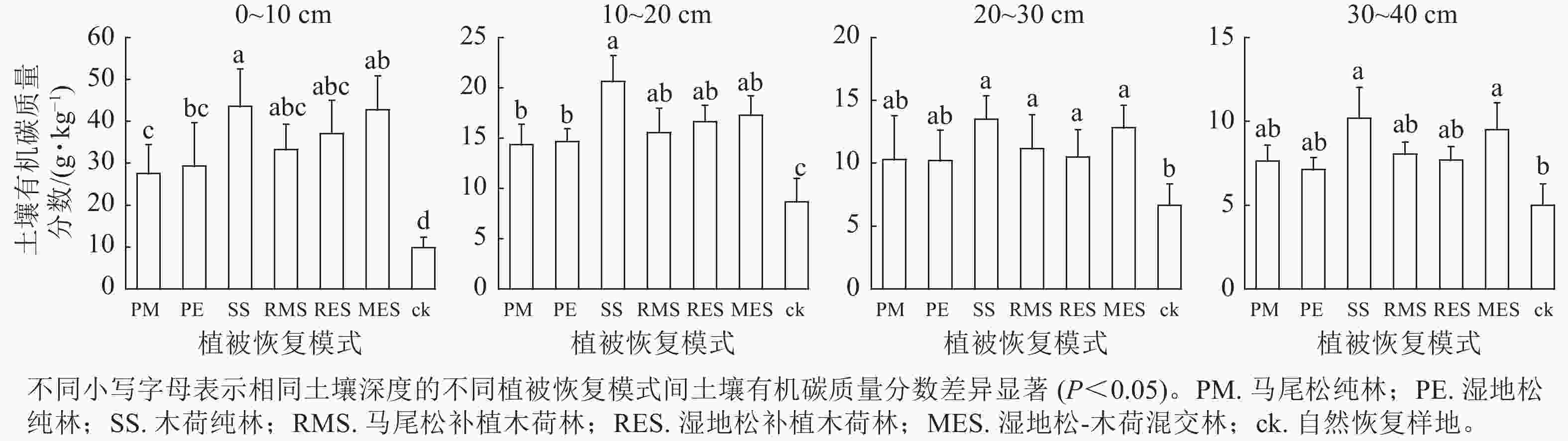
图 1 不同植被恢复模式间土壤有机碳质量分数特征
Figure 1. Characteristics of soil organic carbon contents among different vegetation restoration patterns
-
从图2可以看出:所有植被恢复模式不同土层的土壤有机碳质量分数分布规律基本一致,均表现出较强的表聚效应,即0~10 cm表层土壤有机碳最多,且随着土壤深度的增加土壤有机碳呈现减少的趋势,但不同植被恢复模式土壤有机碳层间差异不同。其中木荷纯林的层间差异最大,0~10 cm土层土壤有机碳质量分数与30~40 cm土层相差33.26 g·kg−1,其次为湿地松-木荷混交林、湿地松补植木荷林、马尾松补植木荷林、湿地松纯林、马尾松纯林,对照不同土层之间的差异最小,仅相差4.90 g·kg−1。不同植被恢复模式0~20 cm土层土壤有机碳质量分数均占0~40 cm土层的60 %以上。
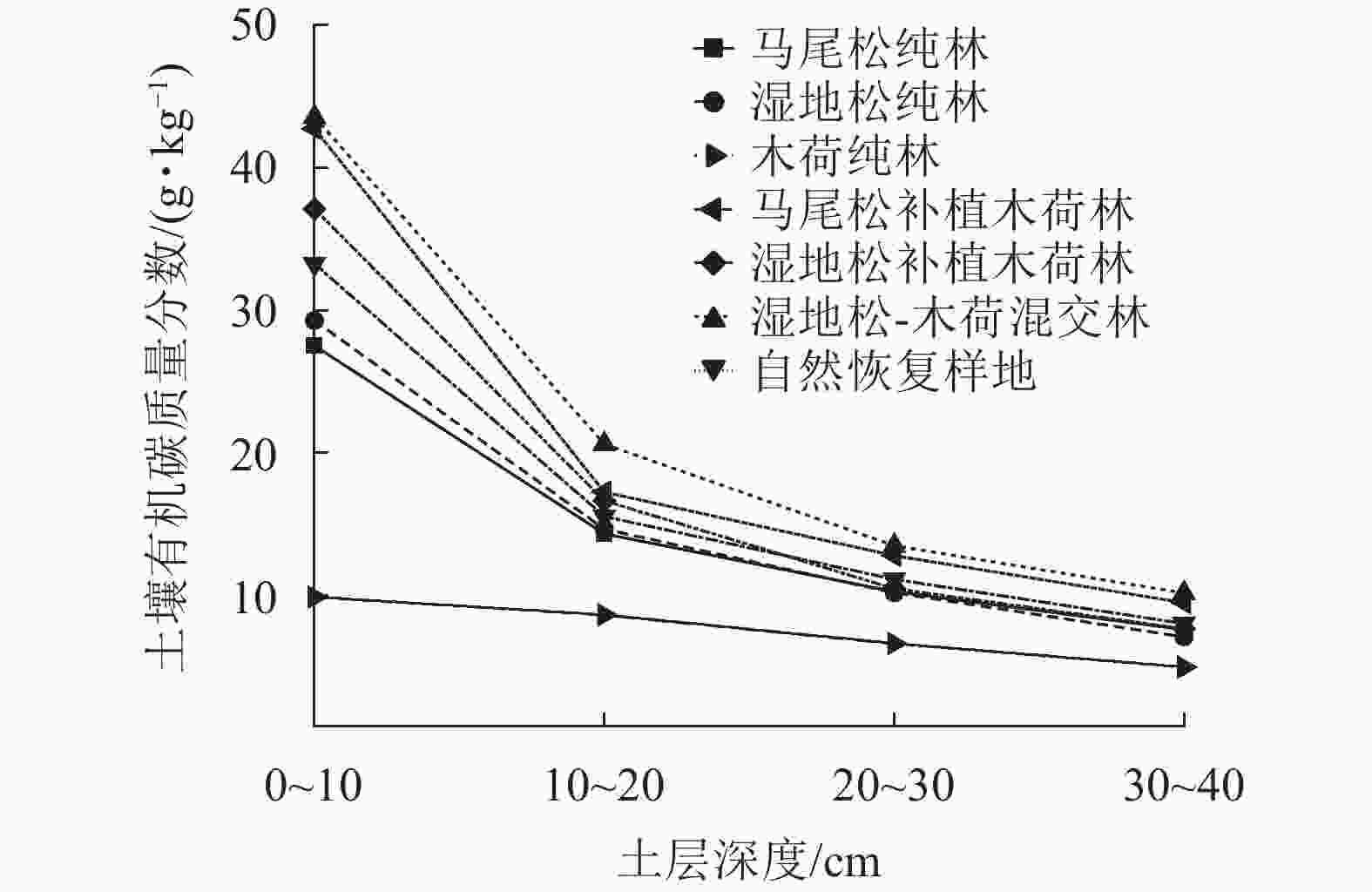
图 2 不同植被恢复模式间土壤有机碳质量分数垂直分布特征
Figure 2. Vertical distribution characteristics of soil organic carbon contents between different vegetation restoration patterns
-
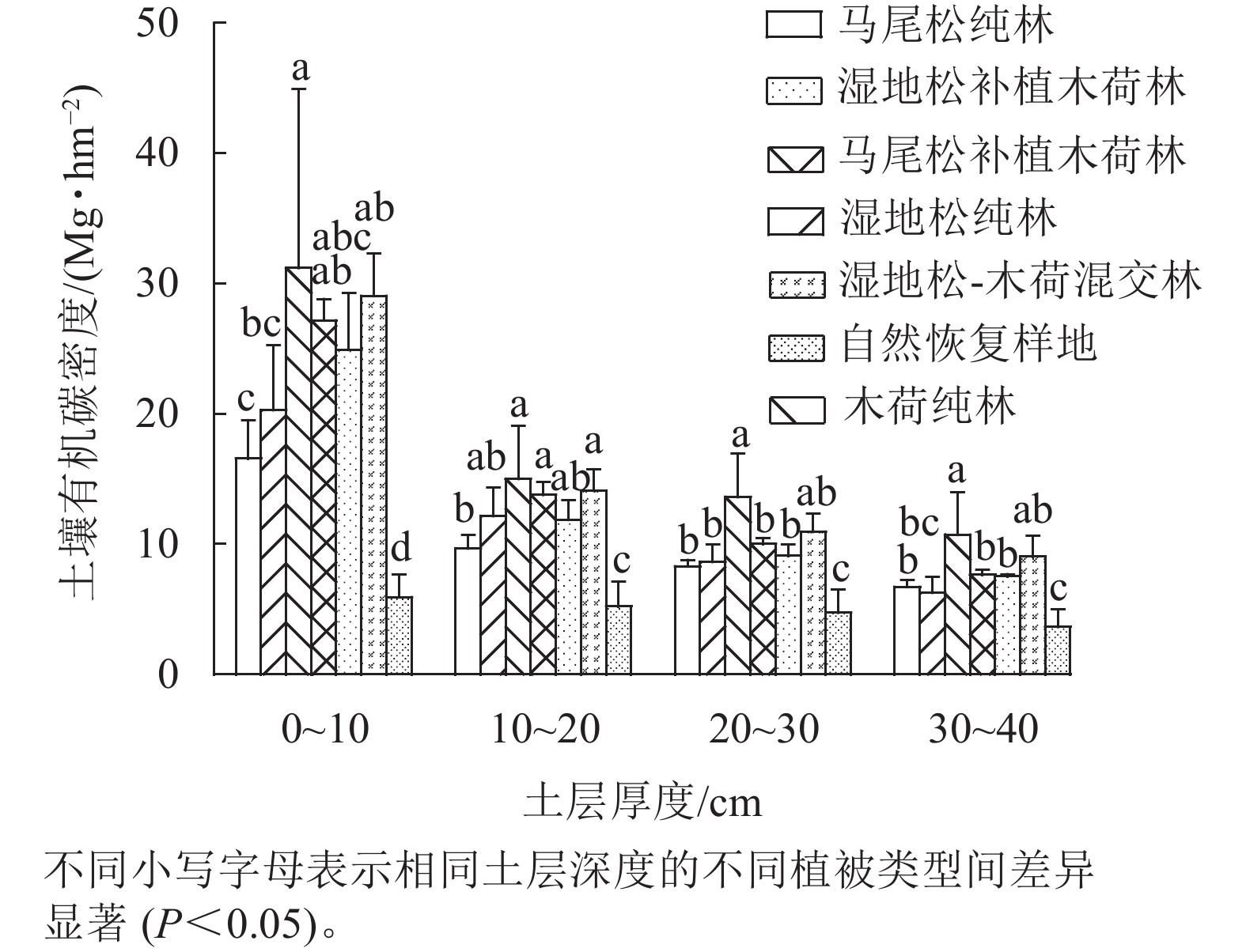
从图3可以看出:不同植被恢复模式之间土壤有机碳密度存在显著差异(P<0.05),其中在0~10、10~20和20~30 cm土层中,6种人工植被恢复模式的土壤有机碳密度均显著大于对照,在30~40 cm土层中除了湿地松纯林外,其余5种人工植被恢复模式的土壤有机碳密度均显著大于对照(P<0.05)。比较6种人工植被恢复模式发现,在0~10 cm土层中,木荷纯林的土壤有机碳密度显著大于湿地松纯林和马尾松纯林,湿地松-木荷混交林的土壤有机碳密度显著大于马尾松纯林(P<0.05);在10~20 cm土层,木荷纯林、湿地松-木荷混交林和马尾松补植木荷林的土壤有机碳密度显著大于马尾松纯林(P<0.05);在20~30和30~40 cm土层,木荷纯林的土壤有机碳密度显著大于马尾松补植木荷林、湿地松补植木荷林、马尾松纯林和湿地松纯林(P<0.05)。总体而言,土壤有机碳密度变化呈现出随土壤剖面深度的增加而减少的趋势。
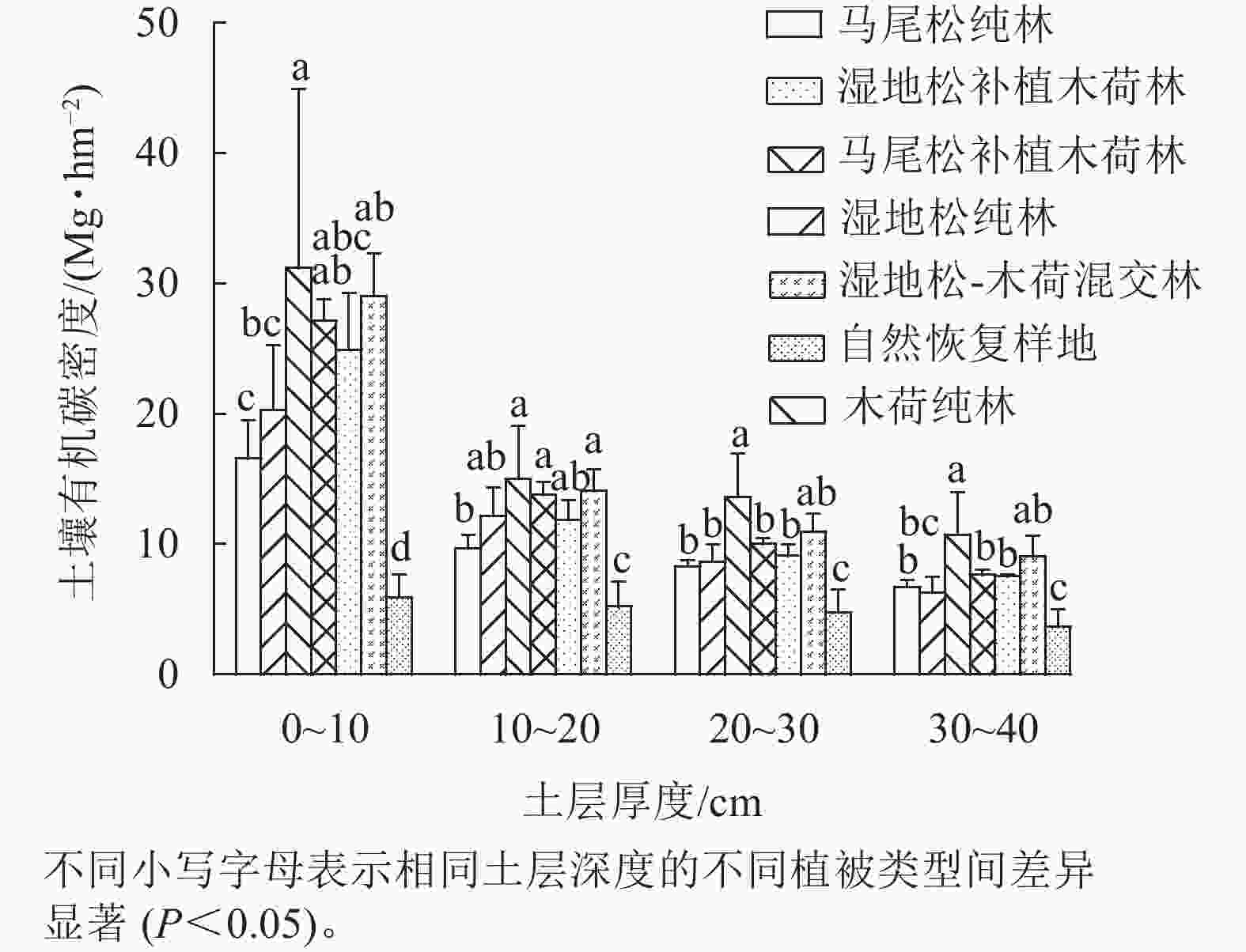
图 3 不同植被恢复模式土壤有机碳密度分布特征
Figure 3. Distribution characteristics of soil organic carbon density in different vegetation restoration patterns
-
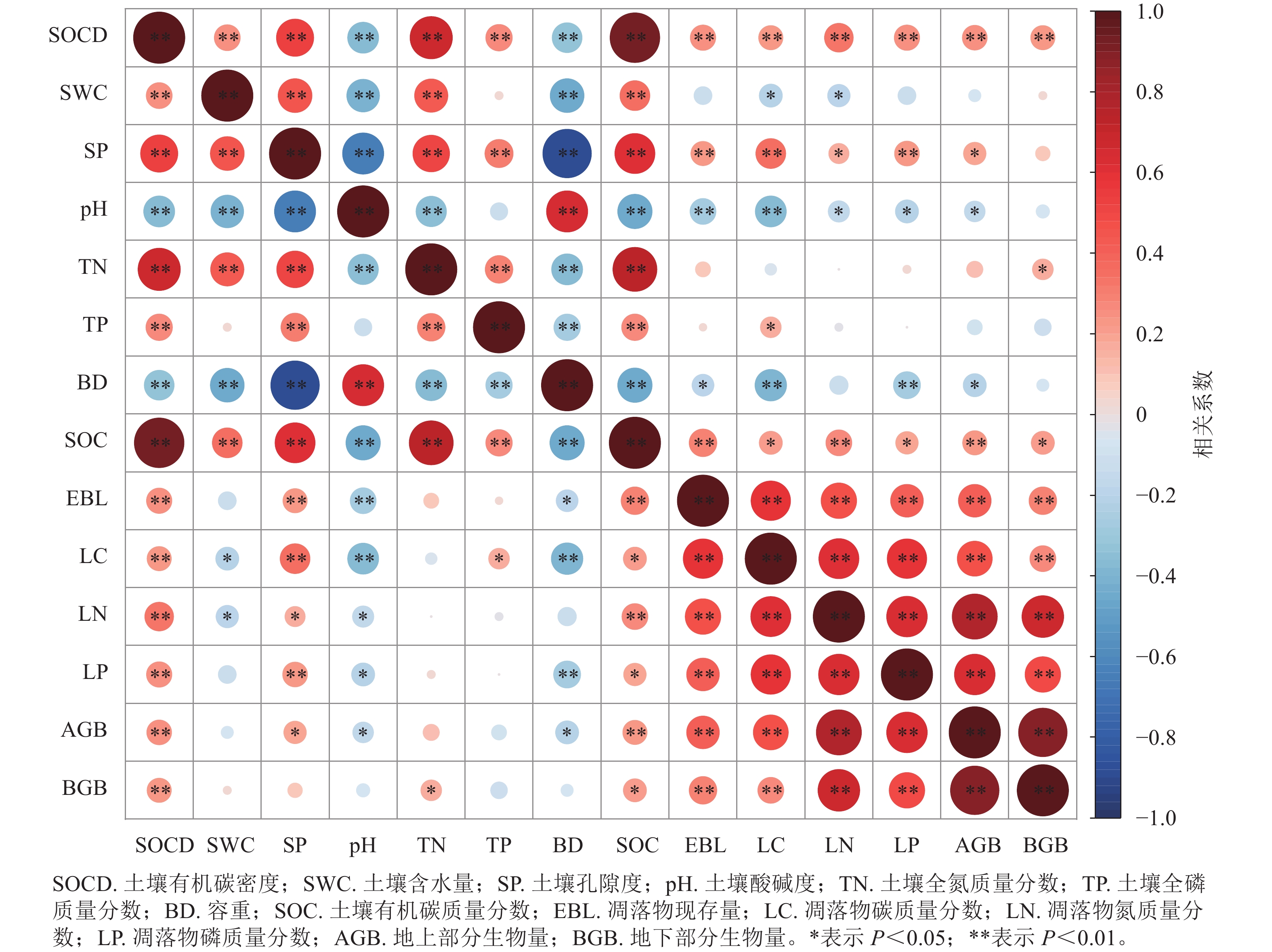
从图4可以看出:土壤有机碳密度与土壤理化性状和植被因子之间均存在显著相关性。其中,土壤有机碳密度与土壤有机碳质量分数(r=0.932,P<0.01)、土壤含水量(r=0.258,P<0.01)、土壤孔隙度(r=0.533,P<0.01)、土壤全氮质量分数(r=0.674,P<0.01)、土壤全磷质量分数(r=0.268,P<0.01)、凋落物现存量(r=0.254,P<0.01)、凋落物碳质量分数(r=0.229,P<0.05)、凋落物氮质量分数(r=0.322,P<0.01)、凋落物磷质量分数(r=0.246,P<0.05)、地上部分生物量(r=0.243,P<0.01)和地下部分生物量(r=0.237,P<0.01)呈显著或极显著正相关,与pH (r=−0.367)和土壤容重(r=−0.336)呈极显著负相关(P<0.01)。土壤有机碳密度与土壤有机碳质量分数相关系数最大,与地下部分生物量的相关系数最小。总体上看,土壤理化性状和植被因子对土壤有机碳的影响与土壤有机碳密度基本一致。

图 4 土壤有机碳密度与土壤理化性状、植被因子间的皮尔逊相关分析
Figure 4. Pearson correlation analysis between SOCD and soil physicochemical properties and vegetation factors
-
表2为研究区经过27 a植被恢复与重建后不同植被恢复模式的土壤有机碳储量。从表2可以看出:0~40 cm土层土壤有机碳储量从高到低依次为木荷纯林、湿地松-木荷混交林、马尾松补植木荷林、湿地松补植木荷林、湿地松纯林、马尾松纯林,与对照相比,土壤有机碳储量分别增加了50.12、42.73、38.20、33.03、26.93和20.85 Mg·hm−2,最大提升了2.46倍,最小提升了1.02倍,说明人工植被恢复对促进土壤有机碳积累效果显著。
表 2 退化红壤区植被恢复土壤有机碳储量
Table 2. Soil organic carbon stocks in revegetated red soil areas with degradation
土层深度/cm 土壤有机碳储量/( Mg·hm−2) 马尾松纯林 湿地松纯林 木荷纯林 马尾松补植木荷林 湿地松补植木荷林 湿地松-木荷混交林 自然恢复样地 0~10 16.56 20.28 31.19 27.18 24.92 29.03 6.06 10~20 9.68 12.16 15.00 13.78 11.88 14.11 5.51 20~30 8.30 8.63 13.62 10.01 9.11 10.96 4.98 30~40 6.72 6.27 10.72 7.64 7.53 9.04 3.86 0~40 41.26 47.34 70.53 58.61 53.44 63.14 20.41 -
研究区经过27 a人工植被恢复后,土壤有机碳质量分数为17.57 g·kg−1,土壤有机碳储量为55.72 Mg·hm−2,与闽西[21]和赣南[12]红壤侵蚀区植被恢复24 a后0~40 cm土层土壤碳密度(分别为27.28和49.45 Mg·hm−2)相比,土壤有机碳储量较高。这主要是与植被恢复类型有关,土壤有机碳主要来源于植被凋落物及地下根系的分解和周转[22]。凋落物的分解速率及地下根系的分布和周转因植被类型不同而有所差异,因此植被类型在很大程度上影响着土壤碳库。土壤有机碳储量即使在同一地区或同一气候条件下, 也可能会因植被类型不同而产生较大的差异[23]。研究结果与湘中红壤侵蚀区植被恢复26 a后0~40 cm土层土壤有机碳相比,土壤有机碳质量分数高于湘中红壤侵蚀区(12.16 g·kg−1)[24],土壤有机碳密度却低于湘中地区(71.40 Mg·hm−2)[13]。这一方面是因为辜翔等[13]的研究中没有去除粒径>2 mm砾石,系统地高估了有机碳储量[25];另一方面是因为在本研究区砾石较多,平均达到35.8%,因此本研究区土壤有机碳质量分数较高,土壤有碳储量却较低。植被恢复过程中,植被层的形成和发育会改变植被层生产力及土壤有机物质积累和分解,进而影响土壤碳质量分数和储量[2]。通常的恢复模式是使用具有高存活率的先锋物种在景观中定居,然后在环境条件允许的情况下逐渐引入当地的顶极物种。本研究区直接利用当地顶极物种[5]来恢复严重侵蚀退化土壤,并取得巨大成功,这对中国南方红壤侵蚀区大规模造林和再造林工程具有一定的参考意义。
-
本研究发现:对比自然恢复地,6种人工植被恢复模式0~40 cm土层土壤有机碳质量分数均有所增加,但随着土层深度的增加差异逐渐减小。这与刘苑秋等[26]对本研究区植被恢复早期(10 a)的研究结果不一致,其研究结果表明:0~40 cm土层内重建森林土壤的有机质质量分数低于对照区。这主要是因为:①自然恢复地虽然没有进行人工重建植被,但在不樵采、不放牧的保护措施下,一些阳性草本植物生长较快,地表植被盖度达90%[26]。在植被恢复初期(10 a),地表植物凋落物量更多,凋落物及地下根系的分解与周转速率更快,土壤有机碳的固持和保护作用更强。②已有研究表明:在植被恢复早期,原先土壤碳的缓慢分解是中国南方红壤侵蚀区有机碳增加的重要原因[27],而人工植被恢复模式对土壤有机碳的消耗远大于自然植被恢复模式,因此在植被恢复早期自然恢复地土壤有机碳质量分数高于人工重建植被。然而随着恢复年限的增加,人工植被恢复模式群落总生物量及各组分生物量增高,地表凋落物现存量逐渐增多,凋落物层质量明显改善,且被土壤微生物分解的速率更快,有机物输入量更大[28];同时地表裸露面积减少, 地表径流侵蚀减弱,土壤环境质量逐渐改善和稳定,土壤团聚体物理保护的增强,减缓了土壤有机碳的损失,因此土壤有机碳质量分数逐渐高于自然恢复地。
研究还发现:6种人工植被恢复模式0~20 cm土层土壤有机碳质量分数也存在差异,其中木荷纯林的土壤有机碳质量分数显著高于马尾松纯林和湿地松纯林。一方面是湿地松纯林和马尾松纯林凋落物较少,且存在大量顽抗的化合物,导致其分解速率较低并且减慢了颗粒有机物向矿质土壤转化的速度。木荷纯林的凋落物产量大,分解速率较快[29],导致木荷纯林表层土壤有机碳质量分数显著高于马尾松纯林和湿地松纯林。另一方面是由于木荷纯林的地下根系发达且向更大范围伸展,有效改善了土壤储水功能和孔隙度[30]。土壤水分得到改善后提高了植物生产力,减缓了有机碳矿化速率[31]。土壤孔隙度增大有利于土壤中水分和气体的运移与交换[32],可以更好地固持有机碳中的颗粒物,引起颗粒有机碳的变化,促进有机碳的积累[33]。然而随着土层深度的增加,发育于同一母质的深层土壤,受植物凋落物、根系的影响程度减弱,使得不同植被恢复模式土壤有机碳质量分数之间的差异也随着土壤深度的增加而减弱[24]。6种人工植被恢复模式在20~40 cm土层土壤有机碳质量分数差异不显著,这也表明:植被恢复过程中土壤有机碳的积累需要一个过程,表层土壤碳库比深层恢复更快。单一的凋落物会降低微生物丰度与多样性,导致其分解残留率显著增高[34],因此虽然阔叶林的凋落物量更多,但是混交林的土壤微生物活性更高,分解凋落物和地下根系的速率更快,导致湿地松-木荷混交林、马尾松补植木荷林和湿地松补植木荷林等3种恢复模式与木荷纯林的0~40 cm土层土壤有机碳质量分数均无显著差异。同一气候条件下, 相同厚度土层土壤有机碳密度的变化取决于土壤有机碳质量分数和土壤容重。受到土壤容重的影响,本研究中不同植被恢复模式土壤有机碳密度的变化规律与土壤有机碳存在些许差异,但总体变化趋势基本相同。此外,本研究区土壤有机碳质量分数分布呈现随土层深度增加而减少的特征,但不同植被恢复模式土壤有机碳的层间差异不同。这与众多研究结果一致[2, 12−13]。这是因为地表凋落物和植物根系分解所形成的土壤有机碳首先进入土壤表层,使得表层土壤有机碳明显高于深层土壤,且林分类型对凋落物产量和分解速率均有显著影响,进而造成土壤有机碳层间差异有所不同[35]。
植被恢复是一个漫长的过程。在这个过程中土壤有机碳质量分数与植被因子、土壤理化性状密切相关,但不同因子对土壤有机碳质量分数影响不尽相同。研究表明:土壤有机碳和氮、磷均存在正相关关系,这是因为土壤碳和氮之间有较强的交互作用,土壤氮的增加会促进植物向土壤中转移更多的有机碳[36],同时有机质质量分数高的土壤中往往含有大量有机磷[32],磷质量分数的提升可促进土壤有机质分解[37]。凋落物氮、磷能为微生物群落提供充足的养分和能源,从而提高微生物活性,促进凋落物的分解[38],因此土壤有机碳与凋落物氮、磷也呈正相关。土壤有机碳质量分数与pH呈显著负相关。这与JIA等[39]的研究结果一致。这是由于较低的土壤pH通常会抑制微生物的活性和数量,从而减慢土壤有机碳的分解速率,有利于土壤有机碳的积累[40]。土壤有机碳密度与土壤有机碳质量分数一样,也与土壤理化性状、植被因子密切相关,且与土壤有机碳的变化规律基本一致。
-
研究区人工植被恢复模式下土壤有机碳质量分数为17.57 g·kg−1,土壤有机碳储量为55.72 Mg·hm−2,显著高于自然恢复地,说明人工植被恢复与重建取得了巨大成功,可以在当地继续推广。在不同人工植被恢复模式0~20 cm 土层的土壤有机碳质量分数之间存在显著差异,其中木荷纯林显著高于马尾松纯林和湿地松纯林,但与湿地松-木荷混交林、湿地松补植木荷林和马尾松补植木荷林均无显著差异,因此在研究区今后的建设中,可通过间伐补植阔叶树等方式对针叶纯林进行改造,进而增强土壤固碳能力。研究区7种植被恢复模式不同土层的土壤有机碳质量分数分布规律基本一致,均表现出较强的表聚效应,表明土壤有机碳主要储存在土壤表层。因此, 应该加强地表植被层的保护,减少人为活动的干扰,促进植物群落恢复,维持土壤碳库的稳定。土壤有机碳密度的变化规律与土壤有机碳质量分数基本一致,是植被因子和土壤理化性状共同作用的结果,在今后的研究中应该综合考虑。
Characteristics and influencing factors of soil carbon stocks in different vegetation restoration models in red soil erosion areas
-
摘要:
目的 探讨江西红壤侵蚀区植被恢复过程中不同植被恢复模式的土壤碳储量特征,为揭示植被恢复对土壤碳汇的影响机制和江西红壤侵蚀区生态修复及碳库管理提供科学支撑。 方法 以经过27 a植被恢复与重建后的6种人工植被恢复模式(马尾松Pinus massoniana纯林、湿地松P. elliottii纯林、木荷Schima superba纯林、湿地松-木荷混交林、马尾松补植木荷林、湿地松补植木荷林)为研究对象,以未加人工干扰的自然恢复地为对照,比较不同植被恢复模式土壤有机碳质量分数、储量的变化特征,并分析其影响因素。 结果 ①相较于对照,在0~20 cm土层中,6种人工植被恢复模式土壤有机碳质量分数均有显著增加(P<0.05);在20~30 cm土层中,除了马尾松纯林和湿地松纯林外,其余4种人工植被恢复模式土壤有机碳质量分数均显著高于对照(P<0.05);在30~40 cm土层中,仅有木荷纯林和湿地松-木荷混交林的土壤有机碳质量分数显著高于对照(P<0.05)。②研究区7种植被恢复模式的土壤有机碳质量分数均表现出较强的表聚效应,但不同植被恢复模式土层间差异不同,其中木荷纯林最大,0~10 cm土层土壤有机碳质量分数与30~40 cm土层相差33.26 g·kg−1,对照最小,仅相差4.90 g·kg−1。③土壤有机碳密度变化规律与质量分数变化规律基本一致,均是植被因子与土壤理化性状共同作用的结果。植被因子中凋落物氮质量分数与土壤有机碳密度的相关系数最大(r=0.322,P<0.01),土壤理化性状中土壤有机碳质量分数与土壤有机碳密度的相关系数最大(r=0.932,P<0.01)。④土壤有机碳储量从高到低依次表现为木荷纯林、湿地松-木荷混交林、马尾松补植木荷林、湿地松补植木荷林、湿地松纯林、马尾松纯林,与对照相比,分别增加了50.12、42.73、38.20、33.03、26.93和20.85 Mg·hm−2,最大提升了2.46倍,最小提升了1.02倍。 结论 人工植被恢复与重建在红壤侵蚀区效果显著,在研究区今后的建设中可继续推广,以促进植物群落恢复,提高和维持土壤碳库的稳定。图4表2参40 Abstract:Objective This study is to explore the soil carbon stock characteristics of different vegetation restoration models in the process of vegetation restoration in red soil erosion areas of Jiangxi Province, so as to provide scientific support for revealing the impact mechanism of vegetation restoration on soil carbon sinks and ecological restoration and carbon pool management in red soil erosion areas of Jiangxi Province. Method The research objects included 6 types of forest stands after 27 years of vegetation restoration and reconstruction: Pinus massoniana pure forest (PM), P. elliottii pure forest (PE), Schima superba pure forest (SS), P. elliottii and S. superba mixed forest (MES), replanting S. superba with P. massoniana (RMS), and replanting S. superba with P. elliottii (RES). Natural restoration sites without artificial disturbance (ck) were used as controls to compare the changes in soil organic carbon (SOC) content and storage under different vegetation restoration models, and analyze their influencing factors. Result (1) Compared with ck, SOC mass fraction increased significantly (P<0.05) in the 0−20 cm soil layer in all 6 artificial vegetation restoration models. In the 20−30 cm soil layer, except for PM and PE, SOC mass fraction of the other 4 vegetation restoration models was significantly higher than that of ck (P<0.05). In the 30−40 cm soil layer, only SS and MES had significantly higher SOC mass fractions than ck (P<0.05). (2) SOC mass fractions of the 7 vegetation restoration models in the study area showed a strong surface aggregation effect, but the differences between soil layers of different vegetation restoration models were different, among which SS had the largest SOC mass fraction, with a difference of 33.26 g·kg−1 between 0−10 cm soil layer and 30−40 cm layer, and ck had the smallest difference, with only 4.90 g·kg−1. (3) The change rules of SOC density and SOC content were basically the same, both of which were the result of the joint action of vegetation factors and soil physicochemical properties. The correlation coefficient between litter nitrogen mass fraction (r=0.322, P<0.01) and SOC density was the largest among vegetation factors, and the correlation coefficient between SOC mass fractions (r=0.932, P<0.01) and SOC density was the largest among soil physicochemical traits. (4) From high to low, SOC storage increased by 50.12 Mg·hm−2 in SS, 42.73 Mg·hm−2 in MES, 38.20 Mg·hm−2 in RMS, 33.03 Mg·hm−2 in RES, 26.93 Mg·hm−2 in PE, and 20.85 Mg·hm−2 in PM, respectively, compared with ck, with a maximum enhancement of 2.46 times and a minimum enhancement of 1.02 times. Conclusion Artificial vegetation restoration and reconstruction have significant effects in red soil erosion areas and can be further promoted in future construction in the research area to promote plant community restoration and enhance and maintain the stability of soil carbon pool. [Ch, 4 fig. 2 tab. 40 ref.] -
图 1 不同植被恢复模式间土壤有机碳质量分数特征
Figure 1 Characteristics of soil organic carbon contents among different vegetation restoration patterns
图 2 不同植被恢复模式间土壤有机碳质量分数垂直分布特征
Figure 2 Vertical distribution characteristics of soil organic carbon contents between different vegetation restoration patterns
图 3 不同植被恢复模式土壤有机碳密度分布特征
Figure 3 Distribution characteristics of soil organic carbon density in different vegetation restoration patterns
图 4 土壤有机碳密度与土壤理化性状、植被因子间的皮尔逊相关分析
Figure 4 Pearson correlation analysis between SOCD and soil physicochemical properties and vegetation factors
表 1 7种植被恢复模式的基本情况
Table 1. General characteristics of seven vegetation restoration patterns
植被恢复模式 面积/hm2 优势种 恢复年限/a 胸径/cm 树高/m 马尾松纯林(PM) 17.16 马尾松 27 11.40±0.75 7.39±0.26 湿地松纯林(PE) 17.74 湿地松 27 17.32±2.05 8.98±0.74 木荷纯林(SS) 11.61 木荷 27 11.63±0.32 9.02±0.76 湿地松-木荷混交林(MES) 5.46 湿地松、木荷 27 13.64±0.47 9.33±0.63 马尾松补植木荷林(RMS) 19.07 马尾松、木荷 27 14.08±0.36 8.39±0.33 湿地松补植木荷林(RES) 14.81 湿地松、木荷 27 14.18±0.44 7.91±0.15 自然恢复样地(ck) 狗牙根、金樱子、狗尾草 27 说明:狗牙根Cynodon dactylon;金樱子Rosa laevigata;狗尾草Setaria viridis。马尾松补植木荷林初植为马尾松,2010年补植木荷;湿地松补植木荷林初植为湿地松,2006年补植木荷。胸径和树高数值为平均值±标准差。  下载: 导出CSV
下载: 导出CSV
表 2 退化红壤区植被恢复土壤有机碳储量
Table 2. Soil organic carbon stocks in revegetated red soil areas with degradation
土层深度/cm 土壤有机碳储量/( Mg·hm−2) 马尾松纯林 湿地松纯林 木荷纯林 马尾松补植木荷林 湿地松补植木荷林 湿地松-木荷混交林 自然恢复样地 0~10 16.56 20.28 31.19 27.18 24.92 29.03 6.06 10~20 9.68 12.16 15.00 13.78 11.88 14.11 5.51 20~30 8.30 8.63 13.62 10.01 9.11 10.96 4.98 30~40 6.72 6.27 10.72 7.64 7.53 9.04 3.86 0~40 41.26 47.34 70.53 58.61 53.44 63.14 20.41
下载: 导出CSV
-
[1] STOCKMANN U, ADAMS M A, CRAWFORD J W, et al. The knowns, known unknowns and unknowns of sequestration of soil organic carbon [J]. Agriculture,Ecosystems &Environment, 2013, 164: 80 − 99. [2] SOUCÉMARIANADIN L N, CÉCILLON L, GUENET B, et al. Environmental factors controlling soil organic carbon stability in French forest soils [J]. Plant and Soil, 2018, 426(1/2): 267 − 286. [3] van der WERF G R, MORTON D C, DEFRIES R S, et al. CO2 emissions from forest loss [J]. Nature Geoscience, 2009, 2(11): 737 − 738. [4] BASTIN J F, FINEGOLD Y, GARCIA C, et al. The global tree restoration potential [J]. Science, 2019, 365(6448): 76 − 79. [5] WEI Xiaohua, LI Qinlin, LIU Yuanqiu, et al. Restoring ecosystem carbon sequestration through afforestation: a sub-tropic restoration case study [J]. Forest Ecology and Management, 2013, 300: 60 − 67. [6] HONG Songbai, YIN Guodong, PIAO Shilong, et al. Divergent responses of soil organic carbon to afforestation [J]. Nature Sustainability, 2020, 3(9): 694 − 700. [7] DAVIDSON E A, JANSSENS I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change [J]. Nature, 2006, 440(7081): 165 − 173. [8] JIN Zhao, DONG Yunshe, WANG Yunqiang, et al. Natural vegetation restoration is more beneficial to soil surface organic and inorganic carbon sequestration than tree plantation on the Loess Plateau of China [J]. Science of the Total Environment, 2014, 485: 615 − 623. [9] SOLLY E F, SCHÖNING I, BOCH S, et al. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands [J]. Plant and Soil, 2014, 382(1): 203 − 218. [10] 李鹏, 刘晓君, 刘苑秋, 等. 退化红壤区不同植被恢复模式的土壤养分空间分布特征[J]. 中南林业科技大学学报, 2023, 43(8): 113 − 124. LI Peng, LIU Xiaojun, LIU Yuanqiu, et al. Spatial distribution characteristics of soil nutrients after vegetation restoration in degraded red soil regions [J]. Journal of Central South University of Forestry &Technology, 2023, 43(8): 113 − 124. [11] 李燕燕, 刘亮英, 吴春生, 等. 亚热带红壤区不同植被恢复类型土壤有机碳δ13C特征[J]. 核农学报, 2020, 34(11): 2561 − 2568. LI Yanyan, LIU Liangying, WU Chunsheng, et al. δ13C characteristic of soil organic carbon under different vegetation restoration in subtropical red soil region [J]. Journal of Nuclear Agricultural Sciences, 2020, 34(11): 2561 − 2568. [12] XIAO Shengsheng, ZHANG Jie, DUAN Jian, et al. Soil organic carbon sequestration and active carbon component changes following different vegetation restoration ages on severely eroded red soils in subtropical China [J/OL]. Forests, 2020, 11(12): 1304[2023-06-10]. doi: 10.3390/f11121304. [13] 辜翔, 张仕吉, 刘兆丹, 等. 中亚热带植被恢复对土壤有机碳含量、碳密度的影响[J]. 植物生态学报, 2018, 42(5): 595 − 608. GU Xiang, ZHANG Shiji, LIU Zhaodan, et al. Effects of vegetation restoration on soil organic carbon concentration and density in the mid-subtropical region of China [J]. Chinese Journal of Plant Ecology, 2018, 42(5): 595 − 608. [14] 吕茂奎, 谢锦升, 周艳翔, 等. 红壤侵蚀地马尾松人工林恢复过程中土壤非保护性有机碳的变化[J]. 应用生态学报, 2014, 25(1): 37 − 44. LÜ Maokui, XIE Jinsheng, ZHOU Yanxiang, et al. Dynamics of unprotected soil organic carbon with the restoration process of Pinus massoniana plantation in red soil erosion area [J]. Chinese Journal of Applied Ecology, 2014, 25(1): 37 − 44. [15] 刘政, 许文斌, 田地, 等. 南方红壤严重侵蚀地不同恢复年限马尾松人工林生态系统碳储量特征[J]. 水土保持通报, 2019, 39(1): 37 − 42. LIU Zheng, XU Wenbin, TIAN Di, et al. Characteristics of ecosystem carbon stocks in Pinus massoniana plantations with different restoration age on severely eroded red soils in southern China [J]. Bulletin of Soil and Water Conservation, 2019, 39(1): 37 − 42. [16] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2008: 30 − 109. BAO Shidan. Soil Agrochemical Analysis [M]. 3rd ed. Beijing: China Agricultural Press, 2008: 30 − 109. [17] CHEN Yuanqi, YU Shiqin, LIU Suqing, et al. Reforestation makes a minor contribution to soil carbon accumulation in the short term: evidence from four subtropical plantations [J]. Forest Ecology and Management, 2017, 384: 400 − 405. [18] 全国森林资源标准化技术委员会. 立木生物量模型及碳计量参数——马尾松: LY/T 2263—2014 [S]. 北京: 中国标准出版社, 2014. National Technical Committee on Forest Resources Standardization. Tree Biomass Models and Related Parameters to Carbon Accounting for Pinus massoniana: LY/T 2263−2014 [S]. Beijing: Standards Press of China, 2014. [19] 全国森林资源标准化技术委员会. 立木生物量模型及碳计量参数——湿地松: LY/T 2261—2014 [S]. 北京: 中国标准出版社, 2014. National Technical Committee on Forest Resources Standardization. Tree Biomass Models and Related Parameters to Carbon Accounting for Pinus elliottii: LY/T 2261−2014 [S]. Beijing: Standards Press of China, 2014. [20] 全国森林资源标准化技术委员会. 立木生物量模型及碳计量参数——木荷: LY/T 2660—2016 [S]. 北京: 中国标准出版社, 2016. National Technical Committee on Forest Resources Standardization. Tree Biomass Models and Related Parameters to Carbon Accounting for Schima superba: LY/T 2660−2016 [S]. Beijing: Standards Press of China, 2016. [21] 谢锦升, 杨玉盛, 解明曙, 等. 植被恢复对侵蚀退化红壤碳吸存的影响[J]. 水土保持学报, 2006, 20(6): 95 − 98, 123. XIE Jinsheng, YANG Yusheng, XIE Mingshu, et al. Effects of vegetation restoration on carbon sequestration in degraded red soil [J]. Journal of Soil and Water Conservation, 2006, 20(6): 95 − 98, 123. [22] LAGANIÈRE J, ANGERS D A, PARÉ D. Carbon accumulation in agricultural soils after afforestation: a meta-analysis [J]. Global Change Biology, 2010, 16(1): 439 − 453. [23] ZHAO Yonggang, LIU Xiaofang, WANG Zilong, et al. Soil organic carbon fractions and sequestration across a 150-yr secondary forest chronosequence on the Loess Plateau, China [J]. Catena, 2015, 133: 303 − 308. [24] 王振鹏, 陈金磊, 李尚益, 等. 湘中丘陵区不同恢复阶段森林生态系统的碳储量特征[J]. 林业科学, 2020, 56(5): 19 − 28. WANG Zhenpeng, CHEN Jinlei, LI Shangyi, et al. Characteristics of forest ecosystem carbon stocks at different vegetation restoration stages in hilly area of central Hunan Province, China [J]. Scientia Silvae Sinicae, 2020, 56(5): 19 − 28. [25] POEPLAU C, VOS C, DON A. Soil organic carbon stocks are systematically overestimated by misuse of the parameters bulk density and rock fragment content [J]. Soil, 2017, 3(1): 61 − 66. [26] 刘苑秋, 杜天真, 郭晓敏, 等. 退化第四纪红黏土重建森林模式及其土壤恢复研究[J]. 水土保持学报, 2004, 18(6): 34 − 37. LIU Yuanqiu, DU Tianzhen, GUO Xiaomin, et al. Study on rehabilitating forest model and soil restoration on Quaternary red soil [J]. Journal of Soil and Water Conservation, 2004, 18(6): 34 − 37. [27] DOU Xiaolin, DENG Qi, LI Ming, et al. Reforestation of Pinus massoniana alters soil organic carbon and nitrogen dynamics in eroded soil in south China [J]. Ecological Engineering, 2013, 52: 154 − 160. [28] 王志齐, 杜兰兰, 赵慢, 等. 黄土区不同退耕方式下土壤碳氮的差异及其影响因素[J]. 应用生态学报, 2016, 27(3): 716 − 722. WANG Zhiqi, DU Lanlan, ZHAO Man, et al. Differences in soil organic carbon and total nitrogen and their impact factors under different restoration patterns in the Loess Plateau [J]. Chinese Journal of Applied Ecology, 2016, 27(3): 716 − 722. [29] WANG Bing, XUE Sha, LIU Guobin, et al. Changes in soil nutrient and enzyme activities under different vegetations in the Loess Plateau area, northwest China [J]. Catena, 2012, 92: 186 − 195. [30] 袁星明, 朱宁华, 郭耆, 等. 南亚热带不同人工林对土壤理化性质的影响及土壤质量评价[J]. 林业科学研究, 2022, 35(3): 112 − 122. YUAN Xingming, ZHU Ninghua, GUO Shi, et al. Effects of different plantations on soil physical and chemical properties and soil quality evaluation in south subtropical zone [J]. Forest Research, 2022, 35(3): 112 − 122. [31] WANG Dong, LIU Yu, WU Gaolin, et al. Effect of rest-grazing management on soil water and carbon storage in an arid grassland (China) [J]. Journal of Hydrology, 2015, 527: 754 − 760. [32] 许婷婷, 董智, 郭建英, 等. 放牧对内蒙古典型草原植物群落特征与土壤有机碳的影响[J]. 草地学报, 2022, 30(9): 2273 − 2279. XU Tingting, DONG Zhi, GUO Jianying, et al. Effects of grazing on plant community and soil organic carbon in the typical steppe of Inner Mongolia, China [J]. Acta Agrestia Sinica, 2022, 30(9): 2273 − 2279. [33] 陈美玲, 赵友朋, 张金池, 等. 凤阳山典型林分土壤有机碳特征[J]. 东北林业大学学报, 2022, 50(10): 69 − 75. CHEN Meiling, ZHAO Youpeng, ZHANG Jinchi, et al. Soil organic carbon characteristics of different forest types in Fengyang Mountain [J]. Journal of Northeast Forestry University, 2022, 50(10): 69 − 75. [34] 王小平, 杨雪, 杨楠, 等. 凋落物多样性及组成对凋落物分解和土壤微生物群落的影响[J]. 生态学报, 2019, 39(17): 6264 − 6272. WANG Xiaoping, YANG Xue, YANG Nan, et al. Effects of litter diversity and composition on litter decomposition characteristics and soil microbial community [J]. Acta Ecologica Sinica, 2019, 39(17): 6264 − 6272. [35] 邓翠, 吕茂奎, 曾敏, 等. 红壤侵蚀区植被恢复过程中土壤有机碳组分变化[J]. 水土保持学报, 2017, 31(4): 178 − 183. DENG Cui, LÜ Maokui, ZENG Min, et al. Dynamies of soil organic carbon fractions with the restoration process of Pinus massoniana plantation in eroded red soil region [J]. Journal of Soil and Water Conservation, 2017, 31(4): 178 − 183. [36] HOPKINS D W, SPARROW A D, ELBERLING B, et al. Carbon, nitrogen and temperature controls on microbial activity in soils from an Antarctic dry valley [J]. Soil Biology and Biochemistry, 2006, 38(10): 3130 − 3140. [37] FISK M, SANTANGELO S, MINICK K. Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests [J]. Soil Biology and Biochemistry, 2015, 81: 212 − 218. [38] 尤龙辉, 叶功富, 陈增鸿, 等. 滨海沙地主要优势树种的凋落物分解及其与初始养分含量的关系[J]. 福建农林大学学报(自然科学版), 2014, 43(6): 585 − 591. YOU Longhui, YE Gongfu, CHEN Zenghong, et al. Litter decomposition and initial nutrient content of major dominant tree species on coastal sandy areas [J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 2014, 43(6): 585 − 591. [39] JIA Guomei, CAO Jing, WANG Chunyan, et al. Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, northwest China [J]. Forest Ecology and Management, 2005, 217(1): 117 − 125. [40] CHEN Shutao, HUANG Yao, ZOU Jianwen, et al. Mean residence time of global topsoil organic carbon depends on temperature, precipitation and soil nitrogen [J]. Global and Planetary Change, 2013, 100: 99 − 108. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230408
 点击查看大图
点击查看大图
计量
- 文章访问数: 2208
- HTML全文浏览量: 375
- PDF下载量: 73
- 被引次数: 0
