




-
钱江源-百山祖国家公园分为钱江源和百山祖2个园区,其中百山祖园区(以下称百山祖国家公园)涵盖了中亚热带东部山地生态系统完整的垂直带谱(低海拔和中山地带均有常绿阔叶林分布),完好地保存了浙闽赣交界山地的代表性和典型性植被和生态系统。其中,百山祖国家公园内保留着大面积迄今未受人为显著干扰的甜槠Castanopsis eyrei-木荷Schima superba和青冈Quercus glauca等常绿阔叶林,这些常绿阔叶林是百山祖国家公园中最具原真性和代表性的植物群落类型之一[1]。
亚热带常绿阔叶林是世界主要森林植被类型之一,主要分布在中国,分布区域约占中国国土面积的1/4,以中亚热带的常绿阔叶林最为典型[2]。由于受到人类干扰的作用,尤其在中国经济发达的东部地区,亚热带原生常绿阔叶林绝大部分退化为次生林或被改造为人工林,老龄林或原生林几乎丧失殆尽[3]。因此,了解百山祖国家公园内的常绿阔叶林的群落结构、物种多样性和生态系统功能,并与周边区域亚热带常绿阔叶林进行对比研究,对于亚热带地区植被恢复、生物多样性保护和生态系统功能提升等均具有重要的理论指导意义。
目前,针对亚热带常绿阔叶林的相关研究主要集中在中国东部亚热带地区,在局域尺度上探讨亚热带常绿阔叶林的群落特征和生境特点[4]、物种多样性和物种共存机制[5]、演替动态与干扰和气候的关系[6−7]以及群落结构和更新[8−9]等方面。张田田等[10]、宋永昌等[11]还在区域尺度上比较了中国亚热带不同区域分布的常绿阔叶林物种组成及群落特征,但尚未包括百山祖国家公园内中山地带和低海拔地带分布的大面积常绿阔叶林。目前,针对百山祖国家公园常绿阔叶林的研究主要以公园内的5和25 hm2常绿阔叶林固定样地为研究平台,在局域尺度上对常绿阔叶林的物种组成、群落特征和群落动态等方面进行了研究[12−13]。因此,为进一步了解百山祖国家公园的常绿阔叶林的特点及与邻近其他地区常绿阔叶林群落特征差异,本研究选择分布于百山祖国家公园内的五岭坑和凤阳山,邻近地区的古田山、九龙山和乌岩岭等自然保护区内以及非自然保护区内的常绿阔叶林为研究对象,设置森林固定监测样地,结合样地内物种组成数据,比较α多样性指数、β多样性指数和生物量等的差异,对于理解亚热带常绿阔叶林的特征以及探讨百山祖常绿阔叶林原生性、完整性和代表性具有重要意义。此外,通过比较百山祖国家公园与临近地区常绿阔叶林的群落结构和物种组成差异,对于理解常绿阔叶林的群落特征、演替动态、植被恢复等均具有理论指导意义。
-
钱江源-百山祖国家公园地处浙江省西南部,面积约754 km2。本研究选取百山祖园区作为研究区域,包含龙泉片区和庆元片区,是中亚热带常绿阔叶林生态系统的典型代表[14]。在百山祖国家公园庆元片区五岭坑(WLK)低海拔区域,分布有大面积以甜槠和木荷为优势种的常绿阔叶林,在龙泉片区凤阳山(FYS)的中海拔地带分布有大面积以木荷、褐叶青冈Cyclobalanopsis stewardiana和甜槠为优势种的常绿阔叶林。
为与临近区域分布的常绿阔叶林进行比较研究,选择浙江省内其他3个国家级自然保护区,分别为古田山国家级自然保护区(GTS)[15]、九龙山国家级自然保护区(JLS)[16]、乌岩岭国家级自然保护区(WYL)[17]以及非自然保护区(FZR)内的常绿阔叶林(表1)。以上研究区域的气候类型均属于中亚热带季风气候。
表 1 样地基本信息
Table 1. Basic information of sample plots
研究区域 样地名称 纬度(N) 经度(E) 样地数量/个 海拔/m 优势种 百山祖国家公园龙泉片区 凤阳山(FYS) 27.912º 119.184º 14 1051 ~1651 木荷、褐叶青冈、甜槠 百山祖国家公园庆元片区 五岭坑(WLK) 27.540º 119.064º 12 651~851 甜槠、木荷、米槠 古田山国家级自然保护区 古田山(GTS) 29.255º 118.130º 2 658~708 甜槠、木荷 九龙山国家级自然保护区 九龙山(JLS) 28.398º 118.841º 4 625~747 木荷、红楠 乌岩岭国家级自然保护区 乌岩岭(WYL) 27.713º 119.655º 6 960~ 1073 甜槠、木荷 非自然保护区 非自保护区(FZR) 27.560º 119.713º 12 381~871 甜槠、木荷、米槠 说明:木荷Schima superba,褐叶青冈Cyclobalanopsis stewardiana,甜槠Castanopsis eyrei,米槠Castanopsis carlesii,红楠Machilus thunbergia。 -
2012—2022年,在5个研究区域共设置50个大小为30 m×30 m的常绿阔叶林样地进行调查,每个研究地点所选样地信息见表1。参照美国热带森林研究中心(Center for Tropical Forest Science, CTFS)的方法[18],将每个样地划分成36个5 m×5 m小样方,调查样地内所有胸径(DBH)≥1 cm的木本植物个体,记录物种名、胸径、树高、分枝、空间坐标及生活状态等信息。同时测定样地内生境条件,包括海拔、坡度、坡向、郁闭度和土壤类型等。
-
根据《中国植物志》和《浙江植物志(新编)》[19]将调查到的物种分为乔木、灌木和小灌木。根据植物的生活型和DBH值将样地中调查到的所有个体划分为成树和幼树,其中乔木物种DBH>10 cm为成树,DBH≤10 cm为幼树;灌木物种DBH>5 cm为成树,DBH≤5 cm为幼树;小灌木物种DBH>2 cm为成树,DBH≤2 cm为幼树[20]。
-
分别计算了样地内所有木本植物、成树和幼树的α多样性指数,包括物种丰富度指数、Shannon-Wiener多样性指数、Simpson生态优势度指数[21]以及Chao多样性指数[22]。使用vegan包中的“diversity”和“estimate”等函数计算α多样性指数。
-
基于胸径的异速生长方程计算样地内所有木本植物物种(DBH≥1 cm)的生物量,每个个体通过累计其主干、枝、叶以及根的生物量获得个体总生物量。生物量(含地上和地下生物量)具体计算公式参考OUYANG等[23]在中国亚热带森林中构建的生物量与个体胸径间的异速生长方程。
-
为了确定不同样地的优势种和植被类型,计算样地内不同物种的重要值,依据重要值大小确定群落优势种和植被类型[24]。
利用方差分析(ANOVA)和最小显著差异法(LSD),分析不同样地之间的所有木本植物、成树和幼树的α多样性指数以及生物量是否具有显著差异。为了比较不同样地之间的物种组成差异,基于样地间的Bray-Curtis相异度指数的主坐标分析(PCoA),将样地内木本植物数据进行降维处理,使用vegan包中的“adonis”函数进行了999次置换方差分析(PERMANOVA),检验不同分组之间的物种组成是否存在显著差异。
-
在50个样地中共调查到
23021 株木本植物,隶属于57科128属304种。其中,常绿阔叶树种占比最高,共172种,隶属于32科60属;落叶阔叶树种共123种,隶属于40科75属;针叶树种占比最少,共3科7属9种。在百山祖国家公园中共调查到10 935株个体,隶属于52科108属241种,优势种为木荷与甜槠,其中成树多度占比34%,幼树多度占比66%。古田山自然保护区样地中优势种为甜槠和木荷,其中,成树多度占比23%,幼树多度占比77%。九龙山自然保护区样地中优势种为木荷和红楠,其中成树多度占比26%,幼树多度占比74%。乌岩岭自然保护区样地优势种为甜槠和木荷,其中成树多度占比24%,幼树多度占比76%。非自然保护区样地优势种为甜槠、木荷和米槠,成树多度占比32%,幼树多度占比68%。 -
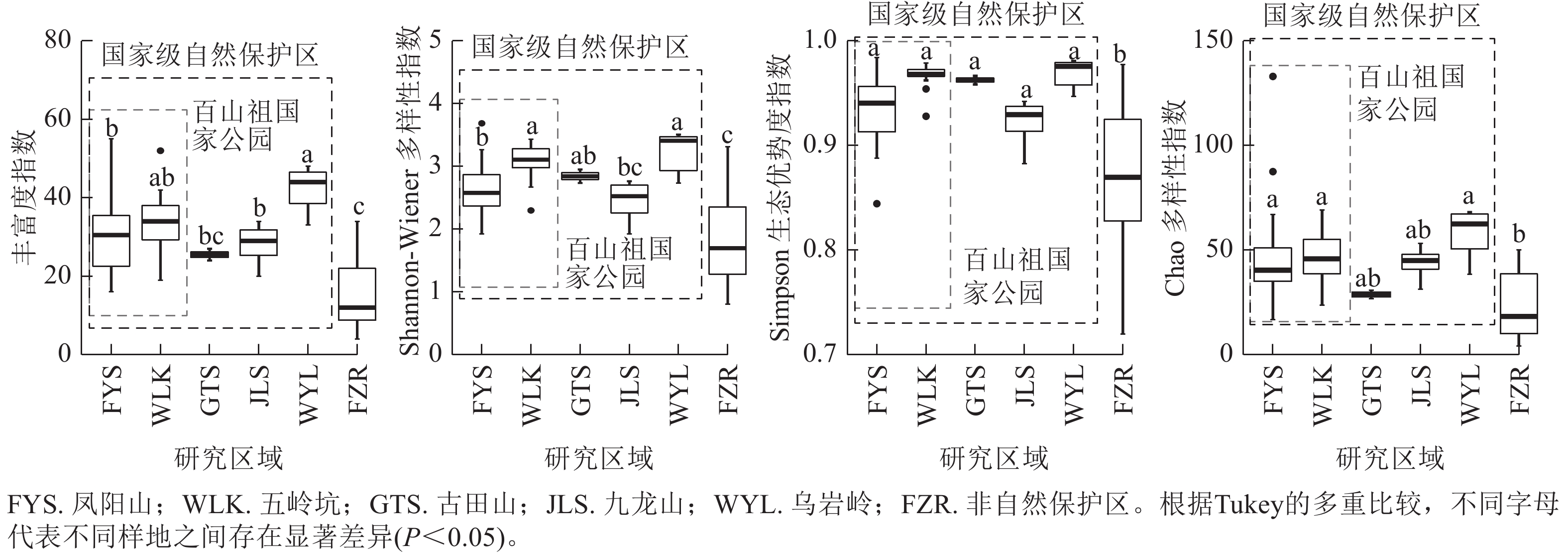
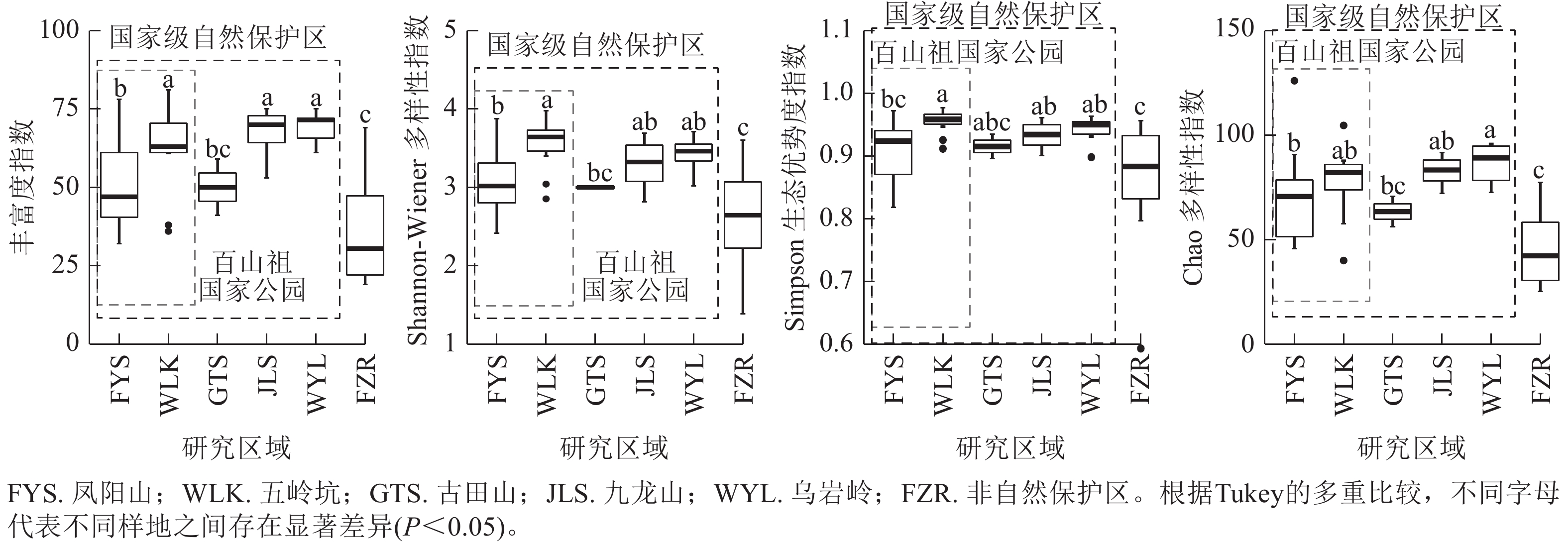
当考虑所有木本植物时,4种α多样性指数差异呈现相同趋势(图1)。百山祖国家公园五岭坑所有木本植物的物种丰富度指数、Shannon指数和Simpson指数都显著高于凤阳山(P<0.05),Chao指数无显著差异,但五岭坑与九龙山和乌岩岭所有木本植物的物种α多样性均无显著差异,凤阳山与古田山所有木本植物的物种α多样性均无显著差异。非自然保护区所有树种的物种α多样性显著低于百山祖国家公园、九龙山和乌岩岭(P<0.05),与古田山无显著差异。
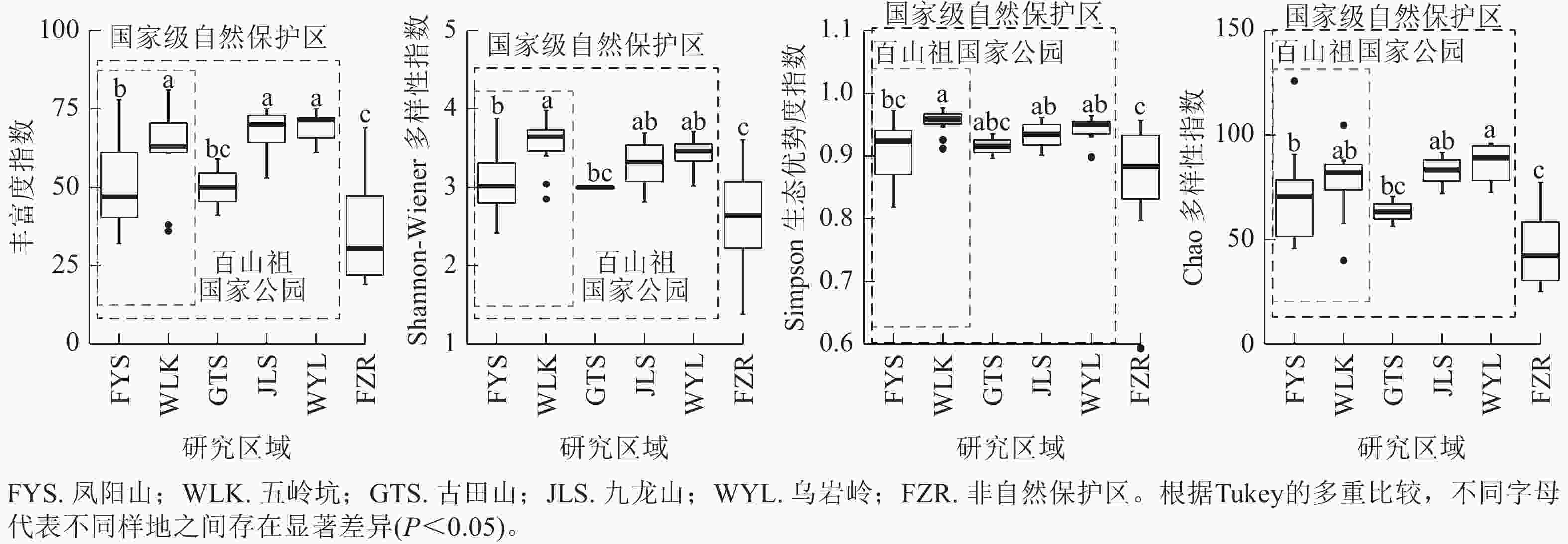
图 1 不同研究区域常绿阔叶林所有木本植物物种α多样性
Figure 1. Differences in species α richness of all woody plants in different plots
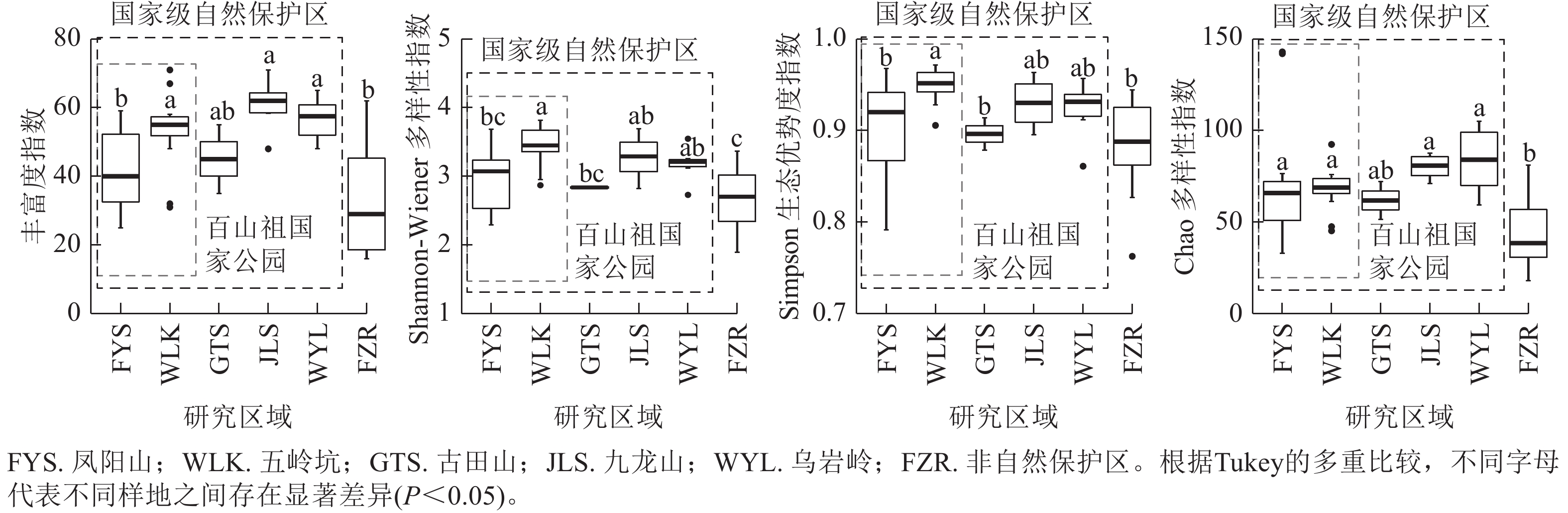
当考虑样地内成树的物种多样性时(图2),百山祖国家公园内五岭坑成树的Shannon指数显著高于凤阳山样地(P<0.05),而物种丰富度,Simpson指数和Chao指数无显著差异。五岭坑与古田山和乌岩岭样地成树的物种α多样性无显著差异。凤阳山成树的物种丰富度和Shannon指数显著低于乌岩岭(P<0.05),与古田山和九龙山无显著差异。非自然保护区成树的物种α多样性显著低于百山祖国家公园和乌岩岭(P<0.05),Simpson指数显著低于古田山和九龙山(P<0.05)。
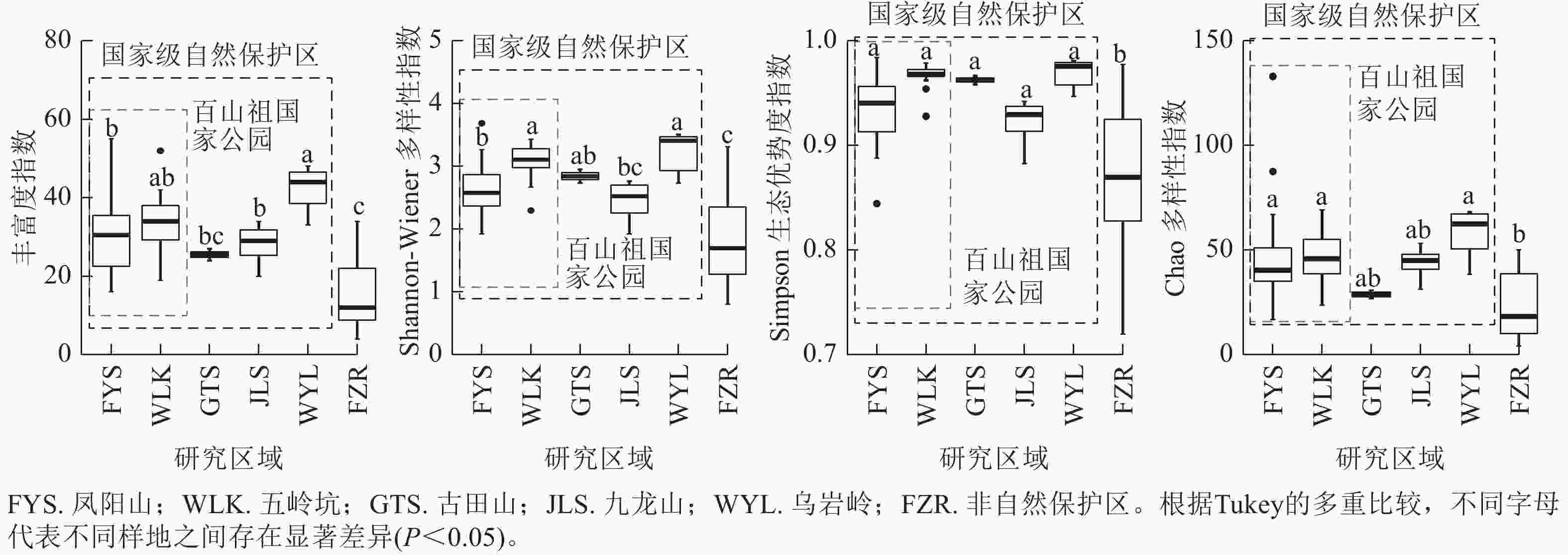
图 2 不同研究区域常绿阔叶林木本植物成树物种α多样性
Figure 2. Differences in species α richness of mature trees in different plots
当考虑样地内幼树的物种多样性时(图3),百山祖国家公园五岭坑幼树的物种丰富度指数,Shannon指数和Simpson指数都显著高于凤阳山样地(P<0.05),Chao指数无显著差异,但五岭坑与九龙山和乌岩岭幼树α多样性无显著差异。凤阳山,古田山和非自然保护区幼树的物种丰富度,Shannon指数和Simpson指数无显著差异。非自然保护区幼树的物种丰富度,Shannon指数和Chao指数显著低于五岭坑、九龙山和乌岩岭(P<0.05)。
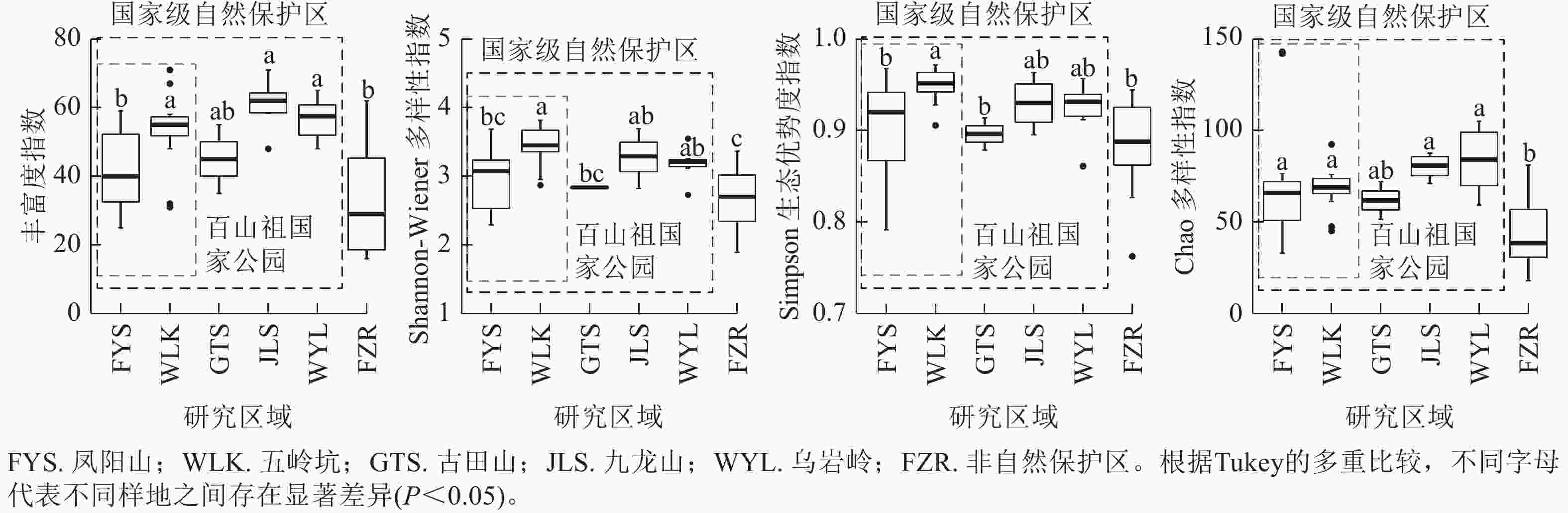
图 3 不同研究区域常绿阔叶林木本植物幼树物种α多样性
Figure 3. Differences in species α richness of saplings in different plots
-
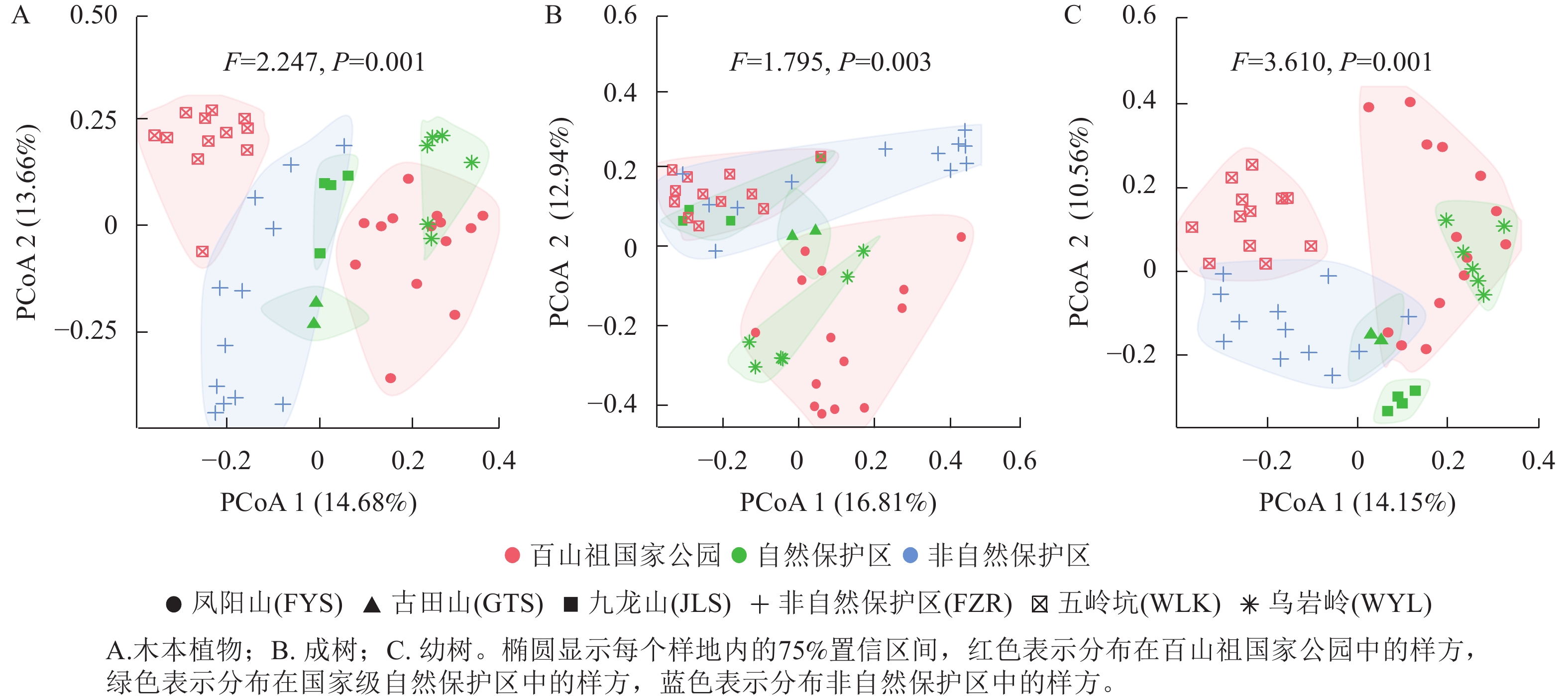
对于所有木本植物的物种组成,百山祖国家公园中凤阳山和五岭坑样地的物种组成存在显著差异(PERMANOVA检验:F=8.138,P=0.001),凤阳山的物种组成与古田山、九龙山和乌岩岭的更为相似,而五岭坑的物种组成与非自然保护区的物种组成更为相似(图4A和表2)。对于成树,凤阳山与乌岩岭的物种组成更相似,具有最低的Bray-Curtis指数值,凤阳山与五岭坑(F=7.261,P=0.001),九龙山以及非自然保护区(F=4.823,P=0.001)的物种组成存在显著差异(图4B和表2),而五岭坑、九龙山以及非自然保护区的物种组成更相似。凤阳山与乌岩岭的幼树物种组成相似,且五岭坑幼树物种与非自然保护区幼树物种组成更相似,具有最低的Bray-Curtis指数值(图4C和表2)。

图 4 不同研究区域常绿阔叶林木本植物物种组成的差异
Figure 4. Differences in species composition of woody plants among different plots
表 2 不同研究区域之间常绿阔叶林木本植物的Bray-Curtis值以及物种组成显著差异(PERMANOVA)的检验
Table 2. Bray-Curtis values of woody plants among different plots and the test of significant differences in species composition (PERMANOVA)
研究对象 项目 Bray-Curtis 平方和 R2 F P 所有木本植物 FYS/WLK 0.706 1.867 0.253 8.138 ≤0.001 FYS/WYL 0.467 0.729 0.149 3.153 0.002 FYS/FZR 0.648 1.436 0.182 5.347 ≤0.001 WLK/WYL 0.637 1.366 0.328 7.807 ≤0.001 WLK/FZR 0.582 1.433 0.220 6.204 ≤0.001 成树 FYS/WLK 0.771 1.904 0.232 7.261 ≤0.001 FYS/WYL 0.628 0.778 0.142 2.977 0.003 FYS/FZR 0.711 1.387 0.167 4.823 ≤0.001 WLK/WYL 0.646 1.285 0.272 5.986 ≤0.001 WLK/FZR 0.631 1.458 0.205 5.689 ≤0.001 幼树 FYS/WLK 0.685 1.656 0.218 6.700 ≤0.001 FYS/WYL 0.443 0.708 0.137 2.869 0.002 FYS/FZR 0.655 1.399 0.166 4.786 ≤0.001 WLK/WYL 0.653 1.321 0.295 6.683 ≤0.001 WLK/FZR 0.575 1.165 0.169 4.469 ≤0.001 说明:本表仅包含具有显著差异的结果。FYS. 凤阳山;WLK. 五岭坑;GTS. 古田山;JLS. 九龙山;WYL.乌岩岭;FZR. 非自然保护区。 -
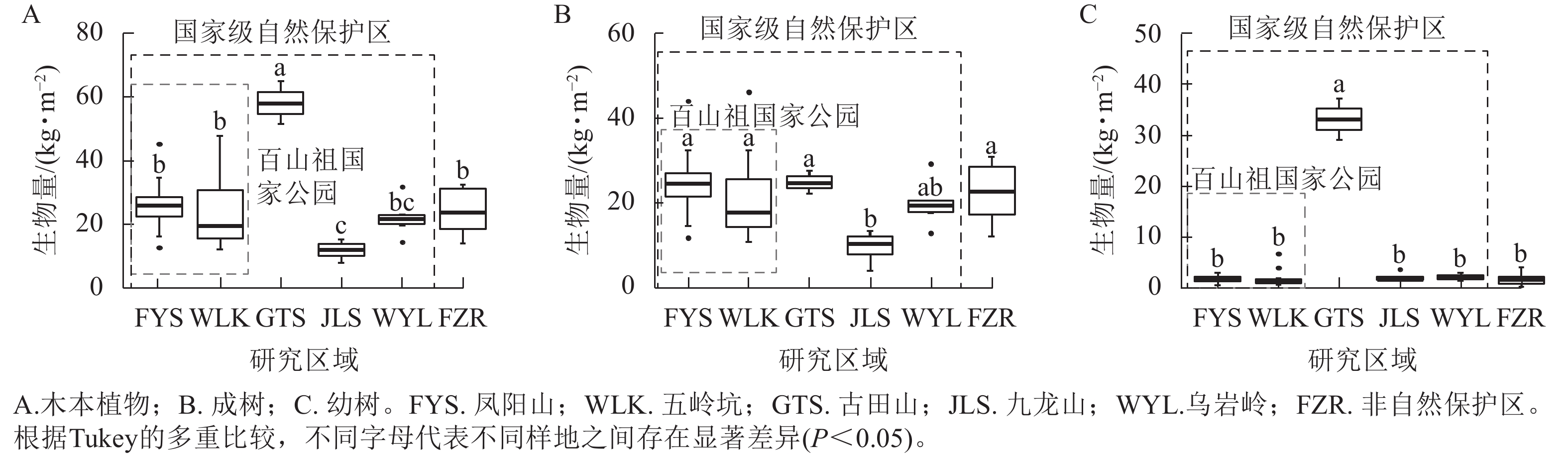
在考虑样地内所有木本植物的生物量时,百山祖国家公园内凤阳山、五岭坑,乌岩岭和非自然保护区之间的生物量无显著性差异,但均显著(P<0.05)低于古田山,高于九龙山(图5A)。凤阳山、五岭坑、古田山、乌岩岭以及非自然保护区之间成树的生物量都没有显著差异,且除乌岩岭外,均显著高于九龙山(图5B)。古田山具有最高的幼树生物量,显著(P<0.05)高于凤阳山、五岭坑、九龙山,乌岩岭和非自然保护区等其他研究区域(P<0.05),且其他样地之间的幼树生物量均无显著差异(图5C)。
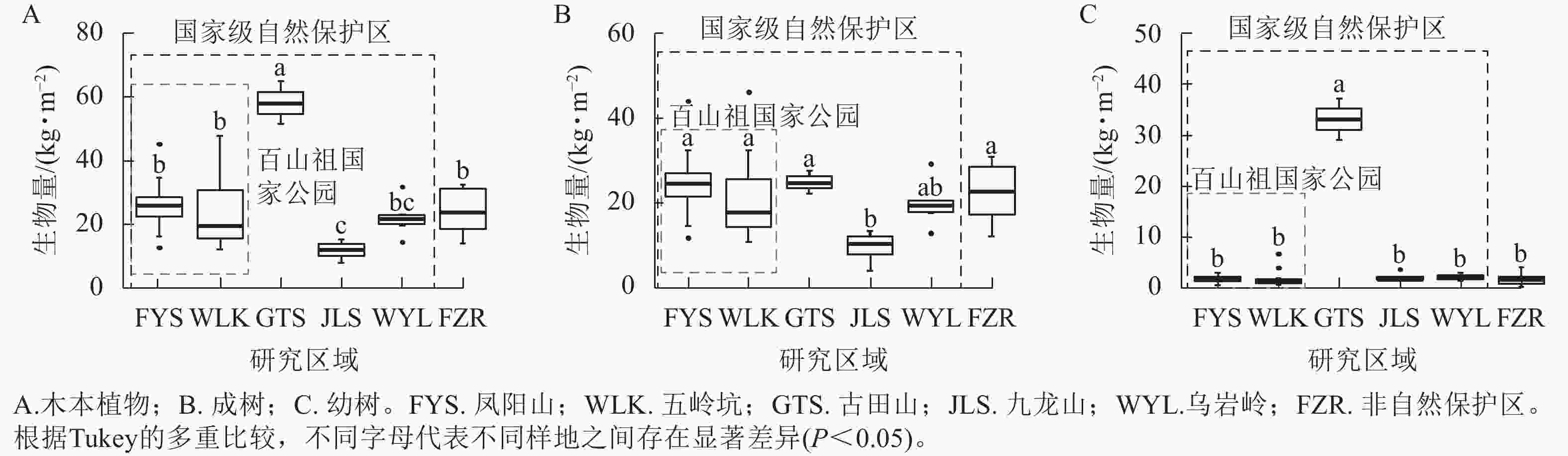
图 5 不同研究区域中木本植物的生物量差异
Figure 5. Differences in biomass of woody plants among different plots
-
中国具有世界上分布最广、类型最为丰富的亚热带森林,其结构复杂,物种丰富,提供了稳定的生态系统服务价值。本研究系统分析并比较了中国亚热带地区百山祖国家公园内常绿阔叶林与其他自然保护区和非自然保护区内常绿阔叶林的群落特征。研究发现百山祖国家公园内五岭坑所有木本植物物种α多样性显著高于凤阳山,这与田磊等[25]的研究结果一致,可能与凤阳山和五岭坑所处的海拔差异有关。海拔通过影响气温和降水等影响物种α多样性,但物种α多样性随海拔的变化关系尚缺乏统一的格局[26−27]。林阳等[28]发现:在百山祖国家公园内不同海拔梯度的木本植物物种α多样性与海拔梯度呈显著负相关。随着海拔的升高,气温降低,常绿阔叶树种的生长受到限制,这可能导致高海拔凤阳山比低海拔五岭坑的物种α多样性低。进一步通过对比分析不同生长型幼树和成树物种α多样性的差异,发现五岭坑幼树的物种α多样性显著高于凤阳山,但2个样地内成树的物种α多样性没有显著差异。这说明低海拔区域幼树的物种多样性显著高于高海拔区域。与成树相比,幼树的物种α多样性可能更容易受到生物和非生物因子的影响[29]。五岭坑样地处于较低海拔范围,与高海拔的凤阳山比较,五岭坑内群落生境更加稳定,可能更有利于幼树的更新和生长[30]。
“中间膨胀效应”假说认为不同物种的分布范围相互重叠,但是由于边界限制,使得不同物种的分布范围在边界处重叠小,在中心地区重叠大[31]。在百山祖国家公园和选择的自然保护区中,处于中间海拔梯度的五岭坑样地、乌岩岭样地以及九龙山样地内,总体上物种α多样性都处于较高水平,符合这一假说,从而导致处于中间海拔样地的物种丰富度更高,多项研究也证实了这一结论[32]。
另外,非自然保护区的物种α多样性显著低于自然保护区,可能与人为干扰对物种多样性的影响有关[33]。通常自然保护区限制人为活动(如砍伐或择伐),同时也对自然保护区内的物种进行特定的保护,因而具有更高的物种α多样性。
-
物种组成差异能够反映群落间的异质性,物种组成相似度越高,群落间异质性和群落生境条件的异质性越小[34]。植物群落所处的生境条件、气候因子以及区域物种库等不同均会影响群落间物种组成的差异。如海拔可以通过影响温度和水分改变群落环境,进而影响群落内物种组成[35]。本研究发现:百山祖国家公园内凤阳山和五岭坑的常绿阔叶林中所有木本植物、成树和幼树的物种组成均存在显著差异,可能与2个区域所处的海拔不同有关。随海拔变化,不同物种对温度和水分等环境因子的耐受性不同,在海拔梯度上的分布范围也会不同[36]。如五岭坑样地主要分布在低海拔区域(海拔为651~851 m),乔木层优势种主要为甜槠和木荷,样地内木本植物的叶生活型以常绿阔叶树种为主(常绿阔叶树种122种、落叶树种39种和针叶树种2种),而处于高海拔的凤阳山样地(海拔为
1051 ~1651 m),常绿阔叶树种中的青冈类和杜鹃类以及落叶树种和针叶树种的种类比例增加(常绿阔叶树种97种、落叶树种69种和针叶树种7种)。同时,除了海拔对物种组成的影响外,纬度梯度也会对物种组成产生影响[37]。李林等[38]发现相近纬度和海拔上的物种组成更相似。本研究中凤阳山和乌岩岭所处海拔和纬度更接近,2个样地所有木本植物、成树和幼树物种组成更加相似,五岭坑与非自然保护区海拔和纬度相近,物种组成也更相似。 -
生物量是衡量森林生态系统生产力的重要指标,同时也是评估森林碳汇的重要参数[39]。在森林生态系统中,气候、土壤理化性质和地形等非生物因子[40]以及群落的演替历史和物种组成等生物因子[41]都是影响生物量的重要因素。如森林生物量随年均气温和年均降水的提高而逐渐升高[42],处于演替前期的次生林相比于演替后期原生林或老龄林具有更低的生物量[43]。本研究中,百山祖国家公园内的凤阳山和五岭坑区域,虽然海拔上存在差异,但2个区域间所有木本植物、成树和幼树的生物量均无显著差异,说明环境因子虽然对2个区域的物种组成和物种多样性产生了较大的影响,但对于生物量并无显著影响。本研究中古田山所有木本植物和幼树的生物量显著高于其他样地,其他各区域间幼树的生物无显著差异。结合样地中不同生长型的树种多度发现:虽然样地中幼树的多度占比均显著高于成树,但古田山幼树多度占比最高。此外,样地内壳斗科植物个体的生物量普遍高于其他物种,可能使壳斗科植物占比高的群落生物量更高。进一步分析各样地幼树中壳斗科个体多度占比发现,古田山壳斗科幼树多度占比高于凤阳山、五岭坑、九龙山、乌岩岭和非自然保护区。因此,古田山具有更高的幼树生物量,也可能与样地内壳斗科物种个体占比较高有关。本研究结果也说明物种组成是影响生物量的主要原因之一。为更全面反映百山祖国家公园与邻近地区常绿阔叶林群落特征比较,后续研究应增加研究样地的数量和分布范围。
-
百山祖国家公园内凤阳山和五岭坑常绿阔叶林群落的物种多样性和物种组成存在显著差异,但生物量并无显著差异。相比于其他区域的常绿阔叶林,五岭坑常绿阔叶林具有更高的物种多样性。同时,国家公园和自然保护区内分布的常绿阔叶林物种多样性显著高于非自然保护区。在物种组成方面,五岭坑常绿阔叶林与同纬度的非自然保护区常绿阔叶林物种组成更为相似,而凤阳山则与乌岩岭常绿阔叶林的物种组成更相似。本研究结果表明:受海拔、纬度、群落演替历史和人类干扰等的影响,百山祖国家公园内分布的低海拔和中山地带常绿阔叶林中的物种多样性、物种组成和生态系统功能等不仅在公园内存在差异,尤其在五岭坑分布的以甜槠-木荷为优势种的典型常绿阔叶林和中山地带分布的以褐叶青冈等为优势种的山地常绿阔叶林,也与亚热带其他地区常绿阔叶林存在差异,说明该公园内保存的亚热带常绿阔叶林具有一定的独特性,具有较高的保护价值。
-
感谢浙江大学毛志斌、韦博良,浙江师范大学林阳,中国计量大学杨中杰,华东师范大学李时轩,温州大学刘维勇、邓文婕、刘腾腾、税章利和惠城阳等,以及赖正林和姜淦冰等人参与野外调查工作。
Differences in community characteristics of evergreen broad-leaved forests between Baishanzu National Park and adjacent areas
-
摘要:
目的 百山祖国家公园具有典型且完整的亚热带森林生态系统,垂直带谱完整,原生常绿阔叶林面积分布广泛。通过比较分析百山祖国家公园常绿阔叶林与邻近其他自然保护区和非自然保护区内分布的常绿阔叶林的群落结构和物种多样性,有助于理解百山祖常绿阔叶林群落特征,为亚热带常绿阔叶林保护和退化植被生态恢复策略提供科学依据。 方法 选择百山祖国家公园(凤阳山和五岭坑)、九龙山、乌岩岭和古田山等4个国家级自然保护区和非自然保护区内的常绿阔叶林为研究对象,设置50个30 m×30 m的森林固定监测样地,针对样地内所有胸径≥1 cm的木本植物,利用方差分析、基于Bray-Curtis相异度指数的主坐标分析和置换方差分析等方法,比较各地区常绿阔叶林中所有木本植物成树和幼树的物种α多样性差异、物种组成差异和生物量差异。 结果 ①50个样地中共发现57科128属304种木本植物;其中百山祖国家公园的样地内有52科108属241种,优势种为木荷Schima superba和甜槠Castanopsis eyrei。②五岭坑的所有木本植物和幼树的物种α多样性最高,且国家公园和自然保护区的所有木本植物、成树和幼树的物种α多样性都高于非自然保护区。③五岭坑和凤阳山之间的所有木本植物、成树和幼树的物种组成均存在显著差异,且与其他地区的物种组成均存在差异。④凤阳山、五岭坑及乌岩岭和非保护区之间所有木本植物、成树或幼树的生物量均无显著差异,但五岭坑和凤阳山的幼树生物量显著低于古田山(P<0.05)。 结论 百山祖国家公园常绿阔叶林中的物种组成和多样性、生态系统功能等不仅在公园内不同海拔区域中存在差异,也与亚热带其他地区常绿阔叶林存在差异,说明该公园内的常绿阔叶林具有一定的独特性。图5表2参43 Abstract:Objective Baishanzu National Park has a typical and complete subtropical forest ecosystem with a comprehensive vertical spectrum and extensive primary evergreen broad-leaved forest. By comparing the community structure and species diversity of the evergreen broad-leaved forest in Baishanzu National Park with those in other national nature reserves and non-protected areas in Zhejiang Province, this study aims to understand the community characteristics of the evergreen broad-leaved forest in Baishanzu National Park and provide a theoretical basis for the protection of subtropical native evergreen broad-leaved forest and the restoration strategy of degraded vegetation. Method This study selected evergreen broad-leaved forests in Baishanzu National Park (including Fengyangshan and Wulingkeng), Jiulongshan, Wuyanling, and Gutianshan National Nature Reserve, as well as a non-nature reserve area as the research sites. A total of 50 plots of 30 m×30 m were set up. By calculating the species diversity index and biomass of all woody plants, mature trees and saplings with DBH ≥1 cm in these plots, through methods including Analysis of Variance (ANOVA), Principal Coordinate Analysis (PCoA) based on the Bray-Curtis dissimilarity index, and Permutational Multivariate Analysis of Variance (PERMANOVA), we compared and analyzed the species α diversity, species composition and biomass differences of all woody plants, mature trees and saplings in evergreen broad-leaved forest in different sites. Result (1) A total of 304 species belonging to 128 genera in 57 families were found in 50 plots, among which 241 species belonging to 108 genera in 52 families were found in Baishanzu National Park. Schima superba and Castanopsis eyrei were the dominant species. (2) The species α diversity of all woody plants and saplings was highest in Wulingkeng of Baishanzu National Park. Additionally, the species α diversity of all woody plants, mature trees, and saplings in national parks and nature reserves were higher than that in the non-nature reserves area. (3) Significant difference in species composition of all woody plants, mature trees and saplings were observed between Wulingkeng and Fengyangshan of Baishanzu National Park, as well as between other sites. (4) No significant difference in biomass of woody plants, mature trees, and saplings was found among Fengyangshan, Wulingkeng, Wuyanling and the non-nature reserves area. However, sapling biomass in Wulingkeng and Fengyangshan was significantly lower than that in Gutianshan. Conclusion The species diversity, species composition, and ecosystem function of the evergreen broad-leaved forest in Baishanzu National Park differ not only among different areas in the same national park but also with the other subtropical regions. This suggests that the original evergreen broad-leaved forests in Baishanzu National Park exhibit a distinct uniqueness. [Ch, 5 fig. 2 tab. 43 ref.] -
Key words:
- Baishanzu National Park /
- broad-leaved evergreen forest /
- species diversity /
- biomass
-
图 1 不同研究区域常绿阔叶林所有木本植物物种α多样性
Figure 1 Differences in species α richness of all woody plants in different plots
图 2 不同研究区域常绿阔叶林木本植物成树物种α多样性
Figure 2 Differences in species α richness of mature trees in different plots
图 3 不同研究区域常绿阔叶林木本植物幼树物种α多样性
Figure 3 Differences in species α richness of saplings in different plots
图 4 不同研究区域常绿阔叶林木本植物物种组成的差异
Figure 4 Differences in species composition of woody plants among different plots
图 5 不同研究区域中木本植物的生物量差异
Figure 5 Differences in biomass of woody plants among different plots
表 1 样地基本信息
Table 1. Basic information of sample plots
研究区域 样地名称 纬度(N) 经度(E) 样地数量/个 海拔/m 优势种 百山祖国家公园龙泉片区 凤阳山(FYS) 27.912º 119.184º 14 1051 ~1651 木荷、褐叶青冈、甜槠 百山祖国家公园庆元片区 五岭坑(WLK) 27.540º 119.064º 12 651~851 甜槠、木荷、米槠 古田山国家级自然保护区 古田山(GTS) 29.255º 118.130º 2 658~708 甜槠、木荷 九龙山国家级自然保护区 九龙山(JLS) 28.398º 118.841º 4 625~747 木荷、红楠 乌岩岭国家级自然保护区 乌岩岭(WYL) 27.713º 119.655º 6 960~ 1073 甜槠、木荷 非自然保护区 非自保护区(FZR) 27.560º 119.713º 12 381~871 甜槠、木荷、米槠 说明:木荷Schima superba,褐叶青冈Cyclobalanopsis stewardiana,甜槠Castanopsis eyrei,米槠Castanopsis carlesii,红楠Machilus thunbergia。  下载: 导出CSV
下载: 导出CSV
表 2 不同研究区域之间常绿阔叶林木本植物的Bray-Curtis值以及物种组成显著差异(PERMANOVA)的检验
Table 2. Bray-Curtis values of woody plants among different plots and the test of significant differences in species composition (PERMANOVA)
研究对象 项目 Bray-Curtis 平方和 R2 F P 所有木本植物 FYS/WLK 0.706 1.867 0.253 8.138 ≤0.001 FYS/WYL 0.467 0.729 0.149 3.153 0.002 FYS/FZR 0.648 1.436 0.182 5.347 ≤0.001 WLK/WYL 0.637 1.366 0.328 7.807 ≤0.001 WLK/FZR 0.582 1.433 0.220 6.204 ≤0.001 成树 FYS/WLK 0.771 1.904 0.232 7.261 ≤0.001 FYS/WYL 0.628 0.778 0.142 2.977 0.003 FYS/FZR 0.711 1.387 0.167 4.823 ≤0.001 WLK/WYL 0.646 1.285 0.272 5.986 ≤0.001 WLK/FZR 0.631 1.458 0.205 5.689 ≤0.001 幼树 FYS/WLK 0.685 1.656 0.218 6.700 ≤0.001 FYS/WYL 0.443 0.708 0.137 2.869 0.002 FYS/FZR 0.655 1.399 0.166 4.786 ≤0.001 WLK/WYL 0.653 1.321 0.295 6.683 ≤0.001 WLK/FZR 0.575 1.165 0.169 4.469 ≤0.001 说明:本表仅包含具有显著差异的结果。FYS. 凤阳山;WLK. 五岭坑;GTS. 古田山;JLS. 九龙山;WYL.乌岩岭;FZR. 非自然保护区。
下载: 导出CSV
-
[1] 蔡焕满, 吴素美, 吴逸卿, 等. 百山祖国家公园五岭坑常绿阔叶林甜槠的种群特征[J]. 浙江林业科技, 2024, 44(1): 1−7. CAI Huanman, WU Sumei, WU Yiqing, et al. The population characteristics of Castanopsis eyrei in evergreen broad-leaved forest in Wulingkeng of Baishanzu National Park [J]. Journal of Zhejiang Forestry Science and Technology, 2024, 44(1): 1−7. [2] 宋永昌. 中国常绿阔叶林: 分类·生态·保育[M]. 北京: 科学出版社, 2013. SONG Yongchang. Evergreen Broad-leaved Forests in China: Classification, Ecology, Conservation [M]. Beijing: Science Press, 2013. [3] HUANG Caishuang, XU Yue, ZANG Runguo. Low functional redundancy revealed high vulnerability of the subtropical evergreen broadleaved forests to environmental change [J/OL]. Science of the Total Environment, 2024, 935 : 173307[2024-07-01]. DOI: 10.1016/j.scitotenv.2024.173307. [4] 余秋伍, 杨菁, 沈国春. 浙江天童常绿阔叶林林冠结构与群落物种组成的关系[J]. 植物生态学报, 2022, 46(5): 529−538. YU Qiuwu, YANG Jing, SHEN Guochun. Relationship between canopy structure and species composition of an evergreen broadleaf forest in Tiantong region, Zhejiang, China [J]. Chinese Journal of Plant Ecology, 2022, 46(5): 529−538. [5] FANG Xiaofeng, SHEN Guochun, YANG Qingsong, et al. Habitat heterogeneity explains mosaics of evergreen and deciduous trees at local-scales in a subtropical evergreen broad-leaved forest [J]. Journal of Vegetation Science, 2017, 28(2): 379−388. [6] JIN Dongmei, YUAN Quan, DAI Xiling, et al. Enhanced precipitation has driven the evolution of subtropical evergreen broad-leaved forests in Eastern China since the early Miocene: evidence from ring-cupped oaks [J]. Journal of Systematics and Evolution, 2024, 62(4): 677−686. [7] 金毅, 陈建华, 米湘成, 等. 古田山24 ha森林动态监测样地常绿阔叶林群落结构和组成动态: 探讨2008年冰雪灾害的影响[J]. 生物多样性, 2015, 23(5): 610−618. JIN Yi, CHEN Jianhua, MI Xiangcheng, et al. Impacts of the 2008 ice storm on structure and composition of an evergreen broad-leaved forest community in Eastern China [J]. Biodiversity Science, 2015, 23(5): 610−618. [8] 米湘成, 王绪高, 沈国春, 等. 中国森林生物多样性监测网络: 二十年群落构建机制探索的回顾与展望[J]. 生物多样性, 2022, 30(10): 211−233. MI Xiangcheng, WANG Xugao, SHEN Guochun, et al. Chinese forest biodiversity monitoring network(CForBio): twenty years of exploring community assembly mechanisms and prospects for future research [J]. Biodiversity Science, 2022, 30(10): 211−233. [9] TONG Xin, NASON J D, DING Yuanyuan, et al. Genetic tracking of density-dependent adult recruitment: a case study in a subtropical oak [J]. Journal of Ecology, 2021, 109(6): 2317−2328. [10] 张田田, 王璇, 任海保, 等. 浙江古田山次生与老龄常绿阔叶林群落特征的比较[J]. 生物多样性, 2019, 27(10): 1069−1080. ZHANG Tiantian, WANG Xuan, REN Haibao, et al. A comparative study on the community characteristics of secondary and old-growth evergreen broad-leaved forests in Gutianshan, Zhejiang Province [J]. Biodiversity Science, 2019, 27(10): 1069−1080. [11] 宋永昌, 阎恩荣, 宋坤. 中国常绿阔叶林8大动态监测样地植被的综合比较[J]. 生物多样性, 2015, 23(2): 139−148. SONG Yongchang, YAN Enrong, SONG Kun. Synthetic comparison of eight dynamics plots in evergreen broadleaf forests, China [J]. Biodiversity Science, 2015, 23(2): 139−148. [12] 刘啸林, 吴友贵, 张敏华, 等. 浙江百山祖25 ha亚热带森林动态监测样地群落组成与结构特征[J]. 生物多样性, 2024, 32(2): 25−35. LIU Xiaolin, WU Yougui, ZHANG Minhua, et al. Community composition and structure of a 25 ha forest dynamics plot of subtropical forest in Baishanzu, Zhejiang Province [J]. Biodiversity Science, 2024, 32(2): 25−35. [13] ZHANG Yun, LI Xinke, KONG Zhaochen, et al. Subtropical forest vegetation development and climate change in Baishanzu area of Zhejiang Province, China, since the Holocene [J/OL]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2022, 608 : 111293[2024-07-01]. DOI: 10.1016/j.palaeo.2022.111293. [14] 米湘成, 余建平, 王宁宁, 等. 基于激光雷达技术估算钱江源国家公园森林的地上生物量[J]. 北京林业大学学报, 2022, 44(10): 77−84. MI Xiangcheng, YU Jianping, WANG Ningning, et al. Utilizing LiDAR technology to estimate forest aboveground biomass in Qianjiangyuan National Park, Jiangxi Province of Eastern China [J]. Journal of Beijing Forestry University, 2022, 44(10): 77−84. [15] 寿佳君, 薛乾怀, 王鑫洋, 等. 浙江古田山亚热带常绿阔叶林叶衰老物候影响因子研究[J]. 热带亚热带植物学报, 2023, 31(2): 181−191. SHOU Jiajun, XUE Qianhuai, WANG Xinyang, et al. Studies on affecting factors of leaf senescence phenology in a subtropical evergreen broad-leaved forest in Gutianshan, Zhejiang Province [J]. Journal of Tropical and Subtropical Botany, 2023, 31(2): 181−191. [16] 刘菊莲, 韦博良, 吴雁南, 等. 浙江九龙山常绿阔叶林不同物种的径级结构及空间关联[J]. 浙江农林大学学报, 2023, 40(3): 598−607. LIU Julian, WEI Boliang, WU Yannan, et al. Size structure and spatial association of different species of an evergreen broad-leaved forest in Jiulong Mountain, Zhejiang Province [J]. Journal of Zhejiang A&F University, 2023, 40(3): 598−607. [17] 戴尊, 陈星, 张建行, 等. 浙江乌岩岭国家级自然保护区叶附生苔类及附主植物多样性[J]. 生物多样性, 2022, 30(1): 124−131. DAI Zun, CHEN Xing, ZHANG Jianhang, et al. Species diversity of epiphyllous liverworts and host plants in the Wuyanling National Nature Reserve, Zhejiang Province [J]. Biodiversity Science, 2022, 30(1): 124−131. [18] CONDIT R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama, and a Comparison with other Plots [M]. Berlin: Springer, 1998. [19] 方精云, 郭柯, 王国宏, 等. 《中国植被志》的植被分类系统、植被类型划分及编排体系[J]. 植物生态学报, 2020, 44(2): 96−110. FANG Jingyun, GUO Ke, WANG Guohong, et al. Vegetation classification system and classification of vegetation types used for the compilation of vegetation of China [J]. Chinese Journal of Plant Ecology, 2020, 44(2): 96−110. [20] LIU Jinliang, ZHONG Yuchen, ZHONG Lei, et al. The asymmetric relationships of the distribution of conspecific saplings and adults in forest fragments [J]. Journal of Plant Ecology, 2020, 13(4): 398−404. [21] WEI Shiguang, LI Lin, BAI Kundong, et al. Community structure and species diversity dynamics of a subtropical evergreen broad-leaved forest in China: 2005 to 2020 [J]. Plant Diversity, 2024, 46(1): 70−77. [22] CHIU C H, WANG Yiting, WALTHER B A, et al. An improved nonparametric lower bound of species richness via a modified good-turing frequency formula [J]. Biometrics, 2014, 70(3): 671−682. [23] OUYANG Shuai, XIANG Wenhua, WANG Xiangping, et al. Significant effects of biodiversity on forest biomass during the succession of subtropical forest in South China [J]. Forest Ecology and Management, 2016, 372: 291−302. [24] 邓文婕, 吴华征, 李添翔, 等. 洞头国家级海洋公园主要植被类型及其特征[J]. 植物生态学报, 2024, 48(2): 254−268. DENG Wenjie, WU Huazheng, LI Tianxiang, et al. Main vegetation types and characteristics in Dongtou national marine park, Zhejiang, China [J]. Chinese Journal of Plant Ecology, 2024, 48(2): 254−268. [25] 田磊, 于明坚, 陈建华, 等. 基于样方法的浙江省11个自然保护区木本植物区系成分分析[J]. 浙江大学学报(理学版), 2015, 42(1): 28−37. TIAN Lei, YU Mingjian, CHEN Jianhua, et al. Analysis of woody flora based on quadrat method in eleven natural reserves of Zhejiang Province [J]. Journal of Zhejiang University (Science Edition), 2015, 42(1): 28−37. [26] ZHANG Wenxin, HUANG Dizhou, WANG Renqing, et al. Altitudinal patterns of species diversity and phylogenetic diversity across temperate mountain forests of Northern China [J/OL]. PLoS One, 2016, 11 (7): e0159995[2024-07-01]. DOI: 10.1371/journal.pone.0159995. [27] GALVÁN-CISNEROS C M, VILLA P M, COELHO A J P, et al. Altitude as environmental filtering influencing phylogenetic diversity and species richness of plants in tropical mountains [J]. Journal of Mountain Science, 2023, 20(2): 285−298. [28] 林阳, 李时轩, 周伟龙, 等. 百山祖国家公园植物群落α和β多样性对海拔梯度的响应[J]. 生态学报, 2024, 44(17): 7700−7712. LIN Yang, LI Shixuan, ZHOU Weilong, et al. α and β diversity patterns of woody plant communities along an elevation gradient in Baishanzu National Park [J]. Acta Ecologica Sinica, 2024, 44(17): 7700−7712. [29] 李大标, 钟毓萍, 龚笑飞, 等. 浙江遂昌马尾松林物种和谱系β多样性驱动因子分析[J]. 广西植物, 2023, 43(7): 1258−1267. LI Dabiao, ZHONG Yuping, GONG Xiaofei, et al. Analysis of species and phylogenetic β diversity drivers in the Masson pine forests in Suichang, Zhejiang Province [J]. Guihaia, 2023, 43(7): 1258−1267. [30] 周荣飞, 吴义松, 蔡焕满, 等. 浙江五岭坑原生常绿阔叶林群落的动态特征[J]. 浙江林业科技, 2022, 42(5): 1−8. ZHOU Rongfei, WU Yisong, CAI Huanman, et al. Dynamic characteristics of primary evergreen broad-leaved forest in wulingkeng mountain, Zhejiang Province [J]. Journal of Zhejiang Forestry Science and Technology, 2022, 42(5): 1−8. [31] COLWELL R K, LEES D C. The mid-domain effect: geometric constraints on the geography of species richness [J]. Trends in Ecology & Evolution, 2000, 15(2): 70−76. [32] 刘秉儒. 生物多样性的海拔分布格局研究及进展[J]. 生态环境学报, 2021, 30(2): 438−444. LIU Bingru. Recent advances in altitudinal distribution patterns of biodiversity [J]. Ecology and Environmental Sciences, 2021, 30(2): 438−444. [33] LI Chaojun, LOU Hezhen, YANG Shengtian, et al. Effect of human disturbances and hydrologic elements on the distribution of plant diversity within the Shamu watershed, Mt. Yuntai Nature Reserve, China [J/OL]. Journal of Environmental Management, 2022, 311 : 114833[2024-07-01]. DOI: 10.1016/j.jenvman.2022.114833. [34] 隆卫革. 桂南不同造林模式对植物物种组成和多样性特征的影响[J]. 安徽农业科学, 2024, 52(8): 106−109. LONG Weige. Effects of different afforestation modes on characteristics of plant composition and diversity at South Guangxi [J]. Journal of Anhui Agricultural Sciences, 2024, 52(8): 106−109. [35] LUO Yusheng, ZHOU Mengli, JIN Shanshan, et al. Changes in phylogenetic structure and species composition of woody plant communities across an elevational gradient in the southern Taihang Mountains, China [J/OL]. Global Ecology and Conservation, 2023, 42 : e02412[2024-07-01]. DOI: 10.1016/j.gecco.2023.e02412. [36] 何远政, 黄文达, 赵昕, 等. 气候变化对植物多样性的影响研究综述[J]. 中国沙漠, 2021, 41(1): 59−66. HE Yuanzheng, HUANG Wenda, ZHAO Xin, et al. Review on the impact of climate change on plant diversity [J]. Journal of Desert Research, 2021, 41(1): 59−66. [37] NISHIZAWA K, SHINOHARA N, CADOTTE M W, et al. The latitudinal gradient in plant community assembly processes: a meta-analysis [J]. Ecology Letters, 2022, 25(7): 1711−1724. [38] 李林, 魏识广, 练琚愉, 等. 亚热带不同纬度植物群落物种多样性分布规律[J]. 生态学报, 2020, 40(4): 1249−1257. LI Lin, WEI Shiguang, LIAN Juyu, et al. Distributional regularity of species diversity in plant community at different latitudes in subtropics [J]. Acta Ecologica Sinica, 2020, 40(4): 1249−1257. [39] 汤明华, 刘娟, 高林, 等. 基于森林资源清查资料的盈江县森林生物量和生长量分析[J]. 西部林业科学, 2024, 53(1): 129−137. TANG Minghua, LIU Juan, GAO Lin, et al. Forest biomass and growth in Yingjiang County based on forest resource inventory data [J]. Journal of West China Forestry Science, 2024, 53(1): 129−137. [40] 窦啸文, 吴登瑜, 张笑菁, 等. 天目山常绿阔叶林胸高断面积生长量影响因子研究[J]. 浙江农林大学学报, 2023, 40(5): 1063−1072. DOU Xiaowen, WU Dengyu, ZHANG Xiaojing, et al. Study on the factors affecting breast-height basal area increment of evergreen broad-leaved forest in Mount Tianmu [J]. Journal of Zhejiang A&F University, 2023, 40(5): 1063−1072. [41] 朱杰, 吴安驰, 邹顺, 等. 南亚热带常绿阔叶林树木多样性与生物量和生产力的关联及其影响因素[J]. 生物多样性, 2021, 29(11): 1435−1446. ZHU Jie, WU Anchi, ZOU Shun, et al. Relationships between tree diversity and biomass/productivity and their influence factors in a lower subtropical evergreen broad-leaved forest [J]. Biodiversity Science, 2021, 29(11): 1435−1446. [42] 杨远盛, 张晓霞, 于海艳, 等. 中国森林生物量的空间分布及其影响因素[J]. 西南林业大学学报, 2015, 35(6): 45−52. YANG Yuansheng, ZHANG Xiaoxia, YU Haiyan, et al. The spatial distribution of China’s forest biomass and its influencing factors [J]. Journal of Southwest Forestry University, 2015, 35(6): 45−52. [43] 陈彭祯霓, 任亮晶, 罗辑, 等. 海螺沟冰川退缩区原生演替序列植被物种多样性与生物量的关系[J]. 应用与环境生物学报, 2022, 28(5): 1129−1136. CHEN Pengzhenni, REN Liangjing, LUO Ji, et al. Relationship between biodiversity and biomass along the primary succession chronosequence in the foreland of the Hailuogou Glacier [J]. Chinese Journal of Applied and Environmental Biology, 2022, 28(5): 1129−1136. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240456
 点击查看大图
点击查看大图
计量
- 文章访问数: 1579
- HTML全文浏览量: 448
- PDF下载量: 78
- 被引次数: 0
