




-
随着人类活动和工业化程度不断加剧,大气环境中二氧化碳浓度增加,导致全球气温升高和降水模式转变,引起了严重且频繁的干旱[1]。高温和干旱导致碳(C)、氮(N)、磷(P)的生物地球化学循环缓慢,有机物分解速率和初级生产力低[2]。碳、氮、磷是植物生长发育所必需的基本化学营养元素,它们在植物的各种生理代谢中发挥着重要作用。这些元素及其比值在植物不同器官间的变化及分配格局,反映了植物对非生物环境胁迫的适应策略。基于地理梯度的植物生态化学计量研究虽已在区域至全球尺度广泛开展,然而受限于器官采样难度及研究成本,相关成果大多数集中在叶片的化学计量特征上,而茎、根等器官的化学计量分异机制仍缺乏系统研究[3]。值得注意的是,由于植物器官养分元素的分配由特定的器官功能主导,并表现为对环境因子的差异化响应模式[4],具体而言,叶片作为光合器官,其元素投资优先满足碳同化与水分平衡,因此对水、光条件敏感;茎作为支撑与输导结构,发育与代谢效率更易受温度调控;根系化学特征由土壤养分有效性直接塑造,同时也被植物体内的碳-养分循环所调控[5−6]。可见,揭示不同器官的化学计量特征如何响应环境梯度,是阐明植物整体资源分配策略与适应机制的关键。
气候与土壤因子被认为是塑造这种器官特异性响应模式的主要驱动力[6−7]。大尺度研究表明:叶片氮、磷常随年平均气温(MAT)和年均降水量(MAP)增加而降低[5]。然而,不同器官对同一环境因子的响应可能迥异。例如,有研究发现:荒漠植物叶片磷受气候因子显著影响,而茎和根则不然[8];另外,HE等[9]研究表明:根系化学计量与土壤磷、pH等局地因子关系更为密切。由于已有研究结论不一致,驱动植物各器官化学计量特征分异的关键环境因子及其相对重要性仍不明确,因此,亟需通过对多个器官的系统比较来加以厘清。
中国东南湿润区到西北干旱区,存在一条由降水量和气温驱动的天然植被、土壤梯度带(以下简称降水梯度带)。该降水梯度带空间跨度约13个纬度(30.34°~42.82°N),年均降水量为44.17~1 522.61 mm,海拔和光合有效辐射(PAR)强度变化较大。这些因素共同驱动梯度带中植被的生理生态过程。狗尾草Setaria viridis作为干旱半干旱地区的优势物种和C4植物研究的模式物种,被广泛运用于植物生理代谢机制探索和生态系统养分循环研究等领域[10],在中国具有广阔的分布区,是研究草本植物生态化学计量学的理想材料之一。基于此,为了阐明狗尾草不同器官碳、氮、磷化学计量特征受降水梯度带环境因子变化的影响,本研究于2024年夏季沿中国东南—西北10个采样点对狗尾草进行生态化学计量学分析,旨在解决以下问题:①沿降水梯度,狗尾草各器官的化学计量变异格局是什么?②植物如何通过器官间差异化的化学计量可塑性,实现整体上对多尺度环境异质性的协同适应?③植物不同功能器官(叶、茎、根)的化学计量特征是否受其特有核心功能驱动,而对不同环境因子产生特异化响应?
-
中国东南—西北是一条由降水量(或干旱程度)驱动的梯度带:受季风气候影响,东南沿海水热充沛,植被以常绿阔叶林为主;向西北至河南及黄土高原,水热条件下降,植被过渡为温带落叶林;受贺兰山阻隔,黄河流域成为半湿润与干旱区分界,渐变为荒漠。本研究的采样地点从东南部浙江天目山至西北部新疆哈密,横跨整个中国,呈现显著的水热梯度,具体为浙江天目山,安徽宣城、六安,河南信阳、三门峡,陕西铜川,宁夏吴忠,甘肃张掖、酒泉,新疆哈密(表1)。
表 1 研究点及其环境特征
Table 1. Description of study sites and their environmental characteristics
区域 研究点 纬度(N)/(°) 海拔/m 年平均气温/℃ 年均降水量/mm 光合有效辐射/ (mol·m−2·d−1) 湿润区 天目山 30.340 1 142.0 24.94 1 522.61 38.01 宣城 30.620 167.2 25.31 1 394.61 38.41 六安 31.533 353.2 25.22 1 134.24 40.03 信阳 32.130 393.1 24.74 1 082.29 40.16 半湿润区 三门峡 34.800 697.4 23.81 555.64 41.09 铜川 35.820 1 087.0 20.29 584.43 44.17 吴忠 37.980 1 811.5 20.50 235.59 49.76 半干旱/干旱区 张掖 38.930 1 408.2 19.55 131.72 50.26 酒泉 39.770 1 436.1 19.18 81.85 51.96 哈密 42.820 577.0 23.37 44.17 54.47 -
2024年7月中旬至8月中旬,在每个样点设置1个10 m×10 m的调查样地,并在样地内选取没有病虫害、植株较完整、生长相对一致的狗尾草20株,连同根系完整挖出。同时,清除样地表面的枯枝落叶及浮土后,按五点取样法在样点采集0~10 cm的土壤样品,将所有土壤混合均匀,装袋打包,用生鲜快递运回实验室。
采集的植物全株用清水冲洗干净,将根、茎、叶剪下,分别包裹后105 ℃杀青15 min,75 ℃烘干至恒量;用电子天平分别称取干质量(精确度为0.000 1 g),得到各器官生物量。将根、茎、叶粉碎后置于密封袋中干燥保存;将采集的土壤去除石块、土壤动物及植物根系后,自然风干,过100目筛后,装于铝盒,用于测定各项土壤指标。
-
植物、土壤研磨后用于碳、氮、磷质量分数测定。采用重铬酸钾-外加热容量法(Walkley-Black法)测定植物及土壤样品的有机碳质量分数。采用凯氏定氮法(Kjeldahl法)测定植物及土壤样品中的全氮质量分数。采用钼锑抗比色法测定样品中的全磷质量分数。计算得出碳氮比(C/N)、碳磷比(C/P)和氮磷比(N/P),用于后续分析。
-
降水梯度带狗尾草化学计量的变异程度用变异系数(VC)=标准差(σ)/平均值(μ)来量化。采用三因素嵌套方差分析(nested-ANOVA)对每个化学计量变异的空间分布(区域、样点、植物个体)进行检验。
主成分分析(PCA)被用于可视化狗尾草器官间化学计量特征的协调模式,选择前2个主成分 (第1主成分和第2主成分) 进行后续分析。采用线性或二次回归模型分析各器官化学计量特征与地理因子(纬度、海拔)的关系。使用分段结构方程模型 (piecewise SEM)分析地理、气候和土壤因子对狗尾草化学计量特征的直接和间接影响。使用 Fisher C 检验评估建模结果的拟合优度。使用R软件中的“rdacca.hp”包进行层次分割(HP),评估气候和土壤因子对狗尾草各器官化学计量特征变异的相对重要性[11−12],并用回归分析检验各器官主成分分析前2轴与环境因子的相关显著性。以上数据处理及图表制作在Excel、Minitab 2021、Origin 2024、R 4.3.1中完成。
-
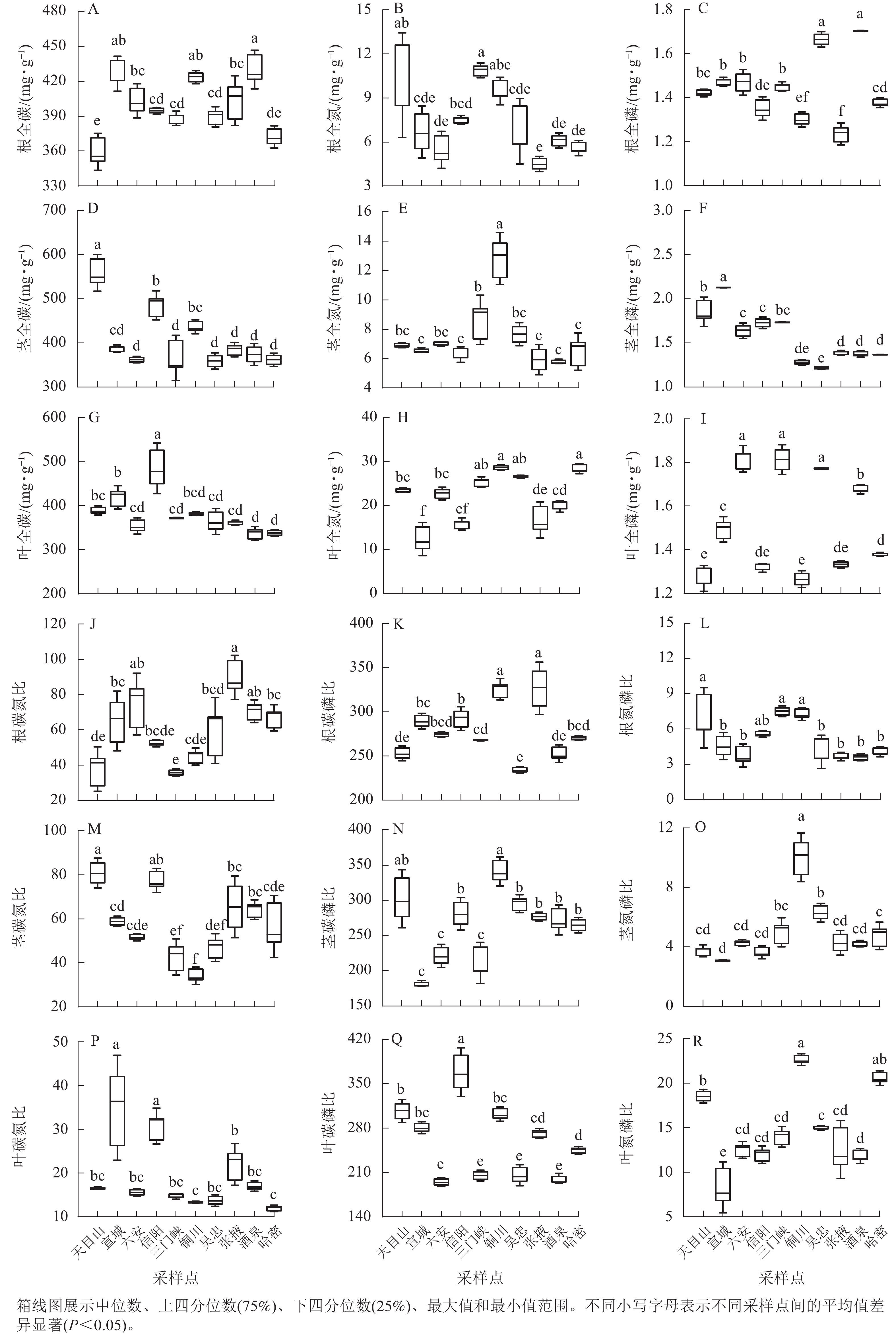
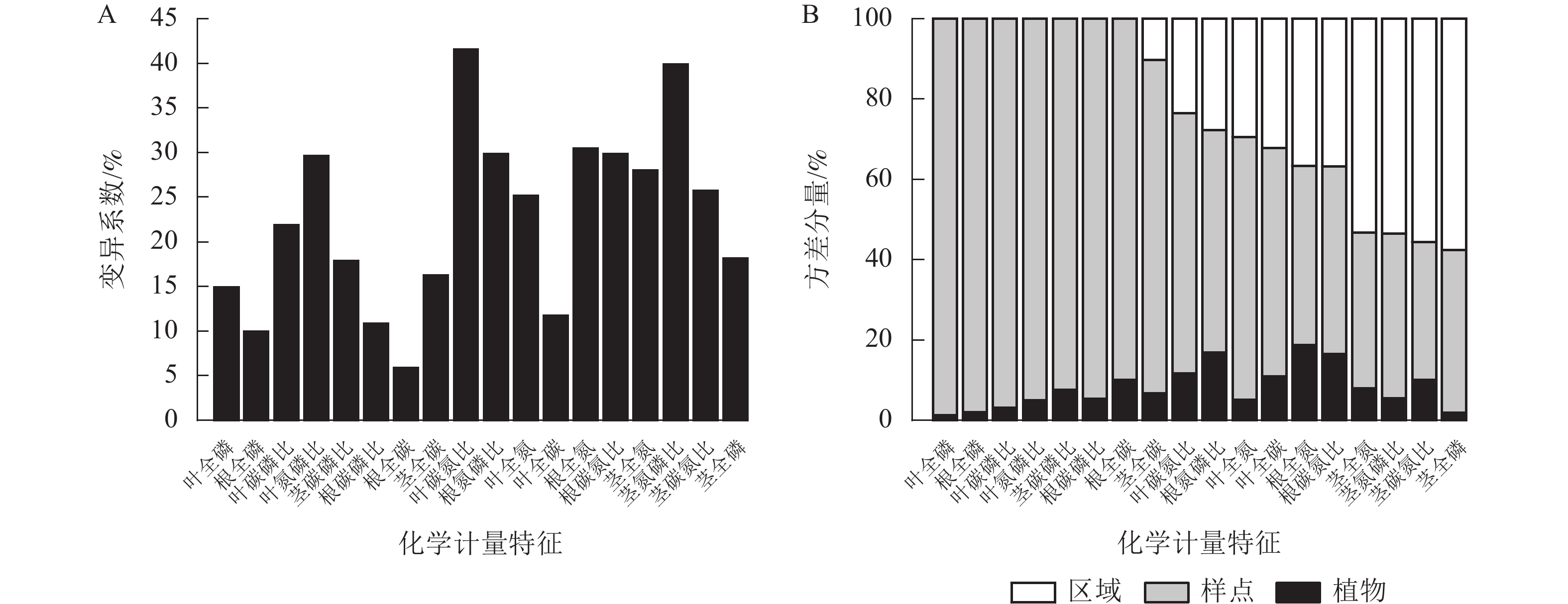
狗尾草化学计量特征沿降水梯度带的变异系数见图1A。在样带水平,所有化学计量特征的变异系数范围为10.00%~40.00%;各器官(根、茎、叶)中全碳的变异系数最小,分别为:根全碳(9.13%)、茎全碳(16.24%)、叶全碳(11.7%)。由于全碳的变异系数小,而全磷的变异系数大,所以在根、茎、叶化学计量特征的比值中碳磷比的变异系数最小,分别为根碳磷比(11.24%)、茎碳磷比(17.85%)、叶碳磷比(21.85%)。
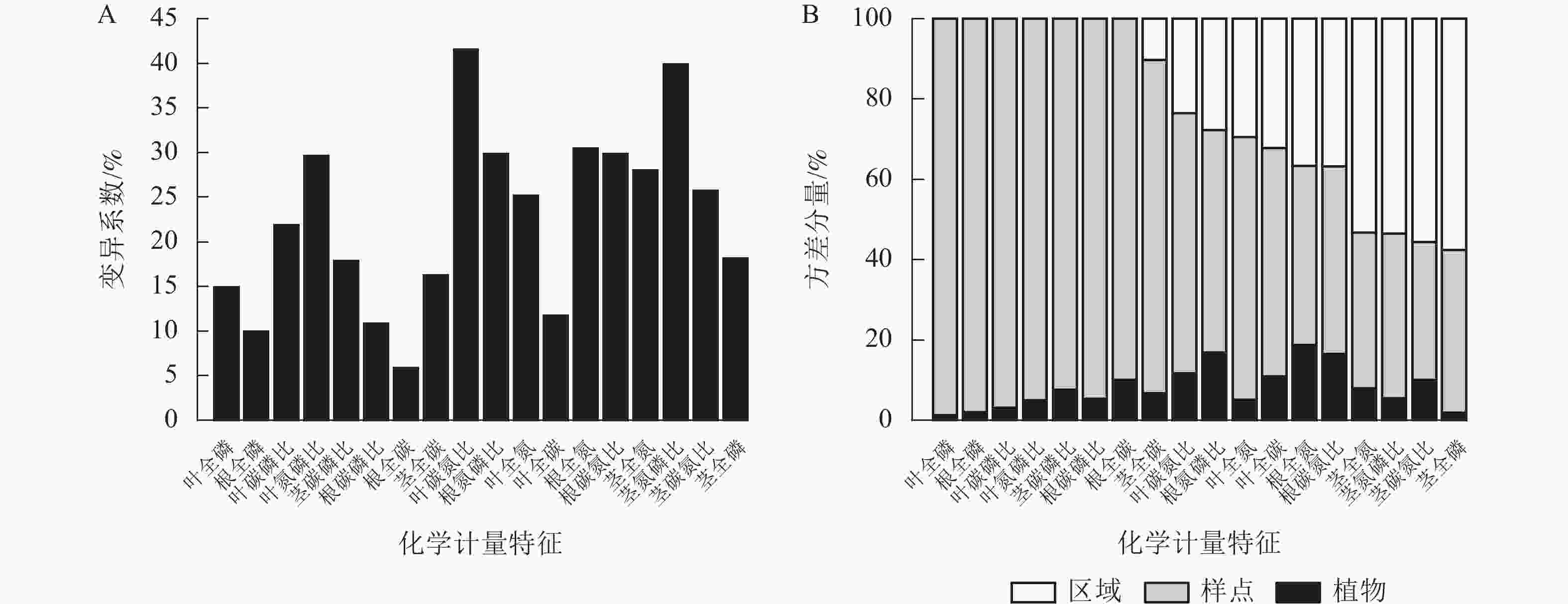
图 1 狗尾草化学计量特征沿降水梯度带的变异系数及方差分量
Figure 1. Variation coefficient and variance components analysis of S. viridis stoichiometric traits along precipitation gradients
方差分量见图1B。狗尾草化学计量特征的样点水平变异整体高于区域水平变异。在所研究的18个化学计量特征中,仅茎全氮、茎全磷、茎氮磷比、茎碳氮比4个化学计量特征的区域水平变异大于样点水平变异。
狗尾草化学计量特征在不同样点间的差异见图2。狗尾草各器官化学计量特征在样点间表现出显著差异。其中,氮和氮磷比均为陕西铜川显著大于其他样点(P<0.05),碳质量分数在各样点间变化较小。碳、磷质量分数及其比值在样点间差异显著,但无明显规律。
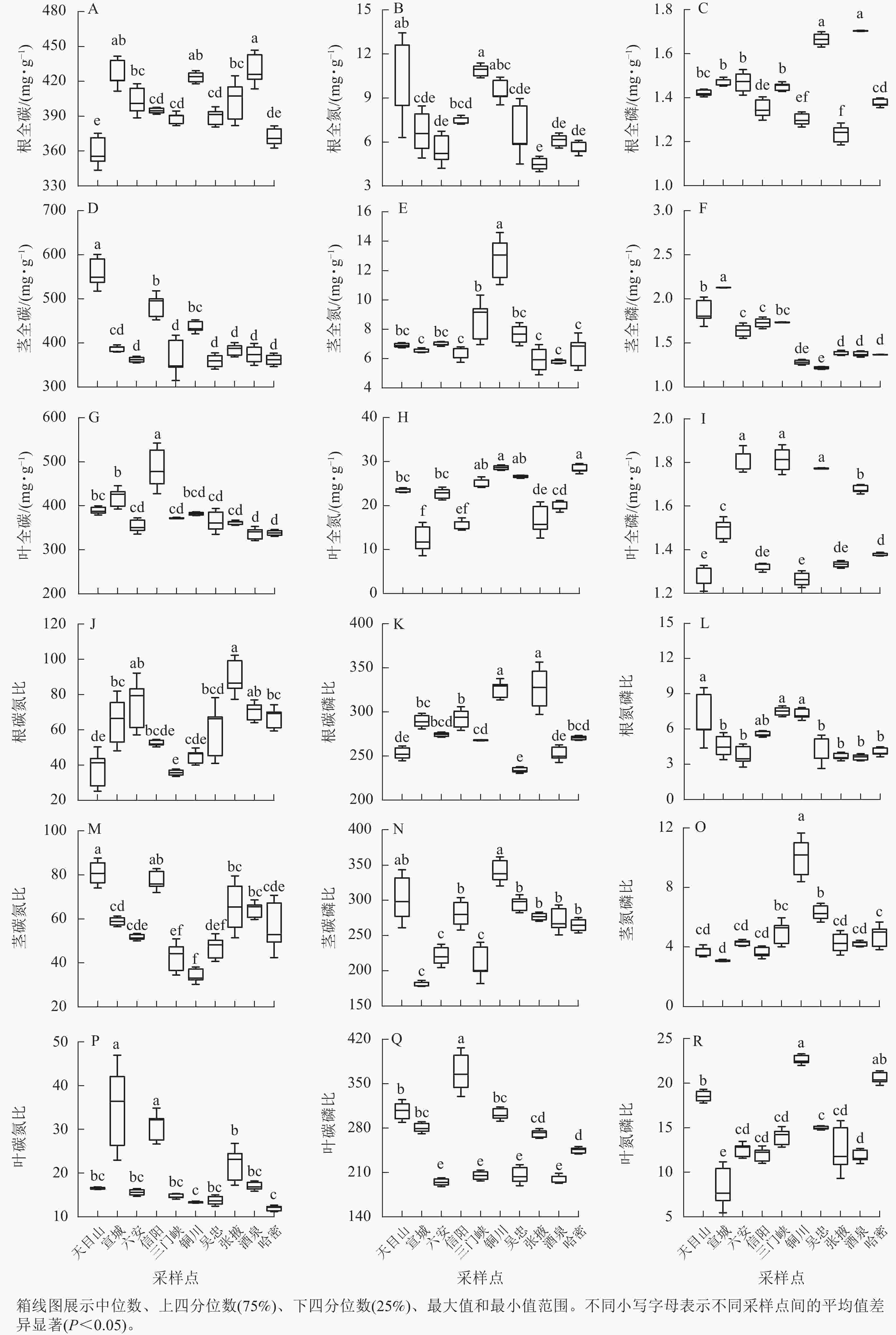
图 2 狗尾草化学计量特征在不同气候区域的差异
Figure 2. Differences in stoichiometric traits of S. viridis along different climate regions
-
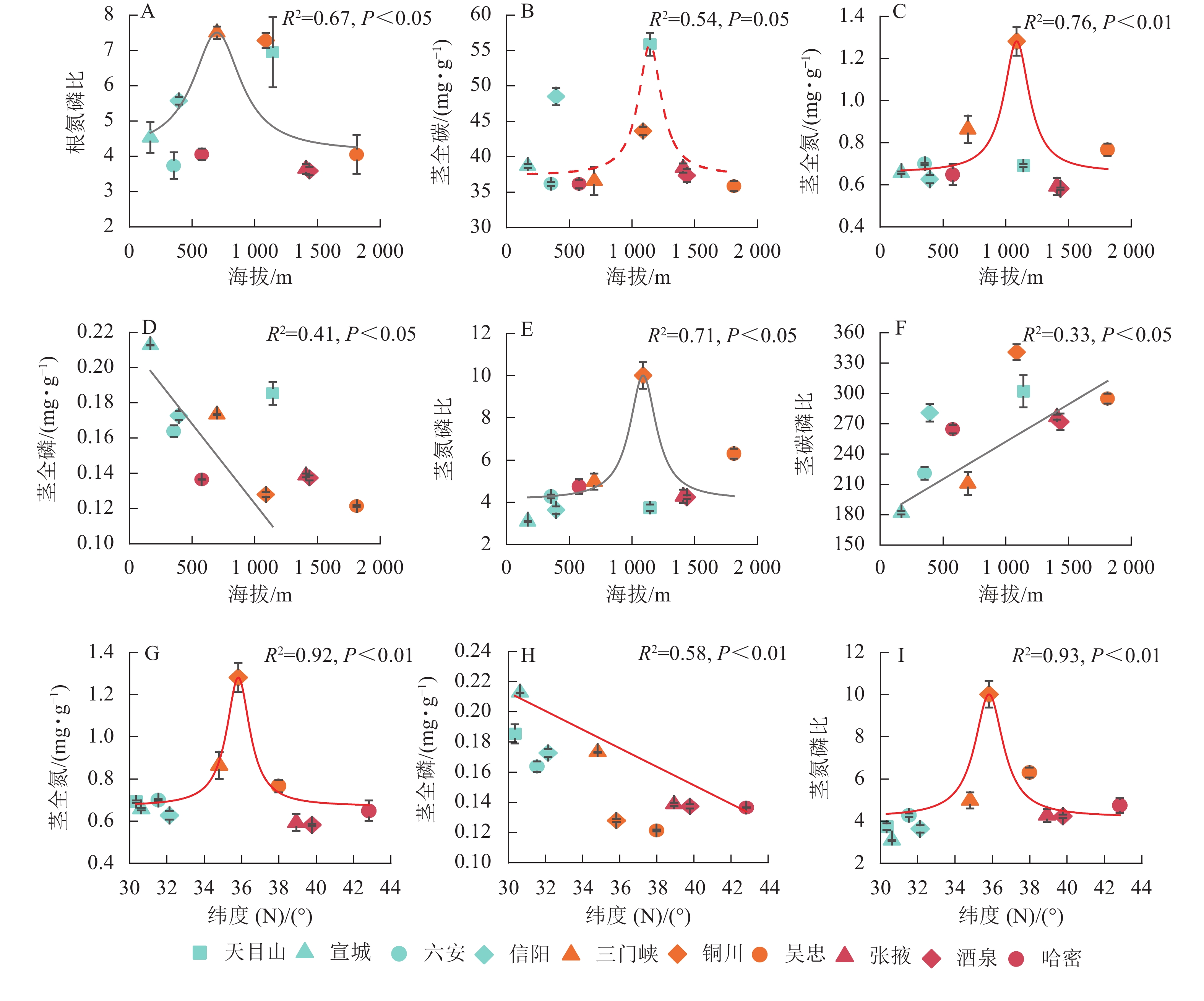
狗尾草化学计量特征与样点地理因子(纬度、海拔)的拟合关系见图3。根氮磷比、茎全碳、茎全氮、茎氮磷比与海拔呈单峰关系,茎全磷与海拔呈显著负相关(P<0.05),茎碳磷比与海拔呈显著正相关(P<0.05);茎全氮和茎氮磷比与纬度呈二次曲线关系(P<0.01);茎全磷与纬度呈显著负相关(P<0.05)。

图 3 狗尾草化学计量特征与地理因子(纬度、海拔)的关系
Figure 3. Relationships between ecological stoichiometric traits of S. viridis and geographical factors (latitude and elevation)
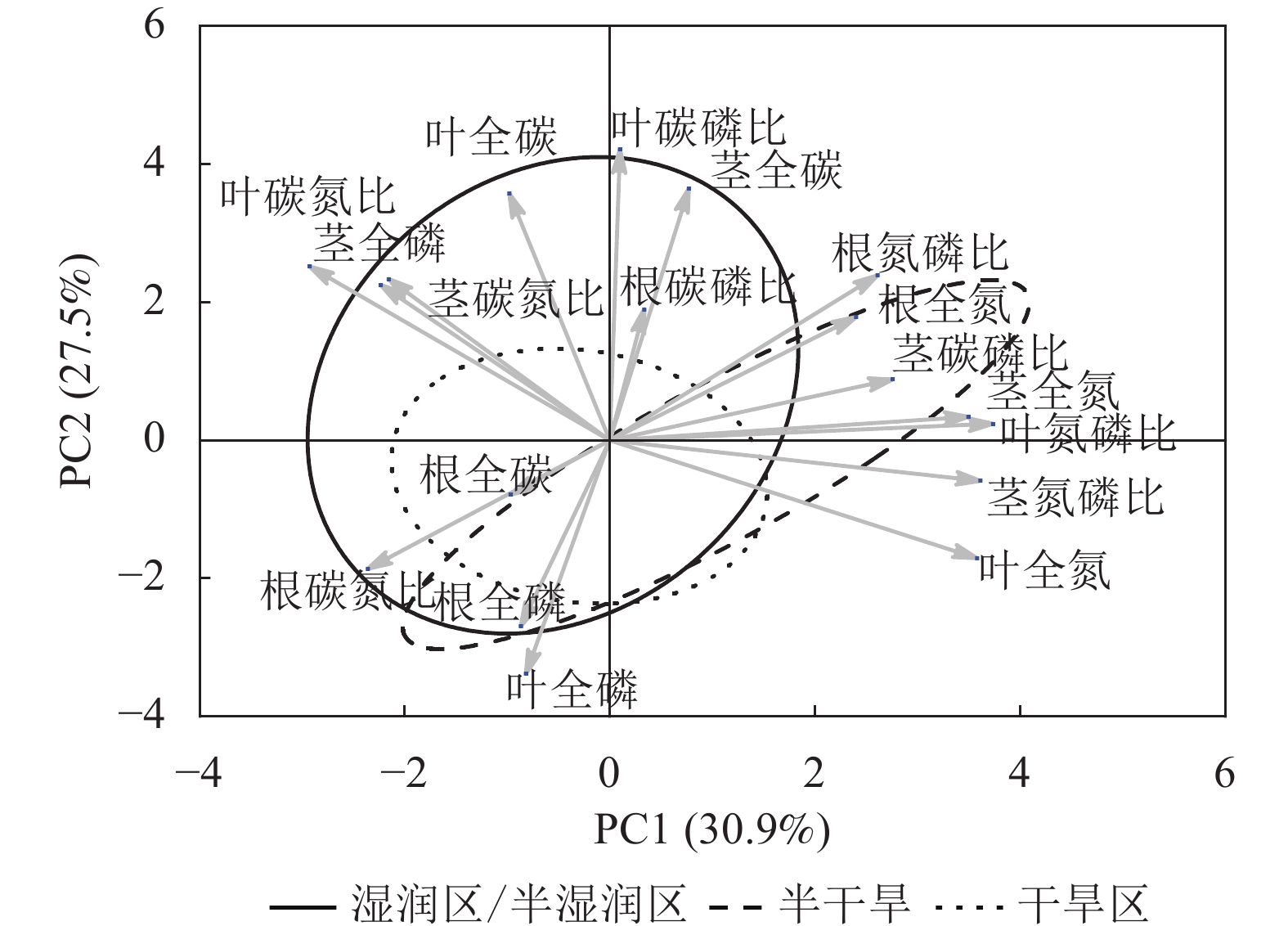
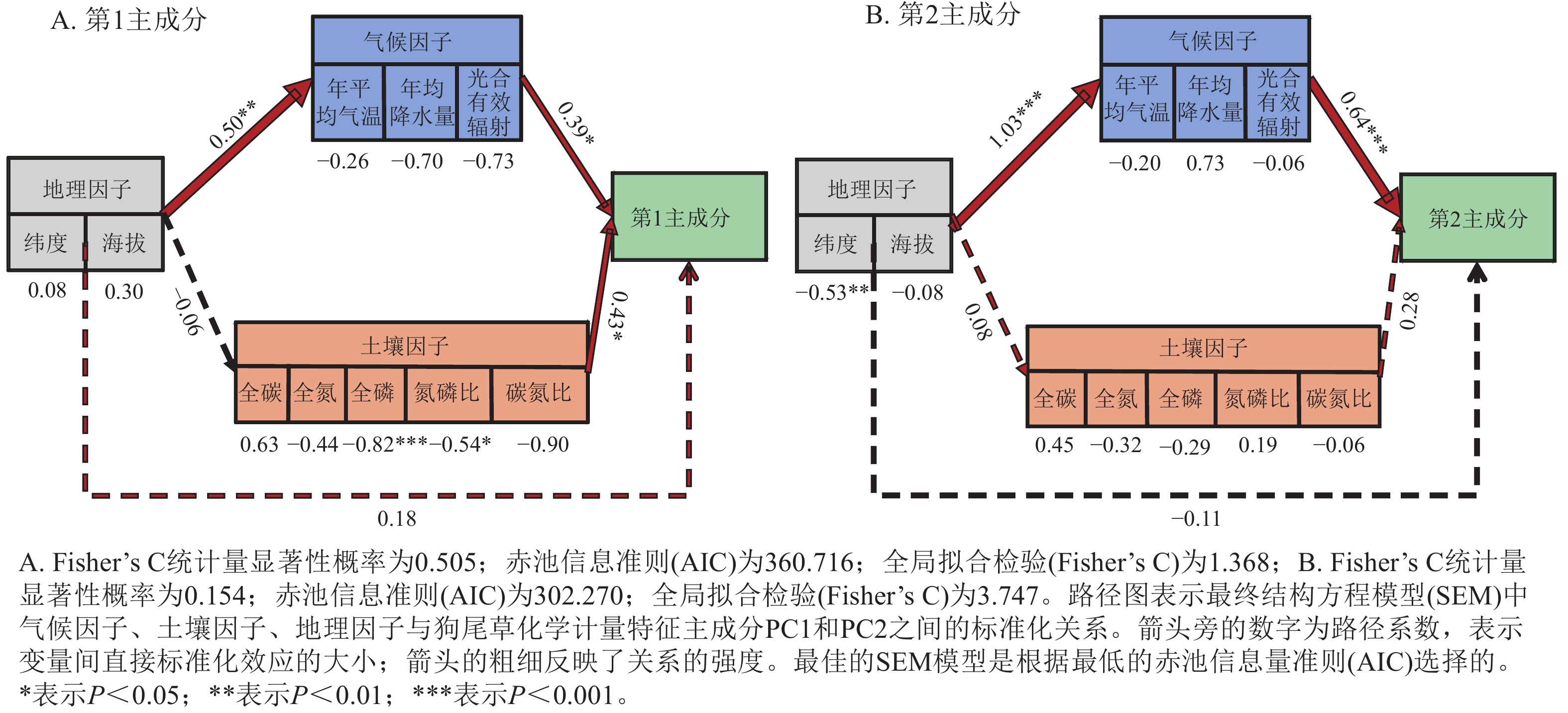
狗尾草化学计量特征的主成分分析见图4。第1主成分的解释率为30.9%,关键化学计量特征为叶氮磷比(载荷为0.88)、茎氮磷比(载荷为0.85)、叶全氮(载荷为0.84)和茎全氮(载荷为0.82);第2主成分的解释率为27.5%,关键化学计量特征为叶碳磷比(载荷为0.94)、茎全碳(载荷为0.81)、叶全碳(载荷为0.80)和叶全磷(载荷为0.75)。根据结构方程模型分析(图5),模型整体拟合良好(卡方检验 P=0.505与0.154)。具体来说,气候因子与土壤因子对狗尾草化学计量特征(第1主成分)均呈显著正向影响(解释率分别为R2=0.39与R2=0.43,P<0.05),而地理因子的直接影响不显著,但地理因子对气候因子存在显著影响(R2=0.50,P<0.01)。对于狗尾草化学计量特征(第2主成分),气候因子对狗尾草化学计量特征呈极显著正向影响(R2=0.64,P<0.001),地理因子的直接影响同样不显著。因此,气候和土壤因子在狗尾草化学计量特征沿降水梯度的变化中起主导作用。虽然地理因子不会直接影响狗尾草化学计量特征变异,但它们可以通过对气候和土壤的影响产生间接效应。
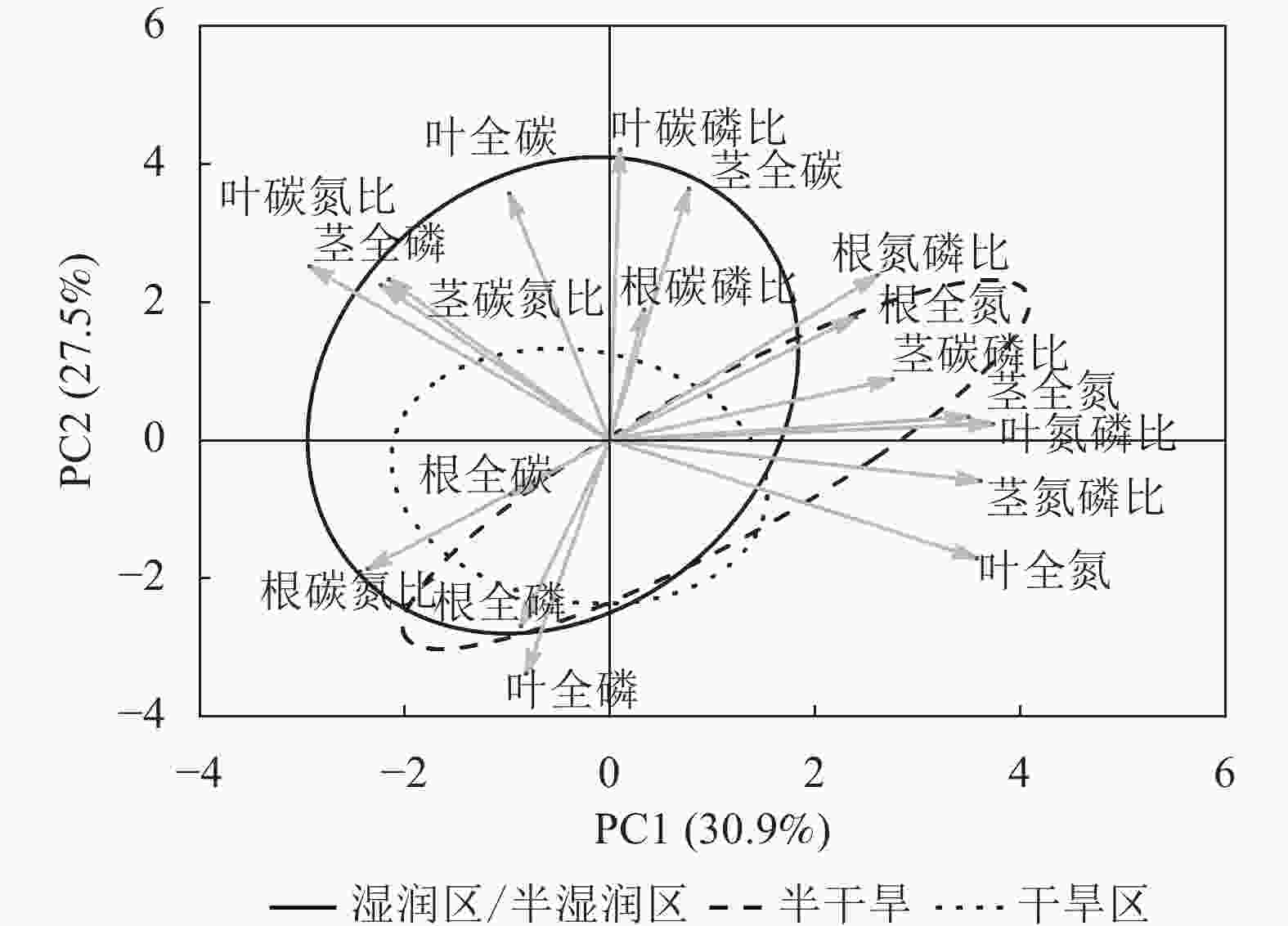
图 4 狗尾草化学计量特征的主成分分析
Figure 4. Principal component analysis of ecological stoichiometric traits in S. viridis
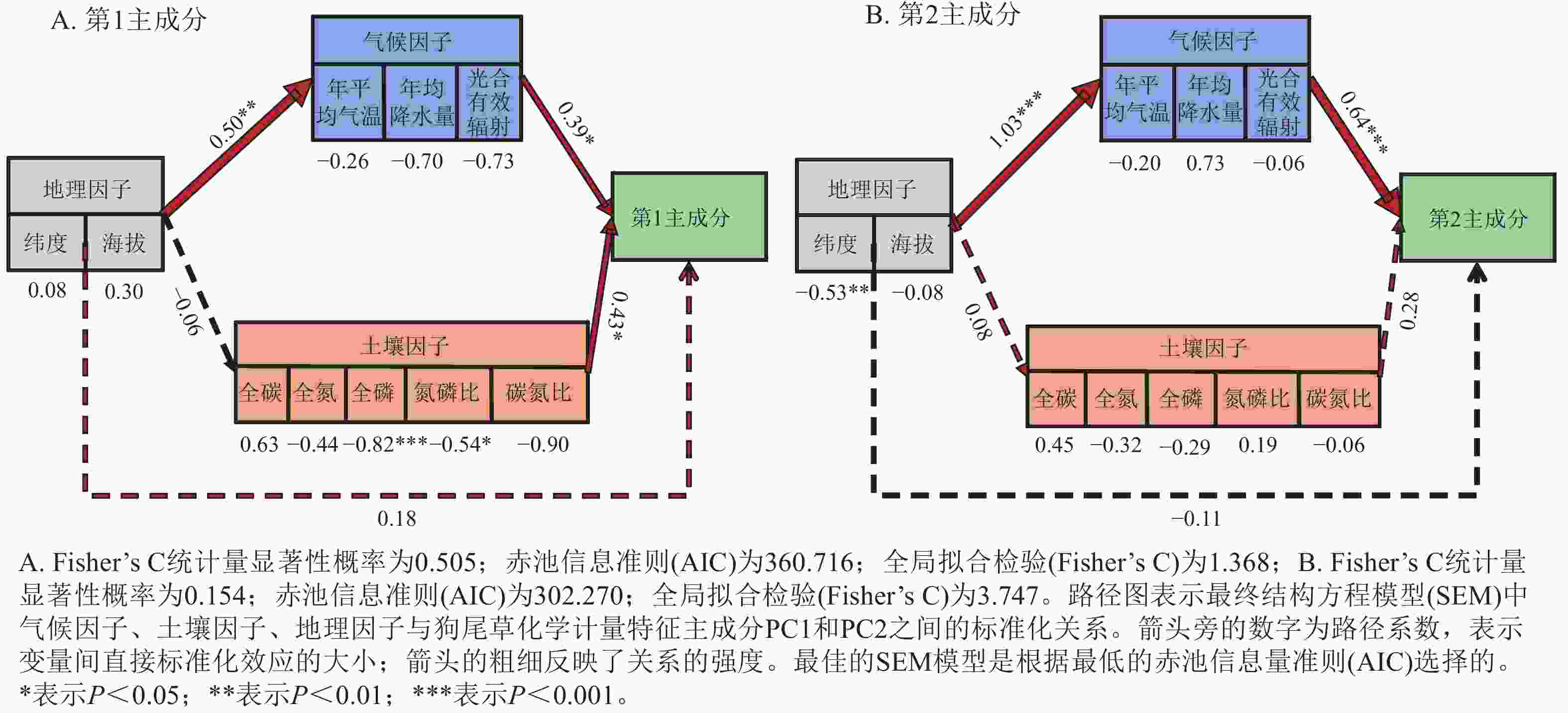
图 5 非生物因子与狗尾草化学计量特征主成分的分段结构方程模型
Figure 5. Segmented structural equation model of the relationships between abiotic factors and ecological stoichiometric traits PC1 and PC2 in S. viridis
-
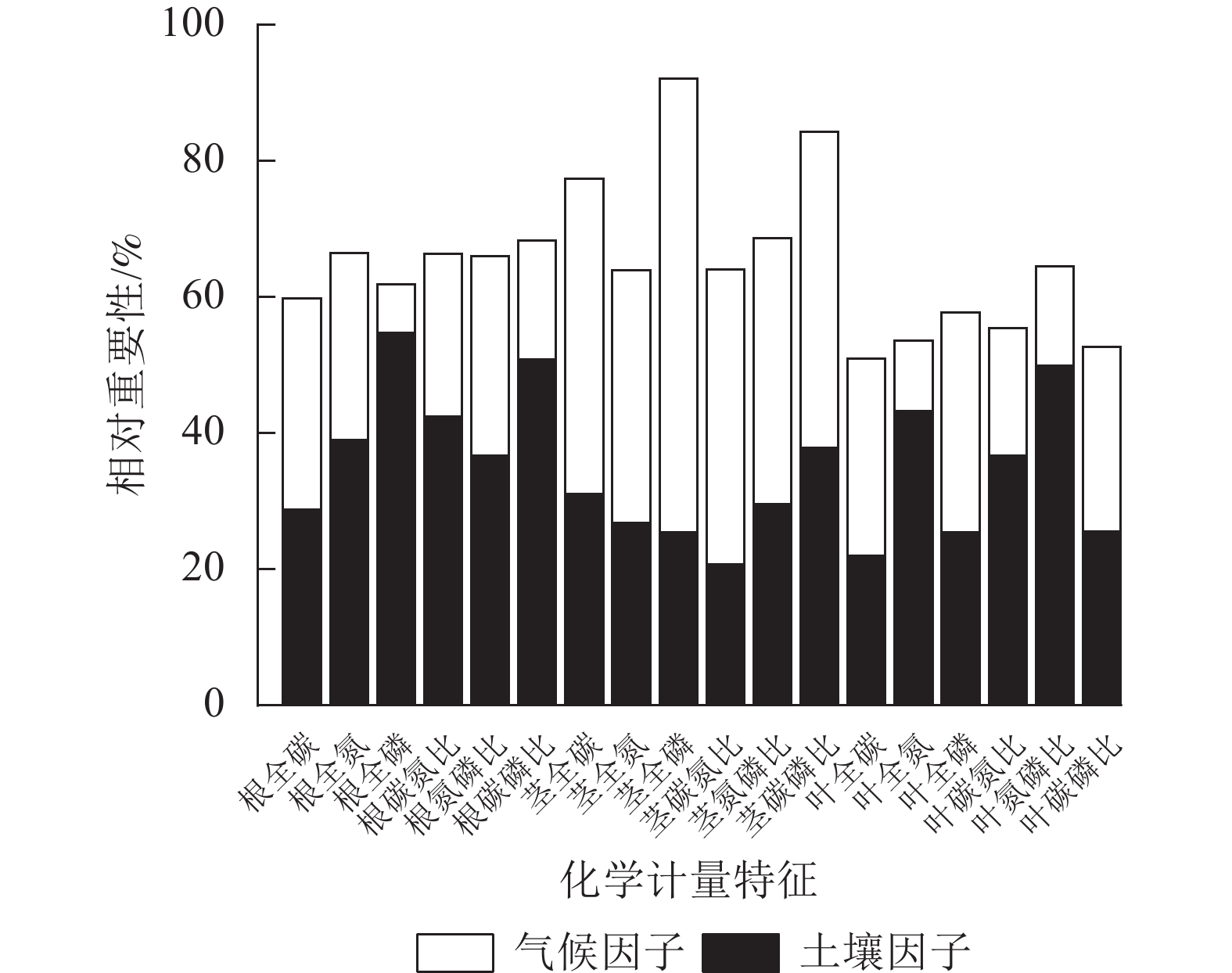
环境(土壤、气候)因子对狗尾草化学计量特征变异的相对重要性见图6。狗尾草各器官化学计量特征变异沿降水梯度受土壤、气候因子影响的总解释率达50%~95%。根化学计量特征变异主要受土壤因子驱动,其中受土壤因子解释率最大的根系化学计量特征为磷(54.67%);茎化学计量特征变异主要受气候因子驱动,其中受气候因子解释率最大的茎化学计量特征为磷(66.72%);叶化学计量特征变异则呈现元素特异性,其中受气候因子驱动的叶化学计量特征为碳(29.02%)、磷(32.27%)及碳磷比(27.04%),受土壤因子驱动的叶化学计量特征为氮磷比(49.94%)、氮(43.26%)、碳氮比(36.64%)。
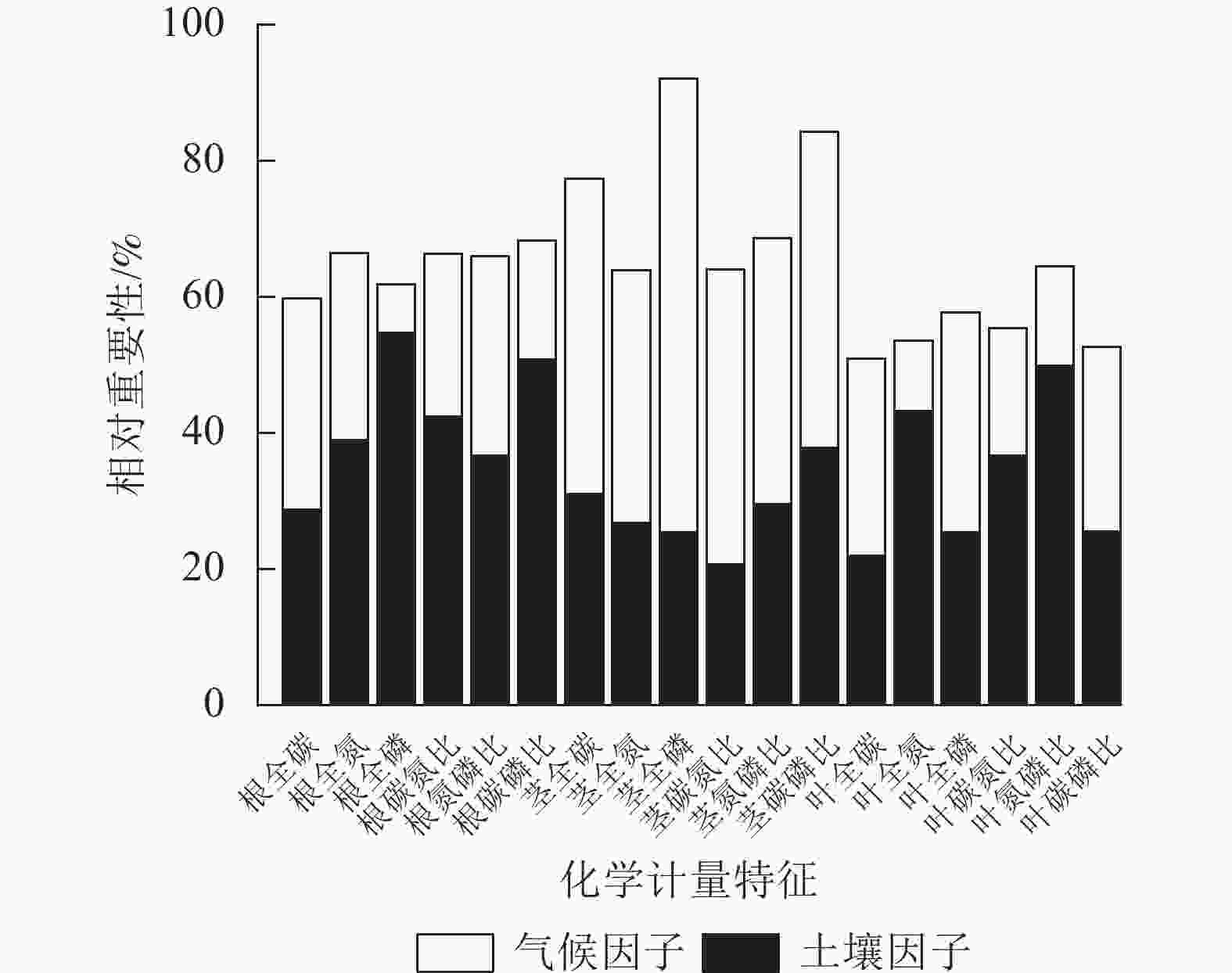
图 6 土壤、环境因子对狗尾草化学计量特征变异的相对重要性
Figure 6. Relative importance of soil and environmental factors on stoichiometric trait variation in S. viridis
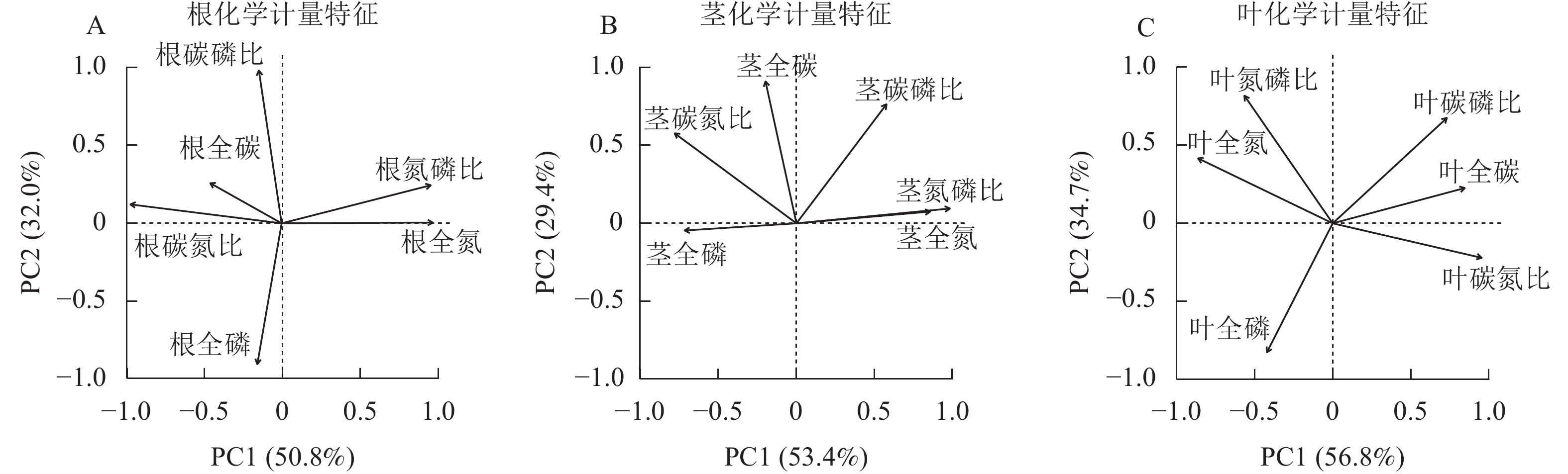
狗尾草根化学计量特征的主成分分析见图7A。第1主成分的解释率为50.8%,关键化学计量特征为根全氮(载荷为0.97)和根碳氮比(载荷为−0.97);第2主成分的解释率为32.0%,关键化学计量特征为根碳磷比(载荷为0.98)和根全磷(载荷为−0.90)。将环境(土壤、气候)因子与根化学计量特征第1主成分和第2主成分进行回归分析,结果表明:光合有效辐射、土壤全碳与第1主成分呈极显著相关(P<0.01,表2),土壤全磷与第1主成分呈显著相关(P<0.05,表2);土壤全氮、全磷与第2 主成分呈显著相关(P<0.05,表2)。
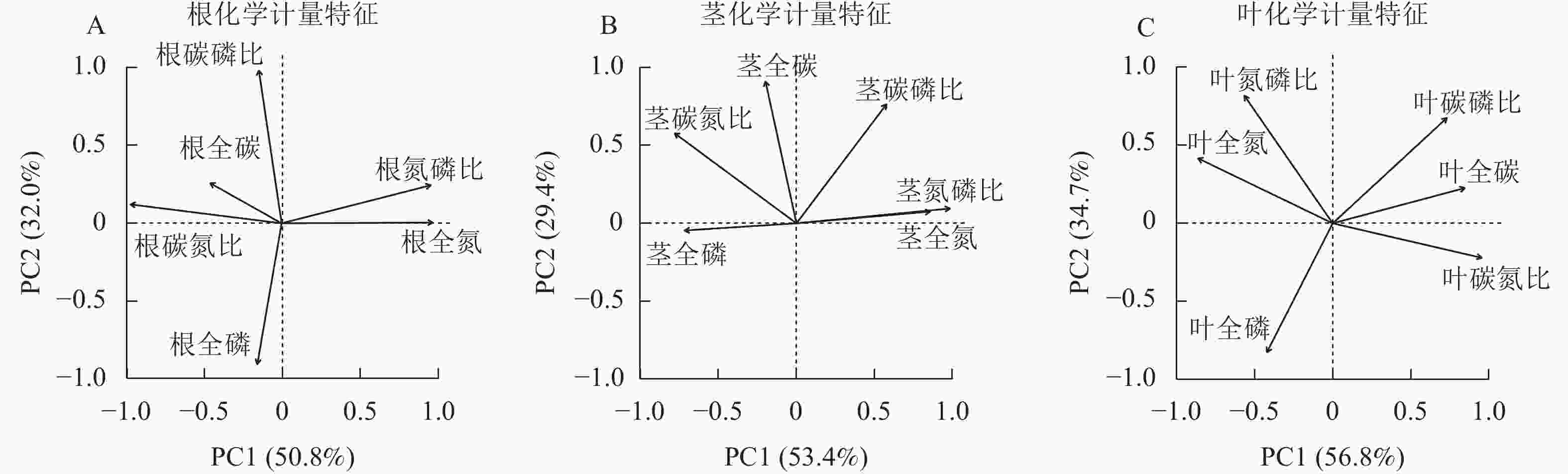
图 7 狗尾草根、茎和叶化学计量特征的主成分分析
Figure 7. Principal component analysis of root, stem, and leaf stoichiometric traits in S. viridis
表 2 狗尾草化学计量特征主成分与环境因子的相关关系
Table 2. Correlations between principal components of stoichiometric traits in S. viridis with environmental factors
环境因子 根化学计量特征 茎化学计量特征 叶化学计量特征 PC1 PC2 PC1 PC2 PC1 PC2 年均降水量 0.13 0.010 0.21* 0.070 0.270** 0.004 5 年平均气温 0.10 0.002 0.32** 0.001 0.120 0.000 4 光合有效辐射 0.23** 0.030 0.10 0.030 0.220** 0.000 1 土壤全碳 0.20** 0.070 0.14* 0.040 0.030 0.050 0 土壤全氮 0.11 0.180* 0.04 0.010 0.050 0.010 0 土壤全磷 0.18* 0.170* 0.08 0.230** 0.006 0.240 0** 说明:*表示P<0.05;**表示P<0.01。 狗尾草茎化学计量特征的主成分分析见图7B。第1主成分的解释率为53.4%,关键化学计量特征为茎氮磷比(载荷为0.98)和茎氮(载荷为0.86);第2主成分的解释率为29.4%,关键化学计量特征为茎碳(载荷为0.91)和茎碳磷比(载荷为0.76)。将环境(土壤、气候)因子与茎化学计量特征第1主成分和第2主成分进行回归分析,结果表明:年均降水量、土壤全碳与第1主成分呈显著相关(P<0.05,表2),年平均气温与第1主成分呈极显著相关(P<0.01,表2);土壤全磷与第2主成分呈极显著相关(P<0.01,表2)。
狗尾草叶化学计量特征的主成分分析见图7C。第1主成分的解释率为56.8%,关键化学计量特征为叶碳氮比(载荷为0.96)和叶全氮(载荷为−0.86);第2主成分的解释率为34.7%,关键化学计量特征为叶全磷(载荷为−0.83)和叶氮磷比(载荷为0.82)。将环境(土壤、气候)因子与叶化学计量特征第1主成分和第2主成分进行回归分析,结果表明:年均降水量和光合有效辐射与第1主成分呈极显著相关(P<0.01,表2),土壤全磷与第2主成分呈极显著相关(P<0.01,表2)。
-
植物化学计量特征变异大小的不同,反映了植物叶片碳、氮、磷的内稳性及其对生态环境变化的敏感性。本研究中,狗尾草氮磷比的变异系数大于氮、磷变异系数,表明沿降水梯度带狗尾草各个器官氮磷比具有较大的波动,且受氮变异主导。这一结果不仅与区域尺度上的同类发现相印证。杨思琪等[13]对天山草本植物的研究表明:氮与磷对环境梯度的响应不同步导致了化学计量比的显著波动。GÜSEWELL[14]的研究进一步证实:大尺度环境因子(如降水)主要通过调控氮循环过程来影响植物氮磷比平衡,其内在机制源于氮、磷生物地球化学循环的根本差异。在本研究区,氮质量分数的变异主要受降水调控的土壤矿化过程及可能的大气沉降等大尺度因素驱动;而磷则因其来源局地化(如母质)及植物对其更强的内稳性,在区域尺度上保持相对稳定,故不足以主导氮磷比的整体变异格局。
-
本研究表明:植物不同器官在地理梯度上的化学计量响应表现出显著的不一致性,其变异并非由地理因子(纬度或海拔)本身直接驱动,而是主要通过气候与土壤因子的协同变化产生。这一结果与全球尺度和区域尺度的多项研究结论相吻合。近年来针对草地与禾本科Poaceae植物的研究表明:器官化学计量特征更主要受到水热条件与土壤养分可利用性的调控,而非地理位置本身的直接效应[15]。在与本研究属同一类功能型的C4草本植物研究中,同样发现植物碳、氮、磷质量分数及其化学计量比沿环境梯度的变化主要由气候与土壤因子共同驱动,而非单一的纬度或海拔因素[16]。本研究结果表明:尽管部分茎化学计量特征与单个地理因子存在显著相关性,但根和叶的碳、氮、磷质量分数及其化学计量比在地理梯度上并未呈现稳定趋势,大多数性状仅与地理因子保持弱相关,说明地理位置更类似于环境差异的“空间载体”,而气候与土壤才是驱动草本植物化学计量格局形成的关键控制因子。
结构方程模型进一步从定量层面印证了这一认识。地理因子对第1主成分与第2主成分的直接路径均不显著,而是主要通过影响气候与土壤因子产生间接效应。该结果与近年来关于草本植物与狗尾草化学计量学的研究结果高度一致:气候因子通过调节碳同化速率、蒸腾过程与生理代谢活动,塑造器官层面的碳、氮、磷分配格局;而土壤碳、氮、磷质量分数及其化学计量比则通过影响根际营养供应和吸收效率,进一步调控根与茎的养分沉积和循环过程[16−17]。在以狗尾草为代表的C4草本系统中,还发现土壤磷有效性及土壤有机质背景对植物化学计量变化具有关键调节作用,支持“气候主导宏观性状变异,土壤主导局地适应分化”的多尺度调控框架[15, 18]。因此,本研究结果为理解典型草本植物(狗尾草)在降水梯度与土壤养分梯度上如何通过器官功能分化实现适应性权衡提供了新的经验证据,也强调了在探讨地理格局时必须将地理位置、气候条件与土壤环境加以区分,而不能简单将“纬度/海拔-性状”视作直接因果关系。
-
本研究发现:主导各器官化学计量特征主要变异模式(第1主成分)的环境因子存在明显差异:叶片化学计量主要受水分和光照驱动(年均降水量、光合有效辐射的载荷值最高且显著),茎化学计量主要受温度驱动(年平均气温的载荷值最高且极显著),而根系化学计量则同时受到光照和土壤碳的影响(光合有效辐射、土壤全碳的载荷值显著)。这一格局与各器官的核心生态功能高度吻合。
本研究揭示:狗尾草根、茎、叶三大器官的化学计量特征对沿降水梯度的环境因子存在明显的差异化响应格局。具体而言,叶片化学计量的主要变异模式(第1主成分解释56.8%的变异)由水分(年均降水量)和光照(光合有效辐射)主导;茎化学计量的主要变异(第1主成分解释53.4%的变异)主要受温度(年平均气温)驱动;根系化学计量的主要变异(第1主成分解释50.8%的变异)则同时受到地上光照(光合有效辐射)与地下土壤碳(土壤全碳)的影响。这一发现与各器官的核心生态功能高度吻合。
研究发现:作为光合作用的主要场所,叶片化学计量对水分和光照的敏感响应反映了其对碳获取过程的直接调控。水分是气孔开闭和光合作用的初级限制因子[19]。较高的年均降水量通常意味着更少的水分胁迫,有利于气孔开放以增加二氧化碳同化,同时增强蒸腾流对氮等矿质养分的运输。这种协同作用最终塑造了叶片碳氮质量分数的核心变异模式,具体表现为本研究第1主成分中叶碳氮比的关键贡献[20]。光合有效辐射作为光合作用的能量来源,其强度直接决定了碳固定的潜力。在高光环境下,植物倾向于投资更多氮资源于光合器官(如增加Rubisco酶),取得最大化光能利用效率。这导致了叶片氮质量分数及碳氮比的系统性变化,符合全球叶片经济学谱的基本规律[21],因此,水分和光照通过共同调控碳同化与氮代谢的效率,成为塑造叶片化学计量大尺度格局的关键环境因子。
另外,茎作为植物的支撑结构与物质输导通道,其化学计量对年平均气温的强依赖性,体现了温度对植物整体代谢速率与生理过程的全局性控制。根据代谢生态学理论,温度是影响生物体生化反应速率(如呼吸、生长)的关键因素[22]。在温暖条件下,植物代谢速率加快,对茎部维管组织的输导效率要求更高,可能驱动其在茎部投资更多与代谢和输导相关的氮、磷资源。这正解释了本研究中茎全氮与茎氮磷比在关键变异轴(第1主成分)上的高载荷。同时,温度也深刻影响生长季长度与物候,调控非结构性碳水化合物在茎中的储存与再动员策略,从而影响其碳化学计量[23]。因此,年平均气温作为一个综合热量指标,通过调节植物的代谢成本、输导需求及物候节律,主导了茎化学计量的宏观变异。
最后,根系同时承担资源吸收与环境感知的双重功能,其化学计量对地上光照(光合有效辐射)和地下土壤碳(土壤全碳)的共同响应,揭示了植物整合地上与地下资源信号的权衡策略。光合有效辐射不仅是光合作用的能量来源,也是指示地上碳同化潜力的关键信号。本研究中根碳氮比、根全氮所表现的关键贡献表明:较高的光合有效辐射通常意味着更强的“源”端碳供应,植物可能将更多的光合产物分配至根系,用于其生长、呼吸或分泌功能,从而直接改变根系的碳、氮化学计量基础[24]。土壤全碳是地下碳循环与养分有效性的重要指标。较高的土壤全碳往往与更活跃的微生物活动和土壤有机质周转相关联,这能改善土壤结构、促进氮等养分的矿化与供应,进而驱动根系调整其吸收策略与化学计量组成以适应局部养分环境[25]。因此,根系化学计量同时受光合有效辐射(地上碳供应潜力)和土壤全碳(地下碳过程与养分环境)的影响,生动体现了植物在协调“觅光”与“觅肥”投资时的整体适应策略。
本研究表明:狗尾草不同器官因固有的生态功能分化(叶主光合、茎主输导与支撑、根主吸收与感知),对环境梯度中的关键因子产生了特异性的敏感响应。这种由器官功能分化介导的差异化环境响应,是植物在变化环境中实现整体适应性生长的重要机制。
-
本研究以中国东南—西北降水梯度带上10个样点的狗尾草为研究对象,旨在分析其不同器官间化学计量特征的变异规律和驱动因素。结果表明:狗尾草氮磷比的变异主要受氮质量分数空间异质性驱动,而磷质量分数则因植物内稳性更强及来源局地化而相对稳定。这一差异根植于氮、磷的生物地球化学循环属性:氮循环受大尺度降水过程调控,而磷更依赖于局地成土母质。研究进一步发现:狗尾草的化学计量特征由多尺度环境因子共同塑造,受多尺度环境因子层级调控[26],即大尺度气候条件筛选出基础的资源利用策略,而局部土壤异质性(尤其是磷的差异)则在此基础上进一步细化出更具体的适应性特征,体现了其功能分化所塑造的特化适应格局。此外,纬度与海拔等地理因子本身并无直接效应,主要通过改变上述水热与土壤条件间接发挥作用,充当了环境梯度的“空间载体”。更重要的是,狗尾草根、茎、叶形成了功能特化的模块化响应策略。叶片化学计量(以碳氮比为核心)响应水分和光照,优化光合碳获取;茎秆化学计量(以氮磷比为核心)主要响应温度,协调代谢与输导;根系化学计量则整合地上光信号与地下土壤碳信息,以平衡吸收与分配。这种高度耦合的“功能−响应”机制是其实现整体适应性的核心。
综上所述,狗尾草通过形成“氮主导、气候驱动、器官功能分化响应”的化学生态策略,实现对大尺度环境异质性的适应。本研究从元素循环−环境驱动−器官功能整合的新视角,深化了对草本植物养分适应机制的理解。
Differences in contributions of climate and soil in precipitation gradient zones to variation in stoichiometric traits of Setaria viridis
-
摘要:
目的 分析广布种狗尾草Setaria viridis器官间的养分化学计量模式及其驱动因子,揭示狗尾草在大区域尺度下的资源获取策略和生长适应性。 方法 选取中国东南湿润区至西北干旱区降水梯度带上10处样点,以广布种狗尾草为目标植物,测定其根、茎、叶的碳、氮、磷化学计量指标,采用单因素方差分析与主成分分析等方法,探究狗尾草化学计量沿降水梯度带的变异规律,用结构方程模型和层次分割法进一步揭示驱动化学计量变异的主要环境因子。 结果 ①狗尾草各器官全碳质量分数变异系数最低(根为9.13%,叶为11.70%),而全氮、全磷变异较大,反映了限制性元素对环境变化的敏感响应。②纬度与海拔等地理因子对化学计量特征无显著直接效应,其影响主要通过调控气候与土壤条件而间接实现。③器官间环境驱动因子存在分化:根化学计量变异主要由土壤因子解释,茎变异主要受气候因子主导,叶化学计量特征则与年均降水量、光合有效辐射和土壤全磷呈显著正相关(P<0.01)。 结论 狗尾草器官间化学计量特征在不同样点间存在显著分异,主要由环境因子(气候、土壤)驱动。植物的化学多样性是由大尺度气候条件筛选出的基础资源利用策略(第1主成分)与局部土壤异质性(尤其是磷)所细化的具体适应性特征(第2主成分)通过层级调控共同决定的。这些发现强调了从器官尺度理解植物适应性分异的重要性,为更精准地预测未来气候变化下植物的响应与适应轨迹提供了新的视角。图7表2参26 Abstract:Objective The objective is to analyze the nutrient stoichiometric patterns among organs of the widespread species Setaria viridis and the driving factors, so as to reveal its resource acquisition strategies and growth adaptability on a large regional scale. Method The 10 sampling sites were selected along a precipitation gradient from the humid southeast region to the arid northwest region of China, with the widespread species S. viridis as the target plant. The carbon (C), nitrogen (N), and phosphorus (P) stoichiometric indices in its roots, stems and leaves were measured. Using one-way ANOVA and principal component analysis, the variation patterns of stoichiometry along the precipitation gradient were investigated. Structural equation modeling (SEM) and hierarchical partitioning were further employed to identify the key environmental factors driving stoichiometric variation. Result (1) The coefficient of variation of total carbon mass fraction in all organs of S. viridis was the lowest (9.13% for roots and 11.70% for leaves), while the variations of total nitrogen and total phosphorus were relatively large, reflecting the sensitive response of limiting elements to environmental changes. (2) Geographical factors such as latitude and elevation had no significant direct effect on the stoichiometric traits. Their influence was achieved indirectly by regulating climatic and soil conditions. (3) The differences in environmental driving factors among organs lay in that the variation in root stoichiometry was primarily explained by soil factors while stem stoichiometry was predominantly driven by climatic factors. Leaf stoichiometric characteristics were positively and significantly correlated with mean annual precipitation, photosynthetically active radiation, and soil total phosphorus. Conclusion The stoichiometric traits of S. viridis organs exhibit significant differences across different sampling sites, primarily driven by environmental factors (climate and soil). The chemical diversity of plants is determined jointly by the basic resource utilization strategies (the first principal component) selected by large-scale climatic conditions and the specific adaptive characteristics (the second principal component) refined by local soil heterogeneity (especially phosphorus), through hierarchical regulation. These findings highlight the importance of understanding plant adaptive differentiation at the organ scale, providing a novel perspective for more accurate prediction of plant response and adaptation trajectories under future climate change. [Ch, 7 fig. 2 tab. 26 ref.] -
Key words:
- ecological stoichiometry /
- Setaria viridis /
- soil factors /
- climate factors /
- geographical gradient
-
图 1 狗尾草化学计量特征沿降水梯度带的变异系数及方差分量
Figure 1 Variation coefficient and variance components analysis of S. viridis stoichiometric traits along precipitation gradients
图 2 狗尾草化学计量特征在不同气候区域的差异
Figure 2 Differences in stoichiometric traits of S. viridis along different climate regions
图 3 狗尾草化学计量特征与地理因子(纬度、海拔)的关系
Figure 3 Relationships between ecological stoichiometric traits of S. viridis and geographical factors (latitude and elevation)
图 4 狗尾草化学计量特征的主成分分析
Figure 4 Principal component analysis of ecological stoichiometric traits in S. viridis
图 5 非生物因子与狗尾草化学计量特征主成分的分段结构方程模型
Figure 5 Segmented structural equation model of the relationships between abiotic factors and ecological stoichiometric traits PC1 and PC2 in S. viridis
图 6 土壤、环境因子对狗尾草化学计量特征变异的相对重要性
Figure 6 Relative importance of soil and environmental factors on stoichiometric trait variation in S. viridis
图 7 狗尾草根、茎和叶化学计量特征的主成分分析
Figure 7 Principal component analysis of root, stem, and leaf stoichiometric traits in S. viridis
表 1 研究点及其环境特征
Table 1. Description of study sites and their environmental characteristics
区域 研究点 纬度(N)/(°) 海拔/m 年平均气温/℃ 年均降水量/mm 光合有效辐射/ (mol·m−2·d−1) 湿润区 天目山 30.340 1 142.0 24.94 1 522.61 38.01 宣城 30.620 167.2 25.31 1 394.61 38.41 六安 31.533 353.2 25.22 1 134.24 40.03 信阳 32.130 393.1 24.74 1 082.29 40.16 半湿润区 三门峡 34.800 697.4 23.81 555.64 41.09 铜川 35.820 1 087.0 20.29 584.43 44.17 吴忠 37.980 1 811.5 20.50 235.59 49.76 半干旱/干旱区 张掖 38.930 1 408.2 19.55 131.72 50.26 酒泉 39.770 1 436.1 19.18 81.85 51.96 哈密 42.820 577.0 23.37 44.17 54.47  下载: 导出CSV
下载: 导出CSV
表 2 狗尾草化学计量特征主成分与环境因子的相关关系
Table 2. Correlations between principal components of stoichiometric traits in S. viridis with environmental factors
环境因子 根化学计量特征 茎化学计量特征 叶化学计量特征 PC1 PC2 PC1 PC2 PC1 PC2 年均降水量 0.13 0.010 0.21* 0.070 0.270** 0.004 5 年平均气温 0.10 0.002 0.32** 0.001 0.120 0.000 4 光合有效辐射 0.23** 0.030 0.10 0.030 0.220** 0.000 1 土壤全碳 0.20** 0.070 0.14* 0.040 0.030 0.050 0 土壤全氮 0.11 0.180* 0.04 0.010 0.050 0.010 0 土壤全磷 0.18* 0.170* 0.08 0.230** 0.006 0.240 0** 说明:*表示P<0.05;**表示P<0.01。
下载: 导出CSV
-
[1] SETH P, SEBASTIAN J. Plants and global warming: challenges and strategies for a warming world[J]. Plant Cell Reports, 2024, 43(1): 27. DOI: 10.1007/s00299-023-03083-w. [2] ELSER J J, BRACKEN M E S, CLELAND E E, et al. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems[J]. Ecology Letters, 2007, 10(12): 1135−1142. DOI: 10.1111/j.1461-0248.2007.01113.x. [3] 任书杰, 于贵瑞, 陶波, 等. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学, 2007, 28(12): 2665−2673. REN Shujie, YU Guirui, TAO Bo, et al. Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC[J]. Environmental Science, 2007, 28(12): 2665−2673. DOI: 10.3321/j.issn:0250-3301.2007.12.001. [4] XIONG Junlan, DONG Longwei, LU Jingli, et al. Variation in plant carbon, nitrogen and phosphorus contents across the drylands of China[J]. Functional Ecology, 2022, 36(1): 174−186. DOI: 10.1111/1365-2435.13937. [5] HONG Jiangtao, WANG Xiaodan, WU Jianbo. Stoichiometry of root and leaf nitrogen and phosphorus in a dry Alpine steppe on the Northern Tibetan Plateau[J]. PLoS One, 2014, 9(10): e109052. DOI: 10.1371/journal.pone.0109052. [6] HUO Chunlin, ZHANG Zhonghua, HU Gang, et al. Altitude-related variation in carbon, nitrogen, and phosphorus contents and their stoichiometry of woody organs in the subtropical mountain forests, South China[J]. Ecology and Evolution, 2025, 15(6): e71451. DOI: 10.1002/ece3.71451. [7] 郭美妙, 张博纳, 汤璐瑶, 等. 降水梯度带共有种旱柳功能性状的区域分异规律[J]. 浙江农林大学学报, 2025, 42(2): 281−290. GUO Meimiao, ZHANG Bona, TANG Luyao, et al. Regional differentiation patterns of common functional traits of Salix matsudana in precipitation gradient zones[J]. Journal of Zhejiang A&F University, 2025, 42(2): 281−290. DOI: 10.11833/j.issn.2095-0756.20240505. [8] HAN Wenxuan, FANG Jingyun, GUO Dali, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2): 377−385. DOI: 10.1111/j.1469-8137.2005.01530.x. [9] HE Mingzhu, ZHANG Ke, TAN Huijuan, et al. Nutrient levels within leaves, stems, and roots of the xeric species Reaumuria soongorica in relation to geographical, climatic, and soil conditions[J]. Ecology and Evolution, 2015, 5(7): 1494−1503. DOI: 10.1002/ece3.1441. [10] 丁潇然. 狗尾草叶水力系统与光合生理沿降水梯度带的调整与协同[D]. 杭州: 浙江农林大学, 2022. DING Xiaoran. Regulation and Coordination in Hydraulic System and Photosynthetic Physiology of Setaria viridis Along a Precipitation Gradient[D]. Hangzhou: Zhejiang A&F University, 2022. DOI: 10.27756/d.cnki.gzjlx.2022.000247. [11] 刘尧, 于馨, 于洋, 等. R程序包 “rdacca. hp” 在生态学数据分析中的应用: 案例与进展[J]. 植物生态学报, 2023, 47(1): 134−144. LIU Yao, YU Xin, YU Yang, et al. Application of “rdacca. hp” R package in ecological data analysis: case and progress[J]. Chinese Journal of Plant Ecology, 2023, 47(1): 134−144. DOI: 10.17521/cjpe.2022.0314. [12] 丁新新, 汤璐瑶, 张博纳, 等. 刺槐小枝功能性状沿降水梯度的适应策略及其驱动机制[J]. 浙江农林大学学报, 2025, 42(3): 503−512. DING Xinxin, TANG Luyao, ZHANG Bona, et al. Adaptive strategies of twig functional traits in Robinia pseudoacacia along a precipitation gradient and their driving mechanisms[J]. Journal of Zhejiang A&F University, 2025, 42(3): 503−512. DOI: 10.11833/j.issn.2095-0756.20240623. [13] 杨思琪, 赵旭剑, 森道, 等. 天山中段植物叶片碳氮磷化学计量及其海拔变化特征[J]. 干旱区研究, 2017, 34(6): 1371−1379. YANG Siqi, ZHAO Xujian, SEN Dao, et al. Leaf C, N and P chemometries and their altitudinal variations in the central Tianshan Mountains[J]. Arid Zone Research, 2017, 34(6): 1371−1379. DOI: 10.13866/j.azr.2017.06.20. [14] GÜSEWELL S. N∶P ratios in terrestrial plants: variation and functional significance[J]. The New Phytologist, 2004, 164(2): 243−266. DOI: 10.1111/j.1469-8137.2004.01192.x. [15] LUO Mengjiao, LIU Jiaxuan, QU Fanzhu, et al. Ecological stoichiometric characteristics and adaptive strategies of herbaceous plants in the Yellow River Delta wetland, China[J]. Biology, 2025, 14(9): 1132. DOI: 10.3390/biology14091132. [16] XU Hongwei, QU Qing, WANG Zhanhui, et al. Plant-soil-enzyme C-N-P stoichiometry and microbial nutrient limitation responses to plant-soil feedbacks during community succession: a 3-year pot experiment in China[J]. Frontiers in Plant Science, 2022, 13: 1009886. DOI: 10.3389/fpls.2022.1009886. [17] ZHANG Xinrui, ZHANG Lin, WANG Zhong, et al. Reviews and syntheses: ecological stoichiometry of carbon, nitrogen, and phosphorus in shrubs and shrublands[J]. EGUsphere, 2025, 848. DOI: 10.5194/egusphere-2025-848. [18] MEI Linlin, YANG Xue, CAO Hongbing, et al. Arbuscular mycorrhizal fungi alter plant and soil C∶N∶P stoichiometries under warming and nitrogen input in a semiarid meadow of China[J]. International Journal of Environmental Research and Public Health, 2019, 16(3): 397. DOI: 10.3390/ijerph16030397. [19] CHAVES M M, MAROCO J P, PEREIRA J S. Understanding plant responses to drought−from genes to the whole plant[J]. Functional Plant Biology, 2003, 30(3): 239−264. DOI: 10.1071/fp02076. [20] WRIGHT I J, REICH P B, WESTOBY M, et al. The worldwide leaf economics spectrum[J]. Nature, 2004, 428(6985): 821−827. DOI: 10.1038/nature02403. [21] REICH P B, WALTERS M B, ELLSWORTH D S. From tropics to tundra: global convergence in plant functioning[J]. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(25): 13730−13734. DOI: 10.1073/pnas.94.25.13730. [22] BROWN J H, GILLOOLY J F, ALLEN A P, et al. Toward a metabolic theory of ecology[J]. Ecology, 2004, 85(7): 1771−1789. DOI: 10.1890/03-9000. [23] KÖRNER C. Carbon limitation in trees[J]. Journal of Ecology, 2003, 91(1): 4−17. DOI: 10.1046/j.1365-2745.2003.00742.x. [24] FRESCHET G T, PAGÈS L, IVERSEN C M, et al. A starting guide to root ecology: strengthening ecological concepts and standardising root classification, sampling, processing and trait measurements[J]. New Phytologist, 2021, 232(3): 973−1122. DOI: 10.1111/nph.17572. [25] BARDGETT R D, MOMMER L, de VRIES F T. Going underground: root traits as drivers of ecosystem processes[J]. Trends in Ecology & Evolution, 2014, 29(12): 692−699. DOI: 10.1016/j.tree.2014.10.006. [26] REICH P B. The world-wide ‘fast-slow’ plant economics spectrum: a traits manifesto[J]. Journal of Ecology, 2014, 102(2): 275−301. DOI: 10.1111/1365-2745.12211. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250505
 点击查看大图
点击查看大图
计量
- 文章访问数: 384
- HTML全文浏览量: 149
- PDF下载量: 98
- 被引次数: 0
