








-
土壤动物是生态系统中分解者的重要组成部分,对土壤的形成、物质循环及能量流动具有重要意义,是土壤肥力的指标之一[1-4]。不同植被类型造成土壤理化性质的异质性[2],会导致生态系统内土壤动物多样性特征的差异[5]。雷竹Phyllostachys violascens为禾本科Poaceae竹亚科Bambusoideae刚竹属Phyllostachys竹种,原产浙西北丘陵平原地带[6],由于雷竹具出笋早、产量高、经济效益显著等特点,被各地广泛引种[7]。四川都江堰在“5·12”灾后重建过程中,为实现农业转型升级,当地政府引进并大力发展雷竹产业,实现了生态保护、农民增收和农村产业转型的目的,并带动当地旅游业的发展。相关学者对雷竹展开了广泛的研究,其研究主要集中在土壤碳库[8]、土壤微生物[9-10]、土壤养分[11-12]等,这些研究丰富了雷竹研究成果,对雷竹产业的发展具有重要意义。目前,有关雷竹的研究主要集中在原产地江浙一带,而有关雷竹异地引种后,对当地生态环境的影响却鲜有报道,因此,本研究选取农耕地和慈竹Neosinocalamus affinis林分别作为引种雷竹前的土地利用方式及乡土竹类进行研究,希望通过对比分析不同生境下的土壤动物群落特征,了解引种雷竹对当地土壤动物群落结构的影响,旨在为雷竹产业的可持续健康发展以及生物多样性保护提供科学依据。
HTML
-
试验地位于成都平原与四川盆周西缘山地接合部的丘陵地区——四川省都江堰市蒲阳镇向峨乡雷竹基地(31°05′07.91″N,103°44′18.71″E),属亚热带气候类型,海拔为759 m。据都江堰市气象局统计,区域内年平均气温为15.1 ℃,年平均降水量为1 225.4 mm,具有明显的春雨期、梅雨期和秋雨期,年平均相对湿度为81%,年平均日照时数为1 024.2 h,无霜期为269 d。样地土壤为砂岩上发育的黄壤,质地为重壤质,pH 6.5~6.8,土壤肥力中等,保肥保水性好。
都江堰向峨乡在5·12地震后,针对灾后重建和农业转型的需要,从浙江省临安市引种目前市场需求较大的雷竹,以期实现农民增收和生态环境建设的目的。该区域雷竹栽植地前期土地利用类型为农耕地,出笋期受人为采伐干扰,其他季节干扰小;慈竹为当地乡土竹类,呈散生丛状分布,轻度人为干扰,林分郁闭度高;农耕地前期农作物为玉米Zea mays,地表无凋落物。本次调查研究样地雷竹林下主要植被为喜旱莲子草Alternanthera philoxeroides,荩草Arthraxon hispidus,稗草Echinochloa crusgalli,常春藤Hedera nepalensis,野艾蒿Artemisia lavandulaefolia,爵床草Justicia procumbens;慈竹林下基本无植被覆盖,主要凋落物为慈竹叶;农耕地的植被以喜旱莲子草为主。各样地详细信息见表 1。
样地类型 海拔/m坡位 坡位 坡向 样地面积/m2 竹龄/a 栽植密度/(株·hm-2) 土壤含水量/% 干扰强度 平均林高/m 胸径/cm 雷竹 759 下 东南 400 5~6 1 650 14.04 中等 8 4.5 慈竹 762 下 东南 400 >20 5 000 16.74 弱 13 6.5 农耕地 756 下 东南 400 12.20 极强 Table 1. Basic conditions of the three plots
-
于2014年10月分别在雷竹、慈竹和农耕地采样,各样地面积设为20 m × 20 m。在各样地中心分别按“品”字型布点,设置3个50 cm × 50 cm(0.25 m2)的小样方,对凋落物层、0~5 cm层、5~10 cm层、10~15 cm层的土壤动物进行手捡,并将收集到的土壤动物放入盛有体积分数为75%的乙醇容器中杀死,对其编号分类并计数,带回实验室在解剖镜下鉴定;在各样点挖土壤剖面,分0~5 cm层、5~10 cm层、10~15 cm层用环刀(r=5 cm,v=100 cm3)自下往上顺次取土样,取土样2个·层-1,用尼龙网包好贴上标签装入黑布袋,带回实验室分别用Tullgren干漏斗和Baermann湿漏斗分离土样中的土壤动物。同时,在雷竹、慈竹林各样点收集10 cm × 10 cm(0.01 m2)面积的凋落物带回实验室进行分离,由于农耕地无地表凋落物,则只分离土壤层的土壤动物。土壤动物的分离均在烘虫箱中进行,控制烘虫箱温度在35~40 ℃。凋落物、干生和湿生的烘虫时间均为48 h。分离出的土壤动物除湿生外,均用盛有体积分数为75%的乙醇培养皿中收集,在解剖镜下观察计数;湿生土壤动物的收集则用清水。凋落物与干生土壤动物隔12 h观察1次,湿生土壤动物隔4 h观察1次,观察间隔时间逐步增长。对观察到的所有土壤动物进行分类并计数。
-
土壤动物的分类鉴定,土壤动物的分离参照肖玖金等[13]和黄玉梅等[14]的方法,将手捡和分离所得的土壤动物置于双目解剖镜(Leica,EZ4HD),主要采用《中国土壤动物检索图鉴》[15]《中国亚热带土壤动物》[16]《昆虫分类检索》[17]和《幼虫分类学》[18]进行分类鉴定,鉴定至纲、目等较高的分类阶元。
-
测定方法按照中国科学院南京土壤研究所编写的《土壤理化分析》进行[19]。
-
类群数量等级:个体数量大于捕获总量的10.0%以上者为优势类群(+++),占1.0%~10.0%者为常见类群(++),不足1.0%者为稀有类群(+)。
群落多样性分析:土壤动物多样性特征主要采用Shannon-Wiener多样性指数H′,Margalef丰富度指数D,Pielou均匀度指数J,Simpson优势度指数C和密度—类群指数IDG来计算[20]。
Shannon-Wiener多样性指数(H′):H′=-∑PilnPi。其中:Pi=ni/N,ni为第i个类群的个体数;N为所有类群的个体数。Margalef丰富度指数(D):D=(S-1)/lnN。其中:S为类群数,N为全部类群的个体总数。Pielou均匀度指数(J):J= H′/lnS。其中:H′为Shannon-Wiener多样性指数,S为类群数。Simpson优势度指数C:C=∑(ni/N)2。其中:ni为第i个类群的个体数;N为所有类群的个体数。密度-类群指数(IDG):IDG=g/G∑DiCi/DimaxC。其中:Di为第i类群个体数;Dimax 为各群落中第i 类群的最大个体数;g为群落中的类群数;G为各群落所包含的总类群数;Ci即在C个群落中第i个类群出现的比率。
群落相似性分析:采用Sorensen相似性指数(Q),即Q=2C/(A+B)。其中:C为2个群落或样地生境共有类群数,A和B分别为生境a和生境b的类群数。
数据的处理和分析采用Excel 2010和SPSS 22.0完成,采用Origin 8.1绘制图形。用单因素方差分析(one-way ANOVA)对不同样地间土壤动物群落组成进行检验。如果差异显著则用最小显著差(LSD)法(方差齐性)或Dunnetts’C法(方差不齐)进行多重比较。利用双因素方差分析(two-way ANOVA)检验土层分布以及样地类型对土壤动物群落结构和多样性的总体影响,显著性水平设定为α=0.05。方差分析时,对不服从正态分布的数据,利用log(x+1)转换,如果仍不服从正态分布,则进行Kruskal Wallis Test(H)非参数检验。
2.1. 调查方法与鉴定
2.1.1. 土壤动物的调查与分离
2.1.2. 土壤动物的鉴定
2.1.3. 土壤理化性质的测定
2.2. 数据分析与处理
-
本次研究调查共捕获土壤动物3 477只,隶属于3门9纲24目(表 2)。优势类群为线虫纲和线蚓科,分别占总体个数的62.26%和24.07%;常见类群为前气门亚目Prostigmata,中气门亚目Mesostigmata,等翅目Isoptera和膜翅目Hymenoptera,占总个数的10.89%;剩余甲螨亚目Oribatida,鞘翅目Coleoptera等构成稀有类群,其个体数占总个体数的2.78%。在雷竹样地中共捕获8纲23目1 816只土壤动物,平均密度3.47×105只·m-2;慈竹样地中捕获9纲24目1 263只土壤动物,平均密度9.65×104只·m-2;农耕地中捕获6纲12目398只土壤动物,平均密度4.58×104只·m-2。
类群 雷竹 慈竹 农耕地 合计 丰富度 平均密度/ (只·2) 百分比/ % 平均密度/ (只·2) 百分比/ % 平均密度/百分比/ (只.m-2) % 密度/(只.m-2) 百分比/ % 线虫纲Gematoda 238 783.78 68.75 37 518.13 38.87 28 520.57 62.32 304 822.47 62.26 +++ 线虫引科Enchytaeidae 71 301.41 20.53 30 251.55 31.34 16 297.47 35.61 117 850.43 24.07 +++ 前气门亚目Prostigmata 16 433.33 4.73 13 600.00 14.09 - - 30 033.33 6.13 ++ 中气门亚目Mesostigmata 5 700.00 1.64 4 700.00 4.87 - - 10 400.00 2.12 ++ 等翅目Isoptera 3 703.10 1.07 3 313.33 3.43 - - 7 016.43 1.43 ++ 膜翅目 Hymenoptera 3 881.97 1.12 1 774.59 1.84 220.00 0.48 5 876.56 1.20 ++ 甲螨亚目Oribatida 1 527.89 0.44 633.33 0.66 - - 2 161.22 0.44 + 鞘翅目 Coleoptera 731.96 0.21 925.33 0.96 - - 1 657.30 0.34 + 蜘蛛目Araneae 1 072.83 0.31 180.43 0.19 36.00 0.08 1 289.26 0.26 + 缨翅目 Thysanoptera 1 133.33 0.33 66.67 0.07 - - 1 200.00 0.25 + 双翅目(幼'Oiptera 266.67 0.08 252.00 0.26 339.53 0.74 858.20 0.18 + 鞘翅目(幼)Coleoptera 500.00 0.14 309.33 0.32 4.00 0.01 813.33 0.17 + 双尾目Diplura 33.33 0.01 566.67 0.59 - - 600.00 0.12 + 综合纲Symhpyla 169.77 0.05 411.53 0.43 - - 581.30 0.12 + 弹尾目 Collembola 333.33 0.10 233.33 0.24 - - 566.67 0.12 + 啮目 Psocoptera 376.86 0.11 174.67 0.18 - - 551.53 0.11 + 鱗翅目(幼)Lepidoptera 33.33 0.01 508.43 0.53 1.33 0.00 543.10 0.11 + 革翅目 Deramptera - - 362.20 0.38 176.43 0.39 538.63 0.11 + 盲蜘目Opiliones 366.67 0.11 100.00 0.10 - - 466.67 0.10 + 輩蠊目 Rlattoptera 401.33 0.12 41.33 0.04 2.67 0.01 445.33 0.09 + 等足目Isopoda 277.33 0.08 77.33 0.08 56.00 0.12 410.67 0.08 + 寡毛纲 Oligochaeta 78.67 0.02 52.00 0.05 85.33 0.19 216.00 0.04 + 唇足纲Chilopoda 169.77 0.05 - - - - 169.77 0.03 + 双翅目Diptera - - 169.77 0.18 - - 169.77 0.03 + 啮 虫 目 Corrodentia - - 133.33 0.14 - - 133.33 0.03 + 鳞翅目 Lepidoptera - - 66.67 0.07 - - 66.67 0.01 + 娱蚣目 Scolopendromorpha 21.33 0.01 20.00 0.02 12.00 0.03 53.33 0.01 + 直翅目 Orthoptera 4.00 0.00 33.33 0.03 4.00 0.01 41.33 0.01 + 倍足目Diplopoda - - 40.00 0.04 - - 40.00 0.01 + 裂盾目 Schizomida 33.33 0.01 - - - - 33.33 0.01 + 地娱蚣目 Teophilomorpha 4.00 0.00 4.00 0.00 4.00 0.01 12.00 0.00 + 石娱蚣目 Lithobiomorpha 1.33 0.00 2.67 0.00 6.67 0.01 10.67 0.00 + 半翅目Hemiptera 2.67 0.00 - - - - 2.67 0.00 + 平均密度/(只_m-2) 347 343.33 96 521.96 45 765.99 489 631.29 总类群数/个 28 30 15 33 说明:“-”表示无或未采集到,+++为优势类群,++为常见类群,+为稀有类群。 Table 2. Compositions of soil fauna community in the three plots
-
3种类型样地的土壤动物水平分布特征显示:雷竹样地土壤动物平均密度显著高于慈竹和农耕地(P<0.05),类群数高于农耕地,低于慈竹样地(P<0.05)(表 2)。方差分析结果显示:各样地间土壤动物平均密度与类群数均存在显著差异(P<0.05)。
参照尹文英[15]对土壤动物体型的划分,将本次研究调查到的土壤动物分为大型和中小型两大类。从图 1可以看出:雷竹样地大型和中小型土壤动物平均密度均显著高于慈竹和农耕地(P<0.05),而类群数高于农耕地,显著低于慈竹(P<0.05)。对大型和中小型土壤动物群落特征进行方差分析,结果显示:各样地大型土壤动物的平均密度和类群数均无显著性差异(P>0.05),中小型土壤动物的平均密度和类群数均存在显著性差异(P<0.05)。

Figure 1. Horizontal distribution of soil fauna in the three habitats
-
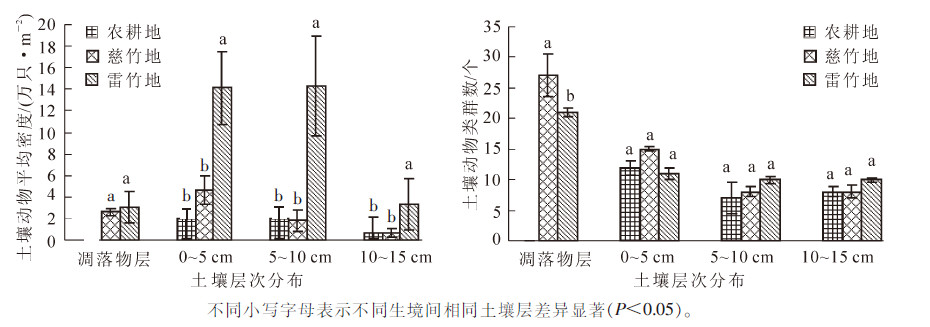
不同的土壤深度,由于含水量、有机质等不同,其土壤动物群落的垂直分布也不同[20-22]。从图 2可以看出:雷竹样地中各层土壤动物的平均密度均高于慈竹和农耕地,其中,雷竹和慈竹凋落物层的平均密度差异达到显著水平(P<0.05);雷竹样地中凋落物层和0~5 cm层土壤动物的类群数低于慈竹,5 cm以下土层雷竹样地高于慈竹与农耕地,其中各样地凋落物层的类群数差异达到显著水平(P<0.05)。

Figure 2. Vertical distribution of soil fauna in each layer of three habitats
3种类型样地中土壤动物的类群数均表现为土壤表层具有较高的类群数,在大于5 cm土壤深度,类群数减少速度趋于平缓;雷竹和慈竹样地中,土壤动物均集中分布在凋落物层。3种类型样地的土壤动物群落分布特征基本满足以下规律:从地表向下,随土层深度的增加,土壤动物数量和类群多样性逐渐减少,并且减少的速度越来越快。
双因素方差分析结果显示(表 3):土层分布对土壤动物的群落结构具有极显著影响(P<0.01),在土壤动物多样性方面,土层分布对密度-类群指数和Margalef丰富度指数具有极显著影响(P<0.01),Shannon-Wiener多样性指数具有显著影响(P<0.05);样地类型对土壤动物群落结构和多样性特征影响水平低于土层分布;样地类型和土层分布的交互作用仅对密度-类群指数有显著影响(P<0.05)。
项目 个体数 类群数 H' J C IDG D 样地类型 1.366 0.836 0.422 0.843 0.554 0.452 1.259 土层 22.518** 34.758** 4.429* 1.124 0.818 47.731** 10.653** 样地类型×土层 0.142 1.744 0.555 0.391 0.351 2.684* 1.188 说明: H'为Shannon-Wiener多样性指数,J为Pielou均匀度指数,C为Simpson优势度指数,IDG为密度-类群指数,D为Margalef丰富度指数。*表示存在显著相关性(P<0.05); **表示存在极显著相关性(P <0.01)。 Table 3. Two-way ANOVA of soil fauna diversity (F)
-
在3种类型的样地中,土壤动物Shannon-Wiener多样性指数表现为雷竹与慈竹、农耕地间均无显著差异(P>0.05),但慈竹与农耕地间存在显著差异(P<0.05);Pielou均匀度指数与Simpson优势度指数表现为各样地间均无显著差异(P>0.05);雷竹与慈竹样地土壤动物的密度-类群指数无显著差异(P>0.05),但雷竹、慈竹与农耕地间存在显著差异(P<0.05);3种类型样地土壤动物Margalef丰富度指数均存在显著性差异(P<0.05,表 4)。
样地类型 H' J C IDG D 雷竹 1.99±0.19ab 0.68±0.06 a 0.22±0.06 a 4.37±0.81a 2.82±0.05 b 慈竹 2.22±0.22 a 0.71±0.08 a 0.19±0.07 a 4.70±0.60 a 3.64±0.50 a 农耕地 1.75±0.05 b 0.77±0.08 a 0.25±0.05 a 0.97±0.29 b 1.84±0.37 c 说明: H'为Shannon-Wiener多样性指数,J为Pielou均匀度指数,C为Simpson优势度指数,IDG为密度-类群指数,D为Mar-galef丰富度指数。同行不同小写字母表示不同生境间差异显著(P<0.05)。 Table 4. Richness, diversity, evenness and dominance of soil fauna in the three habitats
-
不同的立地类型其生境条件不同,必然导致土壤动物的类群分布以及多样性不同,但各生境往往又表现出具有一定的相似性[23]。本研究中,采用Sorensen相似性指数,对3种生境土壤动物类群的相似性指数进行了对比。从表 5可以看出:作为竹类,雷竹与慈竹样地的土壤动物类群具有较高的相似性,且两者间土壤动物相似性指数达到了0.8,而雷竹与农耕地、慈竹与农耕地的相似性则较低。
样地类型 雷竹 慈竹 农耕地 雷竹 1 慈竹 0.814 1 农耕地 0.651 0.652 1 Table 5. Composition index of soil fauna group in the three habitats
3.1. 土壤动物的组成
3.2. 土壤动物群落的水平分布特征
3.3. 土壤动物群落的垂直分布特征
3.4. 土壤动物的多样性特征
3.5. 土壤动物类群的相似性分析
-
本次研究调查中,雷竹林和慈竹林土壤动物优势类群数量分别占总捕获量的89.27%和84.30%,而农耕地中优势类群占总捕获量的97.93%,表明与雷竹和慈竹林相比,农耕地土壤动物数量集中在少数类群上,而雷竹和慈竹林受到的人为干扰小,土壤动物各类群数量较为均匀;研究发现,农耕地中未捕获到蜱螨目Arachnoidea,这除了与农耕地周期性的扰动影响有关外,地表凋落物的缺失也是其主要原因[24]。
雷竹林土壤动物平均密度均显著高于慈竹林和农耕地(P<0.05),这可能与雷竹的引种改变了耕地植物多样性低及凋落物缺乏的状态有关。同时,雷竹栽植后,土壤的扰动减少,土壤的含水量和有机质增加,使得以线虫为主的湿生土壤动物数量大幅度上升[25],其中雷竹林土壤线虫数量分别是慈竹林和农耕地的6.4倍和8.4倍,线蚓数量分别是慈竹林和农耕地的2.4倍和4.4倍。
各样地土壤动物密度剖面及类群分布具有明显的表聚性特征,这与大多数研究结果一致[26-27],同时,受多种因素的影响,土壤动物偶尔呈逆层分布的现象。本研究中,形成这种逆分布现象的原因可能与农耕地的耕作活动有关,导致农耕地10~15 cm土层中土壤动物类群数较5~10 cm高,另外,雷竹地下茎属单轴散生型[28],其散生根系主要分布在0~10 cm层中,从而导致雷竹林土壤动物平均密度在0~5 cm和5~10 cm层较高。
土壤动物的多样性,不仅是生态环境评价的重要指标,还能反映群落的稳定性,因此,研究土壤动物的多样性对于整个生态系统的研究具有重要的意义[29-30]。多样性指数反映群落组成的复杂程度,均匀度指数则能反映各物种个体数目分配的均匀程度,较高的多样性指数和均匀度指数表明该生态系统更长的食物链以及更为复杂的种间关系[20, 31-32]。对各样地土壤动物多样性指数结果分析显示,雷竹和慈竹样地的Shannon-Wiener多样性指数、Margalef丰富度指数均较农耕地高,表明退耕还竹后,竹林生态系统较农耕地能为土壤动物提供了更长的食物链,出现了更多的共生现象,一定程度上提高了土壤动物的多样性和丰富度。雷竹样地的密度-类群指数(IDG)高于农耕地,说明雷竹林生境内土壤动物多度增长的潜力大,雷竹的引种栽培对于提高该地区土壤动物群落多度具有重要作用。3种生境中,雷竹林土壤动物的Simpson优势度指数介于慈竹林和农耕地间,表明雷竹引种后相对于原始迹地农耕地来说,有利于土壤动物群落各类群的生存,群落的优势度越小,生态系统中群落越复杂,且种群越多,群落的优势度也越小[30]。
土壤动物类群的相似性分析表明,雷竹和慈竹样地的相似性最高(为0.814),高于雷竹和慈竹样地与农耕地的相似性(分别为0.651和0.652),这主要是由于雷竹和慈竹具有相似的经营方式和植被生态系统,而农耕地受到人为干扰的强度和频度均较高,导致雷竹和慈竹间具有较高的相似性。
综上所述,就目前而言,引种雷竹进行退耕还竹极大地增加了土壤动物类群数和个体数,雷竹的引种对该地区土壤动物群落特征产生了一定影响。本研究通过与慈竹和农耕地的对照,初步讨论了雷竹的引种对土壤动物群落结构的影响,为了更加准确地掌握雷竹的引种对当地生态环境的影响,还应该加强对雷竹栽植密度、不同经营强度及栽植年限等因素对土壤动物群落结构的影响研究。
 DownLoad:
DownLoad: