







-
光照是植物生存和生长发育最重要的环境因子之一,光照通过改变植物生长的环境因子影响植物光合特性,进而影响植物对碳的吸收和积累[1]。光照过强会导致光抑制,限制植物光合作用,损害植物光合机构,而光照不足也会限制植物光合作用,进而导致植物由于碳饥饿而死亡[2-3]。在遮光条件下,植物会通过降低光补偿点、光饱和点和暗呼吸速率,增大表观量子效率等光合参数来进行光合作用,而最大羧化速率和最大电子传递速率则有所下降[4-5]。落叶植物在春季叶片新生,形态结构逐渐完善,而夏季较高的光照强度和气温会导致植物气孔关闭,使植物产生光合午休现象,秋季叶绿素减少导致光合作用减弱[6]。叶绿素荧光参数能直接反映植物光合作用的实际与最大光合效率、反应中心的开放程度以及热耗散的变化,从而分析对叶片光合机构内部的影响和生理调节能力[7]。如全光照条件下的堇叶紫金牛Ardisia violacea光系统Ⅱ(PS Ⅱ)最大光化学量子产量(Fv/Fm)小于0.80,受到光照胁迫,而遮光处理下没有受到光照胁迫[4]。因此,了解不同季节不同光照强度下植物光合特性对植物分布和保护具有重要意义[8]。
景宁木兰Magnolia sinostellata为木兰科Magnoliaceae木兰属Magnolia落叶灌木或小乔木,种群数量较少,被列为浙江省重点保护野生植物名录(第1批)[9]。景宁木兰分布区极其狭窄,主要分布于中国浙江南部的丽水市、温州市,在海拔900 m以上的灌丛、黄山松Pinus taiwanensis林、落叶阔叶林、杉木Cunninghamia lanceolata林和林缘等生境生长,所处生态环境郁闭度不同会导致景宁木兰所受的光照强度有所差异[10]。因此,本研究通过设置不同遮光处理,探讨不同光照强度下景宁木兰幼苗叶片光合特性的季节性变化,阐明幼苗在不同生长季节对不同光照强度的适应机制和利用对策,为景宁木兰的迁地保护、种群的繁衍复壮以及人工繁育提供理论依据。
-
景宁木兰幼苗由浙江景宁县草鱼塘林场提供。于2020年1月上旬,将长势相似的2年生景宁木兰幼苗移栽到花盆中,每盆添加由V(泥炭土)∶V(水稻土)∶V(河沙)=6∶6∶1混合而成的栽培土壤9.00 kg。土壤pH为6.19,有机质为103.51 mg·g−1,全氮为2.85 mg·g−1,速效磷为0.04 mg·g−1,全磷为0.48 mg·g−1,速效钾为1.16 mg·g−1,全钾为8.40 mg·g−1。2020年3月上旬,设置全光照(光照强度100%,I100)为对照,通过搭建不同层数的黑色遮阳网(规格6针)进行遮光处理,分别是1层遮阳网覆盖(光照强度约40%全光照,I40)和2层遮阳网覆盖(光照强度约10%全光照,I10)。研究在浙江台州学院临海校区试验基地(28°87′N,127°17′E)进行。
-
于2020年4月(春季)、7月(夏季)、10月(秋季)中旬测定景宁木兰幼苗光合参数,选择晴朗的天气,从6:00—18:00,通过Li-6400XT便携式光合作用仪(Li-Cor,美国)透明叶室测定景宁木兰幼苗叶片的光合指标,每隔2 h测定1次。每株测定从上往下的第3对功能叶,每个处理3个重复。测定参数为净光合速率(Pn,µmol·m−2·s−1)、气孔导度(Gs,mol·m−2·s−1)、胞间二氧化碳摩尔分数(Ci,µmol·mol−1)和蒸腾速率(Tr,mmol·m−2·s−1)[11]。
景宁木兰幼苗光响应曲线使用红蓝光源叶室,设定诱导光照强度为2 000 µmol·m−2·s−1,诱导约15~30 min。设置叶室温度为25 ℃,流速控制为500 µmol·m−2·s−1,相对湿度为55%~65%,以二氧化碳(CO2)钢瓶内液态CO2为气源。设置光合有效辐射梯度由高到低分别为2 000、1 500、1 200、1 000、800、600、400、200、150、100、50、20、0 µmol·m−2·s−1[11]。
景宁木兰幼苗CO2响应曲线设定诱导光照强度为1 000 µmol·m−2·s−1,其余条件与光响应条件相同。设置CO2摩尔分数梯度由高到低分别为1 500、1 200、1 000、800、600、400、300、200、150、100、80、50 µmol·mol−1[11]。
-
使用MINI-PAM叶绿素荧光便携式测定仪(MINI-PAM,WALZ,德国)测定叶片充分暗适应30 min后的叶绿素荧光参数。由仪器测定软件直接给出PS Ⅱ最大光化学量子产量(Fv/Fm)、PS Ⅱ实际光化学量子产量(YⅡ)、光化学猝灭系数(qP)、非光化学猝灭系数(qNP)、PS Ⅱ非调节性能量耗散的量子产量(YNO)、PS Ⅱ调节性能量耗散的量子产量(YNPQ)、电子传递速率(ETR)[12]。
-
通过光合3.4.2软件进行光响应曲线和CO2响应曲线的拟合,模型为直角双曲线的修正模型,用Photosynthesis软件获得光响应和CO2响应参数[13-14]。
采用Excel对所测数据进行整理。利用SPSS 21.0软件中的单因素方差分析和Duncan新复极差法比较不同处理间的差异。用双因素方差分析比较光照强度和季节变化对景宁木兰的交互影响。绘图使用Origin 9.1软件。数据均为平均值±标准误。
-
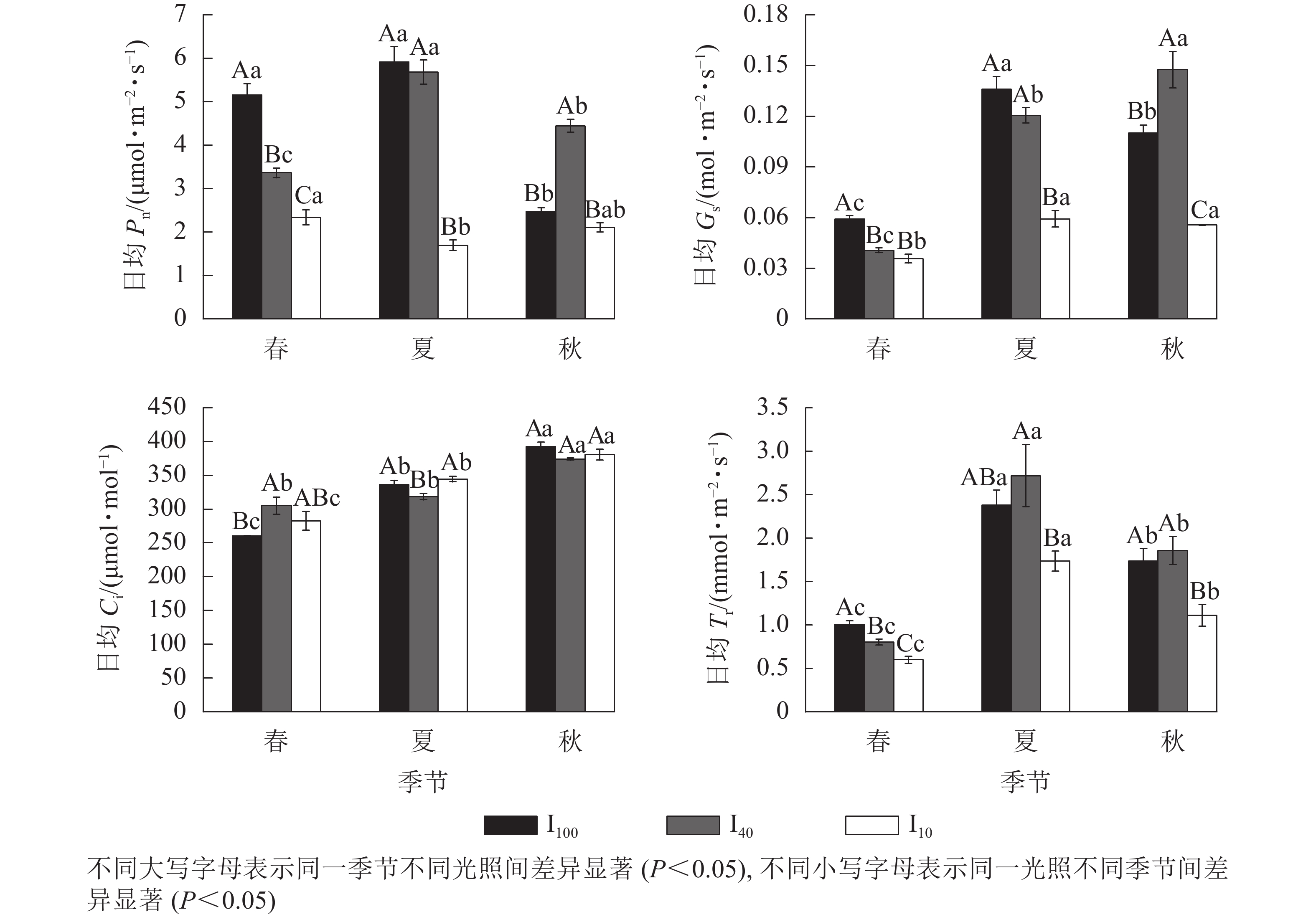
春季、夏季100%全光照(I100)以及夏季40%全光照(I40)下景宁木兰幼苗净光合速率(Pn)日变化均呈“双峰”曲线,第1峰值出现在10:00,在12:00时到达波谷,14:00时出现第2峰值,具有“光合午休”现象,其余Pn日变化均为“单峰”曲线,峰值均在12:00(图1)。春季日均Pn从大到小依次为100%全光照、40%全光照、10%全光照,均达到差异显著水平(P<0.05);夏季10%全光照的日均Pn显著低于100%全光照、40%全光照(P<0.05);秋季40%全光照的日均Pn显著高于100%全光照、10%全光照(P<0.05)(图2)。100%全光照下秋季的日均Pn显著低于春季、夏季(P<0.05);40%全光照下的日均Pn从大到小依次为夏季、秋季、春季,均差异显著(P<0.05);10%全光照下的日均Pn为春季显著高于夏季(P<0.05) (图2)。

Figure 1. Seasonal change of the diurnal changes of net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular carbon dioxide mole fraction (Ci), transpiration rate (Tr) of M. sinostellata seedlings under three light intensities
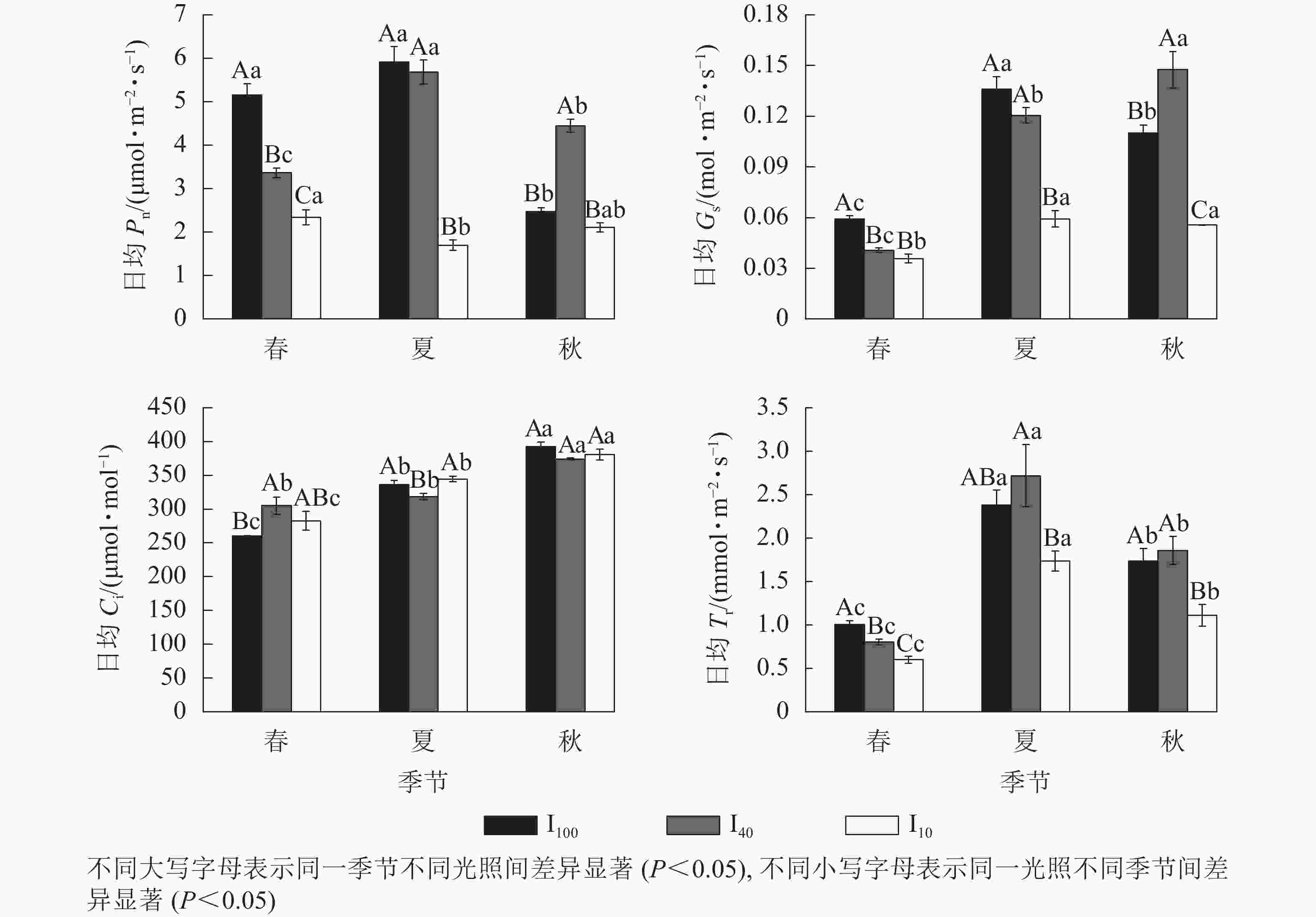
Figure 2. Seasonal change of the diurnal mean values of net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular carbon dioxide mole fraction (Ci), transpiration rate (Tr) of M. sinostellata seedlings under three light intensities
春季、夏季100%全光照和40%全光照下景宁木兰幼苗气孔导度(Gs)日变化均为“双峰”曲线,第1、第2峰值分别在10:00和14:00,波谷在12:00,其余Gs日变化均为“单峰”曲线,其中春季和夏季10%全光照下Gs在10:00达到峰值,而秋季3个光照处理均在12:00达到峰值(图1)。春季100%全光照的日均Gs显著高于遮光处理(P<0.05);夏季10%全光照的日均Gs显著低于100%全光照、40%全光照(P<0.05);秋季的日均Gs从大到小依次为40%全光照、100%全光照、10%全光照,均差异显著(P<0.05)。100%全光照下日均Gs从大到小依次为夏季、秋季、春季,均差异显著(P<0.05);40%全光照下日均Gs从大到小依次为秋季、夏季、春季,均差异显著(P<0.05);10%全光照下春季日均Gs显著低于夏季、秋季(P<0.05) (图2)。
景宁木兰幼苗胞间二氧化碳摩尔分数(Ci)日变化趋势呈现“V型”曲线。春季100%全光照、40%全光照、10%全光照下Ci曲线最低点分别出现在16:00、12:00、14:00;夏季100%全光照、40%全光照、10%全光照下Ci最低点分别出现在12:00、14:00、10:00;秋季3种光照强度下Ci最低点均在12:00 (图1)。春季100%全光照的日均Ci显著低于40%全光照(P<0.05);夏季40%全光照下日均Ci显著低于100%全光照和10%全光照(P<0.05)。在100%全光照和10%全光照下的日均Ci从大到小依次为秋季、夏季、春季,均差异显著(P<0.05);40%全光照下日均Ci秋季显著高于春季、夏季(P<0.05) (图2)。
春季和秋季,景宁木兰幼苗蒸腾速率(Tr)日变化在3种光照强度下均呈“单峰”曲线,其中春季100%全光照和40%全光照下Tr峰值在14:00达到最高值,其余均在12:00达到最高值(图1)。夏季,100%全光照和40%全光照为“双峰”曲线,其中:100%全光照的峰值在10:00、14:00,波谷在12:00;40%全光照的峰值在12:00、16:00,波谷在14:00。10%全光照为“单峰”曲线,峰值在12:00。春季日均Tr从大到小依次为100%全光照、40%全光照、10%全光照,均差异显著(P<0.05);夏季40%全光照下日均Tr显著高于10%全光照(P<0.05);秋季10%全光照下日均Tr显著低于春季、夏季(P<0.05)。不同季节3种光照处理下日均Tr从大到小依次为夏季、秋季、春季,均差异显著(P<0.05) (图2)。
-
从图3可见:夏季10%全光照下的最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)、暗呼吸速率(Rd)均显著小于100%全光照、40%全光照(P<0.05),而表观量子效率(AQY)随着光照强度增加不断下降,3个处理间差异显著(P<0.05)。秋季遮光处理下Pnmax、LSP显著高于100%全光照(P<0.05);100%全光照下LCP、Rd显著大于10%全光照(P<0.05);10%全光照AQY显著大于100%全光照(P<0.05)。100%全光照下Pnmax、LSP从大到小依次为夏季、春季、秋季,均差异显著(P<0.05)。40%全光照下LSP在夏季显著高于春季(P<0.05);夏季LCP、Rd显著高于春季、秋季(P<0.05)。10%全光照下秋季LSP显著高于春季,夏季AQY显著高于春季(P<0.05)。
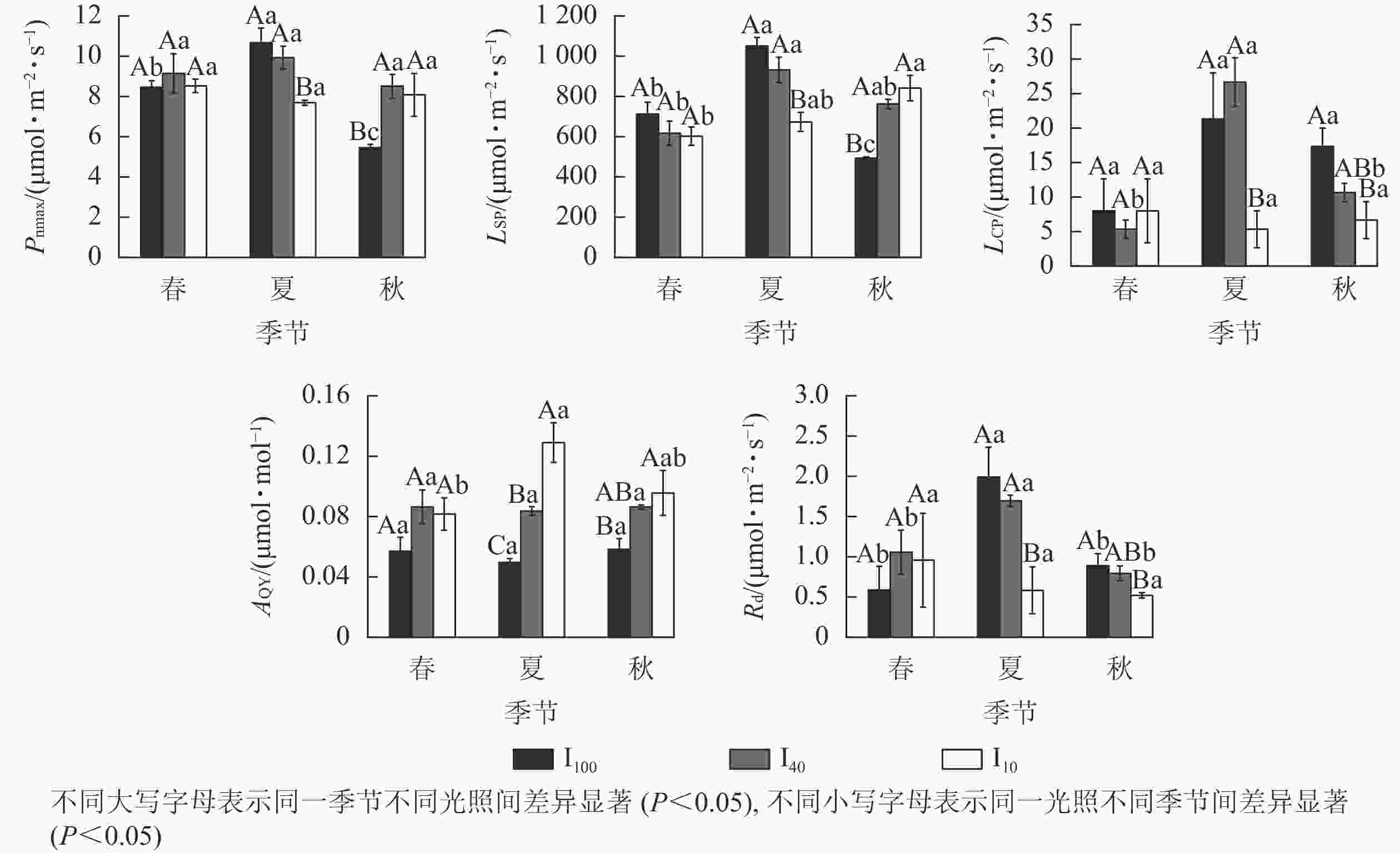
Figure 3. Seasonal change of the light response parameters of M. sinostellata seedlings under three light intensities
-
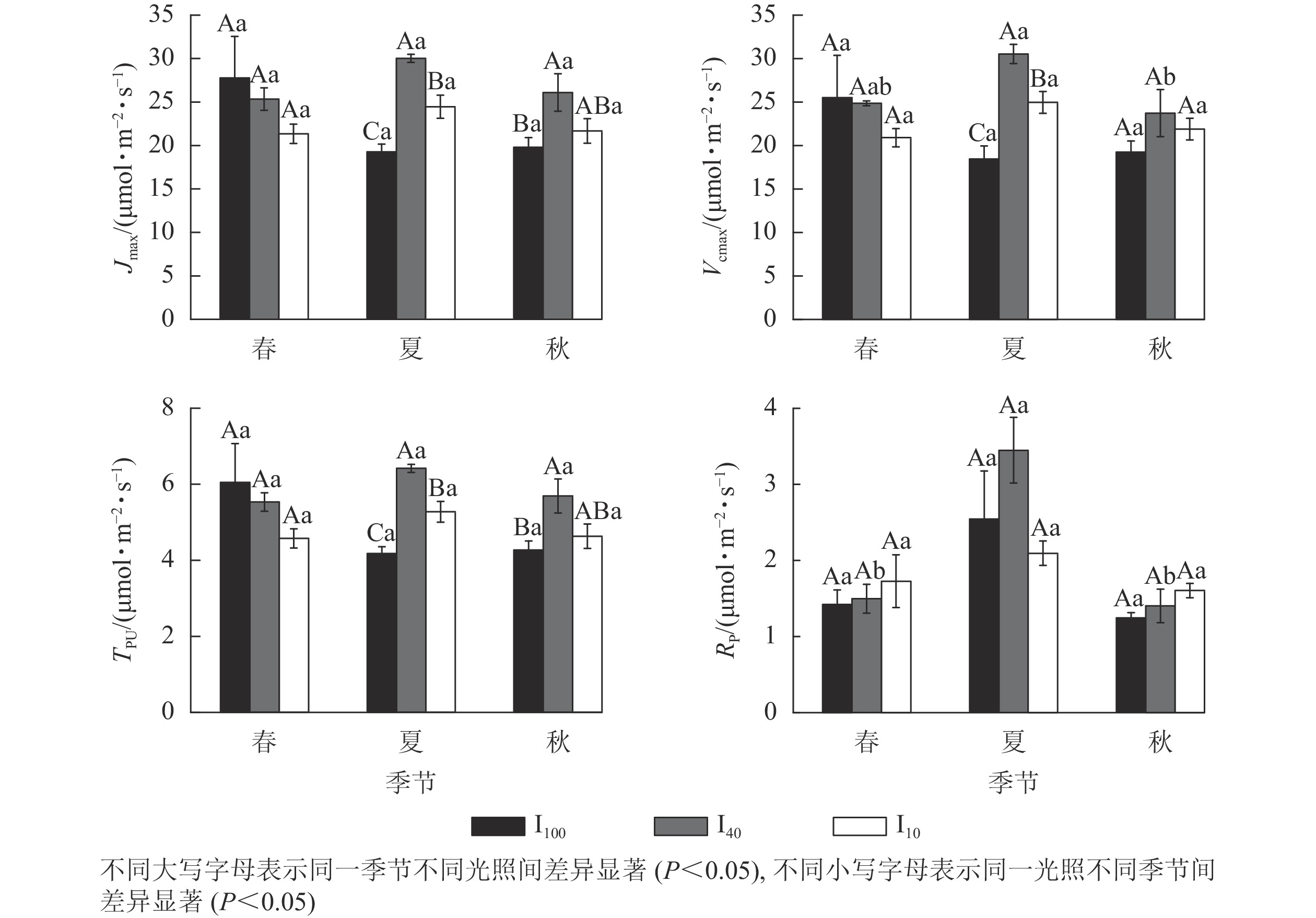
夏季景宁木兰幼苗叶片的最大电子传递速率(Jmax)、最大羧化速率(Vcmax)、磷酸丙糖利用率(TPU)从大到小依次为40%全光照、10%全光照、100%全光照,均差异显著(P<0.05)(图4)。秋季40%全光照下的Jmax和TPU均显著高于100%全光照(P<0.05)。夏季40%全光照下的Vcmax显著高于秋季,光呼吸速率(Rp)显著高于春季、秋季(P<0.05)。
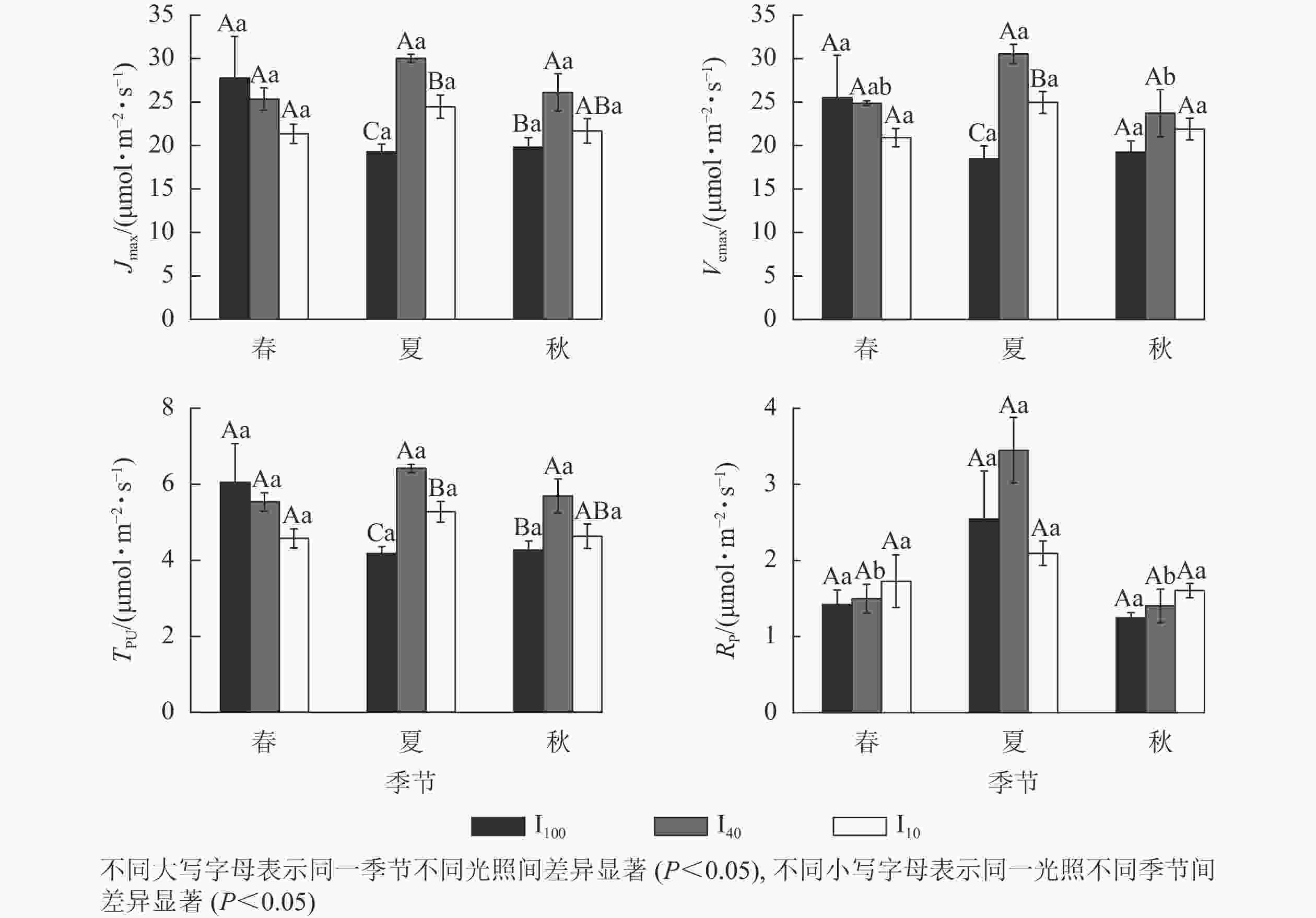
Figure 4. Seasonal change of the CO2 response parameters of M. sinostellata seedlings under three light intensities
-
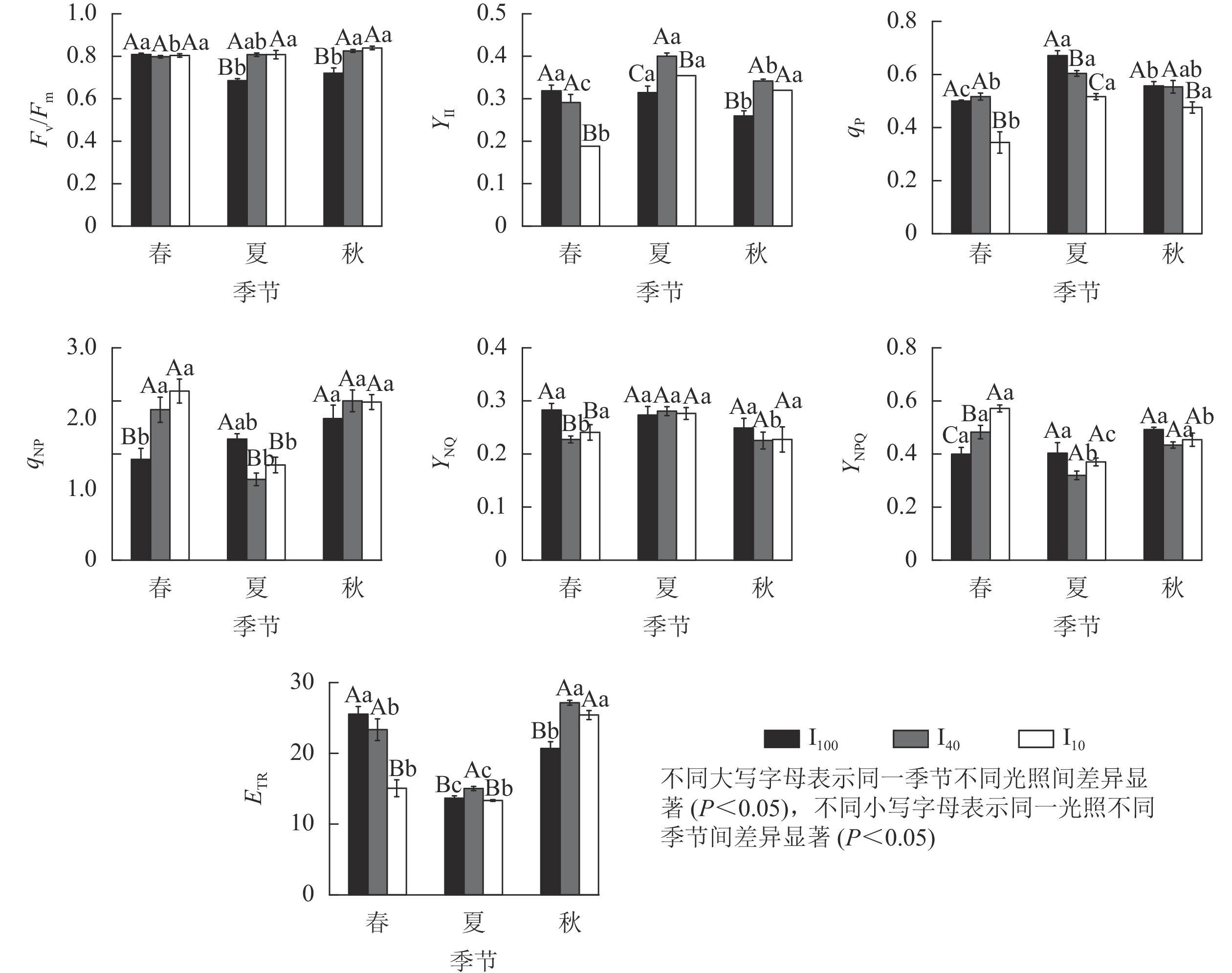
由图5可知:春季,10%全光照下景宁木兰幼苗PS Ⅱ实际光化学量子产量(YⅡ)、光化学猝灭系数(qP)、电子传递速率(ETR)均显著低于100%全光照和40%全光照(P<0.05);100%全光照下的非光化学猝灭系数(qNP)、PS Ⅱ调节性能量耗散的量子产量(YNPQ)显著低于遮光处理(P<0.05);100%全光照下的PS Ⅱ非调节性能量耗散的量子产量(YNO)显著高于遮光处理(P<0.05)。夏季,100%全光照下PS Ⅱ最大光化学量子产量(Fv/Fm)显著低于遮光处理(P<0.05);YⅡ、ETR在40%全光照下显著高于其他2种光照处理(P<0.05);qP、qNP在100%全光照下显著高于遮光处理(P<0.05)。秋季,100%全光照下的Fv/Fm、YⅡ、ETR均显著低于遮光处理(P<0.05);qP在10%全光照下最低,与其他2种光照处理差异显著(P<0.05)。
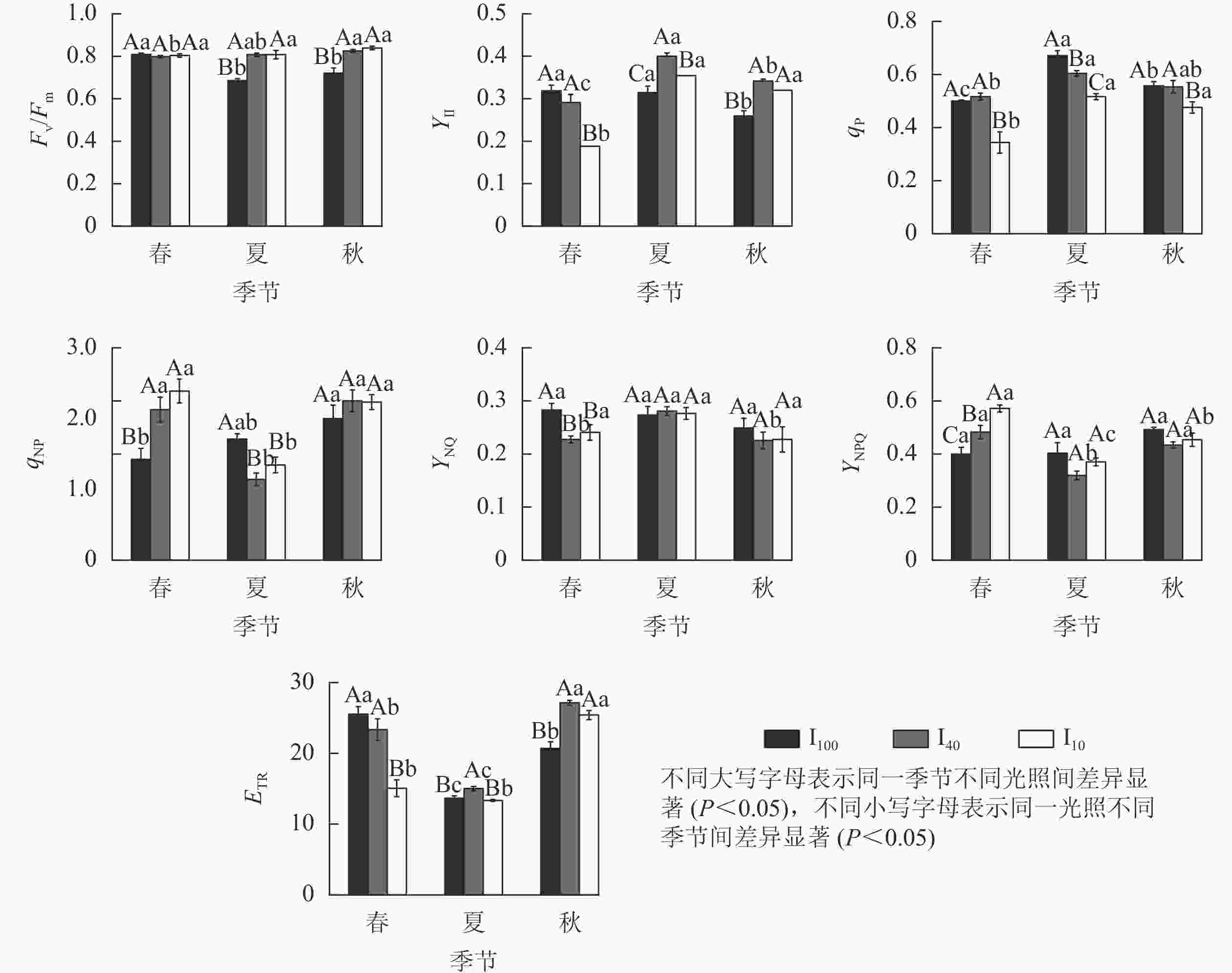
Figure 5. Seasonal change of chlorophyll fluorescence parameters of M. sinostellata seedlings under three light intensities
100%全光照下,景宁木兰幼苗叶片的Fv/Fm在春季显著高于夏季、秋季(P<0.05);YⅡ在秋季则显著低于春季、夏季(P<0.05);qP从大到小依次为夏季、秋季、春季,均差异显著(P<0.05);qNP在秋季显著高于春季(P<0.05);ETR从大到小依次为春季、秋季、夏季,均差异显著(P<0.05)。40%全光照下,秋季的Fv/Fm显著高于春季(P<0.05);YⅡ从大到小依次为夏季、秋季、春季,均差异显著(P<0.05);qP在夏季显著高于春季(P<0.05);qNP在夏季显著低于春季、秋季(P<0.05);YNO在夏季显著高于春季和秋季(P<0.05);YNPQ在夏季显著低于春季和秋季(P<0.05);ETR从大到小依次为秋季、春季、夏季,均差异显著(P<0.05)。10%全光照下春季YⅡ、qP均显著低于夏季和秋季(P<0.05);qNP在夏季显著低于春季、秋季(P<0.05);YNPQ从大到小依次为春季、秋季、夏季,均差异显著(P<0.05);秋季的ETR显著高于春季、夏季(P<0.05)。
-
双因素方差分析(表1)表明:光照强度、季节以及光照强度和季节之间的相互作用对景宁木兰幼苗日均Pn、Gs、Fv/Fm、YⅡ、ETR均有极显著影响(P<0.01),光照强度与光照强度和季节之间的相互作用对LCP、Jmax、TPU有显著影响(P<0.05),而季节与光照强度和季节之间的相互作用对日均Ci、Pnmax、LSP、qNP、YNPQ有极显著影响(P<0.01)。表明不同季节,景宁木兰幼苗叶片日均Pn、Gs、Ci、Pnmax、LSP、Fv/Fm、YⅡ、qNP、YNPQ、ETR等对光照强度变化的响应极显著,LCP、Jmax、TPU等对光照强度变化的响应显著。
参数 F 光照强度 季节 光照强度×季节 日均净光合速率(Pn) 185.425** 44.568** 48.552** 日均气孔导度(Gs) 98.029** 128.380** 21.942** 日均胞间二氧化碳摩尔分数(Ci) 0.468 123.368** 6.179** 日均蒸腾速率(Tr) 13.941** 62.008** 1.589 最大净光合速率(Pnmax) 2.791 8.433** 5.270** 光饱和点(LSP) 1.386 19.936** 14.684** 光补偿点(LCP) 4.986* 6.229** 3.100* 表观量子效率(AQY) 19.642** 1.347 2.857 暗呼吸速率(Rd) 2.753 4.728* 2.465 最大羧化速率(Vcmax) 4.986* 1.619 2.843 最大电子传递速率(Jmax) 5.642* 1.194 3.371* 磷酸丙糖利用率(TPU) 6.043* 1.259 3.426* 光呼吸速率(Rp) 1.261 15.372** 2.129 PS Ⅱ最大光化学量子产量(Fv/Fm) 36.645** 7.022** 11.011** PS Ⅱ实际光化学量子产量(YⅡ) 19.154** 42.680** 19.568** 光化学猝灭系数(qP) 36.964** 37.809** 2.623 非光化学猝灭系数(qNP) 2.855 23.553** 7.280** PS Ⅱ非调节性能量耗散的量子产量(YNO) 2.210 6.257** 1.164 PS Ⅱ调节性能量耗散的量子产量(YNPQ) 4.567* 25.330** 8.287** 电子传递速率(ETR) 15.751** 117.903** 21.406** 说明:*表示差异显著(P<0.05);**表示差异极显著(P<0.01) Table 1. Two-way ANOVA of light intensity and seasonal change on the photosynthetic traits of M. sinostellata seedlings leaves
-
植物叶片的光合生理生态参数可反映植物生理代谢和物质积累的持续能力,也可分析环境因子对植物代谢和生长的影响[15]。景宁木兰幼苗春季、夏季100%全光照以及夏季40%全光照下的Pn日变化均为“双峰”曲线,可见景宁木兰幼苗在光照过强时会出现“光合午休”现象,产生光抑制,光合速率下降。Gs和Ci在同一时间范围内变化趋势一致,说明Pn下降是气孔因素导致的[16]。景宁木兰幼苗春季、夏季100%全光照以及夏季40%全光照下的Gs与Ci在中午的变化趋势相同,可知景宁木兰幼苗出现光合午休的原因是由气孔因素导致,强光照和高温导致气孔关闭,减弱了与外界气体交换的能力,光合能力下降。日均Pn在夏季100%全光照和40%全光照均显著高于10%全光照,而秋季40%全光照显著高于其他2个光照处理,可能是由于夏季光照过强导致100%全光照下植物叶片受到光抑制,并且在经过夏季长时间的高温、高光环境胁迫后,叶片被灼伤,光合机构受到不可逆的损伤,从而限制了其在秋季的光合能力,表明景宁木兰幼苗对强光的适应能力较差,很难与其他伴生植物竞争,这可能是造成景宁木兰在野外濒危的原因之一。适当遮光条件有利于景宁木兰的生长,但在10%全光照环境下,过低的光照会导致Pn处于较低水平,不利于光合产物的合成。
光响应曲线能够直观反映植物光合过程的变化[17]。Pnmax能够体现植物潜在的光合能力,LSP和LCP分别表示植物对强光和弱光的利用能力[18]。景宁木兰幼苗Pnmax和LSP在夏季10%全光照处理下显著低于100%全光照和40%全光照处理,而经过夏季高温和高光的环境胁迫,秋季景宁木兰幼苗Pnmax和LSP在100%全光照下最低,叶片光合机构受到损伤导致Pnmax和LSP下降,而适当遮光下景宁木兰幼苗的Pnmax和LSP能够维持在一个较高水平,这与日均Pn变化情况相似,这与俞芹等[17]在夏季对景宁木兰的研究相似。与100%全光照相比,10%全光照下景宁木兰在夏季和秋季的AQY均显著上升,LCP和Rd显著下降,可知在10%全光照下,景宁木兰幼苗能够提高叶片对光的利用能力,减少由于呼吸作用产生的能量消耗,是对弱光环境的适应性变化。
CO2是植物进行光合作用的底物,其浓度高低会影响植物光合速率。在夏季,3种光照处理下景宁木兰幼苗的Vcmax、Jmax和TPU从大到小均为40%全光照、10%全光照、100%全光照,在秋季,100%全光照下Jmax和TPU也显著低于40%全光照,可能是强光照下景宁木兰幼苗核酮糖-1,5-二磷酸羧化酶活性和数量下降,Jmax降低,从而限制其光合磷酸化和还原型辅酶Ⅱ的再生,并且磷酸丙糖的积累也对光合作用产生负反馈,且经过夏季高温高光环境影响,植物会受到光损伤,而遮光后的景宁木兰幼苗体内核酮糖-1,5-二磷酸羧化酶活性和数量上升,光合能力提高,在40%全光照下3个季节均维持较高的光合速率水平,这与可可Theobroma cacao在强光下的研究结果相似[19]。
Fv/Fm能衡量植物光抑制的程度[20]。在夏季和秋季,100%全光照下景宁木兰幼苗的Fv/Fm分别为0.68和0.72,说明在夏季、秋季100%全光照下的景宁木兰幼苗受到光胁迫,而遮光处理并没有受到光胁迫。YⅡ能反映吸收的光子供给PS Ⅱ反应中心的效率[21]。夏季、秋季遮光处理YⅡ均显著高于全光照,可能是相对于100%全光照环境,遮光下景宁木兰幼苗能将更大比例的光能分配给光化学反应。100%全光照下YⅡ在秋季显著低于春季、夏季,说明全光照下景宁木兰幼苗受到夏季高温高光环境的胁迫。qP反映PS Ⅱ天线色素吸收的光能中用于光化学电子传递的份额,其值越大则PS Ⅱ电子传递活性越高[22]。10%全光照下qP在3个季节均小于其他2个光照处理,说明重度遮光会降低植物的光化学效率,不利于光能转化为化学能。qNP反映植物叶片PS Ⅱ反应中心非辐射能量耗散效率的大小,表示以热能消耗的光能部分,能反映植物的光保护能力[22]。夏季100%全光照下景宁木兰幼苗qNP要显著高于遮光处理,这可能是夏季全光照环境下光照强度充足,叶片吸收的光能过多,需要通过热耗散来消耗光能,保护部分光合机构。在夏季,40%全光照下的YNO和YNPQ显著高于春季、秋季,表明中度遮光下YNO增加,光诱导的YNPQ下降。ETR表示在稳态光合作用过程中通过PS Ⅱ的相对电子数量[23]。春季40%全光照下ETR显著低于100%全光照,而夏季和秋季40%全光照下ETR均显著高于100%全光照,说明经过一段时间的遮光处理后,40%全光照下的ETR明显增加,有利于光合能力的提高。
-
春季、夏季100%全光照以及夏季40%全光照下景宁木兰幼苗具有“光合午休”现象,是气孔因素所致。100%全光照下景宁木兰幼苗在夏季、秋季都受到光胁迫,使叶片受到损伤,导致秋季光合速率明显下降,且100%全光照下景宁木兰幼苗的碳同化能力均被限制。而40%全光照下景宁木兰幼苗具有较高的AQY、Vcmax、Jmax、TPU、YⅡ、ETR,同时具有较低的qP、qNP,对电子的传递速率较高,对CO2利用能力较强,光合速率高。10%全光照下能通过降低LCP、Rd,提高Vcmax、TPU、YⅡ来适应过度遮光环境,但其所处环境光照强度过低,不利于光合产物合成。总之,景宁木兰幼苗不宜在强光下生长,在栽培过程中需要进行遮光处理,但遮光强度不宜过高,建议光照强度保持在自然光的40%以上。
Seasonal changes of photosynthetic characteristics of seedlings of Magnolia sinostellata under different light intensities
doi: 10.11833/j.issn.2095-0756.20210814
- Received Date: 2021-12-24
- Accepted Date: 2022-04-21
- Rev Recd Date: 2022-03-18
- Available Online: 2022-09-22
- Publish Date: 2022-10-20
-
Key words:
- Magnolia sinostellata seedlings /
- light intensity /
- seasonal changes /
- photosynthesis /
- chlorophyll fluorescence
Abstract:
| Citation: | CHEN Chao, JIN Zexin, YUAN Meng, et al. Seasonal changes of photosynthetic characteristics of seedlings of Magnolia sinostellata under different light intensities[J]. Journal of Zhejiang A&F University, 2022, 39(5): 950-959. DOI: 10.11833/j.issn.2095-0756.20210814

|

 DownLoad:
DownLoad: