







-
群落结构特征与种群空间分布格局是森林空间结构的重要研究内容,对开展森林可持续经营及揭示群落演变规律具有重要意义,一直受到林学家和生态学家的广泛关注[1−2]。作为温带地区标志性的地带植被类型,落叶阔叶林广泛分布于中纬度地区。在亚热带高海拔地区,落叶阔叶林是垂直植被体系不可或缺的一部分,其季节性的变换尤为显著[3−4]。针对不同研究范围展开的分析表明:影响落叶阔叶林群落的因素存在差异。如在大范围内,其地理位置与气候条件存在关联;而在小尺度的研究中,则和落叶阔叶林群落的环境异质性以及群落演替有关。落叶树种对于整个群落结构、物种组成、更新状况、养分以及物质循环等方面均具有关键意义。人们逐渐意识到生态转换带的复杂性以及资源丰富性,尤其是植被、动物和微生物组成的复杂森林生态系统对物种变化起到重要作用。近年来,国内学者对暖温带-北亚热带过渡区的群落结构、空间分布格局等开展了诸多研究[5−8]。安徽天马国家级自然保护区位于亚热带向暖温带的典型转折点上,具有独特、罕见且天然的自然资源,也是各类动植物的重要栖息地[9]。已有研究探究了天马国家级自然保护区马鬃岭片区的天然落叶阔叶林结构特征、物种多样性以及种间关系[10−11],而对于种群分布格局和种间空间关联性的研究还较少。本研究使用点格局分析法深入探讨该片区落叶阔叶林的演替动态和物种共存机制,为天马国家级自然保护区森林质量提升及群落结构调整策略制定提供参考。
-
天马国家级自然保护区马鬃岭片区位于安徽省金寨县西南部(31°10′~31°20′N,115°30′~115°50′E),地势呈西北至东南走向,南高北低,平均海拔为900 m。植被类型为落叶常绿阔叶林和落叶阔叶林,乔木树种主要有杉木Cunninghamia lanceolata、黄山松Pinus taiwanensis、马尾松P. massoniana及短柄枹栎Quercus serrata、麻栎Q. acutissima、栓皮栎Q. variabilis、茅栗Castanea seguinii、化香Platycarya strobilacea、大别山山核桃Carya dabieshanensis等,灌木有檵木Loropetalum chinense、杜鹃Rhododendron simsii、荚蒾Viburnum dilatatum、山胡椒Lindera glauca等。列入国家级重点保护的珍贵植物有银缕梅Parrotia subaequalis、大别山五针松P. dabeshanensis、香果木Emmenopterys henryi、连香树Cercidiphyllum japonicum等。该地区属于北亚热带湿润季风气候,全年平均气温为13.3 ℃,夏季平均气温为20.0 ℃,年平均降水量为1 480.0 mm,无霜期为200.0~220.0 d。
-
2018年9月在主要区域设置72 m×72 m的样地,样地中心坐标为31°15′30.2″N,115°41′45.7″E,海拔为1 069~1 113 m,东南坡,坡度为18°~20°,优势树种为短柄枹栎、茅栗、化香,密度为1 537株·hm−2,胸高断面积为37.77 m2·hm−2。使用全站仪定位四至边界,再利用相邻格子法,把该样地分割成9个24 m×24 m的样方,收集每个样方中胸径(DBH)≥2.5 cm林木的信息,包括胸径、树高、空间坐标、海拔、坡度、坡向等。根据样地起测胸径和最大胸径,将林木胸径划分为13个径级组(下限排除法),起始树木胸径为2.5 cm,组距为5.0 cm,从Ⅱ (>7.5~12.5 cm)开始记录,统计各胸径组单位面积株数。
-
树种重要值(VI)表示样地树种的优势程度[12],用来确定空间格局分析的树种。重要值=(相对密度+相对显著度+相对频度)/3。其中:相对密度(%)为样地内某个树种株数与所有树种株数总和之比;相对显著度(%)为某树种胸高断面积与所有树种的胸高断面积总和之比;相对频度(%)为某个树种出现的样方数与所有树种出现的样方数总和之比。
-
为有效研究多变量模式的空间架构,抑制Ripley K函数的累积影响,本研究采用圆环替换原有的圆形,增强g(r)函数的精度[13−14]。通过单变量g11(r)函数和完全空间随机(complete spatial randomness,CSR)零模式的检验,深入探究样地主要树种的空间变化规律,了解其生态系统特征。采用异质泊松(heterogeneous Poisson,HP)零模型来估计强度函数λ值,由于遵循泊松分布,实际的局部领域密度会大大高于理论预测,因此需要考虑一阶效果,而且还需要使用Epanechnikov核来估算。通过使用双变量g12(r),研究样本中物种1和物种2之间的关系,并使用异质泊松零模型来确定它们之间的真正交互。这一方法能够消除栖息地的干扰。在异质泊松零模型的运算中,物种1的位置保持固定,而物种2的位置则进行随机分配,故而,两两树种之间的相互作用并非对称关系[15−16]。
设定1 m×1 m 的栅格,圆环宽度3 m,检验尺度为0~3 m,使用蒙特卡罗模拟(Monte Carlo Simulations)中汇总统计数据199 次的第5位低值和第5位高值,生成了 95%的模拟包络,结合拟合优度(GOF)测试评估距离间隔内的显著性水平[17],如果数据拟合次数大于199,且P<0.05,则零假设在5%的水平拒绝。g11(r)值置于置信区间的上方、内部、下方时,则分别表示物种在该尺度上为聚集、随机、均匀分布;当g12(r)值置于置信区间的上方、内部、下方时,则分别表示物种在该尺度上为正相关性、无相关性、负相关性。
式(1)~(2)中:r代表空间尺度的距离;K(r)表示同一树种在r尺度下的点对数目;K12(r)表示2个不同树种在r尺度上的点对数目;dK(r)表示同一树种在设定圆环宽度下的点对数目;dK12(r)表示2个不同树种在设定圆环宽度下的点对数目;dr代表圆环的宽度(dr=0.2/0.5λ);g11(r)表示圆环中实际观测到的树木数量与预期树木数量的比例;g12(r)表示物种1周围的圆环中实际观测到的物种2的数量与预期数量的比例。
-
样地树种调查统计结果见表1。样地林木DBH≥2.5 cm的树种组成共有17科20属27种,其中壳斗科Fagaceae和蔷薇科Rosaceae树木种类最丰富。壳斗科有栎属Quercus的短柄枹栎、槲栎Q. aliena、锐齿槲栎Q. aliena var. acutiserrata;栗属Castanea的茅栗;蔷薇科含李属Cerasus的尾叶樱(C. dielsiana、稠李Prunus padus、山樱花P. serrulata;石楠属Photinia的中华石楠P. beauverdiana。樟科Lauraceae、山茱萸科Cornaceae、清风藤科Sabiaceae、豆科Fabaceae各有2种,其余科只有1个树种。按照偶见种1~10株·hm−2的标准[18],样地内偶见种有红果山胡椒Lindera erythrocarpa、暖木Meliosma veitchiorum、秀丽槭Acer elegantulum、山樱花、山合欢Albizia kalkora等10种。
序号 树种 密度/
(株·hm−2)胸高断面积/
(m2·hm−2)重要值/% 科 属 1 短柄枹栎Quercus serrata 183 13.53 19.77 壳斗科Fagaceae 栎属Quercus 2 茅栗Castanea seguinii 201 11.91 19.12 壳斗科Fagaceae 栗属Castanea 3 化香Platycaryas trobilacea 123 5.25 10.24 胡桃科Juglandaceae 化香属Platycaryas 4 灯台树Cornus controversa 152 0.64 6.42 山茱萸科Cornaceae 灯台树属Cornus 5 君迁子Diospyros lotus 122 0.59 6.12 柿科Ebenaceae 柿属Diospyros 6 山胡椒Lindera glauca 114 0.25 4.86 樟科Lauraceae 山胡椒属Lindera 7 野鸦椿Euscaphis japonica 106 0.20 4.77 省沽油科Staphyleaceae 野鸦椿属Euscaphis 8 野茉莉Styrax japonicus 102 0.29 4.76 安息香科Styracaceae 安息香属Styrax 9 鹅耳枥Carpinus turczaninowii 106 0.58 4.26 桦木科Betulaceae 鹅耳枥属Carpinus 10 中华石楠Photinia beauverdiana 66 0.26 3.17 蔷薇科Rosaceae 石楠属Photinia 11 槲栎Quercus aliena 29 1.62 2.91 壳斗科Fagaceae 栎属Quercus 12 锐齿槲栎Quercus aliena var. acutiserrata 27 1.36 2.50 壳斗科Fagaceae 栎属Quercus 13 四照花Dendrobenthamia japonica 41 0.22 2.13 山茱萸科Cornaceae 山茱萸属Dendrobenthamia 14 尾叶樱Cerasus dielsiana 33 0.28 1.88 蔷薇科Rosaceae 李属Prunus 15 黄檀Dalbergia hupeana 33 0.19 1.67 豆科Fabaceae 黄檀属Dalbergia 16 刺楸Kalopanax septemlobus 25 0.14 1.19 五加科Araliaceae 刺楸属Kalopanax 17 金缕梅Hamamelis mollis 14 0.02 0.71 金缕梅科Hamamelidaceae 金缕梅属Hamamelis 18 山合欢Albizia kalkora 8 0.14 0.56 豆科Fabaceae 合欢属Albizia 19 山樱花Cerasus serrulata 8 0.08 0.50 蔷薇科Rosaceae 李属Prunus 20 秀丽槭Acer elegantulum 8 0.03 0.46 无患子科Sapindaceae 槭属Acer 21 红果山胡椒Lindera erythrocarpa 8 0.01 0.45 樟科Lauraceae 山胡椒属Lindera 22 华东椴Tilia japonica 6 0.11 0.42 椴树科Tiliaceae 椴树属Tilia 23 暖木Meliosma veitchiorum 8 0.02 0.38 清风藤科Sabiaceae 泡花树属Meliosma 24 红枝柴Meliosma oldhamii 4 0.03 0.24 清风藤科Sabiaceae 泡花树属Meliosma 25 肉花卫矛Euonymus carnosus 4 0 0.22 卫矛科Celastraceae 卫矛属Euonymus 26 青榨槭Acer davidii 4 0.03 0.18 槭树科Aceraceae 槭属Acer 27 稠李Prunus padus 2 0 0.11 蔷薇科Rosaceae 李属Prunus 合计 1 537 37.78 100 Table 1. 1 Composition of tree species in sample plot
样地内短柄枹栎、茅栗、化香的重要值均大于10.00%,分别是19.77%、19.12%和10.24%,3个树种重要值占总重要值的49.13%。3个树种胸高断面积分别为13.53、11.91、5.25 m2·hm−2,占样地总断面积的比例分别为35.81%、31.52%和13.90%,总占比为81.23%,因此,该样地群落类型为栎类与化香落叶阔叶林,优势树种为短柄枹栎、茅栗,亚优势树种为化香。
-
根据样地调查结果统计各胸径组单位面积株数,并绘制样地林木胸径结构图(图1)。由图1可知:样地林木胸径结构呈现倒“J”型分布,其中2.5~7.5、7.5~12.5 cm直径组的林木株数最多,且随着胸径的不断增大,林木株数不断减少。这表明样地林分具备异龄林林分胸径结构的特点。图2 显示:优势树种短柄枹栎的胸径结构大致呈倒“J”型分布,然而茅栗、化香的胸径结构与林地胸径结构不同,呈正态分布;短柄枹栎小径级的林木株数较多,意味着其林内更新情况良好,具有增长的态势;茅栗、化香小径级的林木株数较少,林内更新不佳,增长的趋势并不明显。

Figure 1. DBH structure of forest stands in the sample plot
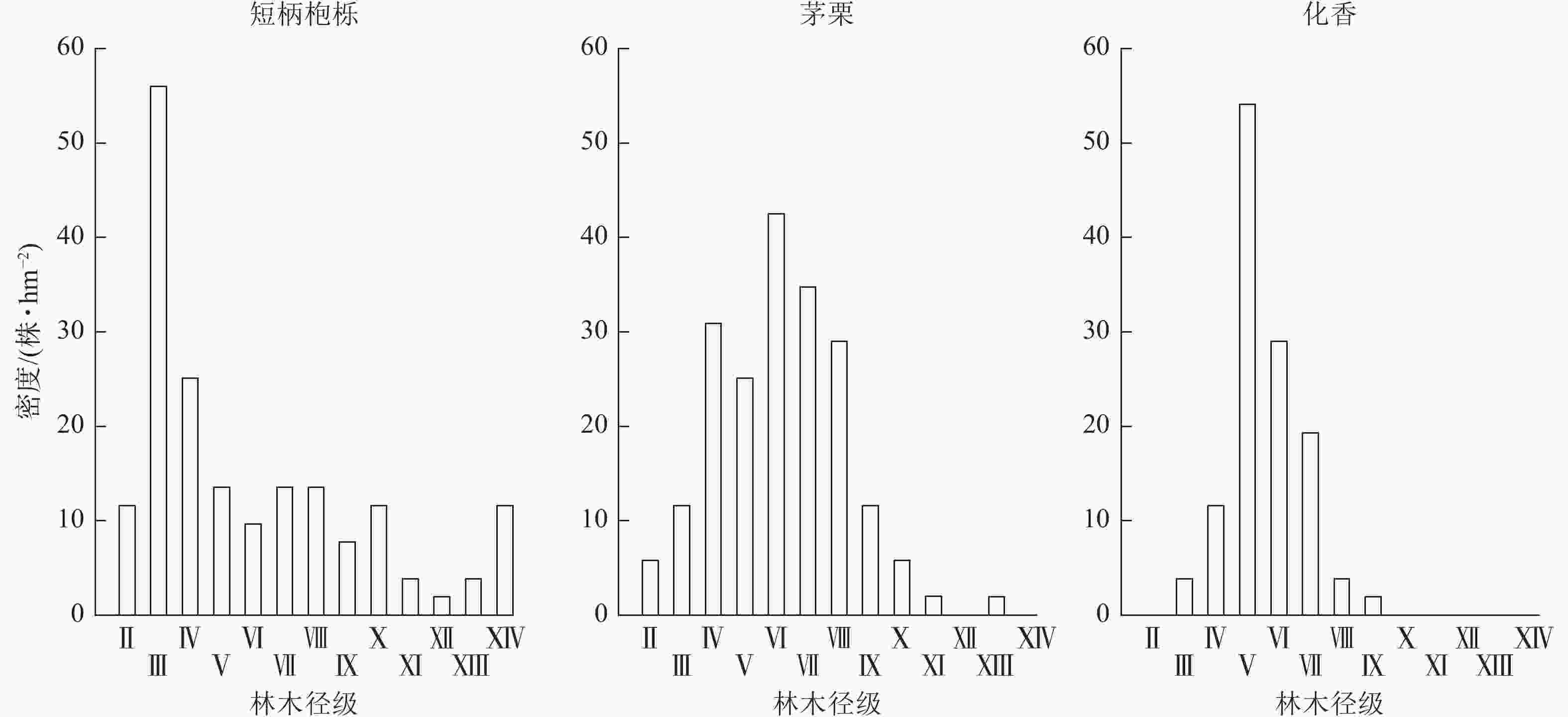
Figure 2. DBH structure of dominant tree species
-
样地所有树种林木2种空间分布函数模型结果如图3。运用单变量 O-ring 函数展开的分析结果表明:在完全随机零模型(CSR)的情况下,样地内所有林木在小尺度0~5 m范围呈现出聚集分布态势,而当尺度为5~30 m时,则呈随机分布。在异质泊松零模型(HP)情况下,所有林木同样在小尺度上表现出聚集分布的特征。随着空间尺度的不断增加,其聚集程度逐渐减弱,最终呈现出不规则随机分布的状态。
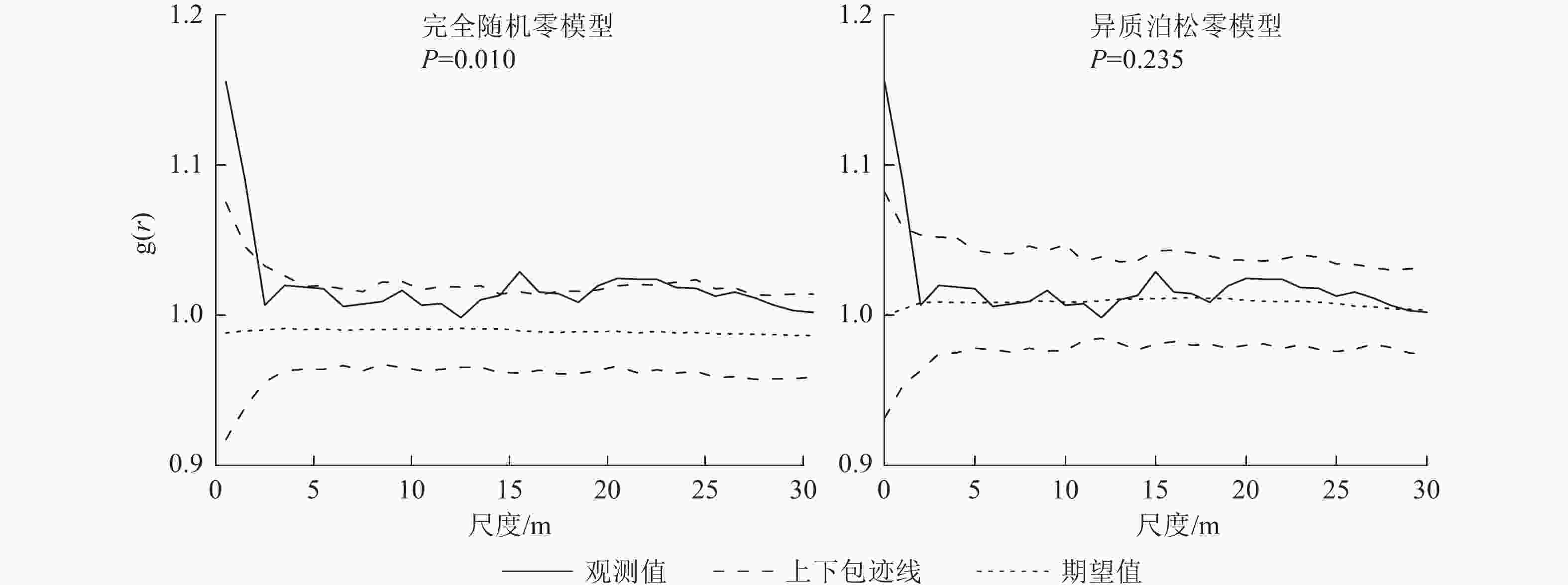
Figure 3. Spital distribution pattern of all tree species under CSR and HP
-
根据完全随机零假设模型、异质泊松零模型,短柄枹栎、茅栗和化香3种优势树种空间分布格局如图4。在完全随机零假设模型情况下,这3种优势树种在0~30 m尺度范围内,起初均表现为聚集分布,伴随尺度的扩大,逐渐转变为随机分布态势。其中,茅栗在1~15、18~30 m尺度上呈现不规则随机分布,在其他尺度上则为聚集分布;化香在2~7、7~21、23~30 m的尺度上呈现不规则随机分布,在其余尺度上为聚集分布;短柄枹栎在5~30 m的大尺度上呈不规则随机分布,在小尺度上主要为聚集分布。在异质泊松零假设模型下进行分析可见:这3种优势树种在0~30 m尺度上起初呈现聚集分布,随尺度的增大,其空间分布格局逐渐趋向于不规则随机分布。
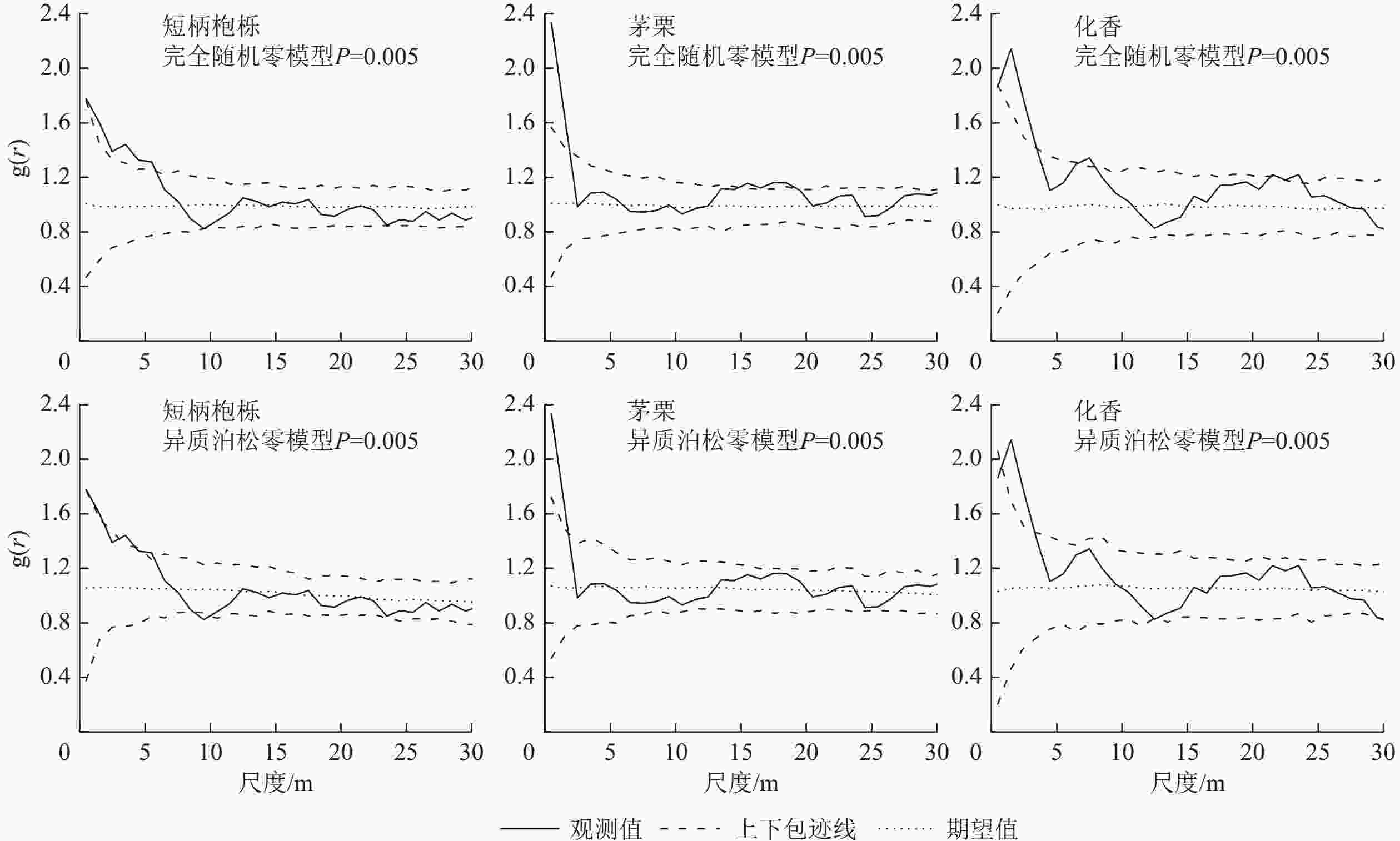
Figure 4. Spatial distribution map of dominant tree species under CSR and HP
-
由表2可见:在异质泊松零模型、GOF检测偏离零模型(P<0.05)检验中,样地内有13个树种对存在显著关联。短柄枹栎、茅栗和化香3种优势树种之间关联性不显著,但3种优势树种与伴生树种在一定尺度上存在关联性。如短柄枹栎与灯台树在0、2 m尺度上呈负相关,在18~20 m尺度上呈正相关,而短柄枹栎与君迁子在2、4 m尺度上呈负相关,26~28 m尺度上呈正相关。同一种优势树种与不同伴生树种在不同尺度上相关性不同。茅栗与野茉莉在0~2、24 m尺度上呈正相关。化香与灯台树在0、0~2、14~16 m尺度上呈正相关,而化香与山胡椒在4、4~6、6 m尺度上呈正相关,在10、10~12 m尺度上呈负相关。伴生树种之间在一定尺度上也存在关联性,如灯台树与山胡椒在0、0~2、2 m尺度上呈负相关,在22、22~24 m尺度上呈正相关。君迁子与野鸦椿在4、4~6、6、12、12~16 m尺度上呈正相关,其他尺度无关联。山胡椒与君迁子在10、10~12 m尺度上呈正相关,野茉莉与君迁子在8~10、14、14~20 m尺度上也呈正相关。
树种1 树种2 P 不同尺度的关联性 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 m 短柄枹栎 灯台树 0.020 − r/− r r r r r r r r/+ +/r r r r r r 短柄枹栎 君迁子 0.010 r r/− r/− r r r r r r r r r r r/+ + +/r 茅栗 野茉莉 0.050 r +/r r r r r r r r r r r r/+ r r r 化香 灯台树 0.005 + + r r r r r r +/r r r r r r r r 化香 山胡椒 0.020 r r + + r − −/r r r r r r r r r r 灯台树 化香 0.045 + r r r r r r r r r r r r r r r 灯台树 山胡椒 0.030 − − r r r r r r r r r + +/r r r r 君迁子 野鸦椿 0.020 r r r/+ + r r + + +/r r r r r r r r 山胡椒 化香 0.050 r r r/+ + r r r r r r r r r r r r 山胡椒 灯台树 0.005 r − r r r + r r r r/+ r + +/r r r r 山胡椒 君迁子 0.030 r r r r r + +/r r r r r r r r r r 野鸦椿 君迁子 0.035 r r r/+ + r r + + +/r r r r r r r r 野茉莉 君迁子 0.050 r r r r r +/r r r/+ + + +/r r r r r r 说明:+. 正相关;−. 负相关;r. 无关联。如果关联性有1个符号(如“+”),则表示对应尺度与上一个尺度和该尺度本身都具有正相关;如果关联性有2个符号(如“−/r”),则表示对应尺度与上一个尺度具有负相关,与该尺度本身无关联。 Table 2. Interspecific associations of major tree species in the sample plot
-
通常认为:在小尺寸上形成的林木空间分布格局首先是受群体本身生态学特征的直接影响,而在较大尺寸上的空间分布格局则更多是由于栖息地差异性的相互作用[19]。本研究结果与阔叶落叶林主要树种径级结构呈倒“J”型的研究结果一致[20−22],但与吴茜等[23]对紫金山针阔混交林主要树种径级结构的研究结果略有差异。有研究表明:空间结构在小尺度上呈集群分布,聚集程度随着尺度的增大而降低[24−27]。本研究样地林木在小尺度上呈聚集分布,伴随空间尺度的增大逐步趋向随机分布。这是由于在样地选择时就考虑到生境的一致性,所有林木均在同一坡面,坡度为18°~20°,样地生境异质性小;另一方面,种子质量较大的树种,种子成熟后依靠外部风力及动物的迁移所能传递的时间受限,大多散落在母树附近,体现在更小空间尺度的集中散布。样地优势树种短柄枹栎和茅栗,果仁质量较大,而化香果实为小坚果,背腹呈压扁状,两端具狭翅,较轻,所以,短柄枹栎、茅栗与化香比较偏向于更小空间尺度上集中散布。在群落演替进程中,天然次生林基本物种空间结构由聚集分布过渡为随意散布,因而,随意散布常呈现在生长发育完善、比较稳定的群落内[28−30]。本研究样地内林木总体上呈现随意散布,也验证了研究区群落结构的稳定性。
种间关联性主要与生长环境、自身特性等相关,正关联表示种群间生长环境具有相似性,负关联则表示与生长环境具有差异性。本研究也发现:样地优势种的种间关系多呈无关联特点,说明群落中的优势种并没有表现出种内竞争关系,不同龄级的个体在较大的空间尺度上相互独立。随着观察尺度的变化,优势种内部可能呈现正关联或无关联,这反映了同一物种在利用环境资源相似性上呈现出种群内部积极的生态关系。这种关系增强了种间竞争力,使得这些种群能够在群落中占据优势地位,从而维持其生存和发展[31−32]。群落中不同空间层次的树种对生长环境因子的需求不同,树种间的生存策略存在差异,竞争关系主要发生在中、下层[33]。本研究优势树种与伴生树种之间存在一定的关联性。具体表现为:种间存在负关联的分别是短柄枹栎与灯台树、短柄枹栎与君迁子、化香与山胡椒、灯台树与山胡椒,位居群落上层的短柄枹栎、化香,可能对下层的灯台树、君迁子、山胡椒个体存在调控作用,因此,种间关系表现为明显的竞争排斥[34]。茅栗与野茉莉、化香与灯台树、君迁子与野鸦椿、山胡椒与化香、山胡椒与君迁子、野茉莉与君迁子各种间存在正关联或无关联,说明它们之间具有一定的依赖性。研究区落叶阔叶林主要树种总体上呈正相关或无关联,且树种数量均多于负相关的树种,表明这些树种的分布格局具有相互独立的特征,反映出群落整体处于相对稳定阶段,物种之间能够共存,竞争排斥现象较弱,从而促进了群落的稳定性,有利于群落的演替过程。这与姜冬冬等[35]的研究结论一致。
-
本研究样地森林群落类型为落叶阔叶混交林,优势树种为短柄枹栎、茅栗和化香,具有北亚热带落叶阔叶林树种组成特征。林木直径结构具有典型的异龄林林分直径结构特点。短柄枹栎林下更新能力较强,为发展种群,茅栗、化香林下更新较差,为衰退种群。在完全空间随机零模型和异质泊松零模型下林木空间分布格局:在0~5 m小尺度上均呈现聚集分布,在其他尺度上呈现随机分布,随着空间尺度的增加逐渐趋于随机分布,群落发育比较成熟,具有地带性稳定群落特征。短柄枹栎、茅栗和化香3种优势树种之间关联性不显著,但与灯台树、君迁子等伴生树种之间存在一定的关联性。
Spatial distribution pattern and correlation of main tree species in deciduous broad-leaved forest in Tianma National Nature Reserve
doi: 10.11833/j.issn.2095-0756.20240486
- Received Date: 2024-08-11
- Accepted Date: 2024-11-26
- Rev Recd Date: 2024-11-20
- Available Online: 2025-05-30
- Publish Date: 2025-05-30
-
Key words:
- deciduous broad-leaved forest /
- spatial distribution pattern /
- interspecific association /
- northern subtropical zone
Abstract:
| Citation: | TAO Tao, GUANG Taijun, HUANG Qingfeng, et al. Spatial distribution pattern and correlation of main tree species in deciduous broad-leaved forest in Tianma National Nature Reserve[J]. Journal of Zhejiang A&F University, 2025, 42(3): 544−553 doi: 10.11833/j.issn.2095-0756.20240486

|



 DownLoad:
DownLoad: