






-
种群指在一定时间内占据特定空间的同种生物的全部个体,是物种生存、适应及进化的基本单元[1],其结构特征因物种不同而有差异,且同一物种在不同发育阶段、不同生境条件下也表现不同。研究种群数量、分布、年龄结构等特征,有助于揭示种群生存现状、预测种群未来数量与结构变化,对理解种群生态过程及与环境的关系意义重大。种群空间格局是种群结构的关键内容,描述个体在生存空间的位置分布与布局特点,由植物生物学特性、种内种间相互作用及外部环境因素共同作用形成[2],一般分为随机分布、聚集分布、均匀分布3种类型。种内关联性体现种群内部个体空间分布的相互关系,反映种群内部生态联系。对种群空间分布格局与种内关联性的研究,是理解生物多样性、群落动态和生态系统功能的关键[3]。点格局分析可揭示种群在不同空间尺度的分布模式,准确重现种群个体分布动态过程及特征参数[4],已成为种群空间分布格局与生物学基础研究的重要方法。环境因子特别是地形因子,在塑造植物的空间格局方面起着决定性作用。地形因子通过改变土壤湿度、土壤养分和光照等综合环境条件,影响物种分布[5]。土壤是植物生存的物质基础,土壤养分的空间异质性对植物群落产生重要影响,不同植物对于养分需求的异质性决定了其适生地区。通过对植物群落空间格局与土壤、地形因子关系的研究,可以揭示植物群落的分布规律及其环境主导因子,深入诠释植物群落与环境的关系。
茂兰喀斯特常绿落叶阔叶混交林是地球上同纬度地区保存面积最大、原生性较强、结构相对稳定的喀斯特森林生态系统。该群落物种组成复杂、类型多样,但植被生存环境相对恶劣。粗糠柴Mallotus philippensis属大戟科Euphorbiaceae野桐属Mallotus,在研究样地内的相对多度大于5%,重要值位于样地内主要优势树种前7位[6],是喀斯特常绿落叶阔叶混交林的重要组成树种,耐荫和耐干旱瘠薄,能够在多种土壤和光照条件下生长,在生态恢复中可作为先锋树种和退化林修复优选树种。研究粗糠柴空间格局形成的生态过程及其影响机制,了解其种群的动态规律,可为喀斯特原生林保护及退化森林恢复提供重要的基础数据和参考依据。本研究以茂兰国家级自然保护区喀斯特常绿落叶阔叶混交林1.28 hm2样地内粗糠柴种群为研究对象,采用成对相关函数g(r)分析其空间分布点格局以及不同径级空间关联性,结合Pearson相关分析法分析该种群分布特征与环境因子(地形和土壤)的关系,探讨该种群分布格局形成的潜在生态过程。
-
茂兰国家级自然保护区坐落于贵州省黔南布依族苗族自治州荔波县东南部,其地理位置为25°09′20″~25°20′50″N,107°52′10″~108°45′40″E。该保护区海拔为430.0~1 078.6 m,属中亚热带季风湿润气候类型,年平均气温为15.3 ℃,其中1月平均气温为5.2 ℃,7月平均气温为23.5 ℃,年降水量为1 700.0 mm,集中分布在4—10月[7]。土壤主要为在碳酸盐类母岩上发育而成的喀斯特地带性土壤——棕黑色石灰土,呈碱性,土壤中的含钙量较高[8]。该地区的植被为常绿落叶阔叶混交林,属隐域性喀斯特森林植被顶极群落类型,从洼地、谷地到山顶均有分布,植被成层现象比较明显,原生性强[9]。研究区已发现维管束植物1 203种,种子植物1 172种,占贵州植物总种数的四分之一[10]。
-
2023年在茂兰国家级自然保护区喀斯特常绿落叶阔叶混交林内设立1块面积为1.28 hm2的固定样地。为便于调查,将该样地划分为128个10 m×10 m的小样方,并进行编号。对样地内胸径大于1 cm的粗糠柴进行每木调查,记录牌号、坐标、胸径、树高、东西冠幅、南北冠幅和生长状况,并挂牌编号和喷漆标记。样地的边界采用罗盘仪和皮尺测定,树高使用布鲁莱斯测高仪测量,胸径使用胸径尺测量,每株树的坐标位置使用测距仪确定。
-
记录每个10 m×10 m小样方的海拔、坡向、坡度、土层厚度等。同时,在每个小样方中随机选择3~5个点,取0~10 cm表层土壤均匀混合在一起,每个样方取500~1 000 g土壤装至一次性塑料盒中带回实验室,烘干打磨再分装,用于测定土壤化学性质。参照文献[11],土壤碱解氮质量分数通过靛酚蓝比色法测定;土壤速效磷质量分数采用氟化铵-盐酸浸提法测定;土壤速效钾质量分数通过乙酸铵浸提-火焰光度计法测定;土壤全氮质量分数使用开氏消煮法测定;土壤全磷质量分数通过HClO4-H2SO4消煮-钼锑抗比色法测定;土壤全钾质量分数采用氢氧化钠熔融法测定;土壤有机质质量分数使用稀释热-重铬酸钾容量法测定;土壤pH和土壤含水率用pH计和水分测定仪测定。
-
根据样地内粗糠柴胸径的调查数据,按照LY/T
2908 —2017《主要树种龄级与龄组划分》[12],再结合样地内粗糠柴的生物学特性和生长情况对粗糠柴种群径级结构进行划分以反映其不同的生长发育阶段。粗糠柴为速生树种且本次调查中该种群的整体胸径都较小,所以将粗糠柴的胸径(DBH)分为4个等级:径级Ⅰ为1.0 cm≤DBH<2.5 cm;径级Ⅱ为2.5 cm≤DBH<4.0 cm;径级Ⅲ为4.0 cm≤DBH<5.5 cm;径级Ⅳ为DBH≥5.5 cm。 -
运用单变量成对相关函数g(r)[13]对4种不同径级的粗糠柴进行空间点格局分析。g(r)由Ripley’ s K函数[14]转化而来,是用指定宽度的圆环区域取代了Ripley’s K的k函数中半径为r的圆环区域,是一个概率密度函数。本研究采用Monte-Carlo模拟方法生成函数的上下包迹线,进行199次拟合检验,以确定95%的置信水平。如果观测值超过上包迹线则呈聚集分布,低于下包迹线则呈均匀分布,位于上下包迹线之间则呈随机分布[15]。
采用双变量成对相关函数gij(r)[13]对粗糠柴种群4个径级两两之间进行空间关联性分析,i和j分别代表不同径级林木个体。在双变量分析中,如果观测值超过上包迹线,意味着2个变量之间存在显著的正相关关系;如果观测值位于上下包迹线之间,表示2个变量之间没有显著关联;如果观测值低于下包迹线,则显示2个变量之间存在显著的负相关关系[16]。
-
采用Pearson相关系数分析不同径级的粗糠柴林木数量与13个环境因子之间的相关性。数据分析和制图在Excel 2018、R 4.3.0和SPSS 27.0.1中进行。
-
经调查,茂兰国家级自然保护区喀斯特森林1.28 hm2样地内共有粗糠柴635株,个体密度为496株·hm−2(表1)。其中,粗糠柴树高集中在[2.0,5.0) m,共331株,占粗糠柴总株数的52.13%;树高<2.0 m的共123株,占比19.37%;≥5.0 m以上的共181株,占粗糠柴总量的28.50%。粗糠柴以径级Ⅰ为主,共466株,占其总个体数的73.39%;径级Ⅱ共101株,占总个体数的15.91%;径级Ⅲ共31株,占比4.88%;径级Ⅳ共37株,占比5.82%。
不同树高的株数及比例 不同径级的株数及比例 树高/m 株数/株 比例/% 径级 株数/株 比例/% <2.0 123 19.37 Ⅰ 466 73.39 [2.0,5.0) 331 52.13 Ⅱ 101 15.91 ≥5.0 181 28.50 Ⅲ 31 4.88 Ⅳ 37 5.82 Table 1. Population characteristics of M. philippensis population
-
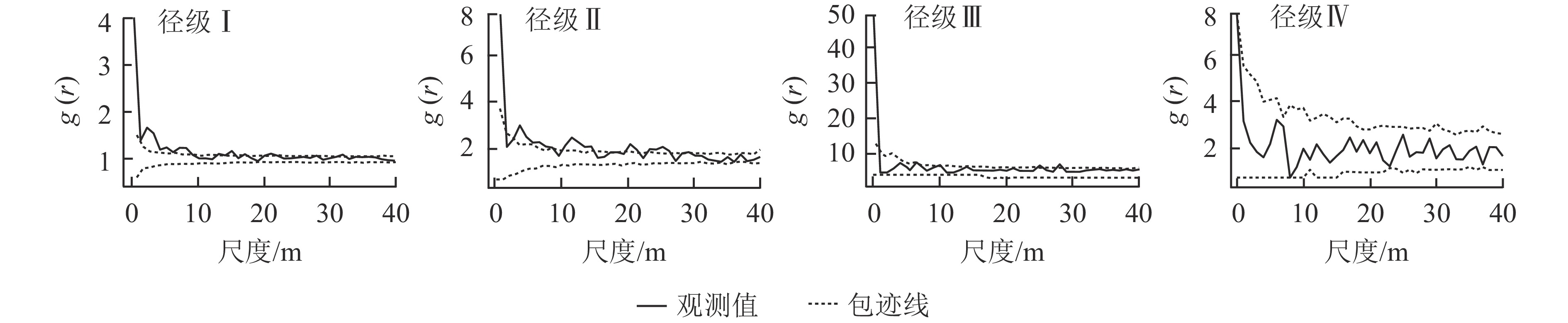
结合粗糠柴种群不同径级林木的空间分布图(图1)和点格局分析结果(图2)得出:在0~9 m尺度范围内,粗糠柴径级Ⅰ表现为聚集分布,超过9 m则表现为随机分布;径级Ⅱ在0~1、3~5、10~15 m尺度内呈现聚集分布,其余尺度内为随机分布;径级Ⅲ除在极小尺度为聚集分布外均为随机分布;径级Ⅳ在整个研究尺度范围内均表现为随机分布。整体来看,随着树龄的增加,粗糠柴种群的空间分布格局由聚集分布逐步转变为随机分布。
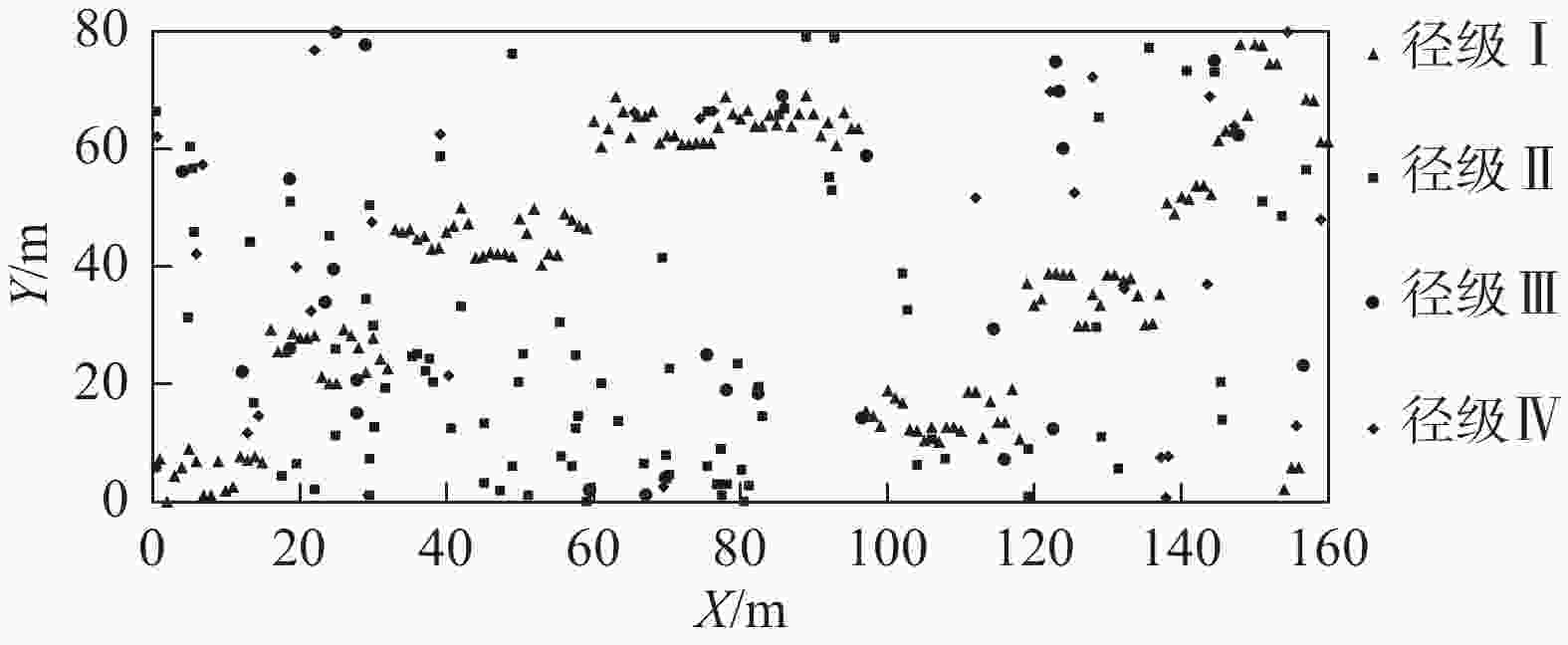
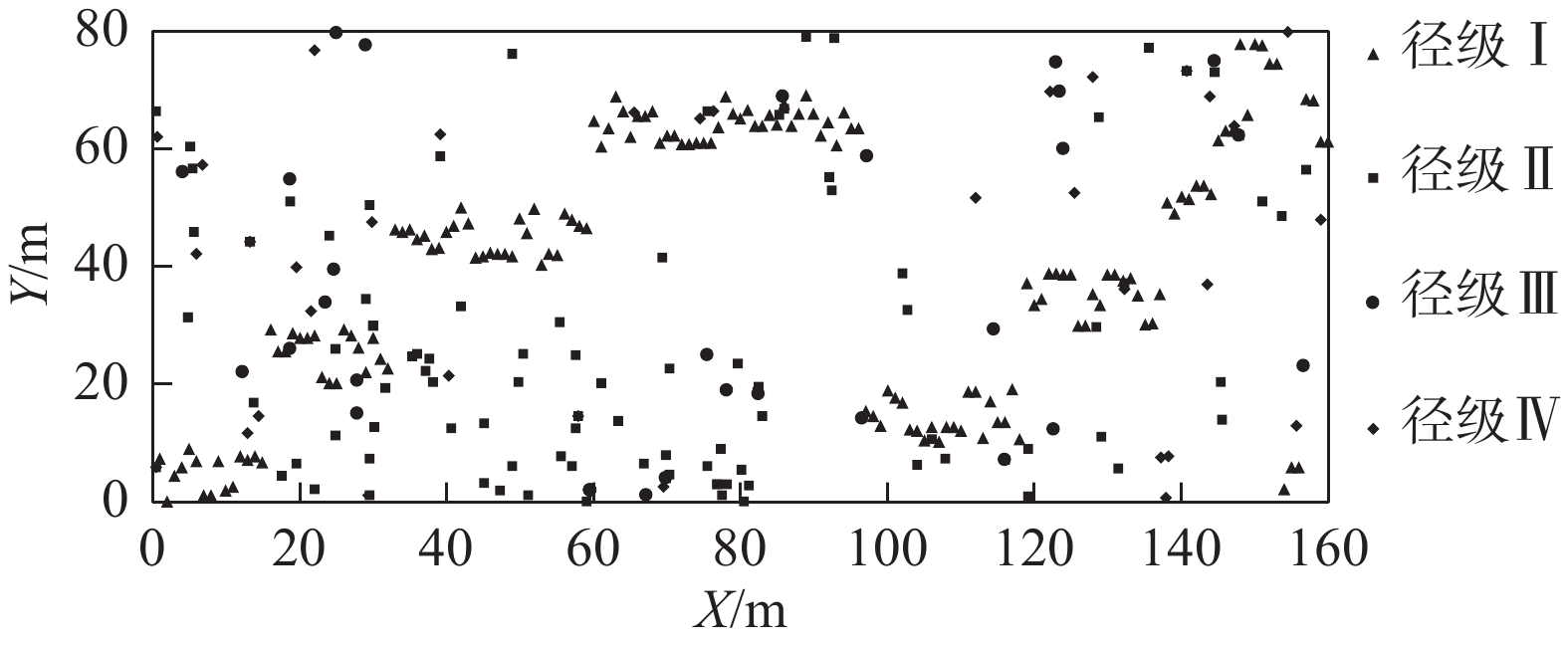
Figure 1. Spatial distribution of trees of different diameter classes of M. philippensis population
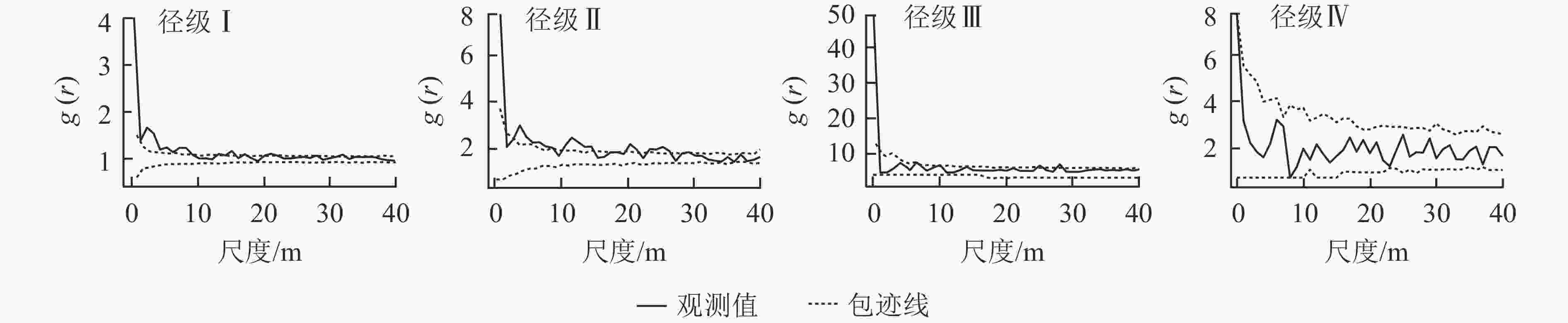
Figure 2. Spatial point patterns of trees of different diameter classes of M. philippensis population
-
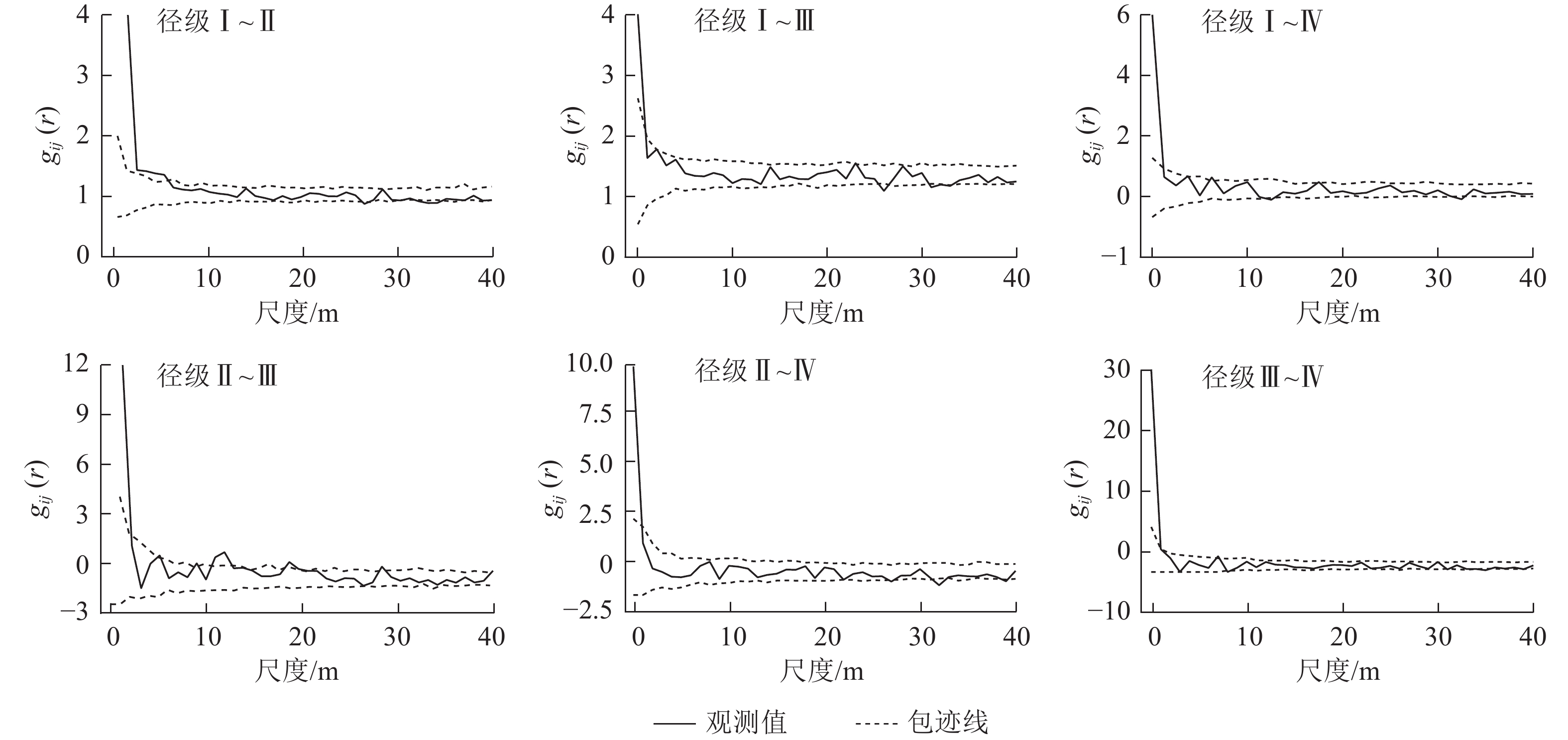
粗糠柴的不同径级之间的关联性存在差异(图3)。径级Ⅰ与径级Ⅱ、径级Ⅲ、径级Ⅳ在0~6、0~1、0~2 m的尺度上呈现显著的空间正相关关系;径级Ⅱ与径级Ⅲ在0~2、10~12 m的尺度上呈现空间正相关关联,与径级Ⅳ的空间正相关关系出现在0~1 m的极小尺度;径级Ⅲ与径级Ⅳ在0~2 m的小尺度上呈现显著的空间正相关关联,其余的尺度上为空间独立。整体来看,4个径级两两之间均表现为小尺度(多为0~2 m尺度)上呈现正关联,随着尺度增大,各径级之间呈现为无空间关联性,即空间独立。
Figure 3. Spatial correlation of trees of different diameter classes of M. philippensis population
-
如表2所示:粗糠柴整体数量与土壤含水率呈极显著正相关(P<0.01),与土壤速效钾、土壤全磷呈显著正相关(P<0.05),其中径级Ⅱ林木数量与海拔呈极显著负相关(P<0.01),与坡度呈显著负相关(P<0.05),与土壤速效氮、速效钾、全氮、全磷呈极显著正相关(P<0.01),与土壤速效磷、有机质呈显著正相关(P<0.05);径级Ⅲ林木数量与土壤含水率、速效钾呈极显著正相关(P<0.01),与土壤速效氮、全氮、全磷呈显著正相关(P<0.05)。
环境因子 径级Ⅰ 径级Ⅱ 径级Ⅲ 径级Ⅳ 整体 土壤速效氮 0.022 0.367** 0.191* 0.088 0.165 土壤速效磷 −0.091 0.192* 0.155 0.086 0.021 土壤速效钾 0.044 0.378** 0.271** 0.135 0.202* 土壤全氮 0.024 0.379** 0.211* 0.082 0.172 土壤全磷 0.074 0.433** 0.227* 0.069 0.227* 土壤全钾 −0.013 0.095 −0.006 −0.127 0.000 土壤有机质 −0.021 0.206* 0.111 0.035 0.066 土壤pH −0.114 −0.012 0.132 0.085 −0.062 土壤含水率 0.143 0.357** 0.274** 0.143 0.273** 土层厚度 0.006 −0.053 −0.058 0.024 −0.016 海拔 −0.023 −0.432** −0.129 0.059 −0.156 坡向 0.079 −0.072 −0.045 −0.070 0.023 坡度 0.039 −0.193* −0.074 −0.004 −0.038 说明:*. P<0.05; **. P<0.01。 Table 2. Correlation analysis between the number of individuals and environmental factors in different diameter classes of M. philippensis population
-
茂兰喀斯特常绿落叶阔叶混交林粗糠柴的胸径以径级Ⅰ (1.0 cm≤DBH<2.5 cm)为主,树高集中在[2.0,5.0) m。粗糠柴种群不同径级林木的株数不同,径级Ⅰ数量最多,径级Ⅲ数量最少,径级Ⅳ次之,呈“L”型分布,说明粗糠柴种群呈增长型,生长潜力大,更新能力强。茂兰国家级自然保护区植被受到的人为破坏少,除自然稀疏外,一般情况下生态环境没有剧烈的变化,粗糠柴种群数量呈稳定增长。
一般认为,不同径级粗糠柴种群个体存在着空间分布格局的差异,这可以更好地利用有限的资源[17]。在种群的幼龄阶段,个体的分布格局主要是聚集分布;随着种群年龄的增加,其分布从聚集分布趋向于随机分布[18]。本研究的结果符合这一规律。范忆等[19]研究了浙江省天目山国家级自然保护区1 hm2样地内紫楠Phoebe sheareri种群的空间分布格局,结果表明:低中龄个体为集群分布,高龄个体逐步转变为随机分布格局。陶涛等[20]研究发现:天马国家级自然保护区马鬃岭区优势树种在0~30 m尺度上起初呈现聚集分布,随尺度的增大,其空间分布格局逐渐趋向于不规则随机分布。这些研究表明:随着时间的推移,植株个体逐渐长大,对环境资源(水分、养分、光照和空间等)的需求也不断增加,导致种内和种间竞争加剧,进而引发自疏效应[21]。随着个体的进一步增大,植物对环境的适应能力增强,分布格局向随机分布转变[22]。赵广东等[23]对川西米亚罗亚高山暗针叶林9.6 hm2样地进行调查分析得出,岷江冷杉Abies fargesii var. faxoniana和糙皮桦Betula utilis不同径级个体的分布格局都有由聚集分布向随机分布发展的趋势。王均伟[24]发现:长白山阔叶红松Pinus koraiensis林主要树种的空间分布格局呈现聚集分布的特点,随着尺度的增大,逐渐转变为随机分布。植物种群的空间分布格局是其为了适应环境而采取的生存策略。这种分布格局并非一成不变,它会随着空间尺度的变化而改变,同时也受到植物自身生物学特性、发育阶段、种内关系和种间关系以及环境因素等多重因素的综合影响[25−26]。
竞争是构建群落和驱动树木空间分布模式的主要机制,在小范围聚集,且聚集程度随着距离增加而降低,这与种子扩散能力有关[27]。粗糠柴小乔木植株高度有限,种子落在母株附近的概率更大,新生个体更容易聚集在母株周围。随着空间尺度的增大,环境异质性开始影响种群,植物在适宜的地理环境中继续生长,最终植株分布趋于随机,如杨晓惠等[28]对小兴安岭典型阔叶红松林幼树种群研究,以及王斌等[29]对广西弄岗北热带喀斯特季节性雨林树种空间分布的研究。在这些因素的综合影响下,种群的空间格局逐渐由聚集分布向随机分布转变。喀斯特地貌地形,如小尺度岩石缝隙、裸露岩石以及溶沟是粗糠柴种群内中小个体小尺度呈现聚集分布大尺度呈随机分布的主要原因。
-
本研究中,4个径级两两之间仅在较小尺度上表现出正相关关系,而在其他尺度上则没有显著关联性。径级Ⅰ与其他龄级的个体竞争较小且幼龄个体的生存能力较弱,所以需要依赖中高龄级个体在生存过程中创造出的良好环境。随着尺度增加,中高龄个体带来的正向效应降低,外界环境的侵害及种内竞争导致的负向效应增加并与正效应相抵消,空间关联性就逐渐转变为不相关。与此相似,曹羚等[30]对新疆塔城地区额敏县新疆野苹果Malus domestica研究中发现年龄相近的个体空间关联性部分呈正关联或无显著关联,随着个体年龄间差距的增加,空间关联性大多数表现为无关联状态,以及王进等[31]报道的湖北木林子保护区的山矾Symplocos sumuntia在4个生长阶段中种内关联主要由正关联逐渐过渡到无关联。虽然有研究指出种内竞争是物种生存的主要压力,但本研究中粗糠柴在4个径级的种内关联性中并没有明显的排斥现象。这可能是因为同种个体具有相似的生物学特性,导致它们对环境资源的利用方式和生态位有所重叠,并形成种群内部的互惠关系,最终促进了种群的共存和发展[32]。
本研究发现:在完全随机分布模型下,4个径级两两之间均表现为小尺度(多为0~2 m尺度)上呈现正关联,随着尺度增大,各径级之间呈现为无空间关联,这很可能是由生态位空间隔离造成的。空间隔离减少了粗糠柴种群之间的竞争,利于该林木生存,同时提高了茂兰喀斯特森林的生物多样性[31]。
-
茂兰喀斯特森林粗糠柴整体的分布与土壤含水率、土壤速效钾、土壤全磷呈显著正相关。径级Ⅱ与海拔、坡度呈显著负相关,与土壤速效氮、速效钾、速效磷、全氮、全磷、有机质呈显著正相关;径级Ⅲ林木数量与土壤含水率以及速效钾、速效氮、全氮、全磷呈显著正相关。张忠华[33]发现:地形因子与土壤因子间的协同作用是影响喀斯特森林种群空间分布的重要因素,地形的起伏对土壤养分的空间分布产生影响;随着环境异质性的产生,种群的结构及空间分布格局也受到影响。李芹等[34]发现:坡度和海拔是喀斯特地区森林植物物种多样性及分布格局产生差异的主要影响因子,其次是坡向和坡位。粗糠柴是喜光植物,低温高湿不利于其光合作用及其他生理过程,阻碍植株生长。喀斯特地区坡度大,加上雨水的冲刷,加剧水土流失,导致土层浅薄,土壤养分低,因此在喀斯特这样土层浅薄且不均匀的特殊地区,坡度对于土层的影响就显得至关重要,进而影响着粗糠柴的生长和分布。适宜的水分供应是树木正常生长的必要条件,充足水分可保障其生理活动顺利进行,促进植株生长与繁殖。另外,喀斯特地区的土壤中富含钙、镁、钾、磷等元素,这些元素也成为影响植物种群分布的重要因素[33]。氮、磷、钾是植物生长必需的大量元素。土壤中的速效氮能直接参与植物蛋白质和核酸的合成,充足的速效氮可促进粗糠柴枝叶生长。土壤速效钾能增强植物的抗逆性,提高植株对干旱、低温等不良环境的适应能力,同时促进光合作用产物的运输和转化,有利于林木的生长和发育。土壤速效磷对植物根系发育至关重要,能刺激根系生长,提高根系对水分和养分的吸收效率。土壤速效磷高的区域往往有更多速生植物,这些植物能够快速占据资源。土壤全氮和全磷的空间分布影响到速效氮、速效磷的长期供应,因此也间接影响植物种群的空间分布格局。土壤有机质高能改善土壤物理性质、增大土壤孔隙度,利于植物根系呼吸与生长,满足林木能量转移、光合作用等需求,为生长提供充足营养。土壤中磷、钾影响植物对相应元素的吸收,并且植物对元素的需求和耐受能力不同,从而影响物种的分布和生长[35]。这与陈宝瑞等[36]在呼伦贝尔草原的研究结论相符。
-
本研究调查茂兰喀斯特森林粗糠柴的种群数量及生长地区的环境因子,将粗糠柴划分为Ⅰ~Ⅳ径级,并对4个径级的粗糠柴种群空间分布格局、种间关联性及其与环境因子的相关性进行了研究,主要研究结论如下:①粗糠柴种群不同径级的植株数量不同。样地中共有粗糠柴635株,径级Ⅰ数量最多,径级Ⅲ数量最少,粗糠柴种群呈增长模型,生长潜力大,更新能力强。②从径级Ⅰ到径级Ⅳ,粗糠柴种群空间分布格局由集群分布向随机分布转化,4个径级两两之间总体上表现为小尺度上呈现正关联,其余尺度上无空间关联性。③粗糠柴整体分布与土壤含水率、土壤速效钾、土壤全磷显著正相关,地形因子中海拔、坡度显著影响径级Ⅱ植株的空间分布。
Spatial distribution pattern and environmental interpretation of Mallotus philippensis population in karst forests
doi: 10.11833/j.issn.2095-0756.20250130
- Received Date: 2025-01-24
- Accepted Date: 2025-06-10
- Rev Recd Date: 2025-06-08
- Available Online: 2025-07-21
- Publish Date: 2026-02-20
-
Key words:
- Mallotus philippensis /
- point pattern /
- interspecific correlation /
- environmental interpretation /
- karst forest
Abstract:
| Citation: | WANG Rong, GUO Falan, QI Yujiao. Spatial distribution pattern and environmental interpretation of Mallotus philippensis population in karst forests[J]. Journal of Zhejiang A&F University, 2026, 43(1): 24−32 doi: 10.11833/j.issn.2095-0756.20250130

|
 DownLoad:
DownLoad: