






-
水稻Oryza sativa是全世界最主要的粮食作物之一,随着经济的快速发展以及人们生活品质的提高,水稻育种的核心目标已由单一追求高产逐渐转向优质稻米品质的协同提升。稻米品质是由外观品质、加工特性、营养组分、蒸煮食味与卫生安全等多维度性状构成的复杂表型系统。在消费升级和“大食物观”的战略驱动下,蒸煮食味已成为决定稻米市场竞争力和居民膳食结构优化的核心指标。蛋白质作为水稻胚乳中的第二大贮藏物质,其含量、组成与理化性质是决定稻米蒸煮食味品质的关键因素[1−2]。研究表明:在直链淀粉含量相近的情况下,稻米蛋白质含量与蒸煮食味品质存在显著的负相关关系[3−4]。
氨基酸是稻米蛋白质合成的重要底物,也是植物运输有机氮的主要形式。氨基酸转运蛋白在植物生长发育中起着关键作用。目前,水稻中已报道多个氨基酸转运蛋白基因。影响水稻产量性状的基因有OsAAP1、OsAAP3、OsAAP4、OsAAP5、OsAAP7、OsAAP13、OsAAP15、OsATL4、OsLHT1,其中OsAAP1通过在水稻生殖生长期调控小穗育性来影响水稻产量[5−6],OsLHT1通过调控穗粒数和千粒重来影响水稻产量[7−8],OsAAP15正向调控小穗的枝梗数与穗粒数的形成,它的功能在低氮及中氮环境下尤为显著[9],OsAAP3、OsAAP4、OsAAP5、OsAAP7、OsAAP13、OsATL4则通过调控水稻分蘖数来影响水稻产量[10−15]。值得注意的是,OsAAP13的启动子序列中含有脱落酸和水杨酸等激素响应元件和胁迫响应元件,说明OsAAP13很可能参与非生物胁迫调控[16];OsAAP3、OsLHT1被证实参与调控水稻叶片的衰老进程,这一过程间接关联光合产物的分配与产量稳定性[17−18]。OsAAP6、OsAAP8是水稻籽粒中淀粉含量的正调控因子和贮藏蛋白含量的负调控因子,对水稻的营养品质有一定影响作用,且OsAAP8在影响籽粒营养物质累积的同时还参与籽粒大小的调控[19−20]。在蒸煮与食味品质方面,敲除OsAAP10、OsAAP11可改善水稻的食味值[21−22]。OsATL6[23]负调控谷氨酰胺和${\mathrm{NH}}_4^+ $从根系到茎的转运,敲除后可促进茎的生长。
综上,现有研究多聚焦于一些氨基酸转运蛋白基因对水稻产量相关性状的影响,而对水稻品质性状调控的研究较少。目前,虽已有研究解析了OsAAP13的时空表达特性及其对水稻产量性状的调控作用,但关于该基因对稻米理化性质及蒸煮食味品质的调控机制尚不清楚。本研究利用CRISPR/Cas9技术,在3个不同的粳稻栽培品种背景下,对在水稻胚乳中高表达的OsAAP13 (LOC_Os04g39489)进行了敲除,重点解析OsAAP13对稻米蛋白质、淀粉及蒸煮食味特性的调控作用,为水稻优质育种提供理论基础和重要材料。
-
供试水稻为3个粳稻品种:‘武运粳30’‘Wuyungeng 30’(WYG30)、‘南粳9108’‘Nangeng 9108’(NG9108)、‘扬粳158’‘Yanggeng 158’(YG158)。分别种植在江苏省扬州市和海南省陵水县,各基因型双行种植,每行种植10株,株行距统一为20 cm×20 cm。全生育期实施标准化田间管理措施。
-
OsAAP13 (LOC_Os04g39489)的编码序列获取自RGAP网站。于OsAAP13第1个外显子中设计了一个靶基因序列:CCGCCGTTCCTCTCATCTCGG,该序列位于转录起始密码子ATG下游37 bp处。利用CRISPR-GE网站(
http://skl.scau.edu.cn/offtarget/ )对该靶位点进行脱靶分析。载体的构建方法和过程参考了文献[21]。利用美国国家生物技术信息中心(NCBI)(http://www.ncbi.nlm.nih.gov )数据库设计合适的检测引物OsAAP13-jc-F: AATCGATCGCCTCCAGTAGA和OsAAP13-jc-R: GGAGATGAAATCACGAGTGG。采用十六烷基三甲基溴化铵(CTAB)法提取转基因植株基因组DNA后,通过PCR扩增及测序分析鉴定突变体类型。 -
通过农杆菌Agrobacterium tumefaciens介导法(武汉伯远生物科技有限公司)获得T0转基因植株。将获得的T0阳性单株种植成T1。针对T1群体,按单株采集叶片并进行DNA提取,采用特异性引物Cas9-F:ACCAGACACGAGACGACTAA;Cas9-R:ATCGGTGCGGGCCTCTTC (用于检测Cas9基因)与Hyg-F:GCTGTTATGCGGCCATTGTC;Hyg-R:GACGTCTGTCGAGAAGTTTC (用于检测潮霉素抗性基因)进行PCR验证。通过目标条带缺失判定为无外源基因整合的纯合突变体。
-
将各水稻品种不同类型的纯合阴性突变单株种植成T3株系,完熟后按系进行混合收种,研磨成粉。使用kjeltec 2300自动氮分析仪(FOSS)测定米粉总氮含量,并基于氮转换系数(6.25)计算蛋白质含量。通过分级提取法分离贮藏蛋白组分,具体操作流程如下:称取100 mg的米粉置于2.0 mL的离心管中。使用1 mL ddH2O提取清蛋白,使用1 mL 1mol·L−1 NaCl提取球蛋白,使用1 mL 乙二醇溶液提取醇溶蛋白,使用1 mL 0.05 mol·L−1 NaOH提取谷蛋白。各组分经30 ℃、200 r·min−1的摇床中振荡2 h后,上清液转移至10 mL的离心管中,在4 ℃条件下以10 000 r·min−1的速度离心15 min,重复3次提取上清液。定量前,上清液冷冻于−20 ℃的环境下。提取完后使用BCA蛋白质定量试剂盒(北京天根公司)进行测定。
-
直链淀粉采用碘蓝比色法测定,参考NY/T 2639—2014《稻米直链淀粉的测定 分光光度法》,并对实验体系做了部分修改,操作流程如下:以4个已知直链淀粉质量分数分别为1.5%、10.4%、16.2%、26.5% 的样品作为标准样品。称取20.0 mg待测样品与标准样品置于2.0 mL离心管中,重复称取3次。离心管中依次加入100 μL无水乙醇和1.8 mL 1.0 mol·L−1的NaOH溶液,充分混匀后用保鲜膜固定在离心管板上,垂直放置于60 ℃摇床中以200 r·min−1的速度振荡1 h完成糊化。取100 μL混合液转移至含有9.0 mL蒸馏水的10 mL离心管,依次添加200 μL 1.0 mo·L−1乙酸钠溶液(pH 4.3)和200 μL质量浓度为 0.02%的碘试剂,充分摇匀并静置显色15 min,使用酶标仪(BioTek)于波长为620 nm下测定混合液的吸光度。利用标准样品计算标准曲线,根据公式计算待测样品的直链淀粉质量分数。
总淀粉采用蒽酮比色法测定[24],并在体系方面做了修改,测定方法如下:预先配制质量浓度为0.4%的蒽酮-硫酸显色剂,在待测样品与标准样品分别加入体积分数为52%的高氯酸30 mL,完全振荡溶解后将溶液滴定至50 mL,并稀释25倍。采用梯度稀释法,将标准淀粉溶液分别稀释至体积分数12.5%、25.0%、50.0%、75.0%和100.0%,各取1 mL稀释液与6 mL显色剂混匀置于10 mL的离心管中,沸水浴显色5 min后在室温中静置放凉。最后,使用蒸馏水对分光光度计进行调零,于波长640 nm处测定混合液吸光度。
-
称取精米30 g,用自来水清洗,加水,至精米∶水(质量比)=1.0∶1.3,用滤纸和橡皮筋密封容器后浸泡30 min,转入蒸锅蒸煮30 min,断热后焖制10 min。打开蒸锅翻动米饭防止粘壁,放置于冷装箱吹风10 min后关盖静置90 min。在此期间预热食味仪(STA1A) 30 min,软件设定为日本粳米模式进行校准。称取冷却好的米饭8 g装入金属环中置于仪器测定稻米食味值。该设备基于近红外光谱技术,通过分析直链淀粉、蛋白质、水分及脂肪酸含量等指标与感官数据的关联模型,最终输出食味值。
-
在水稻成熟期,分别从3个遗传背景下的各突变体及野生型株系中分别选取5个单株,调查统计其株高、分蘖数、每穗粒数、结实率、粒长、粒宽、百粒重。其中粒长和粒宽使用种子测量仪(SC-G型,杭州万深检测科技有限公司)进行分析。
-
在水稻花后第5天分别对3个品种的根、茎、叶、叶鞘组织及种子(5DAF)进行取样,并相继对花后第10天(10DAF)、第15天(15DAF)、第20天(20DAF)的种子进行取样,使用RNA提取试剂盒(A0705A,天根生化科技有限公司)提取总RNA,并使用反转录试剂盒(S20924,全式金生物)合成cDNA。以cDNA为模板,使用CFX96 Real-Time PCR系统(Bio-Rad)和SYBR qPCR Master Mix (Q412-00,Vazyme)进行实时荧光定量PCR (RT-qPCR)检测。所有检测都至少设置了3个生物学重复。以水稻Actin基因(Actin-F:TGCTATGTACGTCGCCATCCAG;Actin-R:AATGAGTAACCACGCTCCGTCA)作为内参。OsAAP13的qPCR的引物为OsAAP13-qF:CAAACGGCCTTGTTCGGCT;OsAAP13-qR: GCAGTTGGACCTCAGGATCG。
-
使用Excel 2021整理数据和作图,SPSS 25.0对数据进行方差分析,显著性水平为0.05。所有数据均以平均值±标准差表示。
-
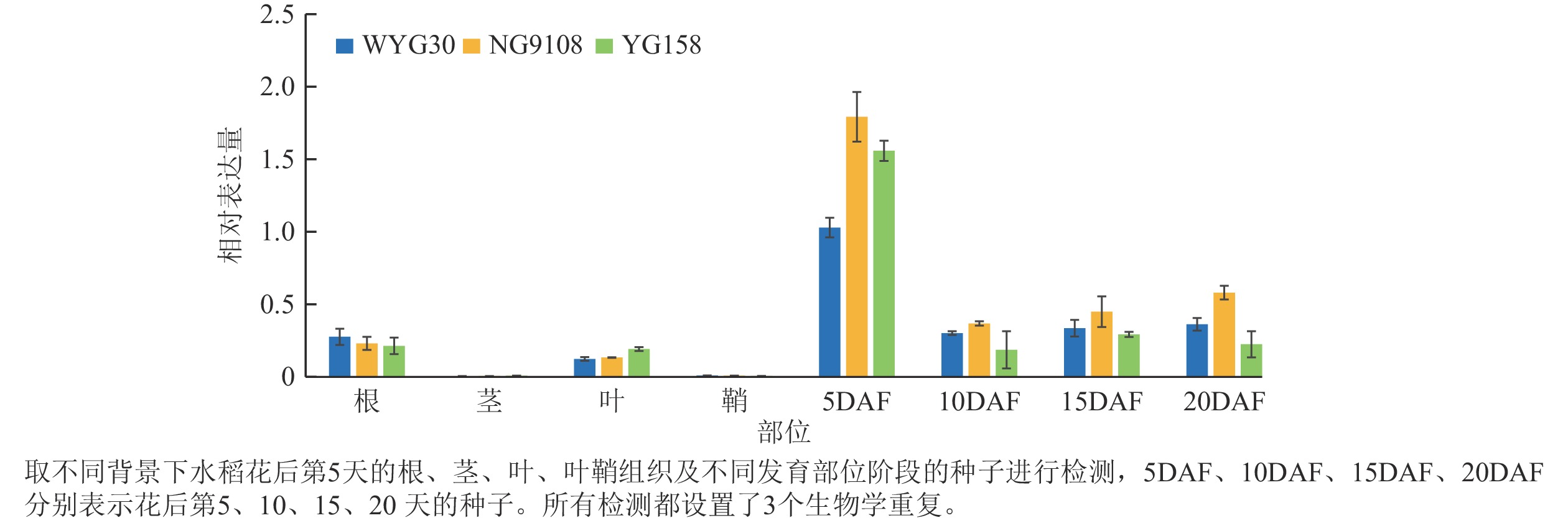
RT-qPCR检测结果表明:OsAAP13在不同水稻品种中均呈现相似的表达趋势,在花后第5 天的种子中表达量较高,随后随着种子成熟的进程显著下调;而在根、叶中则维持较低表达水平,在茎和鞘中基本不表达(图1)。表明OsAAP13在水稻种子发育前期扮演着重要作用。
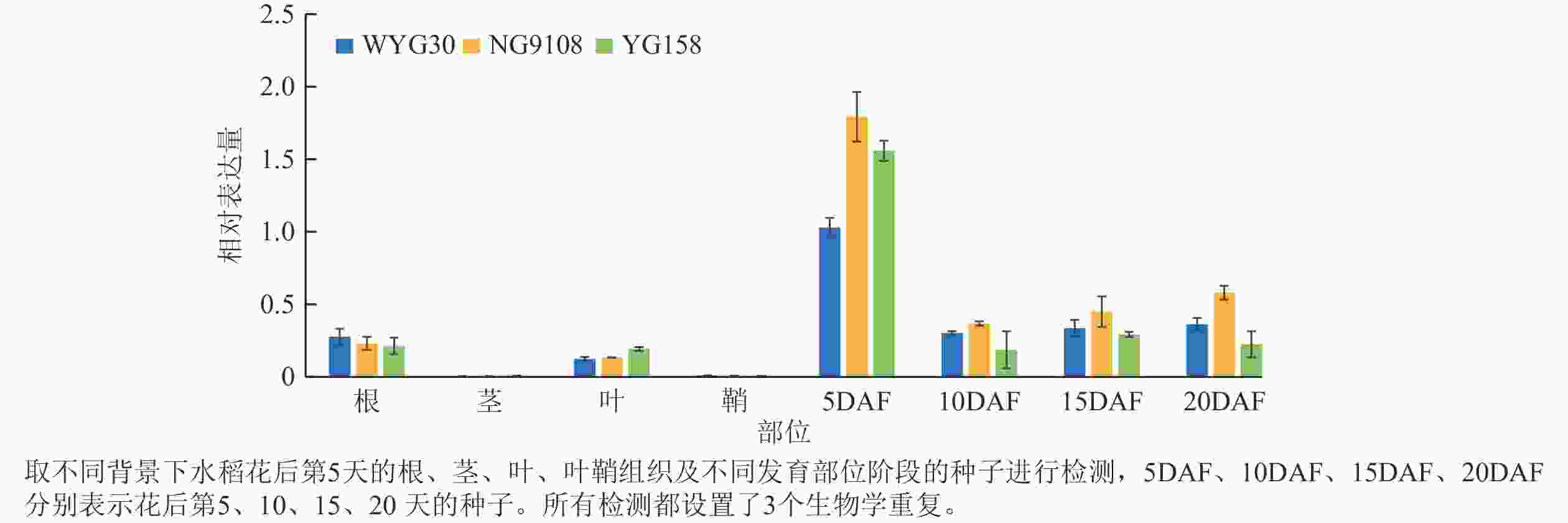
Figure 1. RT-qPCR analysis of OsAAP13
-
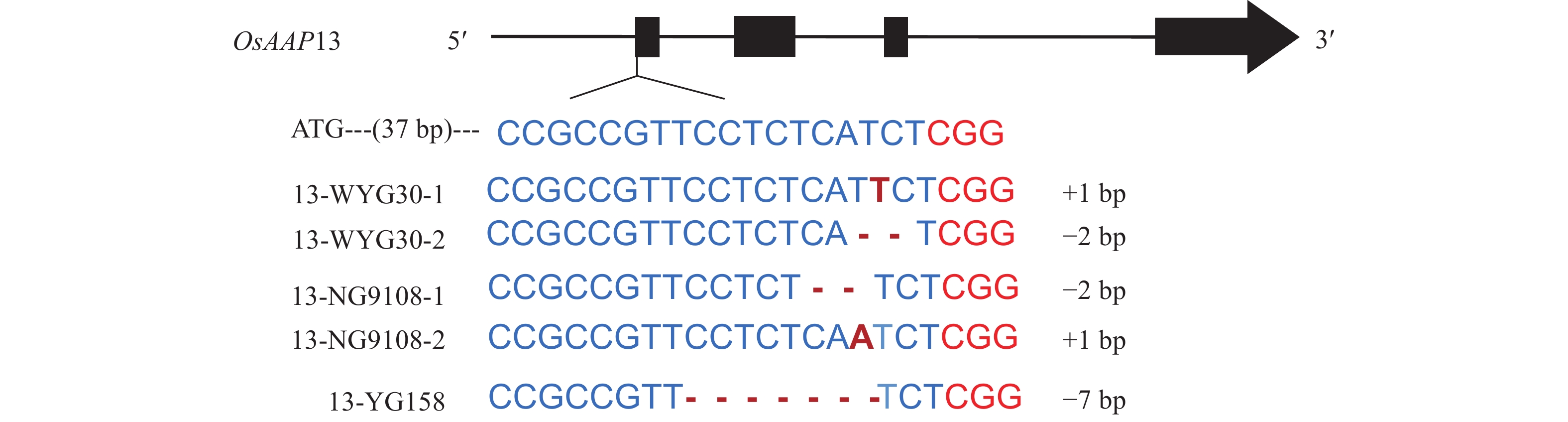
脱靶分析结果如表1所示:OsAAP13靶标在全基因组上的脱靶风险较低。对3个粳稻品种进行OsAAP13进行单独敲除,T0分别获得10、13和13株阳性单株,将获得的T0阳性单株种植成T1株系。利用Cas9引物与潮霉素引物进行阴性单株筛选,对筛选出的阴性单株再进行靶位点序列扩增和测序。最终,WYG30和NG9108分别找出了插入1 bp和缺失2 bp的2种突变类型,YG158找出缺失7 bp的敲除类型(图2)。以上核苷酸的突变均可能造成氨基酸的移码突变。
染色体 位置/bp 序列 脱靶/% 基因 区域 Chr 10 1 934 247 CGACTCCCTTCCTCTCCTCT 0.028 基因间 Chr 1 41 363 176 CGCCGCCGTTCGTCACATCT 0.006 基因间 Chr 3 36 409 463 CGCCGCCGTTCGTCACATCT 0.006 Os03g0862300 内含子 Chr 2 25 001 697 CGCCGCCGTTCGTCACATCT 0.006 基因间 Chr 2 660 940 CGCCGCCATTCGTCACATCT 0.006 基因间 Table 1. OsAAP13 off-target analysis
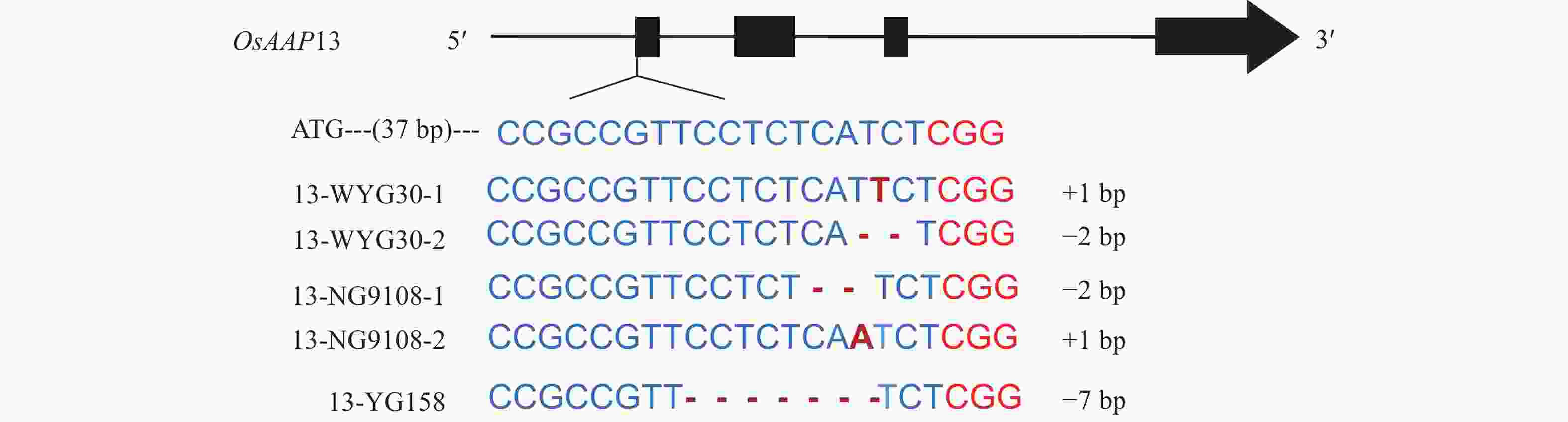
Figure 2. Mutation types of OsAAP13 in different japonica rice culitvars
-
为阐明OsAAP13基因的缺失对水稻农艺性状的影响,考察了3个品种突变体的农艺性状(图3和表2)。结果表明:除13-WYG30-1突变体表现为粒长和百粒重的显著增加外,其余突变体在各农艺性状参数与野生型均无统计学差异。综上,OsAAP13基因敲除对水稻主要农艺性状并无太大影响。

Figure 3. Plants morphology of WT and the OsAAP13 mutants
突变体 株高/cm 分蘖数 每穗粒数 结实率/% 粒长/mm 粒宽/mm 百粒重/g WYG30 100.9±1.3 a 10.0±0.9 a 192.2±16.0 a 93.1±1.4 a 7.21±0.05 b 3.65±0.04 a 2.77±0.14 b 13-WYG30-1 99.8±2.2 a 9.3±2.0 a 185.3±7.1 a 89.3±2.7 a 7.28±0.05 a 3.69±0.07 a 2.82±0.04 a 13-WYG30-2 99.3±1.2 a 10.0±0.8 a 189.3±10.1 a 91.3±1.7 a 7.23±0.15 ab 3.65±0.05 a 2.72±0.14 b NG9108 94.1±2.1 a 10.5±1.4 a 176.8±10.7 a 90.5±1.7 a 7.11±0.1 a 3.85±0.03 a 2.78±0.07 a 13-NG9108-1 92.1±2.4 a 10.7±1.6 a 179.0±19.1 a 88.9±2.6 a 7.16±0.06 a 3.77±0.05 a 2.71±0.05 a 13-NG9108-2 92.3±2.4 a 10.4±1.5 a 185.3±9.5 a 89.3±2.0 a 7.11±0.04 a 3.80±0.01 a 2.71±0.06 a YG158 77.9±3.5 a 8.3±0.6 a 130.2±6.9 a 91.7±2.0 a 7.06±0.29 a 3.64±0.04 a 2.78±0.12 a 13-YG158 78.8±3.1 a 9.0±1.0 a 140.3±23.1 a 89.3±2.9 a 6.98±0.03 a 3.69±0.05 a 2.72±0.04 a 说明:通过单因素方差分析和邓肯多重检验,不同小写字母表示同一品种不同突变体间差异显著(P<0.05)。所有检测都设置了5个生物学重复。 Table 2. Analysis of agronomic characteristics of the wild-type and OsAAP13 mutant lines
-
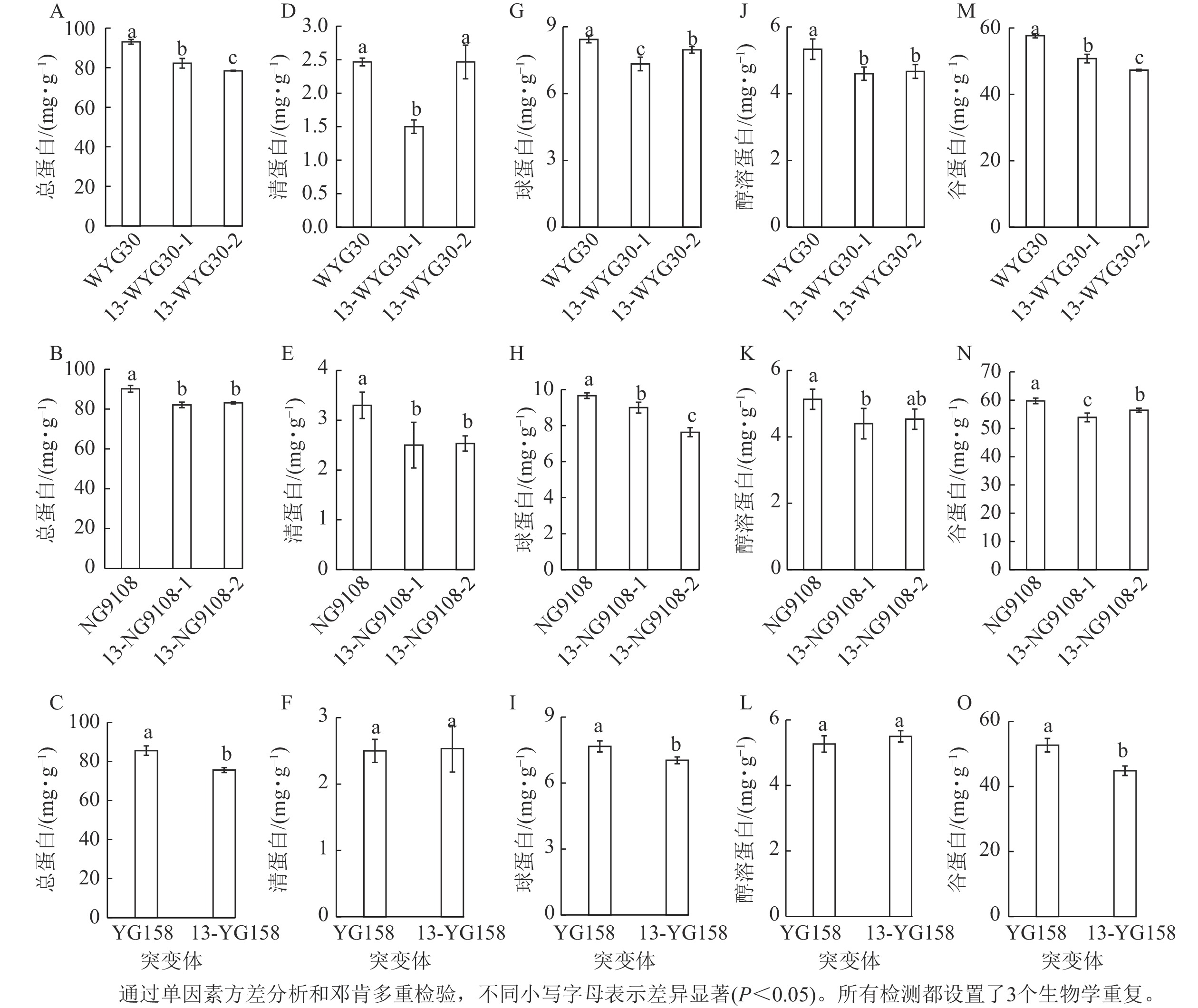
测定3个品种野生型与突变体稻米总蛋白以及组分蛋白,结果表明:WYG30、NG9108和YG158的突变体中的总蛋白质量分数均显著下降(P<0.05),其中WYG30的2个突变体分别降低11.7%和15.8% (图4A),NG9108的2个突变体分别下降9.0%和7.8% (图4B),YG158的突变体下降11.6% (图4C)。4类贮藏蛋白均呈不同程度的下调:清蛋白在NG9108的2个突变体及13-WYG30-1中显著下降(P<0.05,图4D~F);醇溶蛋白在WYG30的2个突变体及13-NG9108-1中显著下降(P<0.05,图4J~L);在贮藏蛋白中占比最多的谷蛋白以及球蛋白在所有突变体中均显著下降(P<0.05),WYG30突变体谷蛋白和球蛋白下降幅度为5.5%~18.0%,NG9108突变体谷蛋白和球蛋白下降幅度为5.5%~21.1%,YG158突变体谷蛋白和球蛋白下降幅度为8.3%~15.0% (图4G~I, M~O)。综上,OsAAP13主要通过调控稻米中的谷蛋白及球蛋白影响总蛋白的积累。
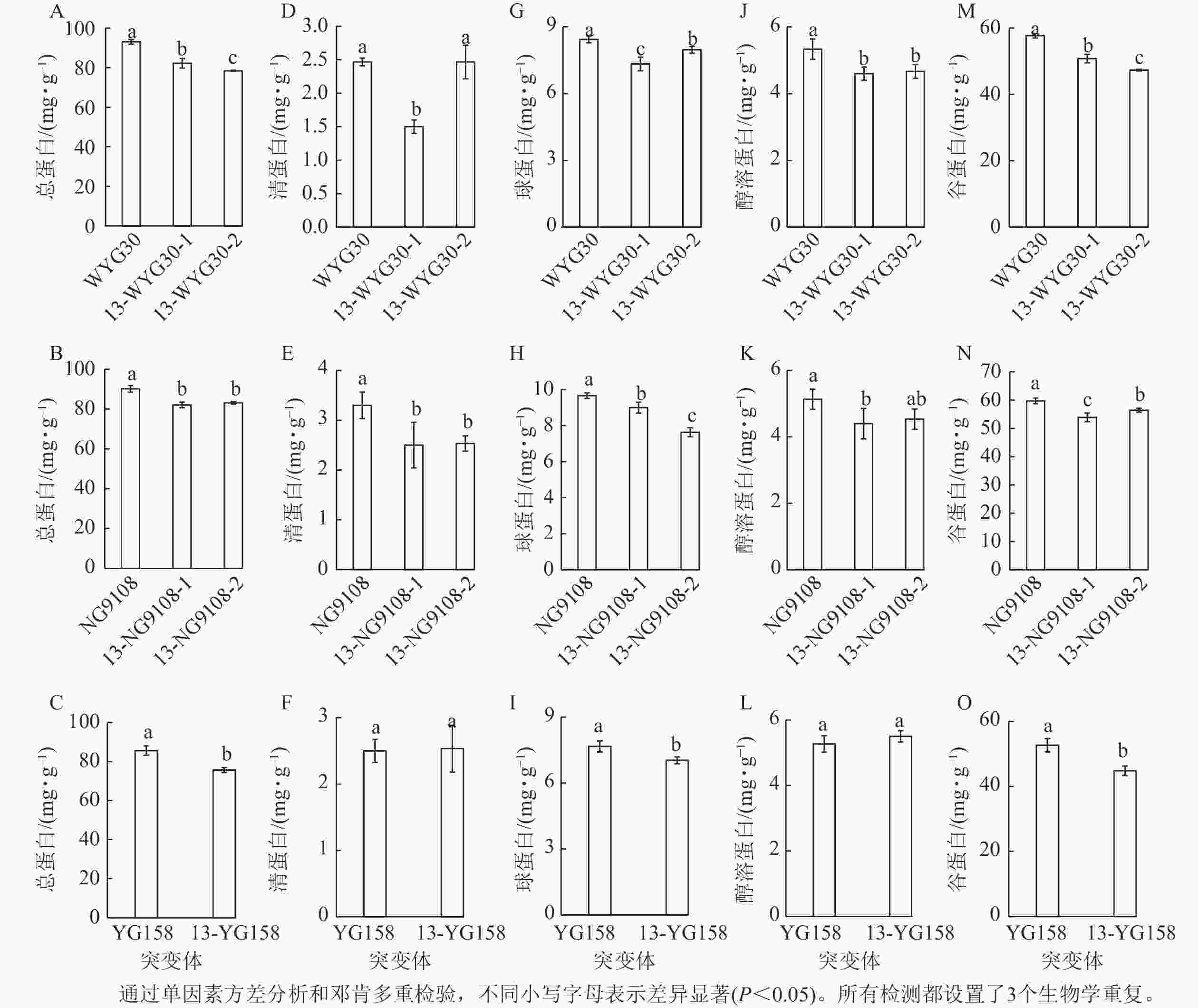
Figure 4. Grain protein contents in wild-type and OsAAP13 mutants
-
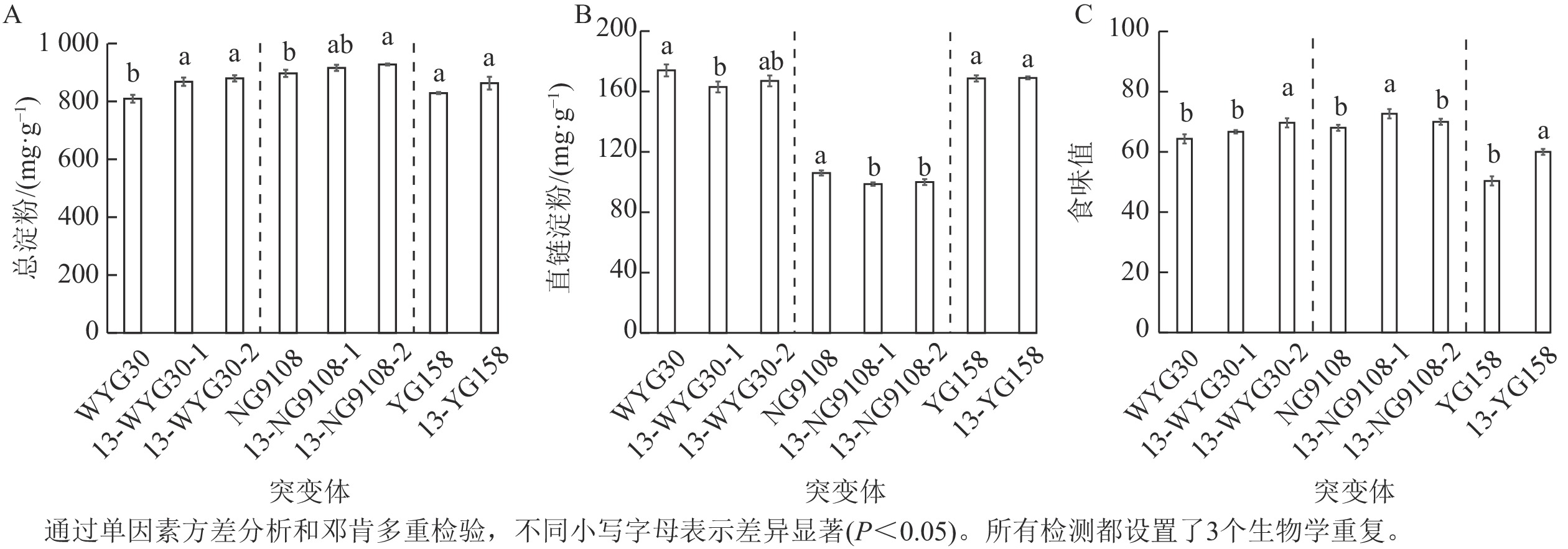
各突变体的总淀粉质量分数有一定的提升:NG9108的2个突变体分别增加了2.1%和3.4%;YG158突变体提升4.1%;WYG30的2个突变体增幅最为显著,达7.3%和8.7% (图5A)。与之相反的是,各突变体直链淀粉质量分数总体却呈小幅度下调趋势,下降幅度为4.0%~6.9%,在13-WYG30-1、13-NG9108-1和13-NG9108-2突变体中下调达显著水平(P<0.05,图5B)。虽然不同品种突变体的变化幅度略有差异,但整体变化趋势一致,表现为总淀粉质量分数上升,直链淀粉质量分数下降。综上,OsAAP13对稻米淀粉合成与代谢具有重要影响。
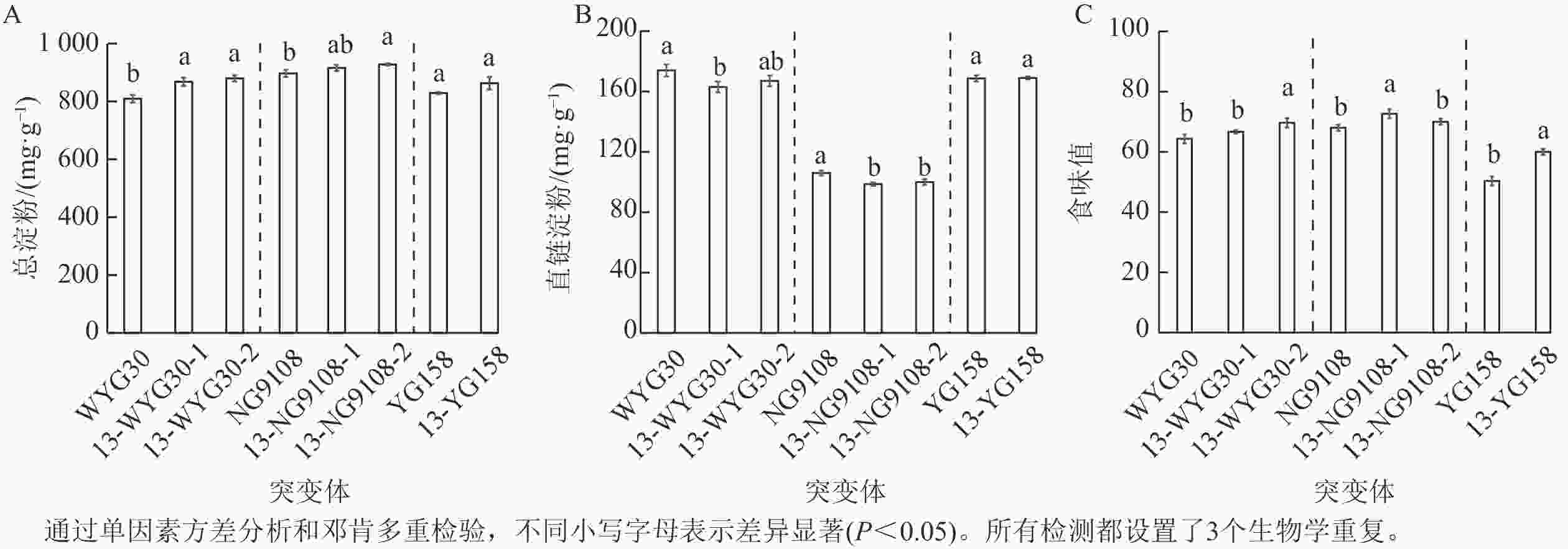
Figure 5. Total starch and amylose contents, and tase scores of OsAAP13 mutants
-
进一步评估不同遗传背景突变体的稻米食味值表明:所有突变体的稻米食味值均发生不同程度提升:WYG30的2个突变体分别提升3.6%和8.3%,NG9108的2个突变体增幅为6.7%和2.9%,YG158的突变体显著提升了11.6% (P<0.05,图5C)。表明OsAAP13基因的缺失可有效改善稻米食味值。
-
氨基酸是植物运输有机氮的主要形式,膜定位氨基酸转运体(AATs)可介导氨基酸从根瘤到种子的移动,在许多生物过程中起到了重要的作用[25]。已有研究表明:不同植物物种之间的氨基酸转运蛋白的氨基酸序列是非常保守的,AATs的同源基因相似性可高达98%,尤其在跨膜结构域的保守性高度一致,对维持转运活性至关重要[10, 26−27]。在本研究中,虽然YG158未获得2个向上的突变体类型,但由于OsAAP13在3个品种中具有相似的表达模式,缺失OsAAP13对3个品种稻米品质的影响也具有相似的变化趋势。因此,可以认为OsAAP13对稻米品质改良具有重要影响。
已有大量研究论述了蛋白质含量对稻米食味值的影响。如YANG等[28]系统测定了近100个粳稻品种的总蛋白含量、组分蛋白含量和食味值,相关分析表明稻米总蛋白和谷蛋白含量与食味值均呈极显著负相关。CAI等[29]分析了近30 a南方稻区籼稻、粳稻主栽品种及骨干亲本的食味品质性状,结果表明,蛋白质含量的降低是南方水稻品种食味品质获得改良的关键因素。以上研究均直接或间接表明稻米蛋白质含量过高对米饭食味具有负面效应。一般情况下优质稻米的蛋白质质量分数低于7%,如日本优质粳稻品种‘越光’‘Koshihikari’,携带一个编码颗粒结合型淀粉合成酶Ⅰ(Granule-Bound Starch Synthase Ⅰ, GBSS Ⅰ) 的Wxb等位基因[30],其稻米蛋白质质量分数低于7%,不仅口感出众,且外观透明,备受欢迎。‘稻花香’‘Daohuaxiang’和‘空育131’‘Kongyu 131’等中国优质水稻品种也表现出类似的品质特性。生产实践中,由于中国施氮水平较高和品种特性等原因,普通稻米的蛋白质含量普遍较高,严重影响了稻米的食味品质。因此,亟待挖掘调控稻米蛋白质含量的基因资源和开发快速改良稻米蛋白质含量的方法。
CRISPR/Cas9技术是近10 a发展起来的革命性技术,可对植物基因组特定位置产生可预见和可遗传的改变,从而实现对植物性状的精准改良,且无转基因争议,应用前景十分广阔[31]。YANG等[28]利用CRISPR/Cas9系统,在2个高产粳稻背景下同时敲除胚乳特异表达的8个谷蛋白合成基因,从而特异性降低了水稻胚乳中谷蛋白的合成,成功筛选出多个稻米蛋白质含量显著下调、米饭食味品质显著提升的新种质材料。此外,在4个高产粳稻品种/品系中水稻氨基酸转运蛋白OsAAP6、OsAAP10和OsAAP13的敲除实验表明,基因敲除可实现快速降低稻米蛋白质含量并改善其蒸煮食味品质[21−22]。本研究利用CRISPR/Cas9技术,在不同粳稻背景下对在胚乳中高表达的OsAAP13基因进行敲除,结果显示:OsAAP13敲除可提升稻米总淀粉含量,降低贮藏蛋白含量,提升米饭适口性。
-
OsAAP13基因主要在水稻花后第5天的种子中高效表达。本研究通过CRISPR/Cas9技术对不同材料进行OsAAP13基因的定向敲除,能抑制种子中贮藏蛋白的积累、促进淀粉的合成和稻米食味值提升。本研究为稻米蒸煮食味品质改良提供重要基因资源和理论基础。
Improving rice grain quality by knocking out OsAAP13 using CRISPR/Cas9 technology
doi: 10.11833/j.issn.2095-0756.20250177
- Received Date: 2025-03-07
- Accepted Date: 2025-07-07
- Rev Recd Date: 2025-07-05
- Available Online: 2025-08-01
- Publish Date: 2025-08-01
-
Key words:
- Oryza sativa /
- rice quality /
- CRISPR/Cas9 /
- OsAAP13
Abstract:
| Citation: | WU Lingjun, SHENG Shujie, SHAN Xinlan, et al. Improving rice grain quality by knocking out OsAAP13 using CRISPR/Cas9 technology[J]. Journal of Zhejiang A&F University, 2025, 42(4): 694−702 doi: 10.11833/j.issn.2095-0756.20250177

|

 DownLoad:
DownLoad: