






-
百山祖冷杉Abies beshanzuensis隶属于松科Pinaceae冷杉属Abies,于1963年被首次发现,并于1976年被正式鉴定及命名为新物种,目前仅自然分布于浙江省庆元县百山祖海拔约1 750 m的区域[1],被列为国家一级重点保护野生植物[2],且被列为全球最濒危的12种植物之一[2–3]。该物种起源古老,是低纬度地区的冷杉属等高寒植物在第四纪冰川末期从低海拔向高海拔山体退缩的佐证,自然生长的成熟个体现仅存3株,被誉为“植物活化石”。由于其环境适应能力弱且自然繁殖能力低,该物种长期处于极度濒危的状态[4]。该物种对于这一地区的植物区系发生和演变,以及植物学、生态学、地理学和气候学等具有很高的科研价值[5]。
生长量是反映植物生长状况最直接的指标之一,被用来直观地显示环境对植物的生存状态造成的影响[6–7]。叶绿素荧光参数是一种非破坏性的测量技术,专门用于评估光系统Ⅱ(PSⅡ)的活性,能够在不破坏叶片细胞结构的前提下,快速获取相关参数信息。此技术目前已在植物生理学研究中得到广泛应用[8–9],尤其在深入探究光合作用机制、植物对环境变化的适应性响应、遗传多样性分析以及生态系统多样性评估等方面发挥了重要作用[10–11]。因此,近年来,叶绿素荧光技术已在多个领域广泛应用,包括高海拔植物光合适应性的研究[12−14]、植物在不同胁迫条件下光合生理响应的探讨[15−17]、不同物种间光合生理特性差异性的分析[18–19]、遮阴对植物光合特性影响的研究[20–21],以及从光合作用角度探讨濒危物种致濒机制的解析等[22−24]。为了改善百山祖冷杉的濒危状况,针对其濒危机制及抢救性保护措施展开了深入研究。近年来,除了传统的嫁接繁育技术,一些利用转录组学、代谢组学等手段对其遗传多样性、濒危机制及保护策略的研究也取得了新的认识[25–26]。选择1年生小枝和叶片作为研究对象的主要原因:其一、叶片在成熟后光合能力强,叶绿素荧光参数相对稳定和可靠;其二、取材量相对较少,对植株损坏较小,尤其是原生母树;其三、2年生及以上年龄枝条上的叶片可能因各种原因引起落叶或损坏,对统计数据会有影响。因此,本研究通过对比自然条件下的百山祖冷杉原生母树与嫁接树1年生枝叶的生长指标,并在主要生长季节持续监测其叶片的叶绿素荧光参数,旨在深入揭示百山祖冷杉原生母树与嫁接树在生长状态及叶绿素荧光参数的差异,进而为百山祖冷杉的精准保护策略提供科学依据。
-
采样点位于浙江省庆元县钱江源-百山祖国家公园,原生母树采样点位于海拔约1 750 m的山地林中,无遮阴、高光强,土壤瘠薄;嫁接树采样点位于海拔约1 550 m的百山祖国家公园庆元保护中心百管站苗圃内(嫁接株不能种植在原生母树周围,否则花粉会对原生母树所结种子产生影响),生境条件相对优越,有大树和西面房屋部分遮阴,土壤较肥厚。选择生长正常的原生母树(树龄约170 a)和以日本冷杉 Abies firma为砧木的嫁接树(树龄约42 a)的健康1年生枝条。于2023年生长季4—9月,每月进行1次采样。
-
2023年8月,选取原生母树与嫁接树长势良好且具有代表性的1年生枝条,使用游标卡尺(精度0.01 cm)测量枝条上叶片的长度和宽度、小枝的长度和直径。使用分析天平(精度0.000 1 g)称量1年生枝条的鲜质量和枝条上叶片鲜质量,并对叶片数量进行计数。均采样3次进行测量,取其平均值。
-
叶绿素荧光测定采用沈宗根等的方法[27],使用德国Mini-PAM便携式调制叶绿素荧光仪测定叶片叶绿素荧光参数。选取百山祖冷杉原生母树与嫁接树1年生枝条上的叶片,将叶片置于黑暗处进行暗处理30 min后,激发光的光强(PAR)梯度设置为0、81、130、195、268、396、540、816、1 190 μmol·m−2·s−1,依次测定叶片的光合量子产量(Y)、电子传递速率(RET)、光化学猝灭系数(CPQ)和非光化学猝灭系数(CNPQ),重复3次。测定时始终使叶片垂直于入射光,为减少误差,不同处理的叶片交叉测定。4—9月每月测定1次,共计6次。
-
使用软件Excel 2019和SPSS 26对数据进行统计分析和方差分析,采用Pearson相关系数对各项指标进行相关性分析,使用软件GraphPad Prism 9.5进行作图和分析。
-
如表1所示:百山祖冷杉原生母树的1年生枝叶生长量指标小枝长、叶片数、叶长、小枝直径、叶鲜质量、小枝鲜质量均显著小于嫁接树(P<0.05)。8月,原生母树的小枝长为56.39 mm±2.66 mm,而嫁接树的小枝长为78.66 mm±5.87 mm;原生母树的小枝直径为3.20 mm±0.15 mm,嫁接树的小枝直径为3.71 mm±0.21 mm;原生母树叶片数为67.33片±2.91片,嫁接树叶片数为79.33片±2.40片;母树叶长为33.76 mm±2.56 mm;嫁接树叶长为45.85 mm±0.38 mm;原生母树叶鲜质量为2.32 g±0.25 g,嫁接树叶鲜质量为4.93 g±0.21 g;原生母树小枝鲜质量为0.36 g±0.03 g,嫁接树小枝鲜质量为0.69 g±0.02 g。而叶宽在原生母树和嫁接树之间无显著差异。
样树 小枝长/mm 小枝直径/mm 叶片数/片 叶长/mm 叶宽/mm 叶鲜质量/g 小枝鲜质量/g 原生母树 56.39±2.66 b 3.20±0.15 b 67.33±2.91 b 33.76±2.56 b 3.49±0.22 a 2.32±0.25 b 0.36±0.03 b 嫁接树 78.66±5.87 a 3.71±0.21 a 79.33±2.40 a 45.85±0.38 a 3.93±0.45 a 4.93±0.21 a 0.69±0.02 a 说明:数据为平均值±标准误,同列不同小写字母表示差异显著(P<0.05,独立样本t检验)。 Table 1. Growth of current-year branches and leaves of the original mother and grafted trees of A. beshanzuensis
-
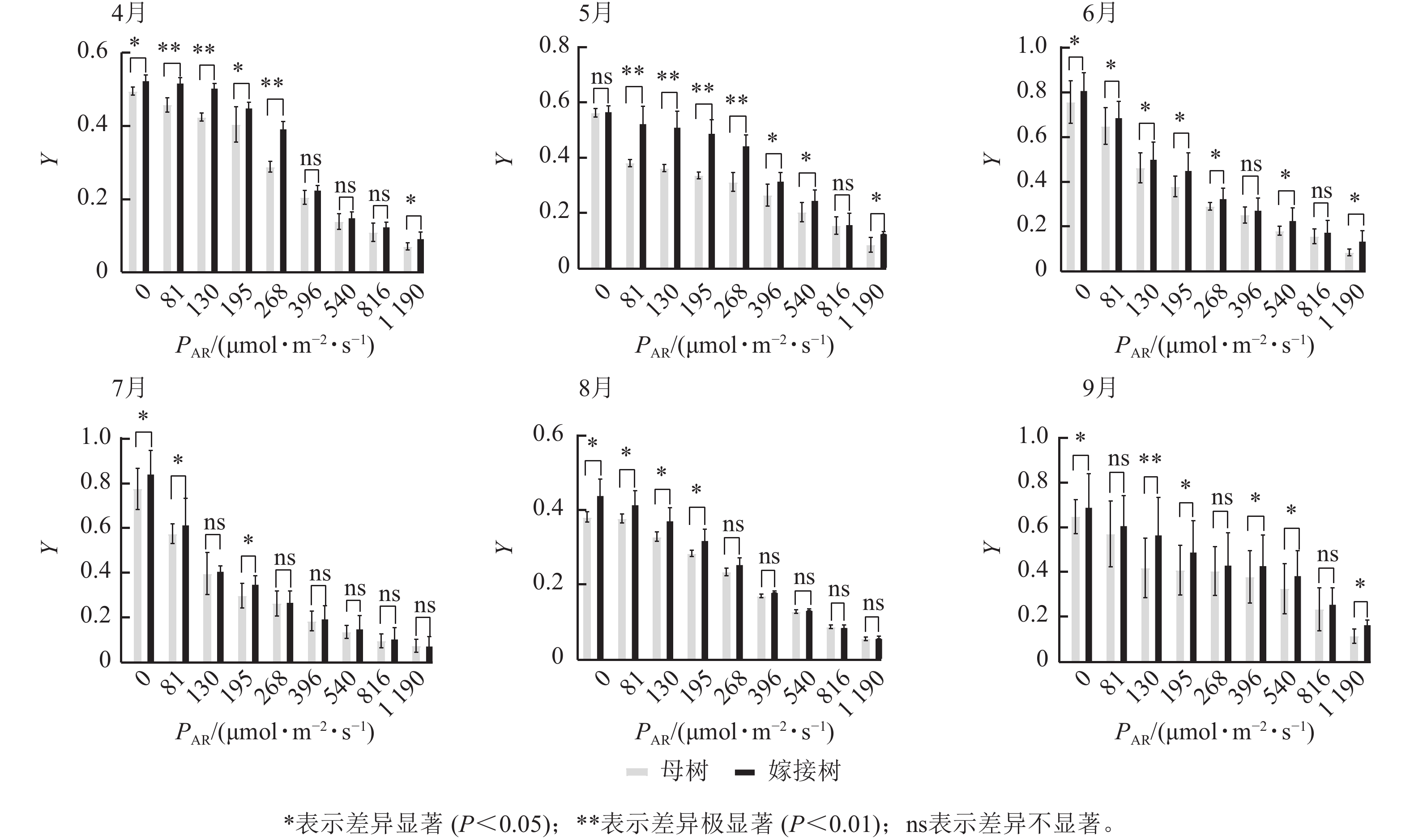
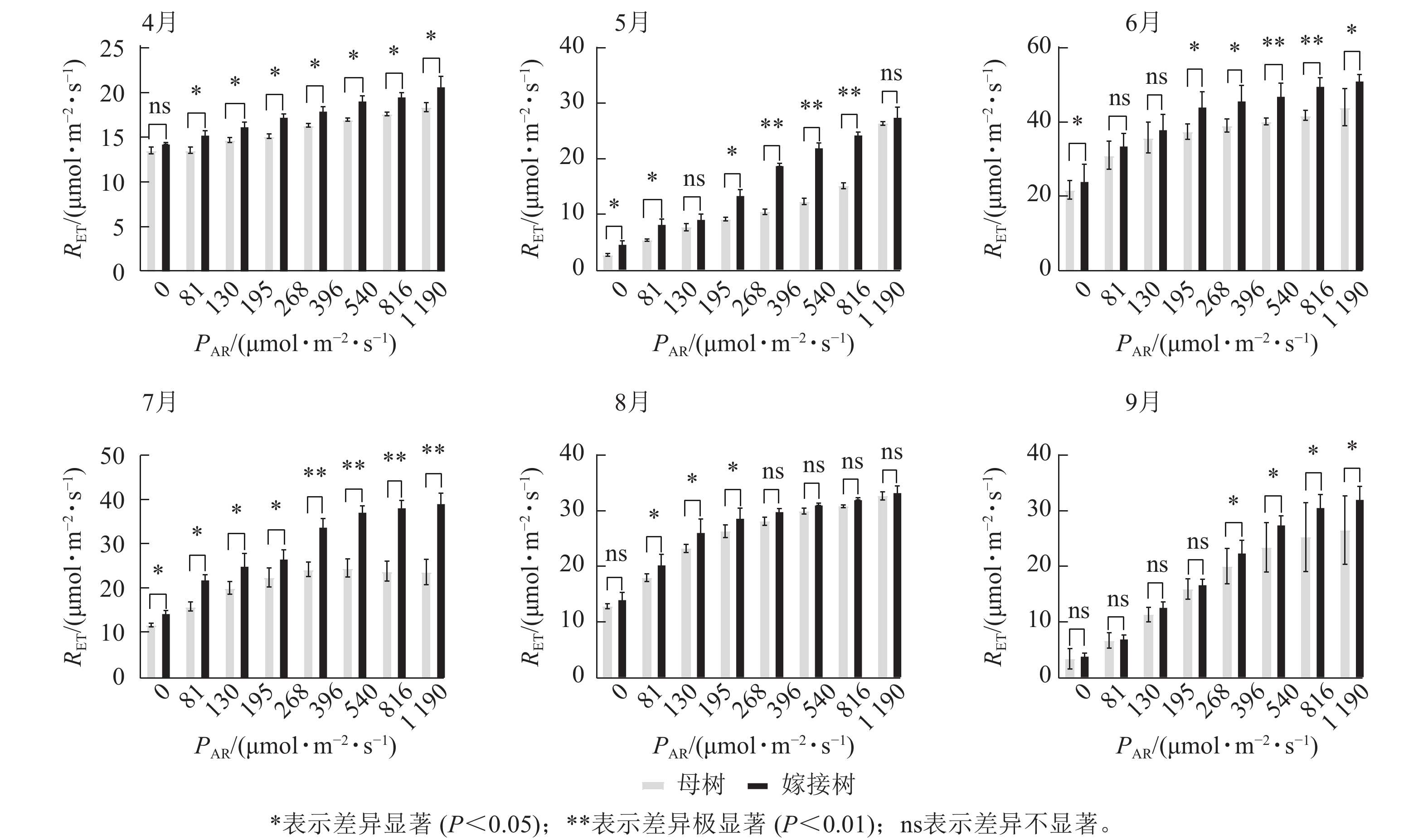
6个月的持续监测(图1)表明:百山祖冷杉原生母树与嫁接树的叶片光合量子产量均随光强的增高而降低。在整个监测期间,同一光强下原生母树的光合量子产量均低于嫁接树:4月,在0、195、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在81、130、268 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);5月,在396、540、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在81~268 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);6月,在0、81、130、195、268、540、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05);7月,在0、81、195 μmol·m−2·s−1等光强条件下差异显著(P<0.05);8月,0~195 μmol·m−2·s−1等光强条件下差异显著;9月,在0、195、396、540、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在130 μmol·m−2·s−1光强条件下差异极显著(P<0.01)。这表明原生母树的光能捕获效率低于嫁接树。图2显示:原生母树与嫁接树的电子传递速率均随着光强的增高而增加,表明百山祖冷杉具有较强的光合能力。然而,在同一光强下,原生母树的电子传递速率均低于嫁接树:4月,除81 μmol·m−2·s−1光强条件外,其余光强条件下均差异显著(P<0.05);5月,在81、130、268 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在396、540、816 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);6月,在81、268、396、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在540、816 μmol·m−2·s−1等光强条件下差异极显著(P<0.05);7月,在81~268 μmol·m−2·s−1等光强条件下差异显著(P<0.05),≥396 μmol·m−2·s−1光强条件下差异极显著(P<0.01);8月,仅在130~268 μmol·m−2·s−1光强条件下差异显著(P<0.05);9月,≥396 μmol·m−2·s−1等光强条件下差异显著(P<0.05)。这表明原生母树的电子传递速率较嫁接树弱。综合以上2个指标发现:原生母树的光合能力显著低于嫁接树(P<0.01)。
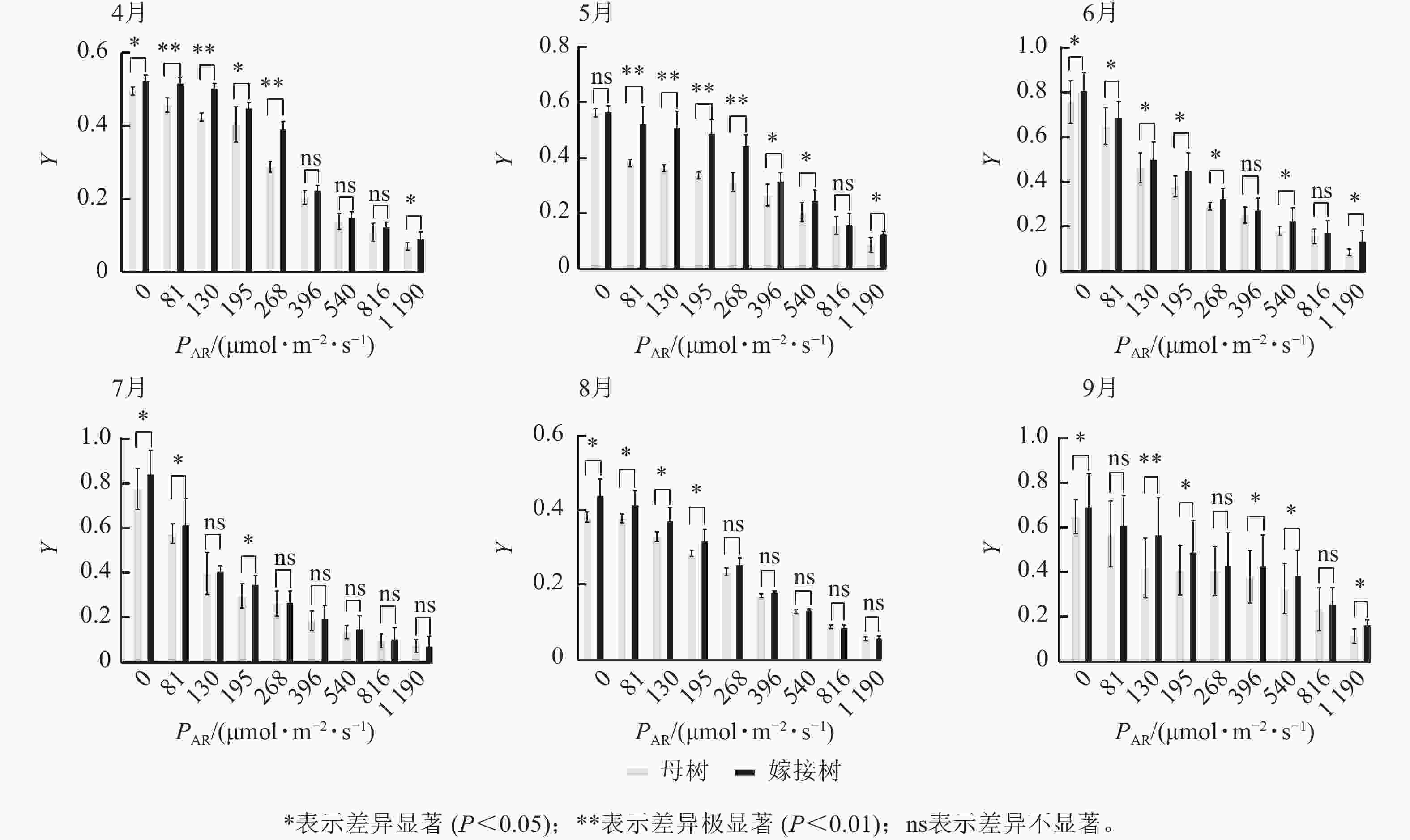
Figure 1. Comparison of the response curves of photosynthetic quantum yield (Y) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
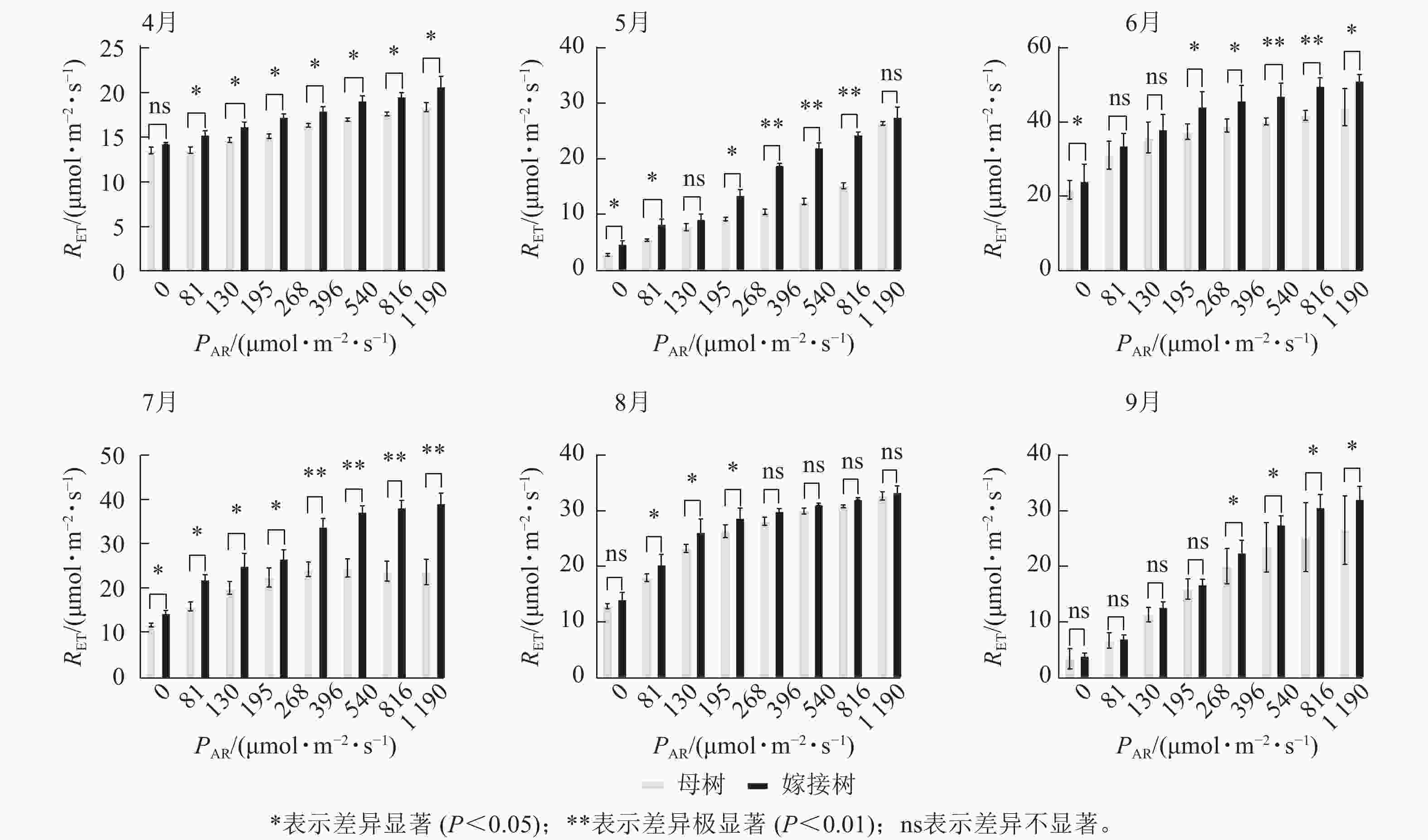
Figure 2. Comparison of the response curves of electron transport rate (RET) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
-
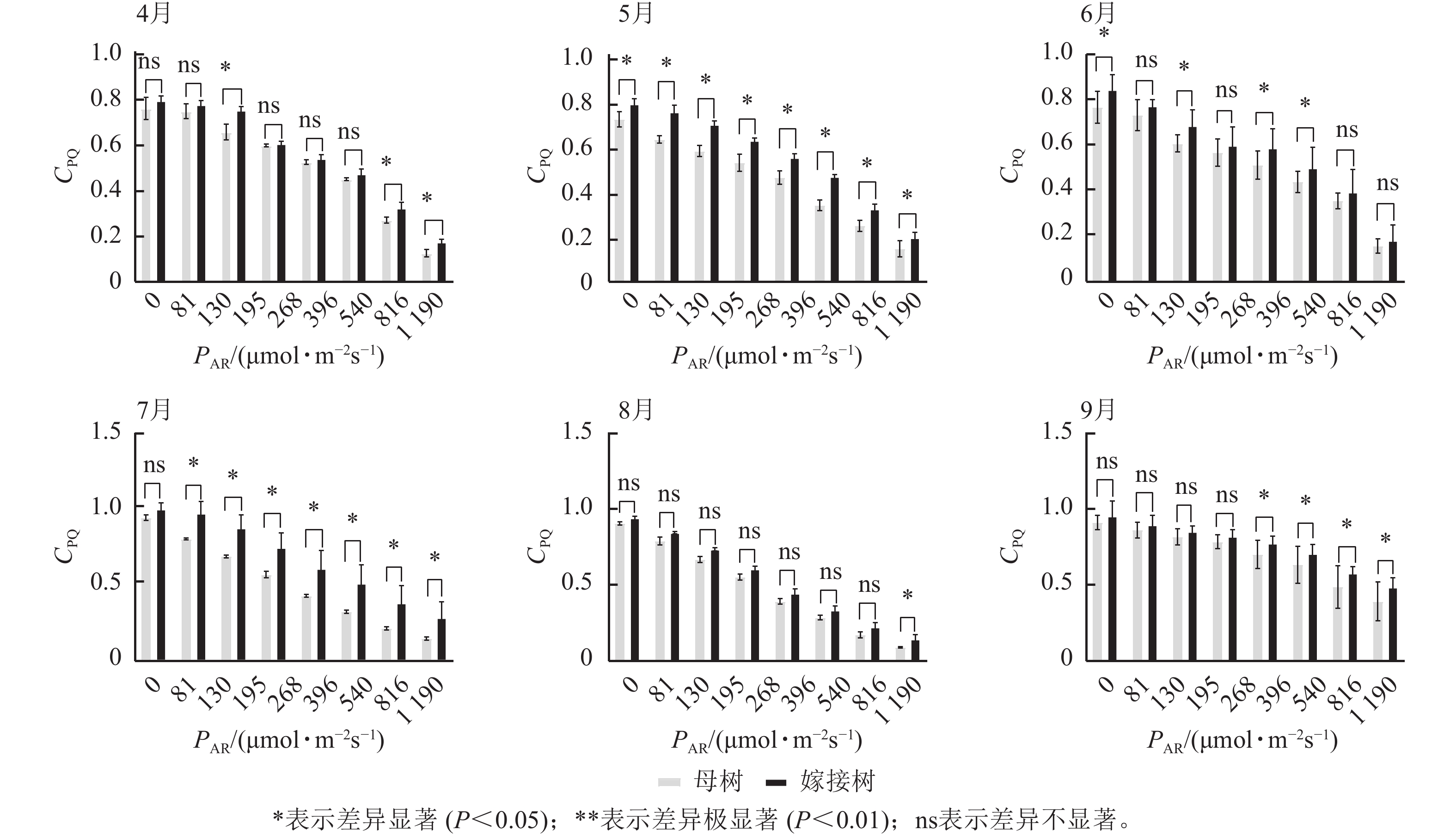
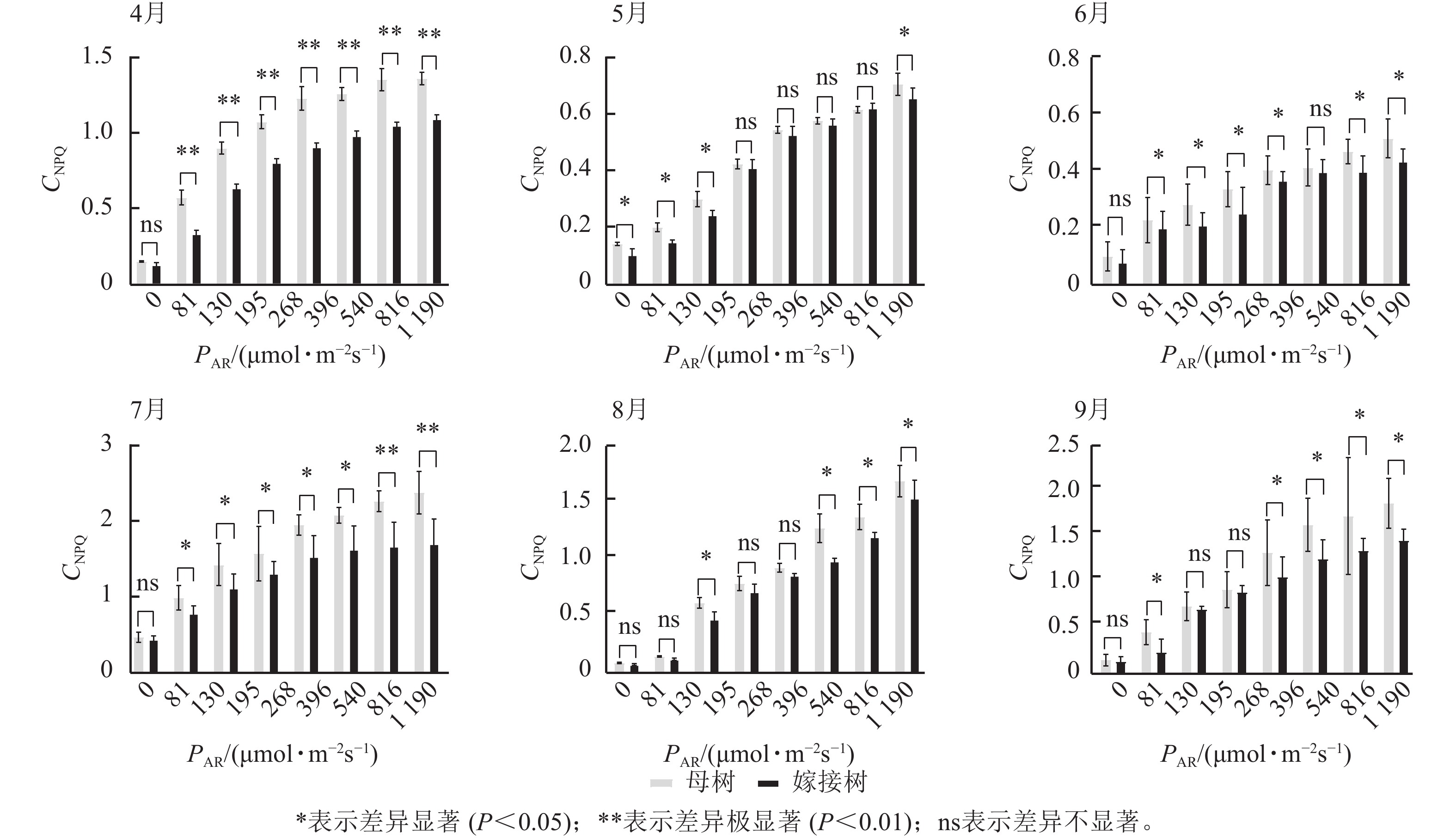
图3显示:原生母树与嫁接树的光化学猝灭系数随着光合有效辐射的升高而降低,且在同一光强下,原生母树的光化学猝灭系数均低于嫁接树:4月,在195、816、
1190 μmol·m−2·s−1等光强条件下差异显著(P<0.05);5月,所有实验的光强条件下均差异显著(P<0.05);6月,在81、195、396、540 μmol·m−2·s−1等光强条件下差异显著(P<0.05);7月,除81 μmol·m−2·s−1外,其他光强下均差异显著(P<0.05);8月,仅1190 μmol·m−2·s−1光强条件下差异显著(P<0.05);9月,≥396 μmol·m−2·s−1光强条件均差异显著(P<0.05)。图4显示:原生母树与嫁接树的非光化学猝灭系数随着光强的升高而升高,光合作用系统完整,并未因受到环境胁迫而破坏。且同一光强下原生母树的非光化学猝灭系数均高于嫁接树:4月,除81 μmol·m−2·s−1外,其他光强下均差异极显著(P<0.01); 5月,在81、130、195、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05);6月,除81、816 μmol·m−2·s−1外,其他光强下均差异显著(P<0.05);7月,在130~540 μmol·m−2·s−1光强条件下差异显著(P<0.05),在816、1190 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);8月,在195和≥540 μmol·m−2·s−1等光强条件下差异显著(P<0.05),其余光强条件下无显著差异;9月,除81、195、268 μmol·m−2·s−1等光强条件下无显著差异外,其他光强条件下均具显著差异(P<0.05)。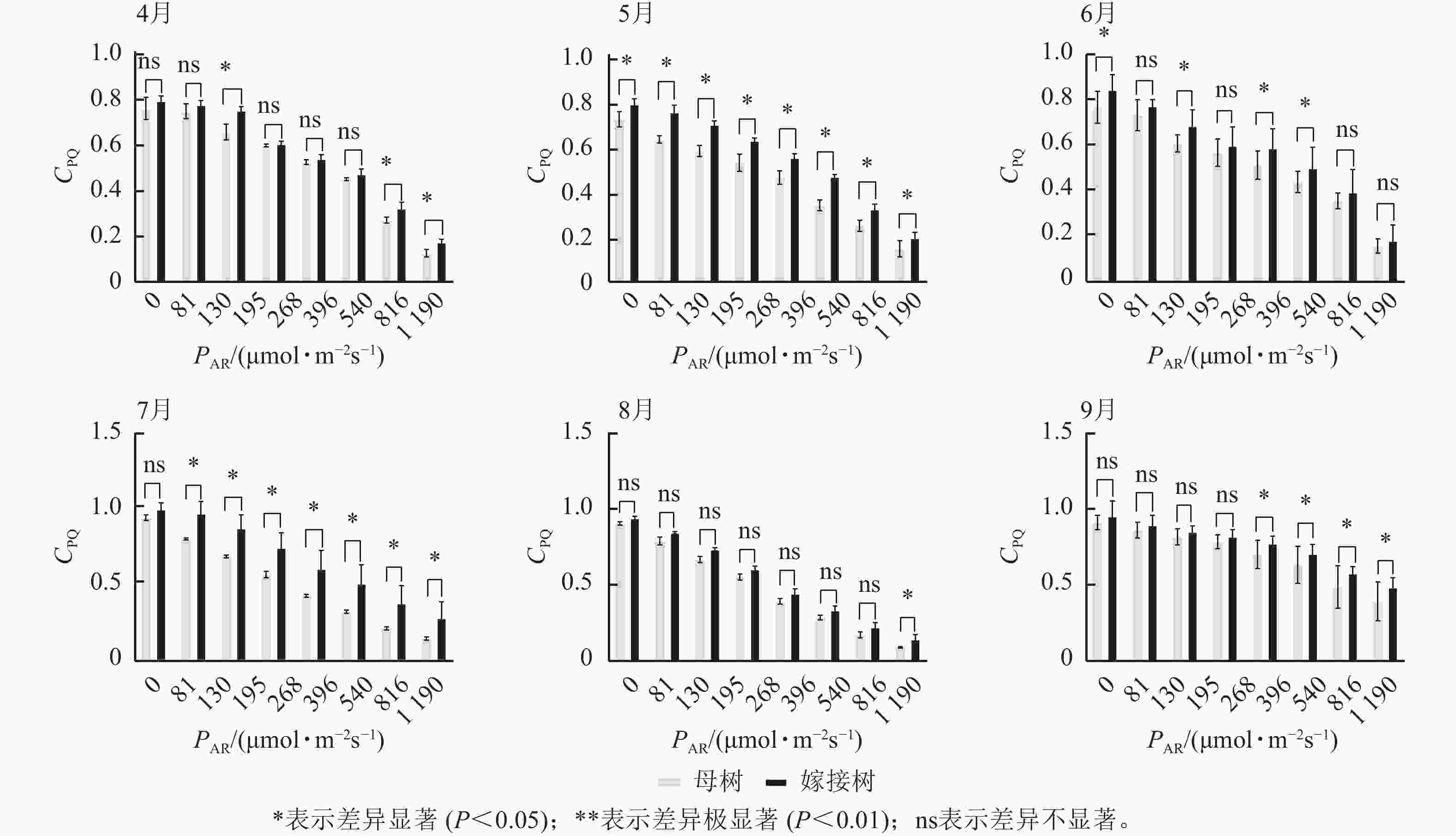
Figure 3. Comparison of the response curves of photochemical quenching coefficent (CPQ) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
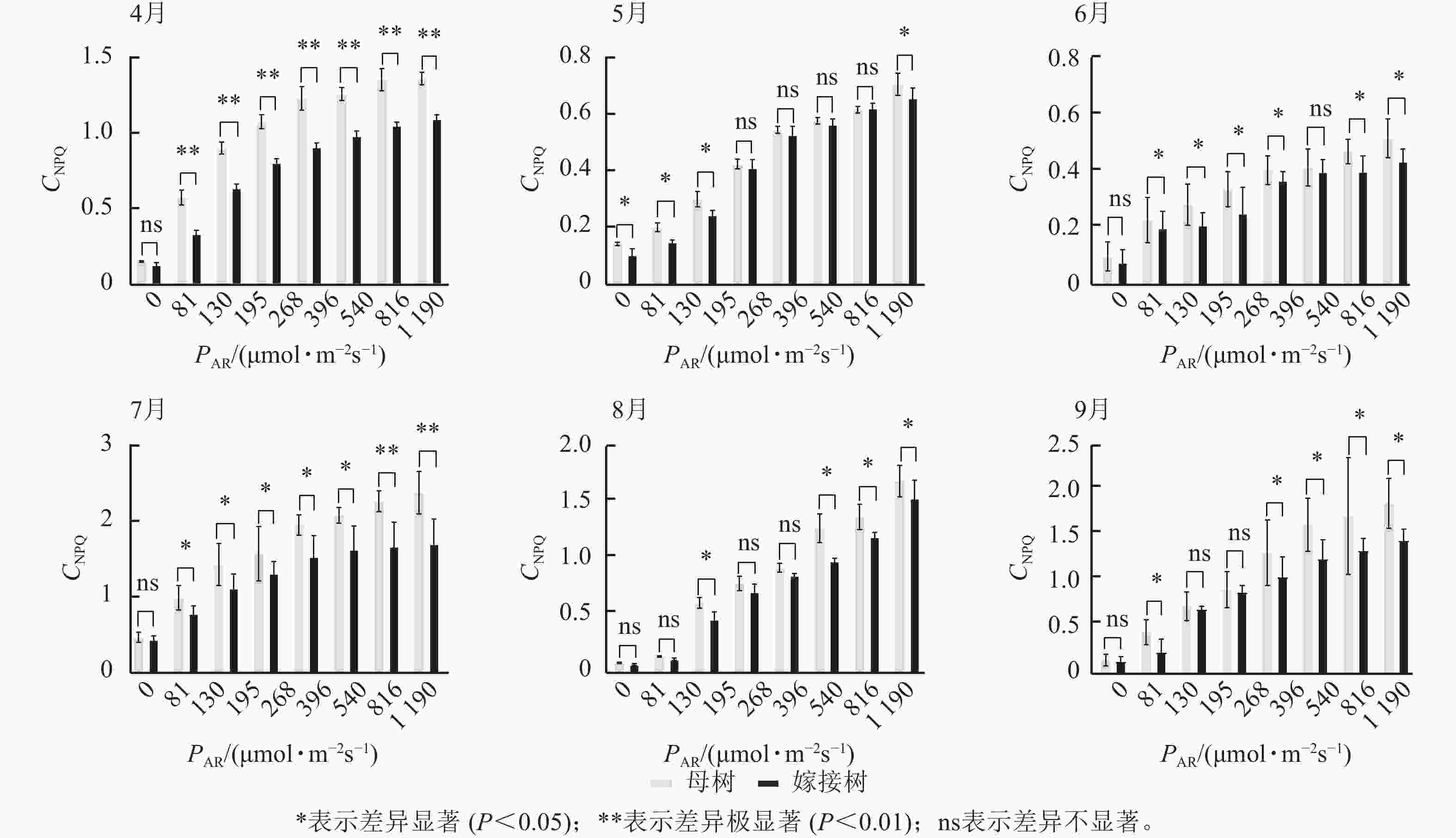
Figure 4. Comparison of the response curves of photochemical quenching coefficent (CNPQ) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
-
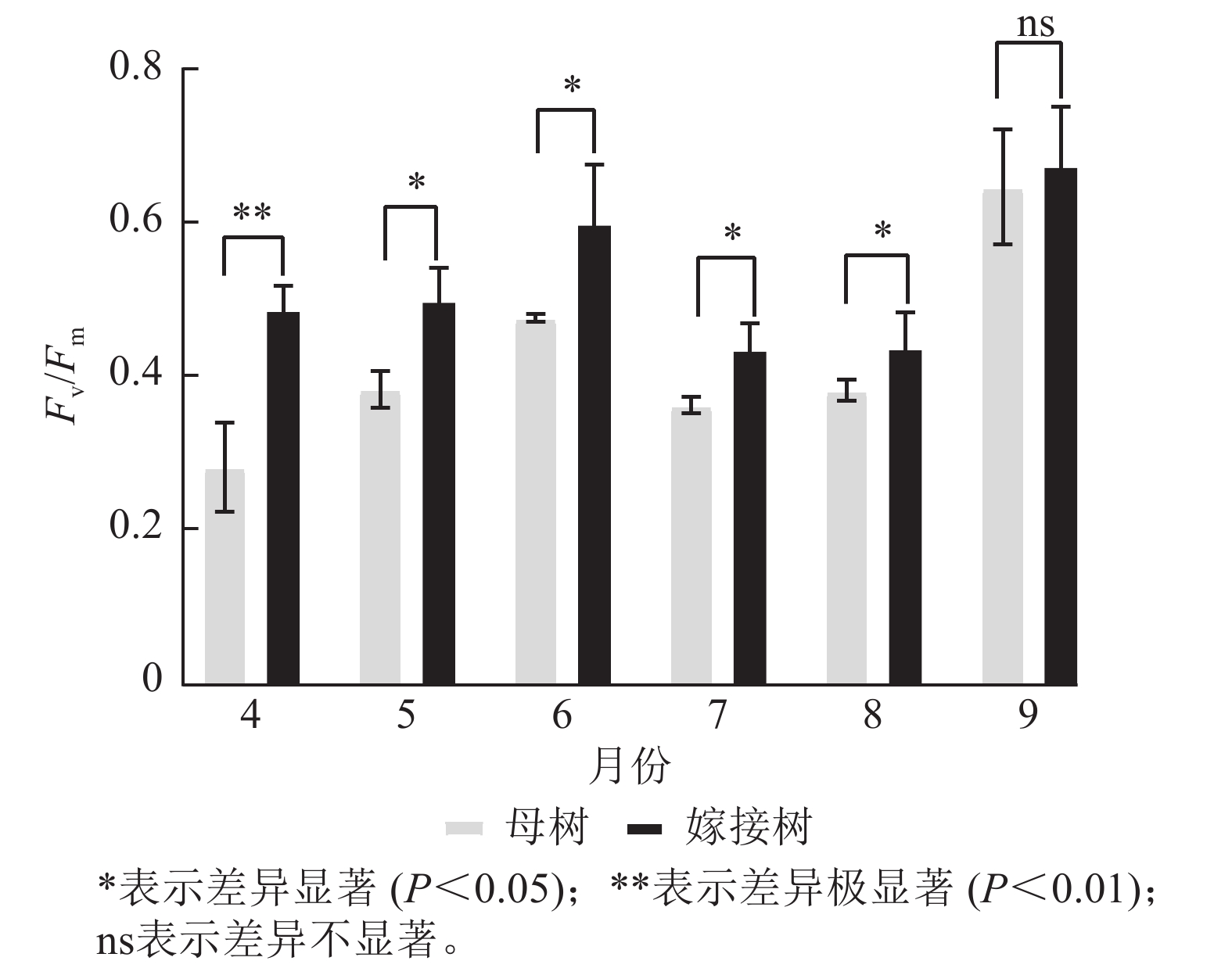
图5显示:原生母树与嫁接树的最大光化学量子产量均在4—6月呈上升趋势:在6—8月呈下降趋势,8—9月开始上升。且嫁接树的最大光化学量子产量均高于原生母树,4月差异极显著(P<0.01),5—8月则差异显著(P<0.05)。
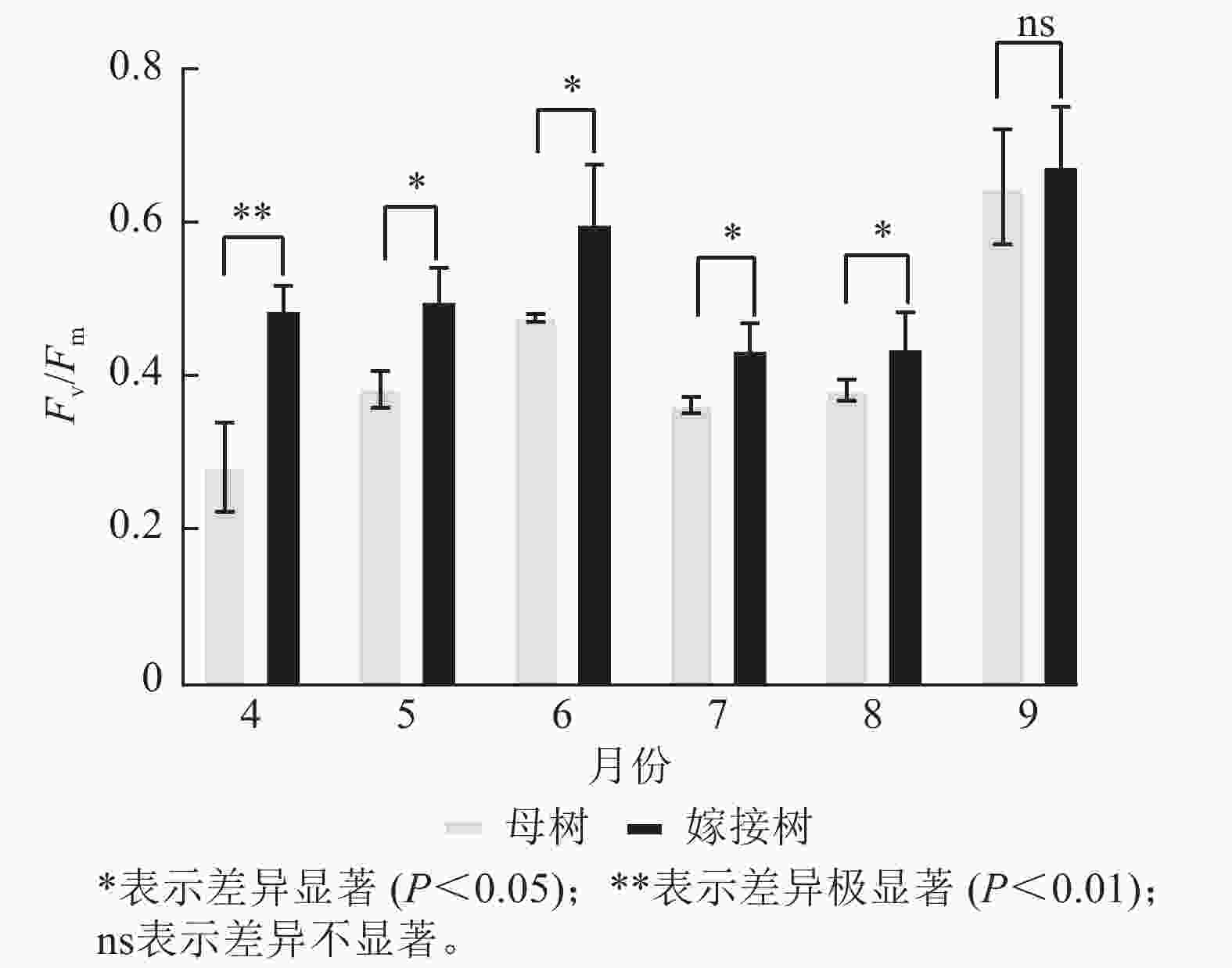
Figure 5. Comparison of the maximum photochemical quantum yield (Fv/Fm) between the original mother and grafted trees of A. beshanzuensis from April to September
-
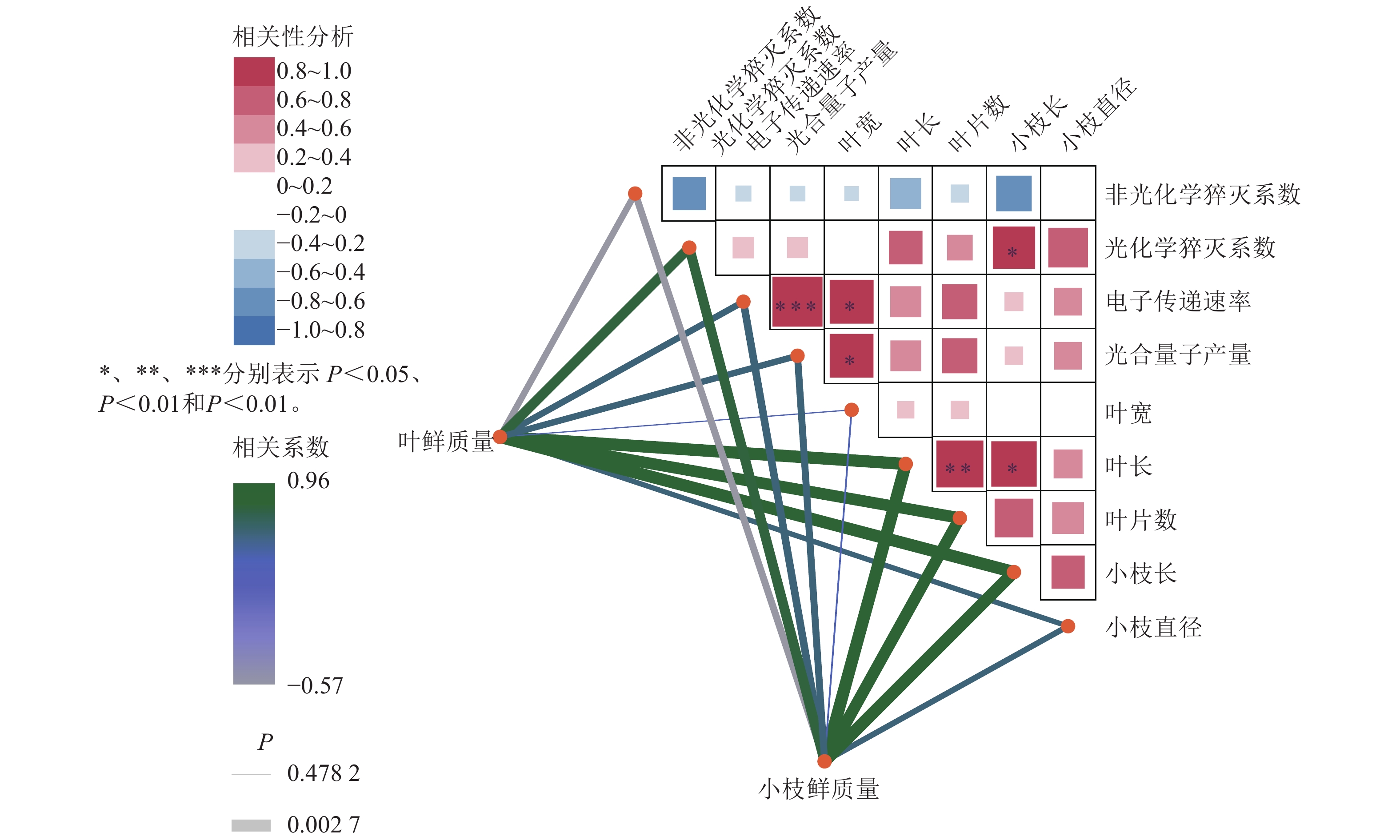
如图6所示:在光合参数之间,电子传递速率与光合量子产量呈极显著正相关(R=0.96,P<0.001);非光化学猝灭系数与光化学猝灭系数、光合量子产量均呈负相关,说明热耗散增强时光化学效率下降,符合植物在逆境下的生理响应。在光合参数与形态指标的关系方面,光合量子产量与叶片宽度、叶片长度呈显著正相关,提示光合效率较高的植株叶片发育较好;非光化学猝灭系数则与多数形态指标呈负相关,可能与胁迫状态下热耗散增强、生长受限有关。在形态指标之间,叶鲜质量与叶长、叶片数、小枝鲜质量、小枝长度与小枝直径等呈显著正相关;叶长与叶片数、小枝长度等呈显著正相关;体现生物量分配的协调性。整体而言,光合效率光合量子产量、光化学猝灭系数与叶片形态正相关,而热耗散非光化学猝灭系数与生长指标负相关,反映植物在光合性能与形态建成上的协同关系,非光化学猝灭系数升高可能指示环境胁迫。
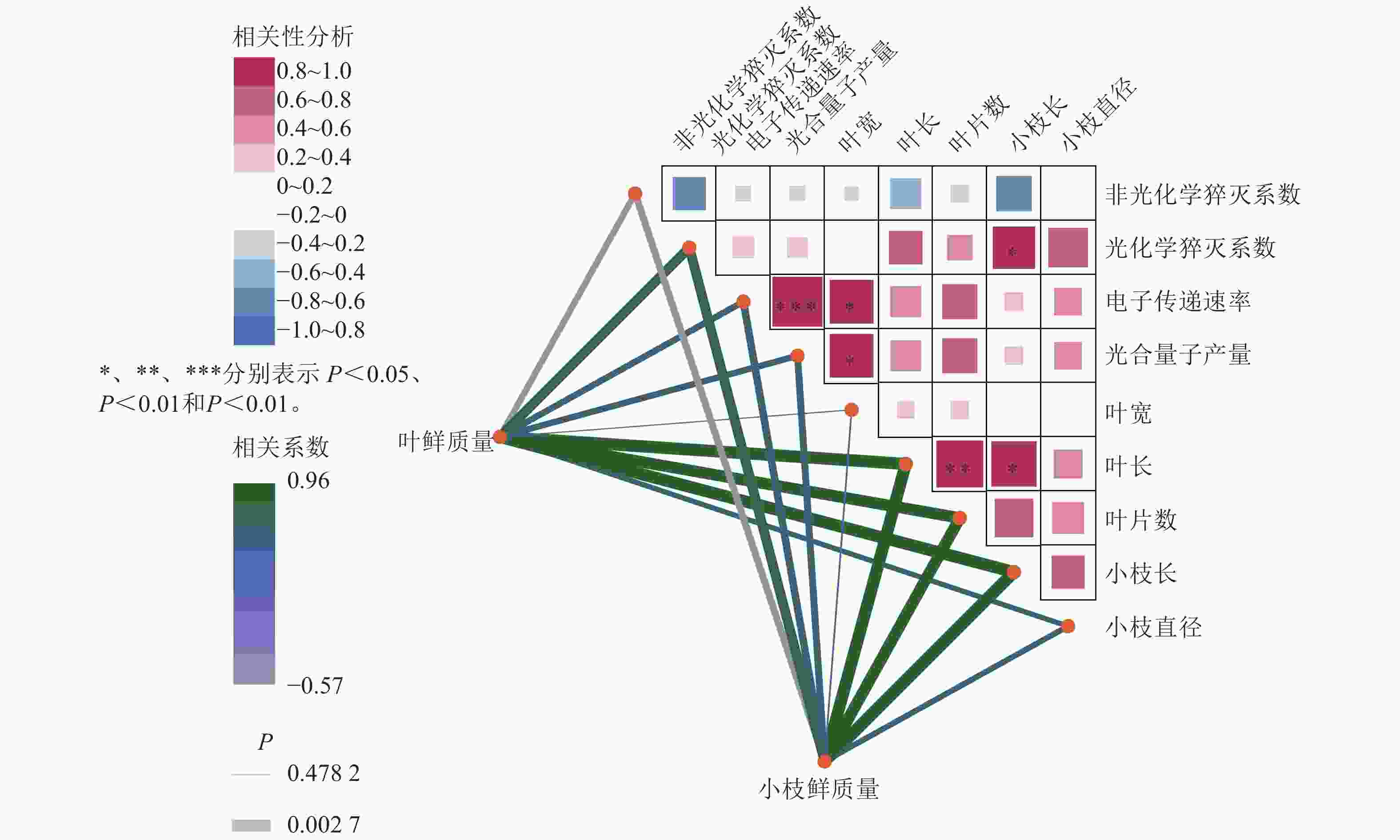
Figure 6. heatmap of correlation networks between the growth parameters of A. beshanzuensis mother trees and grafted trees with chlorophyll fluorescence traits under 268 μmol·m−2·s−1 lightiintensity in August
-
本研究通过比较百山祖冷杉原生母树与嫁接树的生长和叶绿素荧光参数,揭示了嫁接树在光合效率和生长势上的显著优势。这一差异主要源于砧木效应、树龄及生境条件等因素的综合影响。
百山祖冷杉嫁接所用的日本冷杉砧木对其接穗生长可能起到了一定的积极作用。 砧木通常具有更强壮的根系,能更有效地吸收水分和养分[28]。在本研究中,嫁接树各项主要生长指标显著大于原生母树,很可能得益于砧木提供了更充足的资源供给。同时,优良的砧木还能通过调节激素平衡等方式改善接穗的生理状态[29],这为嫁接树拥有更高光化学猝灭系数和电子传递速率,从而将光能更高效地用于光合作用提供了生理学解释。值得注意的是,7月高温天气下,嫁接树的电子传递速率及光化学淬灭系数在>130 μmol m−2 s−1光强条件下等均显著或极显著高于原生母树,而非光化学淬灭系数在>130 μmol m−2 s−1光强条件下显著或极显著低于原生母树,表明嫁接树在高温胁迫下仍能维持较高的光能转化效率。这一差异可能与砧木效应和树龄密切相关。LI等[30]研究表明:耐热砧木可通过维持根系活力、诱导HSP70积累及增强抗氧化酶活性,有效保护PSⅡ免受高温损伤;杏树Armeniaca vulgaris嫁接研究也证实优良砧木能维持高温复合胁迫下更高的光合活性与恢复能力[31]。本研究中嫁接树所采用的日本冷杉砧木可能通过类似机制缓解了高温对光合机构的抑制。此外,树龄是影响植株长势的重要因素之一,高龄树木生理代谢活力普遍下降。王飞等[32]发现沙地云杉Picea mongolica随树龄增长光合性能发生显著下降。百山祖冷杉原生母树树龄约170 a,高龄导致的生理衰退使其对高温胁迫更为敏感。百山祖冷杉嫁接树(42 a)的1年生枝叶生长量和Fv/Fm均大于原生母树,与柑橘‘黄果柑’Citrus ‘Huangguogan’大龄树(280 a)的最大光化学效率和累计光合作用产物低于8 a和30 a植株的研究结果一致[33]。因此,嫁接树凭借砧木保护与树龄优势在夏季高温期表现出更优的光合生理性能,而原生母树受年龄与海拔双重因素制约,光合抑制更为严重。
本研究嫁接树的生长量和叶绿素荧光参数大于原生母树的研究结果,与西滨木槿Hibiscus hamabo根小枝嫁接提高盐胁迫下西洋木槿Hibiscus syriacus生长量、PSⅡ最大光化学量子产量、实际光化学量子产量以及表观电子传递速率的研究结果一致[34];与紫榆Ulmus glabra嫁接树与微繁殖树在高度、胸径和叶片形态方面等研究结果一致,而与其微繁殖榆树的PSⅡ最大光化学量子产量、潜在电子容量和可变初始荧光比较高的结果不同[35],这可能与不同植物物种和不同材料来源有关。
另外,原生母树所处高海拔生境(约1 750 m)是其生长和生理表现较弱的重要原因。高海拔生境导致树体长期面临更强的光辐射等环境胁迫,这与遮阴处理提高了百山祖冷杉光合作用能力的研究结果一致[2]。本研究中原生母树持续较低的最大光化学效率和实际光化学量子产量,以及显著升高的非光化学猝灭系数,是光抑制症状的典型表现[36]。较高的非光化学猝灭系数表明原生母树对比嫁接树启动了强烈的热耗散机制以保护光合机构免受强光破坏,但这不可避免地减少了用于光合碳同化的光能比例,导致光合效率低下[37]。因此,原生母树的表现是其在严峻生境下的一种生存适应,其代价是长势衰退。
本研究在对百山祖冷杉原生母树与嫁接树叶绿素荧光参数的连续6个月监测中发现,其最大光化学量子产量并未随着月份的增加而呈现单一的线性趋势,而是具有先上升、后下降、再上升的变化,夏季7—8月,最大光化学量子产量有明显下降,这可能是因为当气温上升到一定程度, 山顶的气温也不能满足百山祖冷杉对低温条件的需求[38],与百山祖冷杉苗木对高温胁迫的生理响应研究结果一致[10];对高温胁迫下百山祖冷杉的转录组研究中揭示了其相关分子机制[26],提示气候变化下的高温也是其面临的威胁和濒危的重要原因之一。
-
百山祖冷杉嫁接树的1年生小枝长、小枝直径和小枝鲜质量、叶长、叶片数、叶鲜质量等生长指标均显著大于原生母树,表明嫁接树生长量均优于原生母树。通过 4—9月的叶绿素荧光参数测定,嫁接树的光合量子产量、电子传递速率、光化学猝灭系数、最大光化学量子产量均大于原生母树。表明嫁接树的光能捕获效率、电子传递速率、光能转化效率均优于原生母树。嫁接树的非光化学猝灭系数均小于原生母树,表明原生母树的光能热耗散大于嫁接树。同样表明嫁接树的生长状态优于原生母树。百山祖冷杉嫁接树在光合效率和生长势上有显著优势,不仅体现了异砧嫁接在改善接穗生理状态方面的积极作用,也可能得益于其相对优越的较低海拔生境条件。 因此,在制定百山祖冷杉原生母树的精准保护策略时,除考虑嫁接繁殖作为有效的迁地保护手段外,同时应重视通过改善其原生境,如通过适度遮阴和喷雾降低高光强和高温胁迫,从而提升其光合效能、促进其生长和生存能力。
Comparative of growth and chlorophyll fluorescence parameters between original mothers and grafted trees of Abies beshanzuensis
doi: 10.11833/j.issn.2095-0756.20250414
- Received Date: 2025-08-04
- Accepted Date: 2026-04-14
- Rev Recd Date: 2026-04-12
- Available Online: 2026-07-21
- Publish Date: 2026-08-20
-
Key words:
- endangered plant /
- Abies beshanzuensis /
- original mother tree /
- grafted tree /
- growth indices /
- chlorophyll fluorescence parameter
Abstract:
| Citation: | LÜ Hongfei, LI Tao, ZHAO Likang, et al. Comparative of growth and chlorophyll fluorescence parameters between original mothers and grafted trees of Abies beshanzuensis[J]. Journal of Zhejiang A&F University, 2026, 43(4): 784−793 doi: 10.11833/j.issn.2095-0756.20250414

|
 DownLoad:
DownLoad: