






-
土壤真菌作为调控植物生产力的关键因子,是主要的土壤分解者,其菌群多样性受到土壤温度、气候变化及土壤理化性质等的影响。真菌群落组成与土壤异养呼吸强度呈正相关,而土壤温度胁迫导致的真菌群落衰退可能抑制有机质的降解,进而降低土壤-大气系统的碳通量,这种反馈机制深刻影响着陆地生态系统的稳定性,并对全球气候变化具有重要的调节作用[1−3]。土壤真菌的生态营养功能包括腐生营养型、共生营养型和寄生/病原营养型。其中,腐生真菌的α多样性指数与生态系统稳定性呈正相关,而植物病原真菌的丰度则相反,这表明土壤真菌在全球生态系统功能维持中发挥着核心作用,其多样性通过维持植物生长稳态和增强环境胁迫抗性实现[4]。子囊菌门Ascomycota和担子菌门Basidiomycota是土壤真菌的优势类群,它们能够分泌胞外酶降解纤维素、木质素等有机质,促进腐殖化过程,部分类群还能与植物形成外生共生菌根,直接参与养分循环[5−7]。此外,真菌群落还通过跨界互作调控微生物组的装配。JIAO等[8]研究揭示:土壤真菌丰度显著影响细菌群落的构建,随着真菌生物量增加,细菌群落的随机组装减弱,表明真菌-细菌的相互作用在土壤微生物网络形成中具有关键调控作用。
东帕米尔高原平均海拔超过4 000 m,具有低气压、强辐射、昼夜温差大等特征,属于典型的高寒干旱区,年平均气温低于0 ℃,冬季极端低温低于−30 ℃[9−10]。该地区以高原湖盆、宽谷和冰碛地貌为主,分布有戈壁、砾石、草甸、冰川等不同生境。作为典型的高寒生态系统,由于蒸发强烈,土壤普遍存在盐渍化现象,尤其在封闭湖盆区形成大面积盐碱土[11−13]。独特的地理位置和严酷的气候条件使得东帕米尔高原成为研究极端环境下生态系统适应机制的理想区域[14],其土壤真菌群落作为环境变化的敏感指示器,对未来环境监测和生态评估具有重要参考价值。
土壤水分与盐分是调控土壤呼吸及微生物群落结构的关键环境因子[15]。当前水盐梯度对土壤微生物群落的相关研究多集中于温带、亚热带区域,而高寒极端生态系统东帕米尔高原不同生境土壤真菌群落特征及驱动机制仍未见系统报道。鉴于此,本研究选取东帕米尔高原高寒典型生境,基于Illumina NovaSeq高通量测序开展土壤真菌ITS1区测序分析,研究极端高寒环境下土壤真菌适应机制,揭示不同生境土壤真菌多样性、群落结构和功能特征,以期为高原高寒生态系统微生物研究及生态保护修复提供理论依据。
-
本研究选取东帕米尔高原3种典型高寒生境作为研究区域。其一为喀拉库勒湖滨带高寒盐碱地生境(海拔为3 627 m,38°44′86″N,75°04′82″E),该生境受旅游活动干扰与强烈的蒸发积盐作用,呈现典型盐碱化特征,植被覆盖度低[16],其总盐含量和电导率均较高[17]。植物从莎草科Cyperaceae湿生草甸向藜科Chenopodiaceae、蒿属Artemisia等旱生植物演替,表明土壤水体逐渐萎缩,水分条件介于常年积水的湿地与干旱的高寒草甸之间[18]。其二为喀拉苏高寒草甸生境(海拔为3 750 m,38°36′02″N,74°99′08″E),该生境海拔较高,水源以季节性融雪补给为主,土壤排水条件优良且实施围封保育,处于自然恢复阶段,植被长势良好、物种多样性较高,优势植物有兰花棘豆Oxytropis caerulea、岩蔷薇Cistus ladanifer、驼蹄瓣Zygophyllum fabago等;土壤可溶性盐质量分数仅为1.5 mg·g−1,有机质与养分储量充足,土壤持水能力相对偏弱[19]。其三为金草滩河滨带高寒湿地生境(海拔为3 060 m,37°78′32″N,75°23′41″E),该生境为牧草旺盛的高寒湿地,由于常年受地表径流持续补给,土壤水分趋于饱和,盐分胁迫程度较弱,土壤可溶性盐处于中等水平,长期维持湿润稳定的生境条件[18, 20]。
野外采样工作于2023年7月开展,依次采集3种生境15~20 cm土层土壤样品。每个生境设3个重复样地,各样地按S形布点法设5个采样点,采集土壤等量混合,用筛子除去可见的植物根系和石块,得到各生境的3个重复土样。样品置于冰箱运回实验室,在−80 ℃储存用于DNA抽提[21]。
-
利用MoBio Power土壤DNA提取试剂盒12888-50提取土壤DNA,将样本稀释至1 ng·µL−1。以稀释后的基因组DNA为模板,利用真菌通用引物ITS1-1F-F CTTGGTCATTTAGAGGAAGTAA和ITS1-1F-R GCTGCGTTCTTCATCGATGC扩增ITS1序列,PCR采用TransGen AP221-02,TransStart Fastpfu DNA Ploynerase,30 µL反应体系:Phusion High-Fidelity Master Mix(2×) 15 µL,引物 (2 µmol·L−1) 3 µL,基因组(1 ng·µL−1) 10 µL,H2O 2 µL。扩增条件为98 ℃ 1 min,98 ℃ 10 s,50 ℃ 30 s,72 ℃ 30 s,循环30次;72℃ 10 min。PCR产物使用质量分数为2%的琼脂糖凝胶电泳检测扩增目的条带大小,并用Agencourt AMPure XP核酸纯化试剂盒纯化,用质量分数为2%的琼脂糖凝胶电泳检测并等摩尔混合,使用美国Illumina公司的TruSeq® DNA PCR-Free Sample Preparation Kit试剂盒构建文库。构建好的文库经过Qubit和Q-PCR定量,检测合格后,采用Illumina NovaSeq平台进行双末端测序。由陕西佰艾基因信息科技有限公司提供技术支持。
-
基于高质量序列数据,采用Uparse软件在97%的相似性阈值下进行可操作分类单元(OTU)聚类分析,基于OTU聚类结果开展生物信息学分析。采用QIIME分析平台对微生物群落计算α多样性指数,并对真菌群落组成进行解析。使用SPSS Statistics 19软件对α多样性指数进行统计分析,并通过Duncan法进行多重比较。借助Origin 2021软件绘制群落在门和属水平的相对丰度图及聚类热图,以直观展示不同生境的群落结构差异及聚类特征。LEfSe分析可进行2个或多个分组的比较,它强调统计意义和生物相关性,能够在组与组之间寻找统计学差异。采用LEfSe分析筛选组间差异物种,以物种差异贡献分值(LDA)阈值>2、P<0.05为筛选条件,识别各生境的显著差异指示物种。此外,基于FunGuild功能数据库对真菌类群的营养模式和生态功能进行预测和分类注释,同样进行LEfSe分析,设置筛选条件LDA>3,P<0.05。真菌的共现网络图数据分析在R 4.3.1中完成,数据处理使用dplyr,剔除孤立节点后使用igraph软件构建无向加权网络,可视化通过ggraph/ggplot2实现。构建微生物共现网络时,筛选出前50个菌属,随后对丰度数据进行对数转换。采用Spearman秩相关矩阵计算方法,以相关系数绝对值≥0.7为阈值保留边。量化各网络拓扑参数,包括节点数量、边数量、正/负相关计数及平均连接度。
-
由表1可见:ACE与Chao1指数反映的真菌群落丰富度,在高寒湿地和高寒草甸间无显著差异,而高寒盐碱土的真菌群落丰富度显著降低(P<0.05);Shannon指数和Simpson指数表征群落物种多样性,在高寒草甸和高寒盐碱地间无显著差异,而高寒湿地的真菌多样性显著偏低(P<0.05)。3种生境中序列数和OTU数无显著性差异,共有OTU主要为子囊菌门的粪壳菌纲Sordariomycetes和座囊菌纲Dothideomycetes,其次是担子菌门和未培养真菌。
生境类别 序列数 OTU数 Shannon指数 Simpson指数 ACE指数 Chao1指数 高寒盐碱地 166 603±115 93 b 131±56 a 2.79±0.12 a 0.86±0.02 a 134.06±11.91 b 133.77±12.03 b 高寒草甸 185 880±14 394 ab 175±30 a 2.83±0.07 a 0.88±0.01 a 175.61±8.84 a 175.43±8.57 a 高寒湿地 190 886±8 191 a 184±77 a 2.55±0.17 b 0.83±0.01 b 184.93±24.68 a 184.69±24.86 a 说明:同列不同字母表示差异显著(P<0.05)。 Table 1. α diversity index of soil fungi in different habitats
-
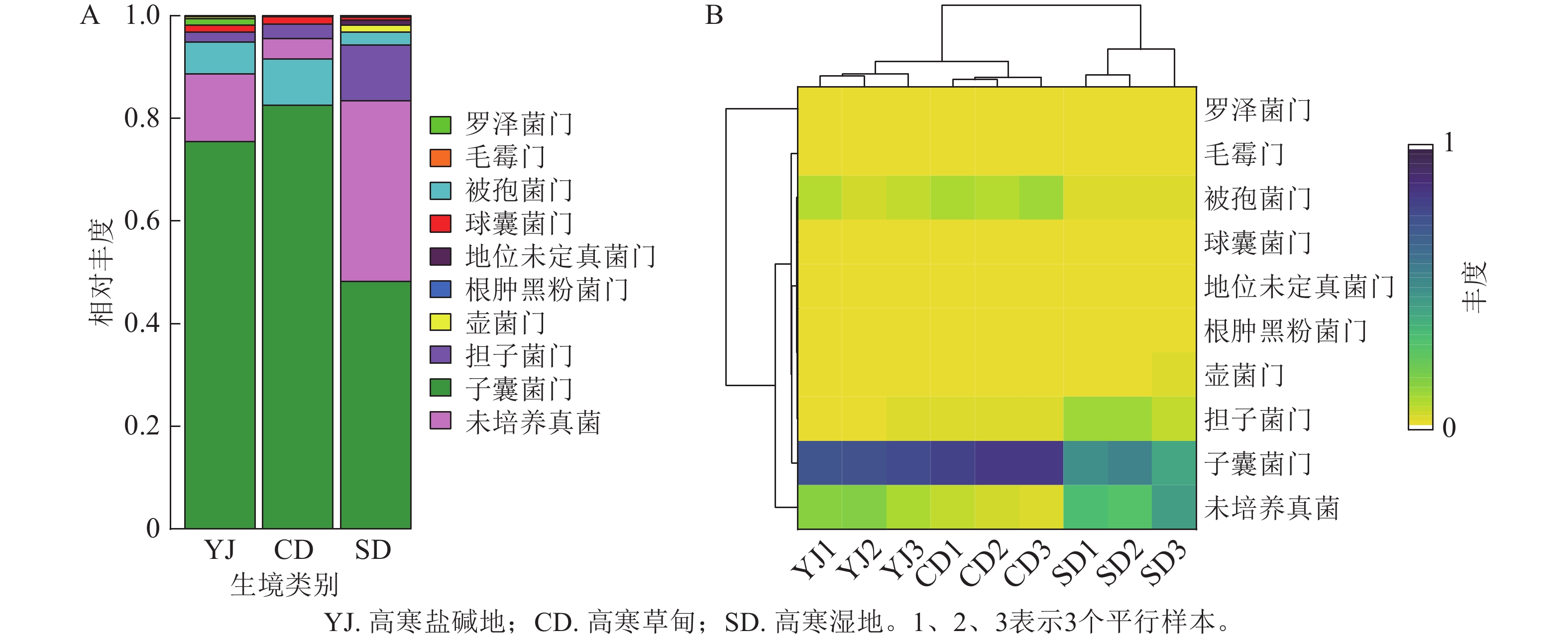
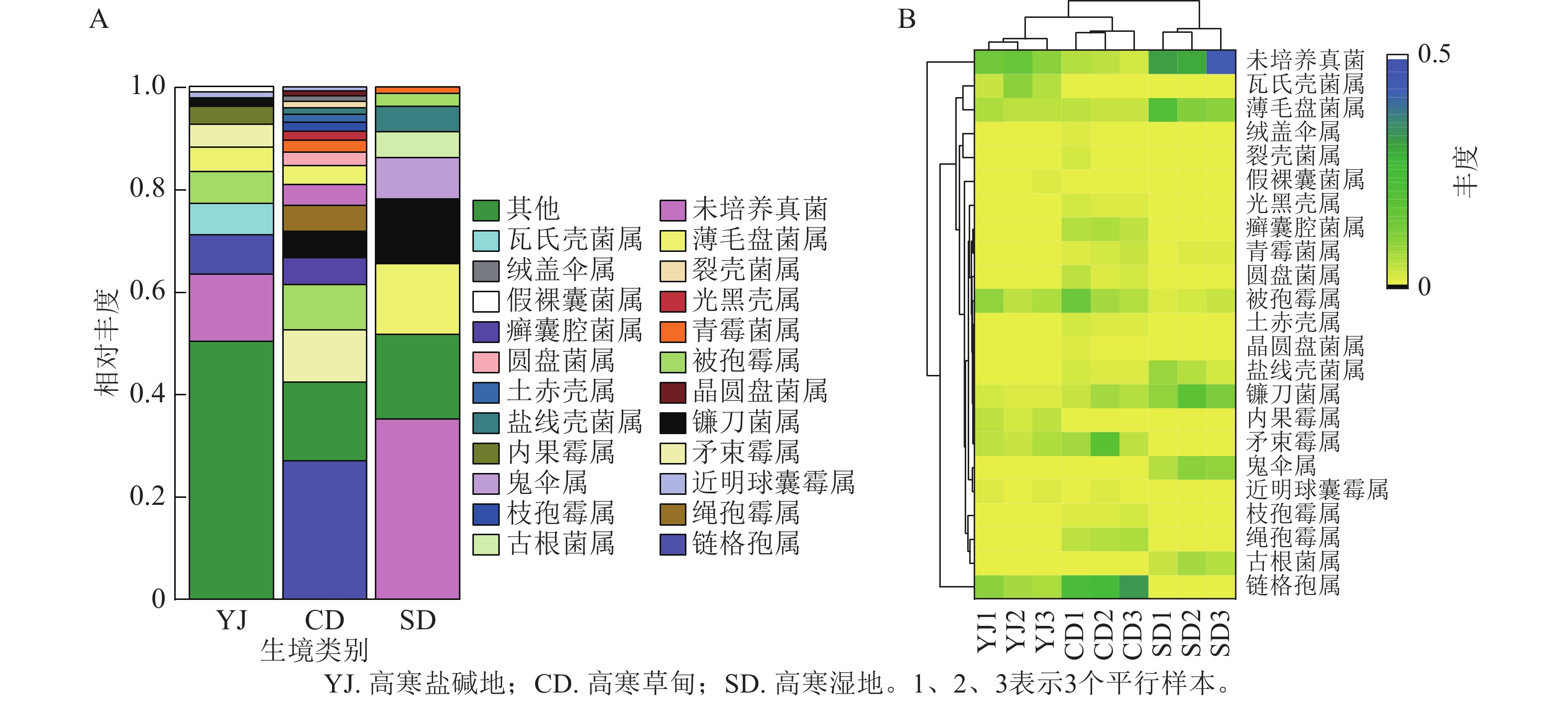
3种生境土壤样品中共检测出真菌10门、30纲、58目、125科、180属、227种。主要类群为子囊菌门,其次是未培养真菌、被孢菌门Mortierellomycota、担子菌门、罗泽菌门Rozellomycota和球囊菌门Glomeromycota等(图1A)。以上菌群在高寒盐碱地土样中占比分别为75.42%、13.20%、6.16%、1.99%、1.32%和1.33%;在高寒草甸土样中占比分别为82.51%、4.00%、8.98%、2.77%、0.11%和1.45%;在高寒湿地土样中占比分别为48.19%、35.30%、2.56%、10.77%、0.18%和0.60%。此外还检测出了占比较少的毛霉门Mucoromycota、根肿黑粉菌门Entorrhizomycota、壶菌门Chytridiomycota。可以看出,子囊菌门在3种生境中均占有绝对优势。聚类分布热图(图1B)显示:高寒盐碱地和高寒草甸真菌群落聚于一支,子囊菌门、未培养真菌、被孢菌门是3种生境中的共有菌群,其中子囊菌门的丰度最高。高寒草甸与高寒湿地的共有菌门是担子菌门。
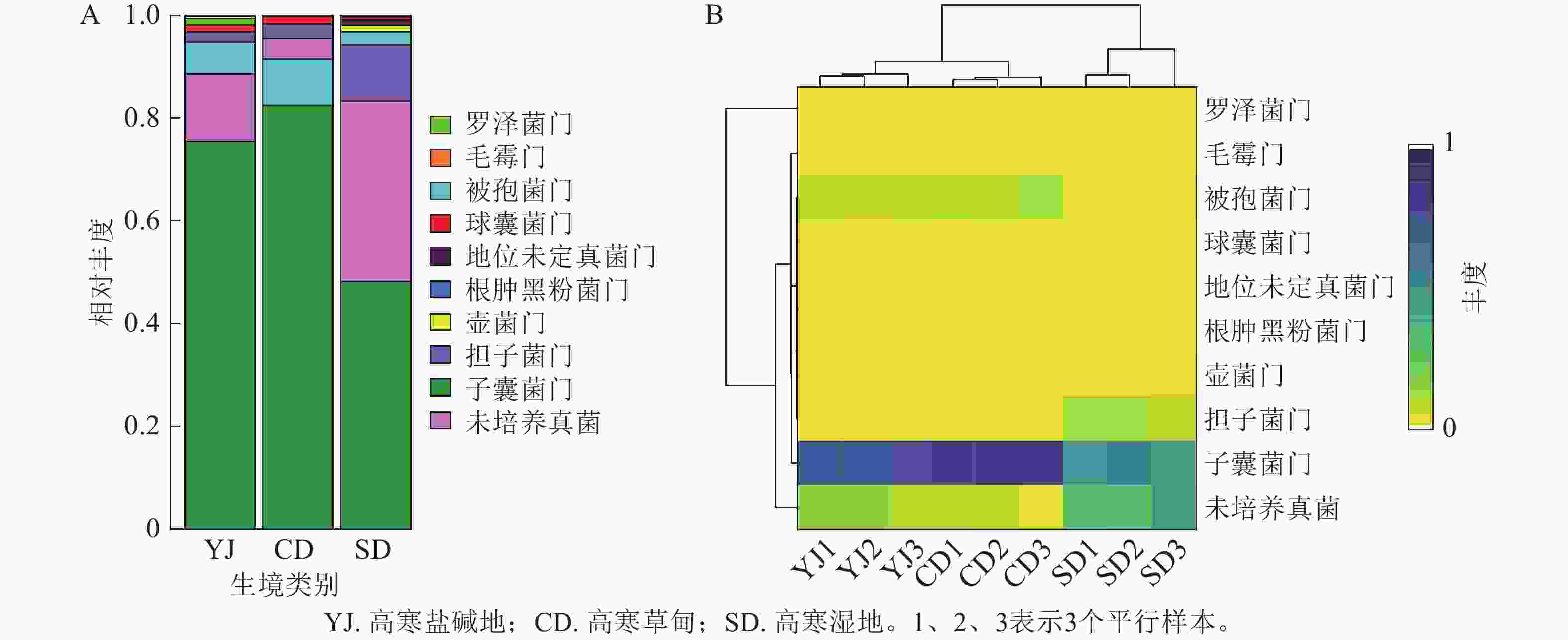
Figure 1. Relative abundance (A) and clustered distribution heat map (B) of fungal community at different taxonomic levels
-
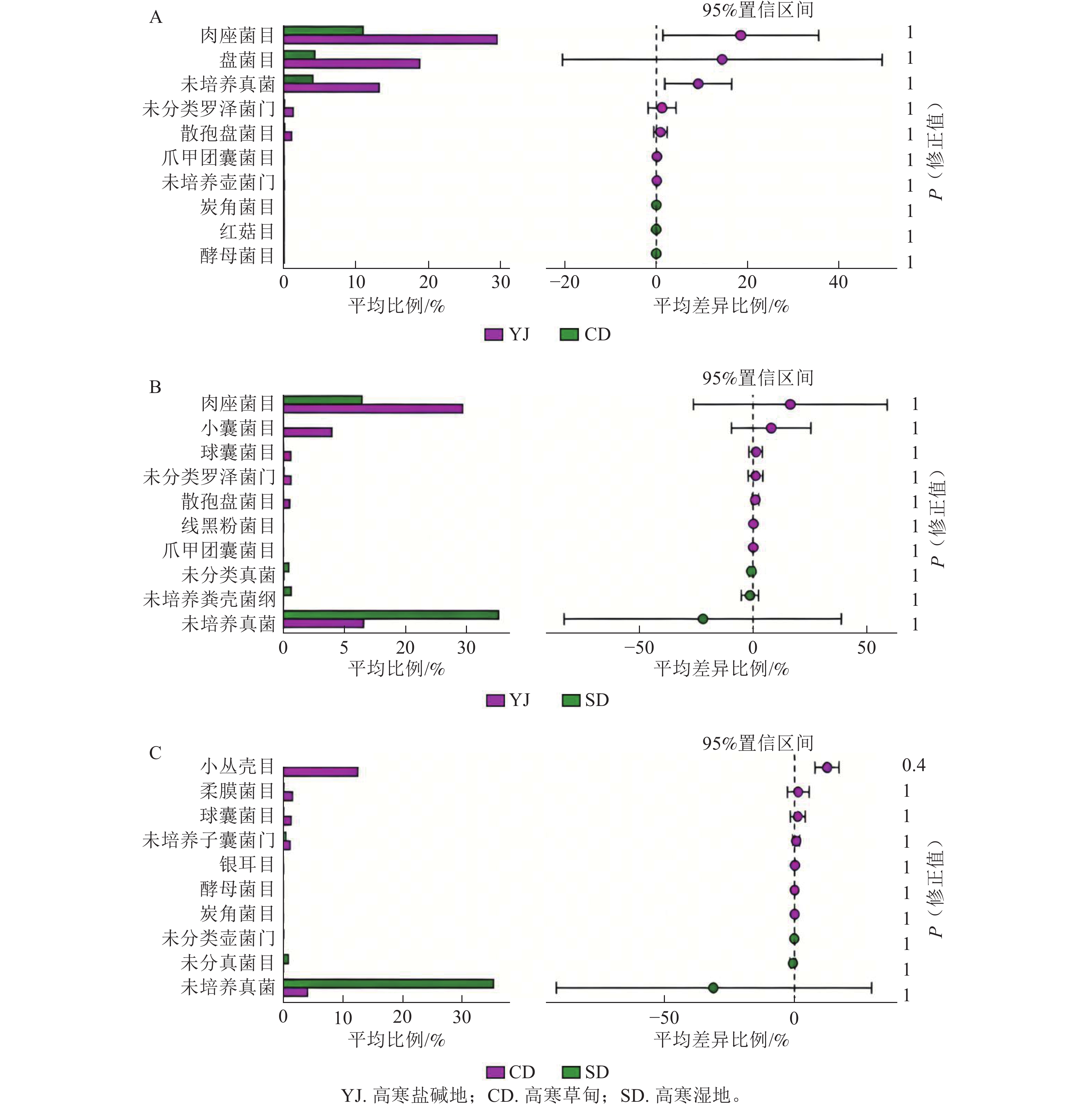
如图2A所示:高寒草甸和高寒盐碱地菌群中肉座菌目Hypocreales、盘菌目Pezizales相对丰度差异显著(P<0.05),均在高寒盐碱地中占优势,其他如散孢盘菌目Thelebolales、爪甲团囊菌目Onygenales、炭角菌目Xylariales、红菇目Russulales、酵母菌目Saccharomycetales等之间差异不显著。高寒盐碱地和高寒湿地菌群中,肉座菌目和小囊菌目Microascales的相对丰度差异显著(P<0.05),均在高寒盐碱地中占优势,其他如球囊菌目Glomerales、线黑粉菌目Filobasidiales、未培养粪壳菌纲Sordariomycetes等之间无显著差异(图2B)。高寒草甸和高寒湿地菌群中小丛壳目Glomerellales的相对丰度差异显著(P<0.05),在高寒草甸中占优势,其他如柔膜菌目Helotiales、银耳目Tremellales等之间差异不显著(图2C)。
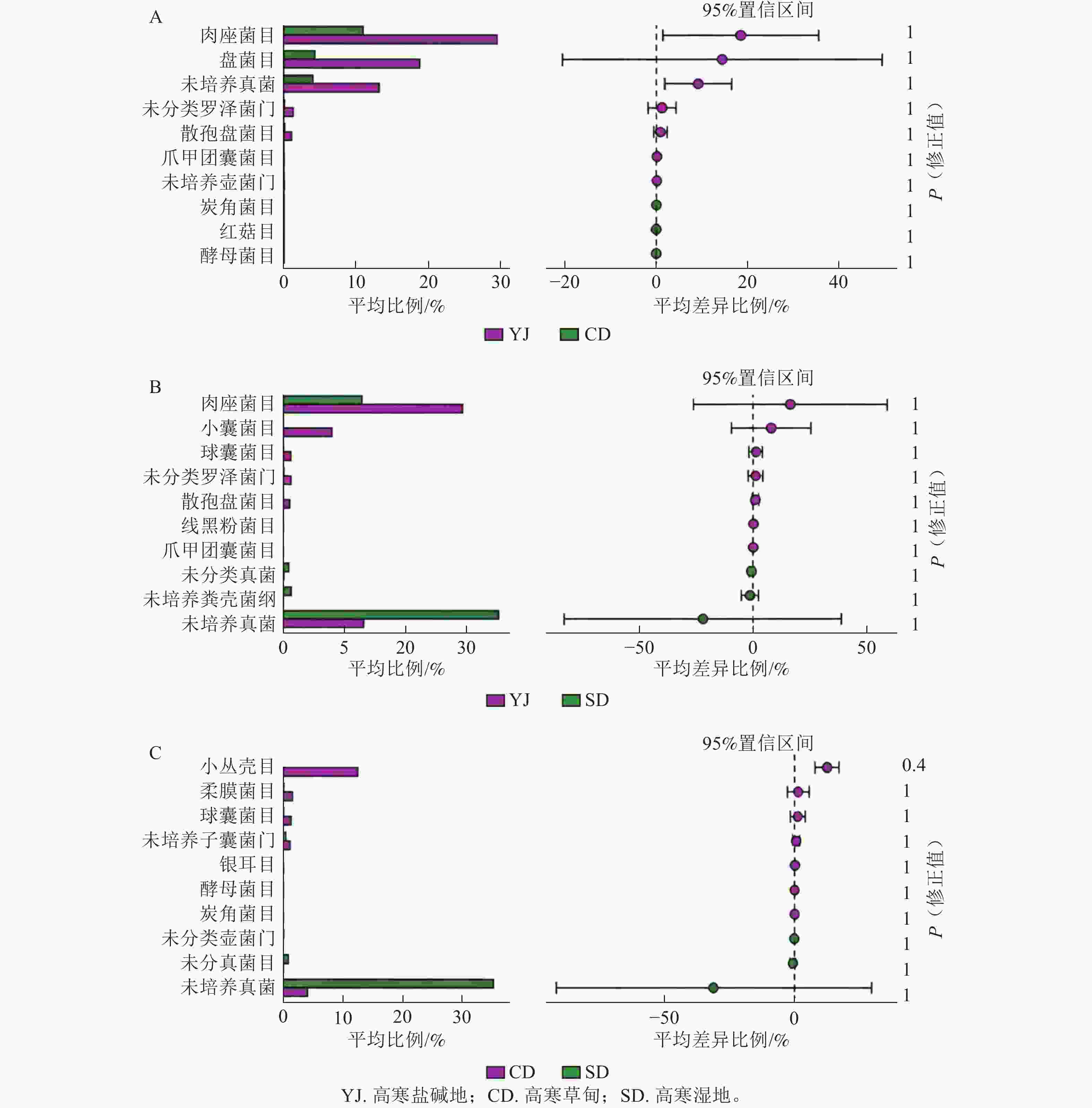
Figure 2. Comparison of relative abundance of soil fungal community at order level in different habitats
-
如图3A所示:3种生境中真菌群落在属水平上差异较大,优势菌属和特有菌属也不尽相同。高寒盐碱地中相对丰度大于1%的属分别为链格孢属Alternaria (7.57%)、近明球囊霉属Claroideoglomus (1.19%)、矛束霉属Doratomyces (4.49%)、内果霉属Enterocarpus (3.50%)、镰刀菌属Fusarium (1.57%)、被孢霉属Mortierella (6.15%)、假裸囊菌属Pseudogymnoascus (1.10%)、薄毛盘菌属Tricharina (4.79%)、瓦氏壳菌属Wallrothiella (6.15%),其中链格孢属和被孢霉属是主要菌群。高寒草甸中相对丰度大于1%的属分别为链格孢属(27.09%)、矛束霉属(10.16%)、镰刀菌属(5.13%)、被孢霉属(8.83%)和薄毛盘菌属(3.66%),其中链格孢属和矛束霉属是主要菌群。高寒湿地中相对丰度大于1%的属分别为镰刀菌属(12.71%)、被孢霉属(2.56%)、薄毛盘菌属(11.82%)和未培养真菌(35.27%),其中除未培养真菌外,镰刀菌属占主导地位。此外,3种生境中还有少量的绒盖伞属Simocybe、裂壳菌属Schizothecium、光黑壳属Preussia、青霉菌属Penicillium、圆盘菌属Orbilia、土赤壳属Ilyonectria、晶圆盘菌属Hyalorbilia、枝孢霉属Cladosporium、古根菌属Archaeorhizomyces等。
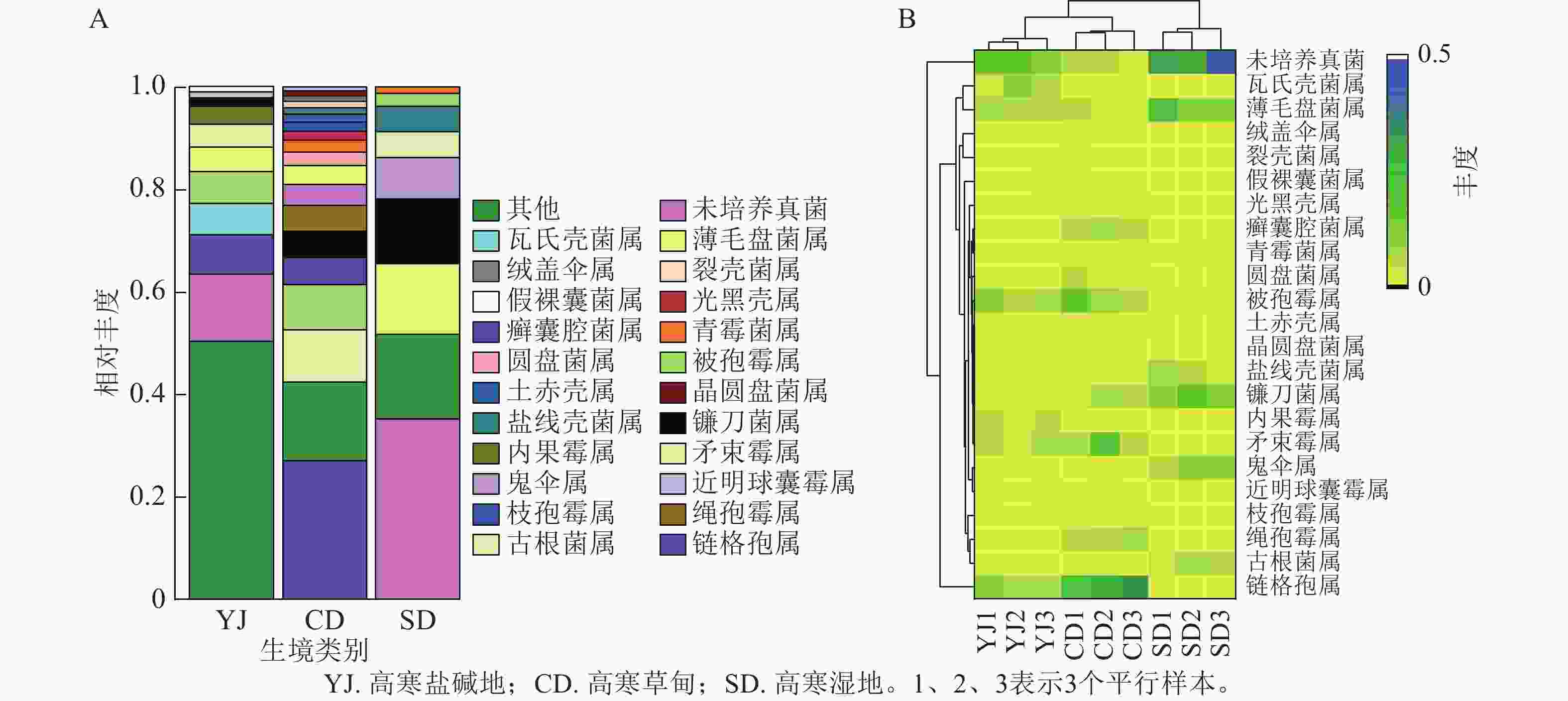
Figure 3. Relative abundance of fungi at the species level (A) and clustered distribution heat map (B)
对属水平相对丰度排名前10的菌群和差异菌群进行关联分析发现:高寒盐碱地和高寒草甸真菌群落聚于一支,它们与高寒湿地菌群存在一定差异(图3B)。薄毛盘菌属、癣囊腔菌属Plectosphaerella、被孢霉属、盐线壳菌属Halobyssothecium、镰刀菌属等22个菌属的相对丰度在3种生境中差异明显。高寒盐碱地中包含瓦氏壳菌属、薄毛盘菌属、被孢霉属、内果霉属、矛束霉属、链格孢属这6个主效菌群。高寒湿地中包含薄毛盘菌属、被孢霉属、镰刀菌属、癣囊腔菌属、矛束霉属、绳孢霉属Chordomyces和镰刀菌属这7个主效菌群。高寒湿地中包含薄毛盘菌属、盐线壳菌属、镰刀菌属、鬼伞属Coprinus和古根菌属Archaeorhizomyces这5个主效菌群。
-
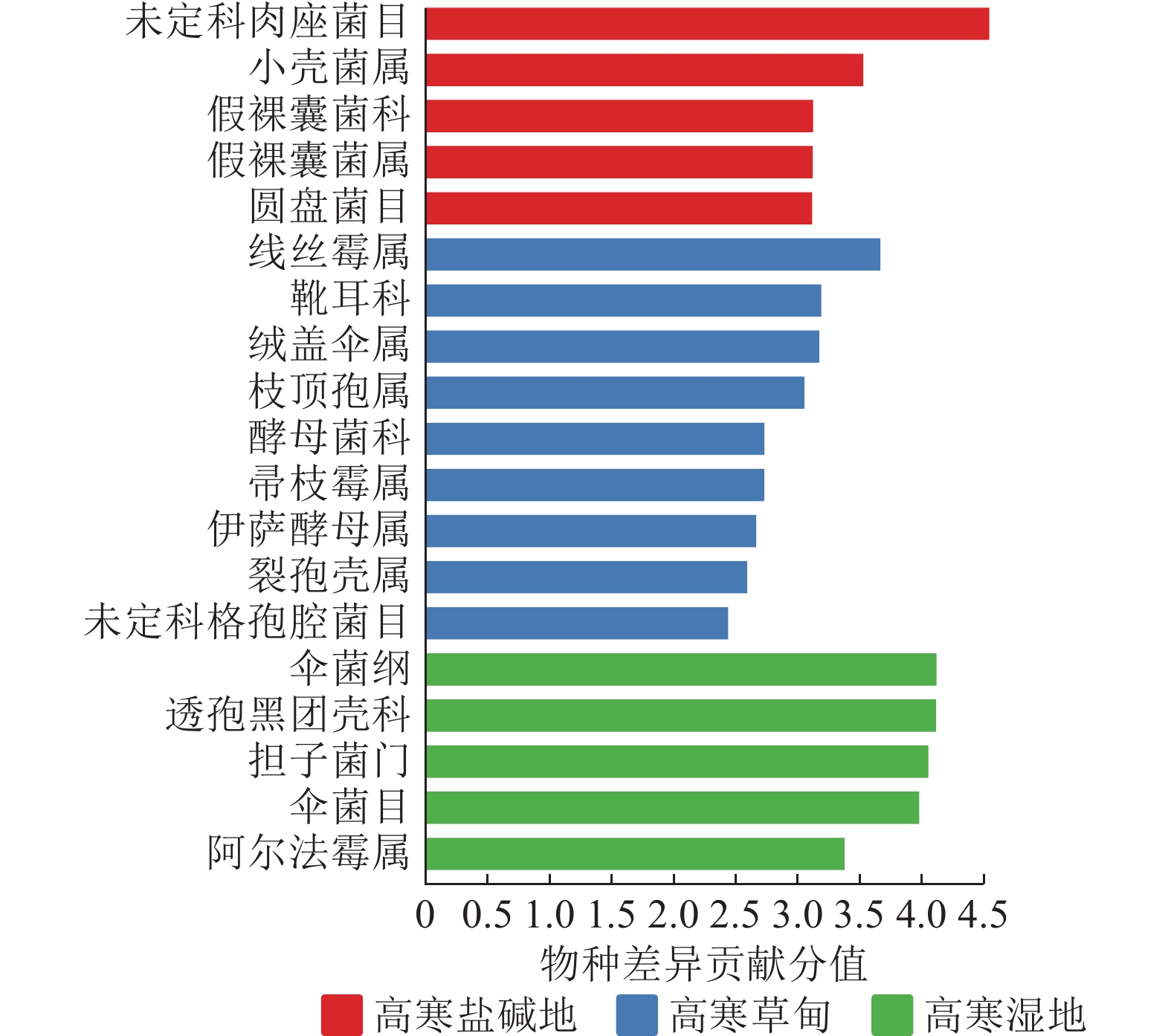
LEfSe物种差异分析(图4)显示:不同生境中菌群相差较大。高寒盐碱地中的主要差异真菌为肉座菌目,其次为小壳菌属Microascus、假裸囊菌科Pseudeurotiaceae、假裸囊菌属Pseudeurotium、圆盘菌目Orbiliales。高寒湿地中的主要差异真菌为伞菌纲Agaricomycetes、透孢黑团壳科Massarinaceae、担子菌门和伞菌目Agaricales,其次为阿尔法霉属Alfaria。高寒草甸未筛选到显著差异真菌,包括线丝霉属Aphanocladium、靴耳科Crepidotaceae、绒盖伞属Simocybe、枝顶孢属Acremonium、酵母菌科Saccharomycetaceae、帚枝霉属Sarocladium、伊萨酵母属Issatchenkia、裂孢壳属Schizothecium及格孢腔菌目Pleosporales等。
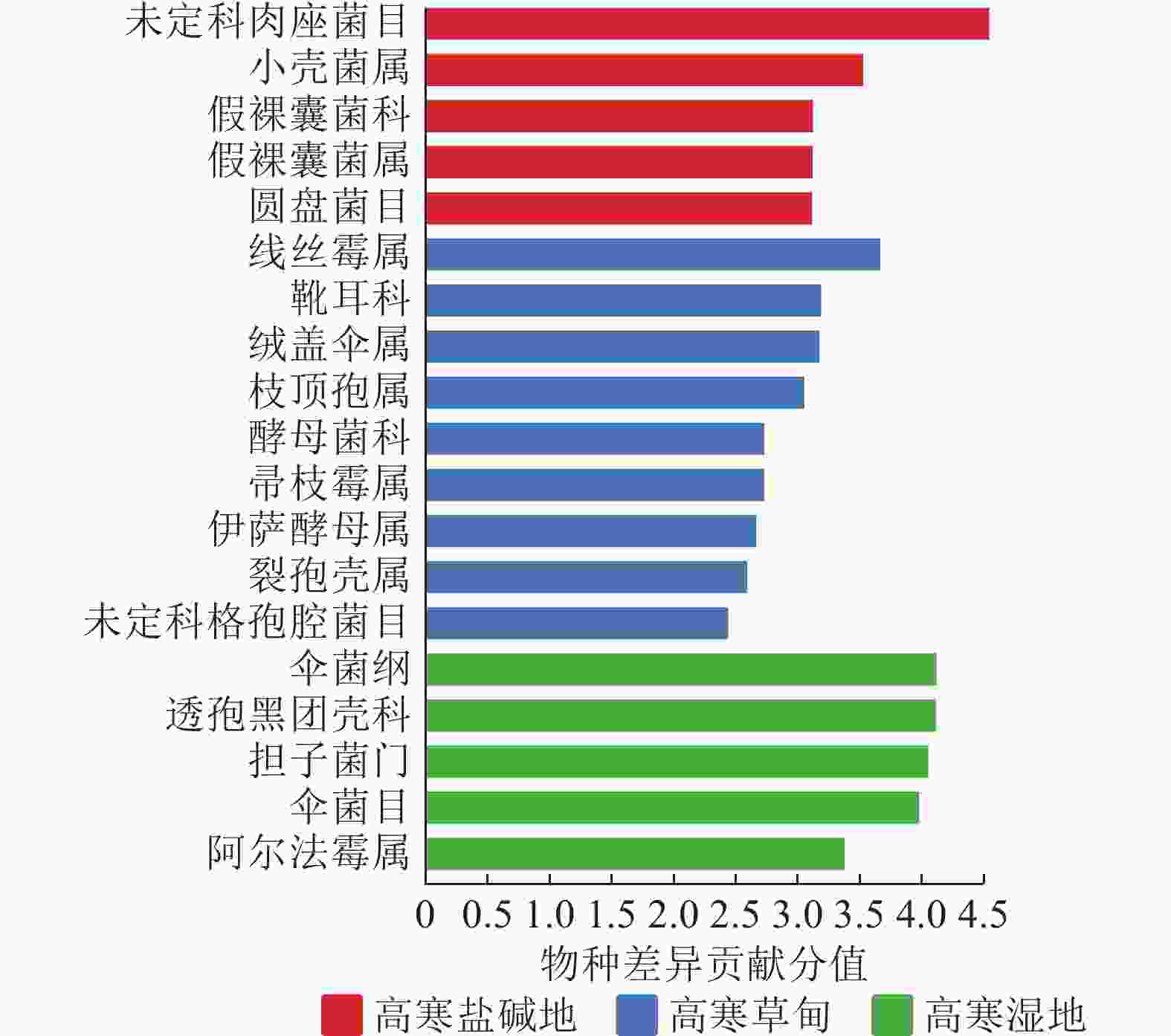
Figure 4. LEfSe analysis of soil fungal community in different habitats
-
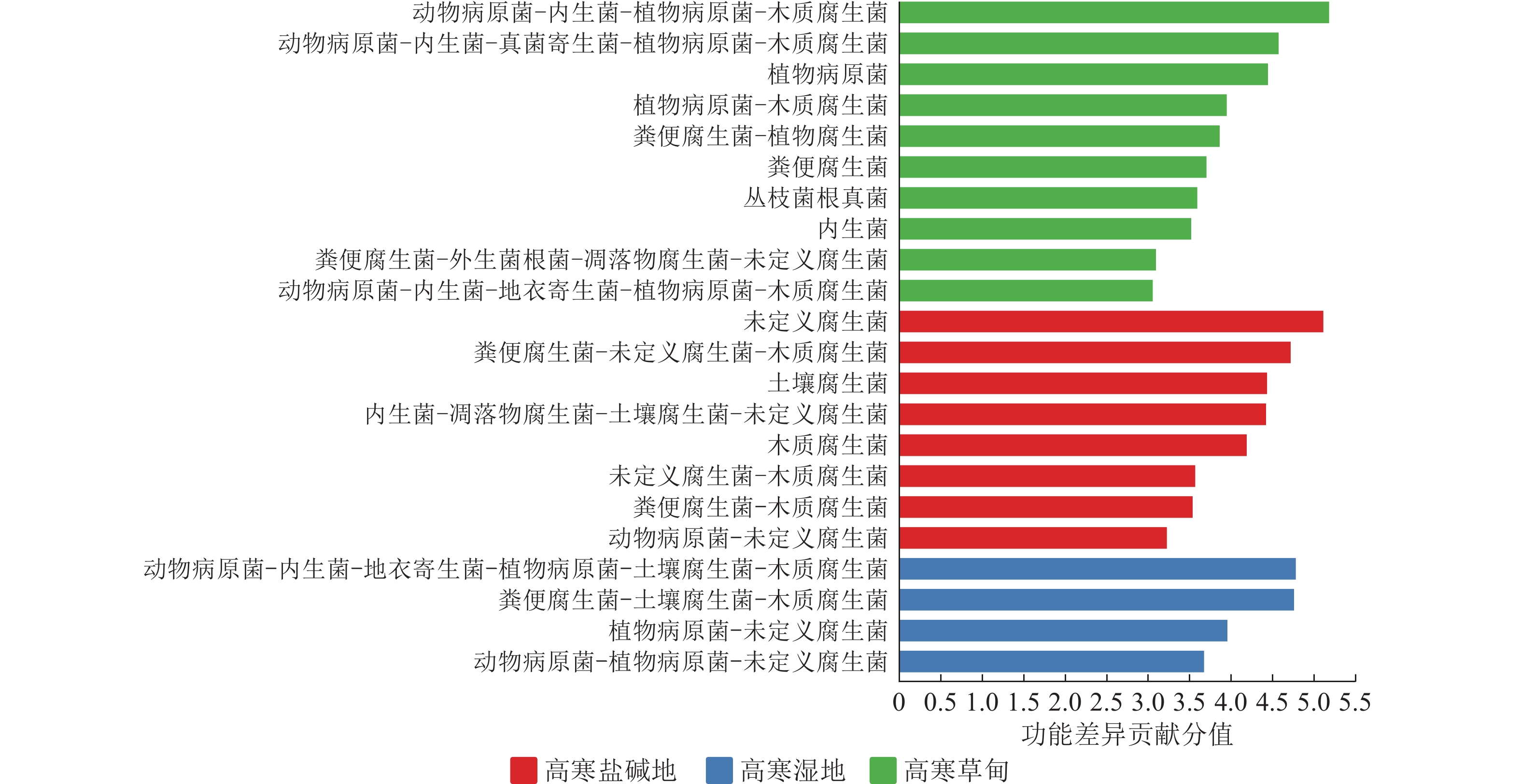
基于FunGuild数据库对所有样本真菌进行物种分类,并注释其生态功能。图5表明:3种生境中菌群营养型差异较大。在高寒盐碱地中,未定义腐生菌、粪便腐生菌-未定义腐生菌-木质腐生菌、土壤腐生菌营养型是最主要的。在高寒草甸中,动物病原菌-内生菌-植物病原菌-木质腐生菌、动物病原菌-内生菌-真菌寄生菌-植物病原菌-木质腐生菌、植物病原菌营养型发挥主要作用。在高寒湿地中,动物病原菌-内生菌-地衣寄生菌-植物病原菌-土壤腐生菌-木质腐生菌、粪便腐生菌-土壤腐生菌-木质腐生菌是重要营养型。综上,尽管腐生功能在3种生境中均普遍存在,但其具体组合与核心营养型存在明显差异。高寒盐碱地以基础腐生型为主,高寒草甸以病原-内生复合型为特征,而高寒湿地则表现为高度多样化的复合营养型。
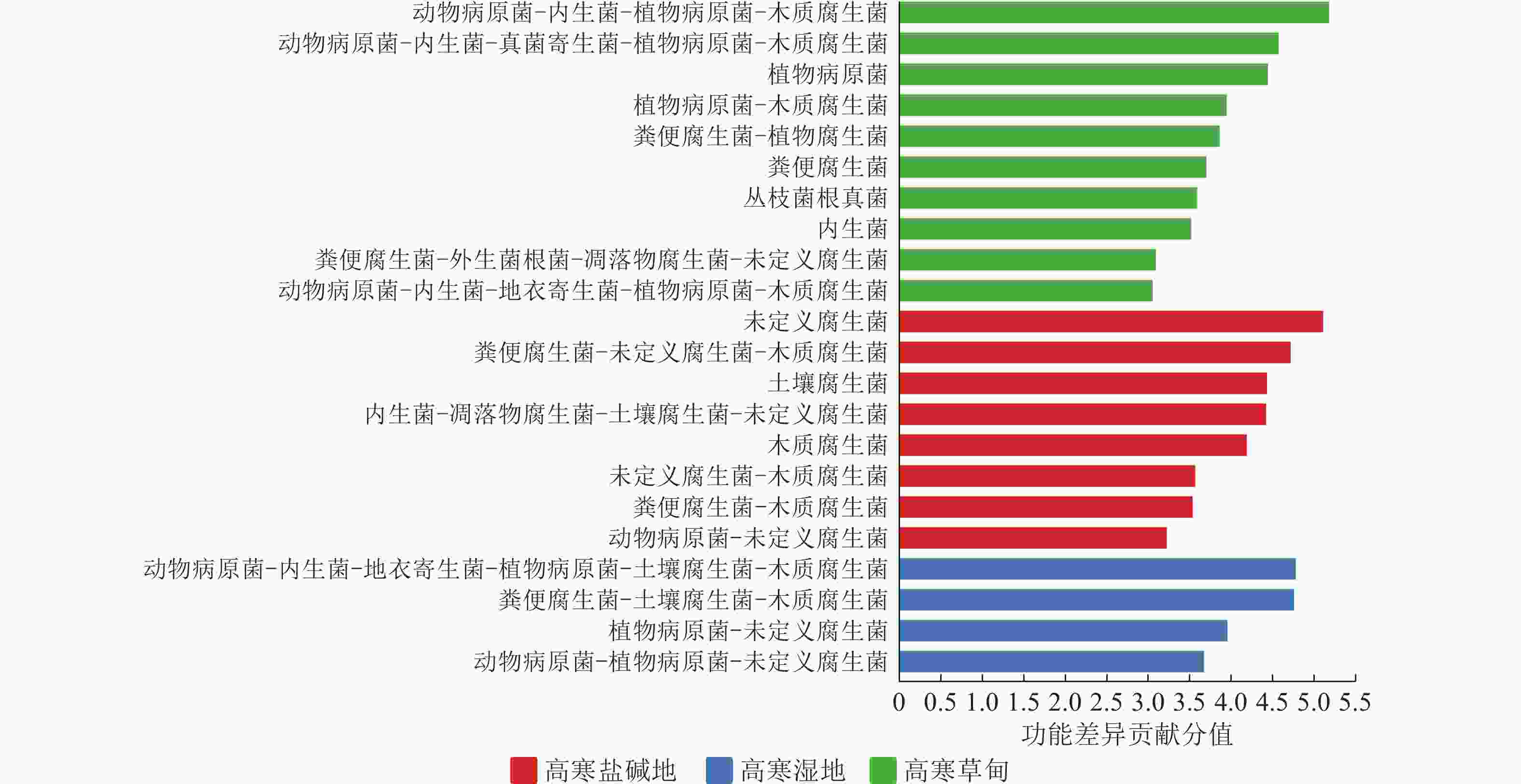
Figure 5. Differences in fungal trophic modes across various habitats based on FunGuild
-
共现网络中(图6),高寒盐碱地有427条边与真菌丰度呈正相关,161条边与真菌丰度呈负相关,平均度数为23.52;高寒草甸中501条边与真菌丰度呈正相关,124条边与真菌丰度呈负相关,平均度数为25.00;高寒湿地中567条边与真菌丰度呈正相关,225条边与真菌丰度呈负相关,平均度数为31.68。所有样本以子囊菌门为绝对主导,其次是担子菌门。此外,还有少量的毛霉菌门Mucoromycota、球囊菌门和罗泽菌门成员,但壶菌门在高寒草甸中不存在。所有样本共有优势菌属分别为子囊菌门的薄毛盘菌属和毛霉菌门的被孢霉属。然而,不同样本的核心菌群组成存在明显差异。高寒盐碱地的核心菌群以子囊菌门的链格孢属、瓦氏壳菌属、矛束霉属为主;高寒草甸的核心菌群以子囊菌门的链格孢属、枝孢霉属、癣囊腔菌属、绳孢霉属、矛束霉属为主;高寒湿地的核心菌群以古根菌属、盐线壳菌属、鬼伞属为主。

Figure 6. Co-occurrence network of fungi community
-
土壤微生物多样性及其互作网络是生态系统的关键调控因子,其中土壤真菌作为陆地生态系统的重要功能组分,对维持生态系统健康、生产力及生物地球化学循环具有不可替代的作用[22]。本研究中高寒湿地与高寒草甸的真菌群落丰度差异不显著,均高于高寒盐碱地。但在群落多样性方面,高寒盐碱地与高寒草甸差异不显著,均高于高寒湿地。这与宋珊珊等[23]关于草甸草原根际土壤真菌多样性与植物地上生物量呈正相关、陈焘等[24]关于高寒草甸土壤真菌丰富度显著高于草甸草原和荒漠草原的研究结果相似。本研究中3种生境间真菌群落的α多样性差异较小,这与张甜等[21]关于高寒草甸和高寒湿地的细菌α多样性显著高于高寒盐碱地的研究结果形成鲜明对比。这一现象与汉鑫等[25]关于根际/非根际土壤真菌α多样性无显著差异,但与GONG等[26]关于土壤生态恢复过程中真菌群落结构较细菌更稳定的结果相吻合,进一步支持了LI等[27]关于真菌互作网络拓扑稳定性高于细菌的研究论断。
子囊菌门是3种生境土壤真菌群落的绝对优势类群,与多项研究吻合[2, 28]。作为真菌界多样性最丰富的类群,子囊菌门主营腐生生活方式,是土壤有机质分解的关键菌群,通过高效降解木质素和纤维素等难分解有机物,在土壤养分循环过程中发挥着核心生态功能[29−30]。被孢菌门是高寒盐碱地和高寒草甸的优势菌门,显著高于高寒湿地,该结果与张阳灿等[31]对高寒荒漠生态系统的研究结果一致,也与徐欢[32]对高寒重度退化草原被孢菌门数量增多的研究结果类似。结合3种生境环境特征推测,真菌群落结构差异与生境类型介导的土壤水分梯度相关。高寒盐碱地和高寒草甸具有更低的土壤含水量和更好的透气性,从而形成了更适宜好氧性被孢菌门生长的微环境。被孢菌门作为重要的土壤有益真菌,兼具固氮解磷效能,还可促植株生长、提升抗逆能力,多重生态功能使其在特定生境中具备更强竞争优势[33]。此外,壶菌门在高寒草甸中缺失,推断是草甸土壤含水量低于湿地和盐碱地,而该菌群多依赖湿润环境,草甸环境可能超出壶菌门的生存阈值[34]。这与李文娟等[35]在滨海湿地盐地碱蓬Suaeda salsa根际土壤的研究结果相似。而担子菌门在高寒湿地中丰度最高,这与纳帕海高原湿地和青藏高原高寒湿地的微生物群落研究结果相印证[36−37]。其形成机制可能与高寒湿地旺盛的牧草生长提供了丰富的木质素和纤维素基质有关,因担子菌门中的优势类群白腐真菌对这类难降解有机底物具有特异性,从而在资源富集区域形成生态位优势[38−39]。
不同生境真菌群落结构和共现网络表明:高寒盐碱地以链格孢属为绝对优势菌属,以被孢霉属、薄毛盘菌属和矛束霉属为次优势菌属;高寒草甸则以链格孢属和矛束霉属为优势类群,以被孢霉属、枝孢霉属、癣囊腔菌属和镰刀菌属为次优势菌属;高寒湿地中以镰刀菌属与薄毛盘菌属为优势类群,以古根菌属、盐线壳菌属和鬼伞属为次优势菌属。被孢霉属隶属于毛霉亚门Mucoromycota被孢霉目Mortierellales,其部分物种可产生厚垣孢子等特殊结构以适应极端环境[40]。而链格孢属、矛束霉属、薄毛盘菌属和镰刀菌属等均归属于子囊菌门,这些真菌类群可促进养分循环和微生物群落演替[38]。部分薄毛盘菌属可与植物形成菌根共生关系,其生态分布具有显著的生境特异性,尤其在高寒湿地、高山草甸及青藏高原地区的腐殖质层或贫瘠土壤中呈现优势分布[41]。链格孢属和瓦氏壳菌属大多具有耐盐碱和干旱特性,能在碱性环境中维持细胞代谢平衡,但链格孢属和镰刀菌属作为典型的兼性寄生菌,既可营腐生生活,也能作为植物病原体存在[42−43]。枝孢霉属与癣囊腔菌属、薄毛盘菌属有相似的特征,可与优势植物形成共生关系[44−45];古根菌属含有形成植物根系组合的隐蔽丝状物种,其群落丰度与腐生真菌及外生菌根真菌类群存在显著的共变化模式;盐线壳菌属为水生菌,适于湿地高含水量环境;鬼伞属为担子菌门成员,是一种典型的腐生菌,可高效分解湿地丰富的植物残体与有机质,提升湿地碳汇能力[46−48]。
3种生境的共现网络结构虽都密集,但核心菌属和相互作用仍呈现差异化。从平均度数看,高寒湿地>高寒草甸>高寒盐碱地,这表明湿地生境中真菌物种间的相互作用更复杂,可能源于该区域土壤较高的含水量和有机质含量,及相对稳定的温湿度条件,促使物种间形成更密集的协作或竞争关系。从高寒盐碱地→高寒草甸→高寒湿地,最核心菌属由链格孢属+被孢霉属逐渐转向镰刀菌属+薄毛盘菌属+鬼伞属,说明真菌功能从抗逆性腐生营养型菌群向腐生+共生营养型菌群转变,以适配植物残体分解与植物-真菌互作的需求。
现有研究证实:草甸土与盐碱土的真菌丰度及多样性接近,这符合退化土壤微生物群落由“细菌型”向“真菌型”转换的理论[21, 49]。本研究中3种生境放牧干扰强度存在差异,高寒盐碱地因营养贫瘠、植被覆盖度低,放牧活动少,同时受喀拉库勒湖旅游区人为干扰导致土壤结构退化。相较之下,高寒草甸与高寒湿地生境牧草产量较高,适度放牧活动输入了丰富的碳源和腐殖质,为细菌生长创造了有利条件。在营养贫瘠的环境中,细菌群落因营养竞争敏感性更易发生衰退,而真菌网络结构赋予其更强的抗干扰韧性,其通过产生大量孢子和菌丝片段实现快速扩散和定殖,或通过分泌胞外酶降解木质素/纤维素等有机物,即使局部菌丝受损仍能维持整体功能稳定性,从而在不同生境中维持较高的生态竞争力[50−52]。这些机制共同解释了高寒盐碱地中细菌α多样性显著低于真菌的现象,也印证了真菌群落在不同生境中表现出的显著生态稳定性。需要指出的是,本研究3个样点的土壤水分与盐分梯度主要基于地理位置、植被特征以及已发表的文献推断得出,未能对采样当季的土壤理化性质(如含水量、电导率)进行实测。未来的研究将包含对土壤理化性质的同步监测,以获取更直接的环境数据。
-
东帕米尔高原高寒盐碱地、高寒草甸和高寒湿地3种生境土壤的真菌群落分布具有一定的稳定性,但也存在一定差异。高寒湿地与高寒草甸的真菌群落丰度均高于高寒盐碱地,而高寒盐碱地与高寒草甸的真菌多样性均高于高寒湿地。土壤真菌群落组成在门和属水平均有差异,核心类群是子囊菌门,共有类群是薄毛盘菌属。此外,高寒盐碱地和高寒草甸的优势菌群还有被孢菌门、链格孢属和被孢霉属,高寒湿地中是担子菌门和镰刀菌属。各生境中真菌生态营养型以腐生型为主,高寒盐碱地主要为基础腐生型,高寒草甸为病原-内生复合型,高寒湿地为高度多样化的复合营养型。综上,高寒区不同生境真菌群落共现网络结构与核心菌群特征,是真菌对环境适应性与生态功能需求共同塑造的结果。
Community structure of soil fungi in 3 typical alpine habitats of the Eastern Pamir Plateau
doi: 10.11833/j.issn.2095-0756.20250527
- Received Date: 2025-10-05
- Accepted Date: 2026-05-26
- Rev Recd Date: 2026-05-20
-
Key words:
- Eastern Pamir Plateau /
- high-throughput sequencing /
- soil /
- fungal community /
- ecological function
Abstract:
| Citation: | ZHANG Tian, WANG Jilian, LI Mingyuan, et al. Community structure of soil fungi in 3 typical alpine habitats of the Eastern Pamir Plateau[J]. Journal of Zhejiang A&F University, 2026, 43(X): 1−12 doi: 10.11833/j.issn.2095-0756.20250527

|

 DownLoad:
DownLoad: