




-
全球城市化率持续上升,噪音污染和土地利用变化使得生物的栖息地逐渐缩小,如何有效保护生物多样性成为环境保护的热点话题之一[1]。城市绿地为各种生物提供庇护所,能让城市产生巨大的生态服务效应[2]。鸟类作为城市中最常见的物种之一,会随着城市化发展程度而变化[3]。研究表明:城市化程度与鸟类数量呈负相关[4],但不同城市绿地间会存在差异[5],当绿地能够提供多样且高质量的生境时,对城市化敏感的鸟类数量会有所增加[6]。此外,鸟类迁徙行为使城市中鸟类群落组成在不同时间发生变化[7]。目前大部分研究只专注于单一绿地鸟类群落的变化,但鸟类群落往往受到不同绿地生境特征以及周边环境的复合影响。因此,本研究选取昆明主城区公园绿地、街旁绿地和社区绿地,分析3种游憩性绿地繁殖季节与非繁殖季节不同居留型鸟类多样性差异,探究绿地环境因子对鸟类的影响,以期为提高城市鸟类绿地利用率和多样性提供参考。
-
昆明市位于中国西南云贵高原中部(102°10′~103°40′N,24°23′~26°22′E),建成区总面积446.13 km2。北部山地环抱,南部面临滇池,总体地形北高南低,自北向南成阶梯状逐级下降。昆明属低纬度高原山地季风气候,冬无严寒,夏无酷暑,四季如春,是众多鸟类的越冬地和迁徙期的中转站,鸟类资源丰富。本研究按照城市绿地系统分类,选择昆明市建城区44个绿地作为调查样地,其中公园绿地18个,街旁绿地18个,社区绿地8个(表1)。
表 1 调查样地基本信息
Table 1. Basic information of survey site
序号 绿地名称 绿地类型 面积/hm2 样线数量/条 序号 绿地名称 绿地类型 面积/hm2 样线数量/条 1 郊野公园 公园绿地 82.90 7 23 低碳中心 街旁绿地 1.32 1 2 黑龙潭公园 公园绿地 86.40 6 24 金汁河桥 街旁绿地 1.87 1 3 金殿森林公园 公园绿地 71.30 6 25 西京寺 街旁绿地 2.91 1 4 洛龙公园 公园绿地 40.00 10 26 葵花公社 街旁绿地 2.16 1 5 渔浦寒泉森林生态湿地公园 公园绿地 41.20 10 27 白邑寺 街旁绿地 2.43 1 6 隅山公园 公园绿地 11.80 6 28 金树春带状公园 街旁绿地 1.45 1 7 瀑布公园 公园绿地 32.50 8 29 董家湾 街旁绿地 0.93 1 8 月牙潭公园 公园绿地 16.40 5 30 杰霖园 街旁绿地 0.79 1 9 昙华寺公园 公园绿地 10.20 5 31 市级行政中心 街旁绿地 1.74 1 10 官渡森林公园 公园绿地 24.10 6 32 永昌湿地 街旁绿地 1.51 1 11 如意公园 公园绿地 27.00 8 33 龙马路 街旁绿地 2.60 1 12 龙江公园 公园绿地 5.37 5 34 华夏天景湾 街旁绿地 1.53 1 13 翠湖公园 公园绿地 24.20 5 35 前卫西路 街旁绿地 1.00 1 14 海洪湿地公园 公园绿地 20.60 5 36 蓝光欢乐城 街旁绿地 1.74 1 15 西亮塘湿地公园 公园绿地 48.40 8 37 博众花园 社区绿地 0.53 1 16 斗南湿地公园 公园绿地 52.70 9 38 水晶骊城 社区绿地 0.91 1 17 捞鱼河公园 公园绿地 55.20 7 39 东旭俊城 社区绿地 1.39 1 18 西华园 公园绿地 6.79 3 40 关上法治 社区绿地 1.27 1 19 银杏公园 街旁绿地 2.48 1 41 金泰国际 社区绿地 1.60 1 20 郎溪街 街旁绿地 2.44 1 42 五华广场 社区绿地 1.27 1 21 鼓楼公园 街旁绿地 0.95 1 43 五星彩园 社区绿地 0.58 1 22 洛羊街道办事处 街旁绿地 3.10 1 44 碧鸡名城 社区绿地 1.39 1 -
鸟类调查使用样线法。根据选取绿地的可游览面积和绿地形状设置调查样线,每条样线长度为100 m,样线之间首尾间隔100 m以上,与绿地边界有50 m的缓冲距离。44块绿地共145条样线。于2021年7月至2022年8月每月调查1次,每次调查时选取晴朗无风的天气,调查时间为7:00—11:00和15:00—17:00的2个时间段。调查期间,使用BOSMA野狼望远镜,以2 km·h−1的速度沿固定线路行进,记录样线两侧25 m范围内观察到的鸟类种类、数量、行为、停留地点以及所处植被生境。鸟类识别、分类依据《中国鸟类野外手册》[8]和《中国鸟类分类与分布名录》[9]。
在每条鸟类调查样线的一侧设置1个20 m×20 m的大样方进行乔木调查,在大样方内4个角和中心设置5 m×5 m的中样方进行灌木调查,在中样方内选取1 m×1 m的小样方进行草本调查。
-
选取城市绿地内部和外部环境因子探究不同绿地鸟类群落存在差异的原因。内部环境因子包括绿地归一化植被指数(NDVI)和归一化建筑指数(NDBI)、面积、周长、形状、水体面积以及乔木、灌木、草本的丰富度和多样性指数。外部环境因子包括按绿地边界50、100、200、400 m缓冲区内部NDVI和NDBI指数,以及绿地距离市中心、山体和滇池最近的距离。
使用ArcGIS计算城市绿地面积和形状、水体面积、各绿地中心到山体、滇池和市中心的距离,提取绿地外部50、100、200、400 m缓冲区的NDVI和NDBI指数。使用方差膨胀因子(VIF)检验所有环境因子的多重共线性,将VIF>5的因子剔除,得到最终进行分析的环境因子(表2)。
表 2 绿地环境因子
Table 2. Environment factor in green spaces
分类 指数名称 描述 内部环境因子 绿地面积/m2 单个绿地占地大小 绿地形状 单个绿地外轮廓形状 水体面积/m2 单个绿地内部水体占地面积 植被指数 单个绿地内部植被覆盖及其生长活力 乔木多度 单个绿地中乔木的总数量 乔木多样性 单个绿地在乔木的多样性指数 灌木多度 单个绿地中灌木的总数量 灌木多样性 单个绿地灌木的多样性指数 草本多样性 单个绿地中草本植物物种数量 外部环境因子 距山体距离/m 单个绿地中心距山体最近的距离 距滇池距离/m 单个绿地中心距滇池最近的距离 距市中心距离/m 单个绿地中心距市中心最近的距离 50 m缓冲区归一化建筑指数 单个绿地50 m缓冲区内建筑密度 400 m缓冲区归一化建筑指数 单个绿地400 m缓冲区内建筑密度 -
采用多度、Shannon-Wiener多样性指数、物种丰富度和Pielou均匀度指数表征昆明游憩性绿地鸟类多样性,分析3种游憩性绿地繁殖季节和非繁殖季节不同居留型鸟类4种指数差异情况;使用冗余分析(RDA)研究绿地内部环境因子对3种游憩性绿地不同居留型鸟类繁殖季节和非繁殖季节鸟类物种组成的影响;使用多元线性回归(MLR)研究绿地外部环境因子对3种游憩性绿地不同居留型鸟类繁殖季节和非繁殖季节鸟类物种组成的影响。不同绿地鸟类多样性、多元线性回归使用R 4.2.1中Vegan包、Ggplot 2包进行分析,使用Canoco 5进行RDA分析。
-
昆明市44块样地共记录到鸟类176种31 058只,隶属于17目53科。国家一级重点保护动物1种,为黄胸鹀Emberiza aureola;国家二级重点保护动物12种,分别为白胸翡翠Halcyon smyrnensis、黑鸢Milvus migrans、红隼Falco tinnunculus、游隼Falco peregrinus、苍鹰Accipiter gentilis、凤头鹰Accipiter trivirgatus、普通鵟Buteo japonicus、蛇雕Spilornis cheela、滇䴓Sitta yunnanensis、银耳相思鸟Leiothrix argentauris、黑喉噪鹛Pterorhinus chinensis、蓝喉歌鸲Luscinia svecica;列入国际自然保护联盟(IUCN)濒危物种红色名录的有2种,为近危物种(NT)滇䴓和极危物种(CR)黄胸鹀Emberiza aureola。
公园绿地记录到鸟类17目50科168种26 239只,国家一级重点保护动物1种,国家二级重点保护动物12种,鸟类居留型物种数以留鸟最多,其次是夏候鸟和旅鸟,冬候鸟最少。街旁绿地记录到鸟类14目37科80种3 663只,国家二级重点保护动物2种,鸟类居留型物种数以留鸟最多,其次是夏候鸟和旅鸟,冬候鸟最少。社区绿地记录到鸟类6目23科35种1 156只,其中国家二级重点保护动物1种,鸟类居留型物种数以留鸟最多,其次是夏候鸟,旅鸟最少,未见冬候鸟(表3)。
表 3 不同繁殖季节游憩性绿地鸟类物种组成
Table 3. Species composition of birds in recreational green spaces during different breeding seasons
季节 绿地类型 留鸟/种 冬候鸟/种 旅鸟/种 夏候鸟/种 国家二级重点保护动物/种 INCU濒危物种红色名录/种 总体 公园绿地 97 18 18 35 12 2 街旁绿地 44 6 8 22 2 0 社区绿地 24 0 3 5 1 0 繁殖季节 公园绿地 76 13 13 25 6 1 街旁绿地 35 1 7 17 1 0 社区绿地 14 0 3 5 0 0 非繁殖季节 公园绿地 76 15 11 28 9 1 街旁绿地 39 5 4 11 1 0 社区绿地 23 0 2 2 1 0 -
如图1所示:3种游憩性绿地的鸟类多度、物种丰富度、Shannon-Wiener多样性指数和Pielou均匀度指数存在差异。公园绿地鸟类物种丰富度、多度和Shannon-Wiener多样性指数均最高,其次是街旁绿地,社区绿地最低。Pielou均匀度指数呈相反情况,社区绿地最高,其次是街旁绿地,公园绿地最低。
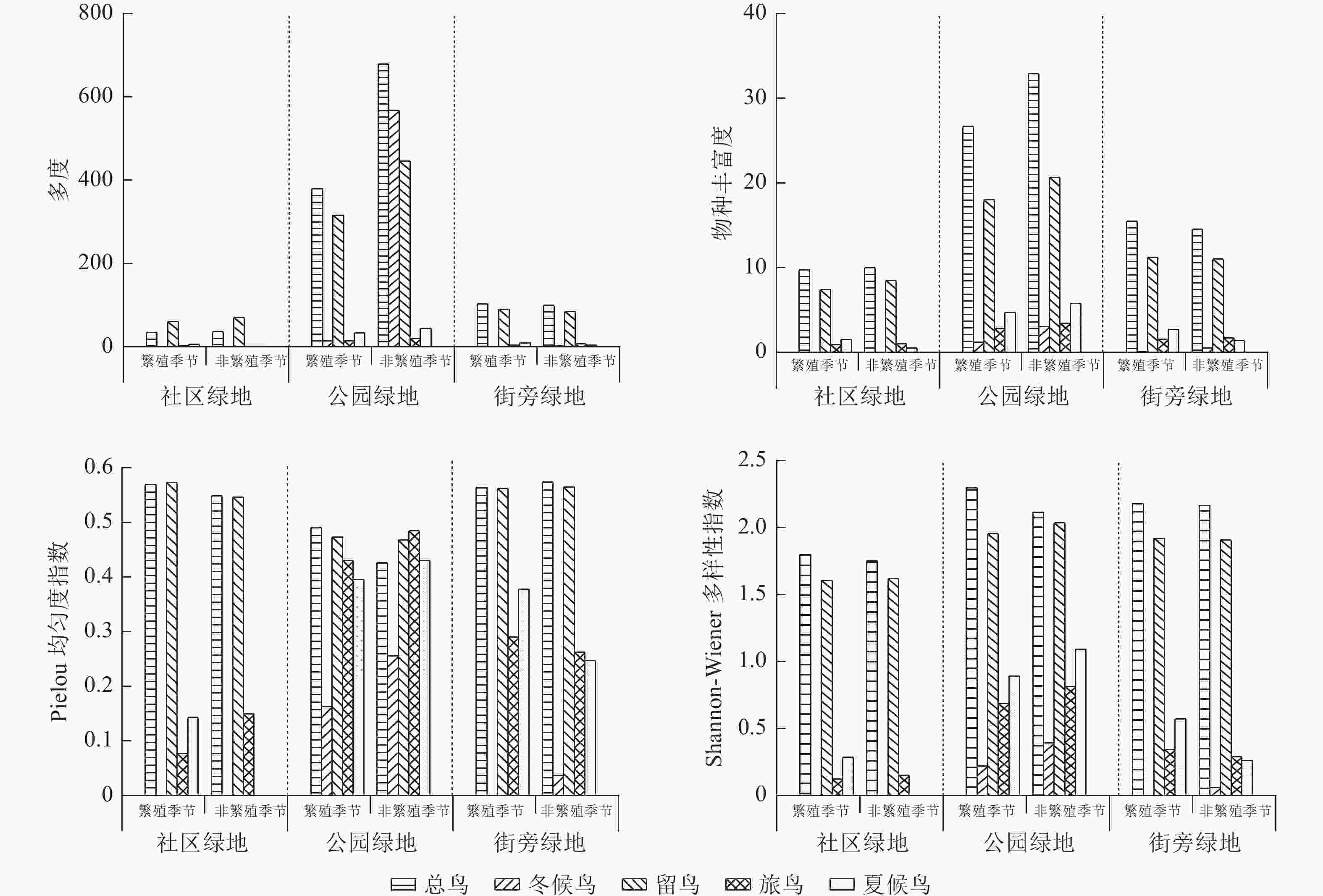
图 1 不同繁殖季节游憩性绿地鸟类多样性
Figure 1. Diversity of birds in recreational green spaces during different breeding seasons
从Pielou均匀度指数看,物种丰富度、多度和Shannon-Wiener多样性指数较低的鸟类,均匀度指数急速上升。从鸟类不同居留型来看,大部分情况下,留鸟的物种丰富度、多度和Shannon-Wiener多样性指数最高,冬候鸟最低,但在公园绿地中,冬候鸟多度超过留鸟。此外,不同居留型鸟类还受繁殖季节和非繁殖季节的影响。通常情况下,非繁殖季节鸟类物种丰富度和多度比繁殖季节多,但社区绿地和街旁绿地夏候鸟物种丰富度出现相反情况。公园绿地鸟类的Shannon-Wiener多样性指数均是非繁殖季节大于繁殖季节,但社区绿地的旅鸟和街旁绿地的冬候鸟出现了相反的情况。在Pielou均匀度指数中,3种绿地大部分鸟类非繁殖季节大于繁殖季节,社区绿地冬候鸟、夏候鸟,街旁绿地夏候鸟则出现了相反的结果。
-
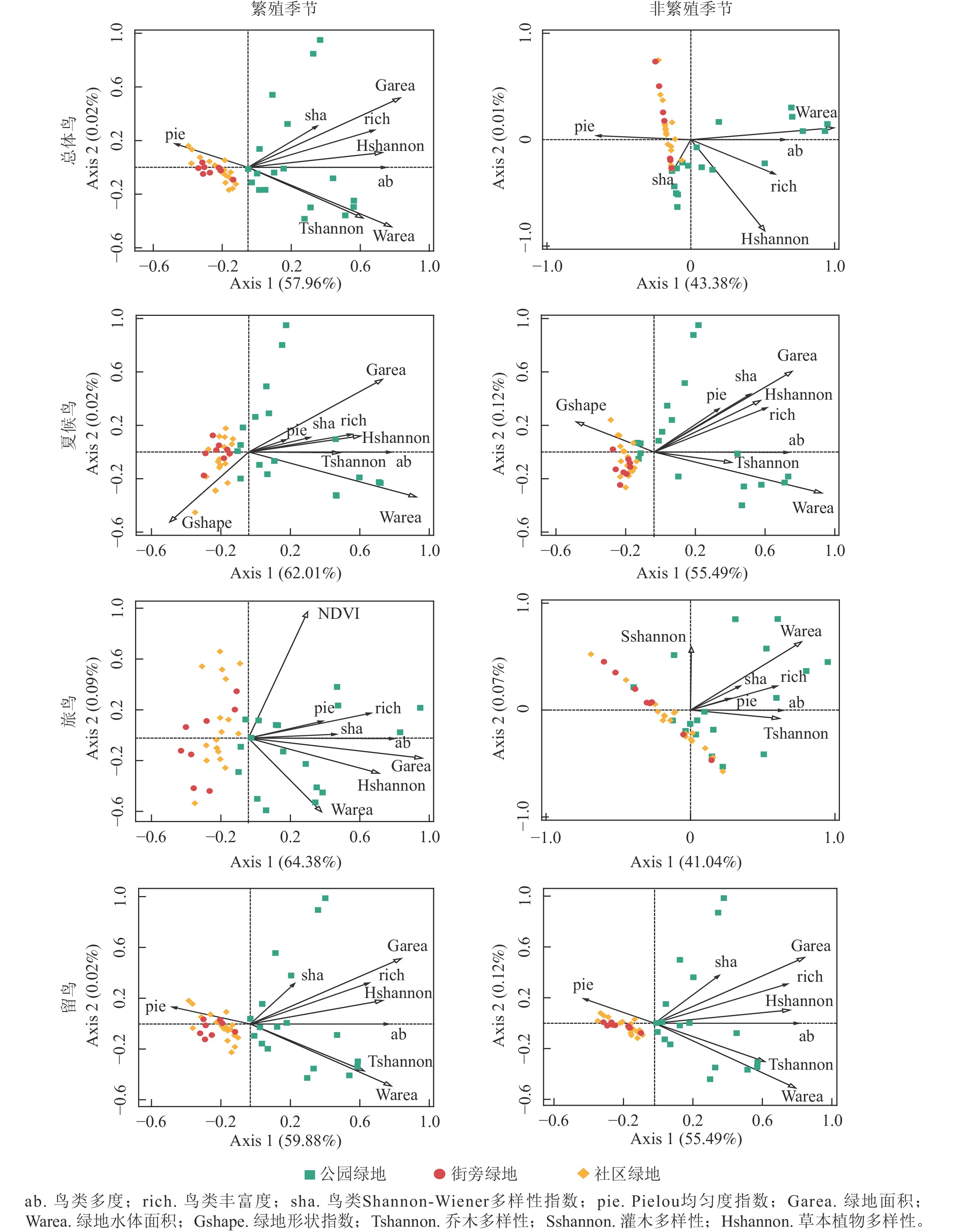
使用降趋对应分析(DCA)查看各轴梯度长度信息均小于3,适合用RDA分析。结果显示冬候鸟物种多样性在2个季节中均与内部环境因子无显著影响,故不论述。
如图2所示:公园绿地受到内部环境因子的影响最多。鸟类物种丰富度、多度、Shannon-Wiener多样性指数与鸟类绿地面积、草本植物多样性、乔木多样性、水体面积呈正相关,鸟类Pielou均匀度指数不受内部环境因子的影响。

图 2 不同繁殖季节鸟类多样性与内部环境因子的RDA分析
Figure 2. RDA analysis of bird diversity and internal environmental factors in different breeding seasons
从不同居留型来看,其他居留型鸟类的物种丰富度、多度、Shannon-Wiener多样性指数和Pielou均匀度指数均受内部环境因子的影响,但留鸟Pielou均匀度指数不受内部环境因子的影响。其中绿地形状指数与夏候鸟Pielou均匀度指数呈负相关,留鸟Shannon-Wiener多样性指数与水体面积呈负相关。此外繁殖季节受到内部环境因子的影响比非繁殖季节多。
-
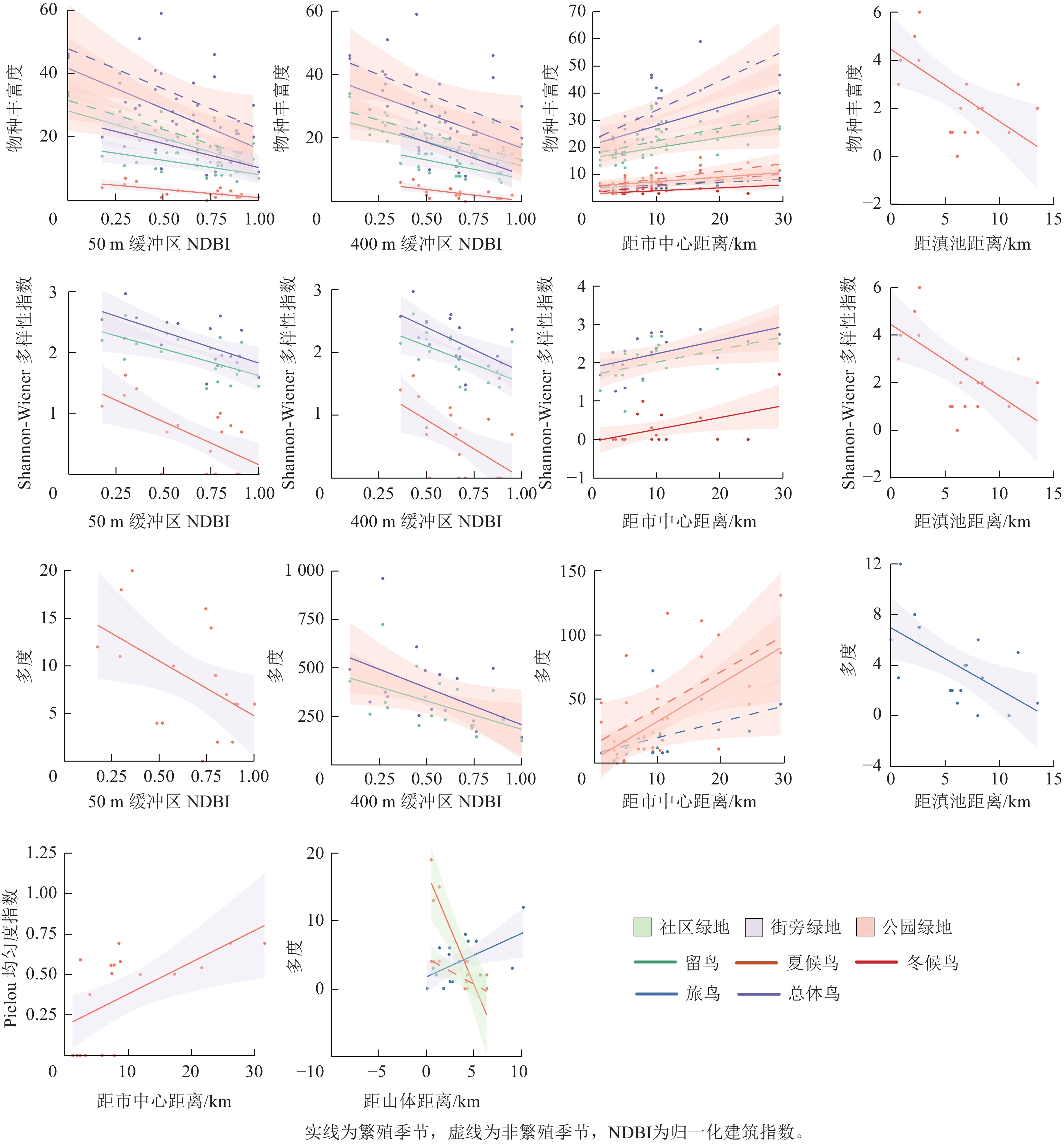
以外部环境因子(50 m缓冲区归一化建筑指数、400 m缓冲区归一化建筑指数、距市中心距离、距滇池距离、距山体距离)作为自变量,Shannon-Wiener多样性指数、Pielou均匀度指数、物种丰富度作为因变量,使用多元线性回归拟合3种游憩性绿地繁殖季节和非繁殖季节不同居留型鸟类多样性与外部环境因子的关系。
从图3可见:外部环境因子50、400 m缓冲区归一化建筑指数、距滇池距离与鸟类多样性均呈负相关,距市中心距离与鸟类多样性均呈正相关,距山体距离与街旁绿地旅鸟多度呈正相关,与社区绿地夏候鸟多度呈负相关。在3种游憩性绿地中,街旁绿地受外部环境因子的影响最多,其次是公园绿地,社区绿地最少。

图 3 鸟类多样性与外部环境因子多元线性拟合模型
Figure 3. Heat map of correlation between environmental variables and monthly bird population
从不同居留型来看,5种外部环境因子对夏候鸟多样性均有影响,留鸟物种丰富度和Shannon-Wiener多样性指数与50、400 m缓冲区建筑指数呈负相关,冬候鸟物种丰富度和Shannon-Wiener多样性指数与距市中心距离呈正相关,旅鸟多度与距滇池距离呈负相关,旅鸟Pielou均匀度指数与距山体距离呈正相关。繁殖季节鸟类多样性与50、400 m缓冲区建筑指数和距滇池距离呈负相关,与距市中心距离呈正相关,距山体距离会对不同鸟类产生不同影响。非繁殖季节鸟类多样性与50、400 m缓冲区建筑指数和距山体距离呈负相关,与距市中心距离呈正相关,距滇池距离的环境因子并未对非繁殖季节的鸟类造成影响。
-
本研究结果表明:公园绿地中,国家二级保护动物的数量最多,其次是街旁绿地,社区绿地最低。这些鸟类对生境要求高,均在靠近山体的大面积公园绿地中出现,该环境能让鸟类很好地躲避人类以及其他动物的干扰[10]。3种游憩性绿地中公园绿地的鸟类丰富度、多度和Shannon-Wiener多样性指数最高,街旁绿地次之,社区绿地最低,这与绿地面积越大,物种越多的研究结果一致[11]。街旁绿地和社区绿地的夏候鸟数量比冬候鸟类数量多,但公园绿地中冬候鸟数量比夏候鸟数量多,这主要是因为冬候鸟偏向于绿地以及水体面积较大的城市绿地[12]。
本研究表明:公园绿地的冬候鸟数量超过留鸟,这可能是因为大量冬候鸟冬季来昆明驻足过冬,导致数量剧增[13]。通常情况下,3种绿地中非繁殖季节鸟类要比繁殖季节多,主要是由于非繁殖季节鸟类无需育雏,可选择的休憩觅食场地多[14]。但是社区绿地和街旁绿地的夏候鸟、冬候鸟在繁殖季节出现的频次比非繁殖季节高,这可能是因为繁殖季节,鸟类为了短时间内获取食物[15],会选择城市化程度较高的社区绿地和街旁绿地。
-
本研究表明:除绿地形状指数以外,其他内部环境因子都对不同居留型鸟类多样性具有正面影响,水体面积对鸟类多样性影响最大,因为水体能为绿地鸟类提供多样的异质性生境[15]。绿地面积也是影响鸟类的最主要环境因子,可为鸟类提供休憩和觅食的场所[16]。其次是乔木和草本多样性指数,植被多样性较高的地区鸟类物种丰富度更高,绿地内部的植被指数体现植被覆盖程度,表明植被指数高的绿地更能吸引鸟类驻足[17]。
从不同绿地类型看,公园绿地对鸟类多样性具有促进作用,因为公园绿地有利鸟类生存的内部环境因子比另外2种绿地丰富,很多鸟类偏向于这样的绿地。街旁绿地和社区绿地的面积小,城市化和人为干扰较强,影响鸟类生活[18]。
绿地形状指数对夏候鸟产生负面影响,这与之前的研究结果不一致,可能是因为形状指数越高,绿地内部各生物越稳定,夏候鸟会被原本绿地内的常驻鸟驱逐[19]。在繁殖季节中,留鸟的Shannon-Wiener多样性指数与水体面积呈负相关,可能是因为水体周围被大量来自昆明的冬候鸟占据[20],留鸟只能选择远离水体的区域。繁殖季节中,鸟类对绿地内部环境因子要求高,因为育雏需要隐蔽场所,绿地面积与水体面积的增加会减少人为干扰[21]。乔木与草本植物多样性增加,可为鸟类提供更加安全的场所。在进行RDA分析时,发现冬候鸟多样性与各环境因子没有显著性,但是实地调查时发现:冬候鸟对水体要求很高[22],靠近滇池的绿地中能发现大量像红嘴鸥Chroicocephalus ridibundus、黑水鸡Gallinula chloropus、骨顶鸡Fulica atra等冬候鸟,而其他绿地中发现较少。
-
外部环境因子主要显示绿地周边环境对鸟类的影响。50、400 m缓冲区归一化建筑指数、距市中心距离对鸟类多样性均有负面影响。有研究显示:建筑指数越高,鸟类多样性指数就逐渐降低[23]。建筑指数升高,人为干扰因素随之增加,噪音、人流量和车流量大,导致城市敏感物种减少,而适应城市环境的鸟类数量增加甚至占据生态位。距市中心距离对公园绿地所有居留型鸟类均造成负面影响,公园绿地为它们提供类似自然的栖息环境,但距市中心越近,就打破了原本自然化的栖息地,使得很多鸟类选择逃离。同时距市中心越远,街旁绿地中夏候鸟的Pielou均匀度也越高,因为没有城市化的影响,鸟类都在绿地中发展了自己的生态位[24]。距山体距离对街旁旅鸟有正面影响,对社区夏候鸟具有负面影响,旅鸟将绿地作为短暂驻足地,对场地的选取以偏向自然为主;社区绿地中居住的都是对城市高度适应的鸟类,距山体越近意味着城市食物来源就会减少[24],影响夏候鸟的数量。距滇池距离越近,鸟类多样性就越高,这是因为水体能够增加景观的异质性,吸引更多鸟类驻足[25]。
公园绿地鸟类受城市外部环境影响最多,其次是街旁绿地,社区绿地最少。这是因为公园绿地面积大,植物和生境群落丰富,很多对城市敏感的鸟类选择公园绿地栖息[26]。社区绿地在居住区内部,面积小且城市化程度最高,能在社区绿地驻留的鸟类大部分是已经适应城市环境的,对周边城市化的变化基本不敏感。街旁绿地面积介于公园绿地和社区绿地之间,它的城市化程度没有社区绿地高,所以鸟类群落多样性相对较高,但它们对外部城市化的影响也会做出较强反应[27]。繁殖季节鸟类比非繁殖季节鸟类更容易受到外部环境因子的影响,这是因为繁殖季节鸟类为了保护幼鸟,会选择城市化较低的地区繁育[28]。
-
昆明市3种游憩性绿地中鸟类多样性最高的是公园绿地,其次是街旁绿地,社区绿地最少,这与城市内部环境因子和外部环境因子有关。大面积的绿地和水体以及丰富的植物群落均对鸟类多样性的提高具有积极作用,应加强游憩性绿地内部环境的保护,增强绿地内部生境异质性,为不同鸟类提供更适宜的栖息条件。绿地周边的建筑指数和距市中心较短的距离会对鸟类造成较强的负面影响,建议拓宽绿化隔离带,加强各绿地间的连通性,减少建筑密度和城市化影响。距滇池距离越近,鸟类多样性越高,应减少滇池附近过度开发,保护原有生态环境。山体是对城市化敏感鸟类的主要栖息地,山体附近绿地建设应提供足够的水源与隐蔽场所,满足鸟类的基本生境需求。
Response of bird diversity to environmental factors in recreational green spaces in Kunming City
-
摘要:
目的 分析昆明市不同游憩性绿地鸟类群落组成差异及其原因,研究城市绿地环境对鸟类群落的影响。 方法 2021年7月至2022年8月对昆明市3种游憩性绿地(建城区公园绿地、街旁绿地和社区绿地)进行鸟类调查,使用冗余分析和多元线性回归,分析3种游憩性绿地繁殖季节和非繁殖季节不同居留型鸟类多样性与绿地环境因子的关系。 结果 ①3种游憩性绿地共记录到鸟类176种31 058只,隶属于17目53科,其中国家一级重点保护动物1种,国家二级重点保护动物12种。②公园绿地鸟类物种丰富度、多度和Shannon-Wiener多样性指数均最高,街旁绿地其次,社区绿地最低,但Pielou均匀度指数呈现相反情况。③冗余分析显示:除绿地形状指数外,其他内部环境因子对提高鸟类多样性均具有促进作用。④多元线性回归表明:50和400 m缓冲区内建筑指数与鸟类多样性呈负相关;距市中心距离与鸟类多样性呈正相关;距滇池距离与街旁绿地鸟类多样性呈负相关;距山体距离与街旁绿地鸟类多样性呈正相关,与社区绿地鸟类多样性呈负相关。 结论 未来游憩性绿地建设应减少周边建筑对公园绿地和街旁绿地的影响,增加异质景观生境,重点营造社区绿地内部植被群落丰富度。图3表3参28 Abstract:Objective The objective of this study is to investigate bird communities in parks, roadside green spaces, and community green spaces in Kunming urban construction zone, analyze the differences in bird community composition across different recreational green spaces and their root causes, and explore the impact of urban green space environments on bird communities. Method From July 2021 to August 2022, birds were investigated in three types of recreational green spaces in Kunming City. Redundancy analysis and multiple linear regression were used to analyze the relationship between the diversity of resident birds and environmental factors during breeding and non-breeding seasons. Result (1) A total of 31 058 individuals in 176 species were recorded, belonging to 17 orders and 53 families, which included 1 species under national first-class key protection and 12 species under national second-class key protection. (2) Parks had the highest richness, abundance, and Shannon-Wiener diversity index of bird species, followed by roadside green spaces, and community green spaces had the lowest. However, the Pielou evenness index showed the opposite trend. (3) Redundancy analysis indicated that other internal environmental factors, except for the green space shape index, had a promoting effect on improving bird diversity. (4) Multiple linear regression showed the building index in buffer zones of 50 and 400 m was negatively correlated with bird diversity. The distance from the city center was positively correlated with bird diversity. The distance from Dian Lake was negatively correlated with bird diversity in roadside green spaces. The distance from the mountain was positively correlated with bird diversity in roadside green spaces, but negatively correlated with bird diversity in community green spaces. Conclusion In the future construction of recreational green spaces, it is necessary to reduce the impact of surrounding buildings on parks and roadside green spaces, increase heterogeneous landscape habitats, and focus on enhancing the richness of vegetation communities inside community green spaces. [Ch, 3 fig. 3 tab. 28 ref.] -
Key words:
- birds /
- season /
- recreational green spaces /
- residence type /
- Kunming City
-
图 1 不同繁殖季节游憩性绿地鸟类多样性
Figure 1 Diversity of birds in recreational green spaces during different breeding seasons
图 2 不同繁殖季节鸟类多样性与内部环境因子的RDA分析
Figure 2 RDA analysis of bird diversity and internal environmental factors in different breeding seasons
图 3 鸟类多样性与外部环境因子多元线性拟合模型
Figure 3 Heat map of correlation between environmental variables and monthly bird population
表 1 调查样地基本信息
Table 1. Basic information of survey site
序号 绿地名称 绿地类型 面积/hm2 样线数量/条 序号 绿地名称 绿地类型 面积/hm2 样线数量/条 1 郊野公园 公园绿地 82.90 7 23 低碳中心 街旁绿地 1.32 1 2 黑龙潭公园 公园绿地 86.40 6 24 金汁河桥 街旁绿地 1.87 1 3 金殿森林公园 公园绿地 71.30 6 25 西京寺 街旁绿地 2.91 1 4 洛龙公园 公园绿地 40.00 10 26 葵花公社 街旁绿地 2.16 1 5 渔浦寒泉森林生态湿地公园 公园绿地 41.20 10 27 白邑寺 街旁绿地 2.43 1 6 隅山公园 公园绿地 11.80 6 28 金树春带状公园 街旁绿地 1.45 1 7 瀑布公园 公园绿地 32.50 8 29 董家湾 街旁绿地 0.93 1 8 月牙潭公园 公园绿地 16.40 5 30 杰霖园 街旁绿地 0.79 1 9 昙华寺公园 公园绿地 10.20 5 31 市级行政中心 街旁绿地 1.74 1 10 官渡森林公园 公园绿地 24.10 6 32 永昌湿地 街旁绿地 1.51 1 11 如意公园 公园绿地 27.00 8 33 龙马路 街旁绿地 2.60 1 12 龙江公园 公园绿地 5.37 5 34 华夏天景湾 街旁绿地 1.53 1 13 翠湖公园 公园绿地 24.20 5 35 前卫西路 街旁绿地 1.00 1 14 海洪湿地公园 公园绿地 20.60 5 36 蓝光欢乐城 街旁绿地 1.74 1 15 西亮塘湿地公园 公园绿地 48.40 8 37 博众花园 社区绿地 0.53 1 16 斗南湿地公园 公园绿地 52.70 9 38 水晶骊城 社区绿地 0.91 1 17 捞鱼河公园 公园绿地 55.20 7 39 东旭俊城 社区绿地 1.39 1 18 西华园 公园绿地 6.79 3 40 关上法治 社区绿地 1.27 1 19 银杏公园 街旁绿地 2.48 1 41 金泰国际 社区绿地 1.60 1 20 郎溪街 街旁绿地 2.44 1 42 五华广场 社区绿地 1.27 1 21 鼓楼公园 街旁绿地 0.95 1 43 五星彩园 社区绿地 0.58 1 22 洛羊街道办事处 街旁绿地 3.10 1 44 碧鸡名城 社区绿地 1.39 1  下载: 导出CSV
下载: 导出CSV
表 2 绿地环境因子
Table 2. Environment factor in green spaces
分类 指数名称 描述 内部环境因子 绿地面积/m2 单个绿地占地大小 绿地形状 单个绿地外轮廓形状 水体面积/m2 单个绿地内部水体占地面积 植被指数 单个绿地内部植被覆盖及其生长活力 乔木多度 单个绿地中乔木的总数量 乔木多样性 单个绿地在乔木的多样性指数 灌木多度 单个绿地中灌木的总数量 灌木多样性 单个绿地灌木的多样性指数 草本多样性 单个绿地中草本植物物种数量 外部环境因子 距山体距离/m 单个绿地中心距山体最近的距离 距滇池距离/m 单个绿地中心距滇池最近的距离 距市中心距离/m 单个绿地中心距市中心最近的距离 50 m缓冲区归一化建筑指数 单个绿地50 m缓冲区内建筑密度 400 m缓冲区归一化建筑指数 单个绿地400 m缓冲区内建筑密度
下载: 导出CSV
表 3 不同繁殖季节游憩性绿地鸟类物种组成
Table 3. Species composition of birds in recreational green spaces during different breeding seasons
季节 绿地类型 留鸟/种 冬候鸟/种 旅鸟/种 夏候鸟/种 国家二级重点保护动物/种 INCU濒危物种红色名录/种 总体 公园绿地 97 18 18 35 12 2 街旁绿地 44 6 8 22 2 0 社区绿地 24 0 3 5 1 0 繁殖季节 公园绿地 76 13 13 25 6 1 街旁绿地 35 1 7 17 1 0 社区绿地 14 0 3 5 0 0 非繁殖季节 公园绿地 76 15 11 28 9 1 街旁绿地 39 5 4 11 1 0 社区绿地 23 0 2 2 1 0
下载: 导出CSV
-
[1] 李翔宇, 张喆, 马红艳, 等. 城市鸟类群落与绿地关系研究进展[J]. 世界林业研究, 2022, 35(5): 49 − 53. LI Xiangyu, ZHANG Zhe, MA Hongyan, et al. Research progress in relationship between urban bird communities and green spaces [J]. World Forestry Research, 2022, 35(5): 49 − 53. [2] 李晓军. 甘肃酒泉城市公园鸟类群落结构及多样性分析[J]. 干旱区资源与环境, 2022, 36(6): 95 − 104. LI Xiaojun. The analysis of bird community structure and diversity in the city park of Jiuquan, Gansu Province [J]. Journal of Arid Land Resources and Environment, 2022, 36(6): 95 − 104. [3] 赵雪琴, 吴岚, 王玉玉, 等. 北京温榆河公园不同建成时间城市绿地鸟类群落组成的空间变化与栖息地质量评价[J]. 生态学杂志, 2023, 42(12): 3008 − 3018. ZHAO Xueqin, WU Lan, WANG Yuyu, et al. Spatial variations of bird community composition and habitat quality evaluation of urban green scape at different construction times in Wenyu River Park in Beijing [J]. Chinese Journal of Ecology, 2023, 42(12): 3008 − 3018. [4] GONZALEZ-LAGOS C, CARDADOR L, SOL D. Invasion success and tolerance to urbanization in birds [J]. Ecography, 2021, 44(11): 1642 − 1652. [5] 郑英茂, 刘娟, 潘成椿, 等. 九龙山国家级自然保护区鸟类组成特征分析[J]. 浙江农林大学学报, 2020, 37(6): 1167 − 1176. ZHENG Yingmao, LIU Juan, PAN Chengchun, et al. Analysis of avian composition characteristics in Jiulongshan National Nature Reserve [J]. Journal of Zhejiang A&F University, 2020, 37(6): 1167 − 1176. [6] de LEON L F, SHARPE D M T, GOTANDA K M, et al. Urbanization erodes niche segregation in Darwin’s finches [J]. Evolutionary Applications, 2019, 12(7): 1329 − 1343. [7] IKNAYAN K J, HEATH S K, TERRILL S B, et al. Patterns in bird and pollinator occupancy and richness in a mosaic of urban office parks across scales and seasons[J/OL]. Ecology and Evolution, 2024, 14: e109583[2023-12-01]. doi: 10.1002/ece3.10958. [8] 约翰·马敬能, 卡伦·菲利普斯, 何芬. 中国鸟类野外手册[M]. 长沙: 湖南教育出版社, 2000. JOHN Mackinnon, KAREN Phillipps, HE Fen. A Field Guide to the Birds of China [M]. Changsha: Hunan Education Press, 2000. [9] 郑光美. 中国鸟类分类与分布名录[M]. 北京: 科学出版社, 2005. ZHENG Guangmei. A Checklist on the Classification and Distrbution of the Birds of China [M]. Beijing: Science Press, 2005. [10] ZHANG Wenwen, ZHOU Ying, FANG Xuelan, et al. Effects of environmental factors on bird communities in different urbanization grades: an empirical study in Lishui, a mountainous area of eastern China [J/OL]. Animals, 2023, 13(5): 882[2023-12-01]. doi: 10.3390/ani13050882. [11] 朱光, 王雪, 张文文, 等. 城市景观格局对鸟类群落的影响: 以南京溧水区为例[J]. 生态与农村环境学报, 2022, 38(3): 327 − 333. ZHU Guang, WANG Xue, ZHANG Wenwen, et al. Effects of urban landscape pattern on bird community: a case study of Lishui District, Nanjing [J]. Journal of Ecology and Rural Environment, 2022, 38(3): 327 − 333. [12] MEISSNER W, WITKOWSKA M. The effect of the temperature on local differences in the sex ratio of Mallards Anas platyrhynchos wintering in an urban habitat[J/OL]. Acta Oecologica-International Journal of Ecology, 2023, 119: 103900[2023-12-01]. doi: 10.1016/j.actao.2023.103900. [13] PACHECO-MUNOZ R, CEJA-MADRIGAL A, PEREZ-NEGRON E, et al. Wintering town: individual quality of migratory warblers using green urban areas in a Neotropical City[J/OL]. Frontiers in Ecology and Evolution, 2022, 9: 779507[2023-12-01]. doi: 10.3389/fevo.2021.779507. [14] ZHOU Daqing, CHU L M. Do avian communities vary with season in highly urbanized Hong Kong? [J]. Wilson Journal of Ornithology, 2014, 126(1): 69 − 80. [15] 梁凤, 周立志. 环巢湖湿地公园鸟类多样性的空间变化[J]. 生态学杂志, 2023, 42(7): 1687 − 1698. LIANG Feng, ZHOU Lizhi. Spatial changes of bird diversity in the wetland parks alongside Chaohu Lake [J]. Chinese Journal of Ecology, 2023, 42(7): 1687 − 1698. [16] 金秋爽, 朱浩, 王爽, 等. 杭州市绿地中常见园林树种果实引鸟特征研究[J]. 浙江农林大学学报, 2022, 39(6): 1359 − 1368. JIN Qiushuang, ZHU Hao, WANG Shuang, et al. Preference of birds to fruit characteristics of common landscaping trees in campus green spaces in Hangzhou [J]. Journal of Zhejiang A&F University, 2022, 39(6): 1359 − 1368. [17] PACHECO-MUNOZ R, AGUILAR-GOMEZ M A, SCHONDUBE J E. Overwintering in a megacity: urban green areas and migratory birds in Mexico City [J/OL]. Urban Forestry & Urban Greening, 2022, 73: 127614[2023-12-01]. doi: 10.1016/j.ufug.2022.127614. [18] 谭昕蔚. 南京城市公园鸟类嵌套格局及其影响因素[D]. 南京: 南京师范大学, 2021. TAN Xinwei. Nestedness and Influencing Factors of Bird Assemblages in Nanjing Urban Parks [D]. Nanjing: Nanjing Normal University, 2021. [19] GRZEDZICKA E. Bird feeder explorers are not attracted by the seeds of invasive weeds in winter [J/OL]. Diversity-Basel, 2024, 16: 812[2023-12-01]. doi: 10.3390/d16020081. [20] CRISTALDI M A, GODOY I N, LEVEAU L M. Responses of urban bird assemblages to land-sparing and land-sharing development styles in two Argentinian Cities [J/OL]. Animals, 2023, 13(5): 894[2023-12-01]. doi: 10.3390/ani13050894. [21] HUANG Peilin, ZHENG Dulai, YAN Yijing, et al. Effects of landscape features on bird community in winter urban parks[J/OL]. Animals, 2022, 12(23): 3442[2023-12-01]. doi: 10.3390/ani12233442. [22] LAKATOS T, CHAMBERLAIN D E, GARAMSZEGI L Z, et al. No place for ground-dwellers in cities: a meta-analysis on bird functional traits [J/OL]. Global Ecology and Conservation, 2022, 38: e02217[2023-12-01]. doi:10.1016/j.gecco.2022.e02217. [23] TODD E C D. Foodborne disease and food control in the Gulf States [J]. Food Control, 2017, 73: 341 − 366. [24] YORIO P, MARINAO C, KASINSKY T, et al. Patterns of plastic ingestion in Kelp Gull (Larus dominicanus) populations breeding in northern Patagonia, Argentina[J/OL]. Marine Pollution Bulletin, 2020, 156: 111240[2023-12-01]. doi: 10.1016/j.marpolbul.2020.111240. [25] ZHAO Zixi, BORZEE A, LI Jinghao, et al. Urban bird community assembly mechanisms and driving factors in university campuses in Nanjing, China [J/OL]. Animals, 2023, 13(4): 673[2023-12-01]. doi: 10.3390/ani13040673. [26] 隋金玲, 李凯, 胡德夫, 等. 城市化和栖息地结构与鸟类群落特征关系研究进展[J]. 林业科学, 2004, 40(6): 147 − 152. SUI Jinling, LI Kai, HU Defu, et al. Progresses of studies on biodiversity of urban birds [J]. Cientla Silvae Sinicae, 2004, 40(6): 147 − 152. [27] STROHBACH M W, LERMAN S B, WARREN P S. Are small greening areas enhancing bird diversity? Insights from community-driven greening projects in Boston [J]. Landscape and Urban Planing, 2013, 114: 69 − 79. [28] KAUSHIK M, TIWARI S, MANISHA K. Habitat patch size and tree species richness shape the bird community in urban green spaces of rapidly urbanizing Himalayan foothill region of India [J]. Urban Ecosystems, 2022, 25(2): 423 − 436. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230604
 点击查看大图
点击查看大图
计量
- 文章访问数: 1166
- HTML全文浏览量: 293
- PDF下载量: 33
- 被引次数: 0
