




-
化学除草剂的危害愈来愈受到人们重视,利用植物化感作用对田间杂草进行生态防控被认为是环境友好,促进农业可持续发展的新技术,并已被列入当代农业生态学研究的重要研究方向[1-2]。香豆素(C9H6O2)是香豆素类物质中形式最简单的化合物。邬彩霞等[3]采用色谱分离等技术,发现黄花草木樨Melilotus officinalis中化感物质含量最高的是香豆素。黄花草木樨对藜Chenopodium album[4],稗草Echinochloa crusgalli[5],苏丹草Sorghum sudanense[6]和多花黑麦草Lolium multiflorum[7]等多种杂草均存在较强的抑制作用。此外,覃逸明等[8]研究发现:在凤丹Paeonia ostii根际土壤中存在的5种化感物质中,香豆素抑制作用最强。CHON等[9]研究发现:与肉桂酸、香草酸等其他酚酸类化感物质相比,香豆素对苜蓿Medicago sativa的化感抑制作用最强。RAZAVI等[10]研究发现:当香豆素质量浓度达100 mg·L-1时,可完全阻碍生菜Lactuca sativa种子萌发及其幼苗根、茎生长。上述研究均表明:香豆素有作为新型除草剂的开发利用潜力,而有关香豆素对入侵植物的化感作用研究尚未见报道。节节麦Aegilops tauschii为禾本科Gramineae山羊草属Aegilops植物,最早起源于西亚及东欧等地,现已成为世界恶性杂草[11]。1955年,节节麦在中国河南省新乡地区首次被发现,如今已广泛入侵至渭河流域陕西、甘肃,黄河流域河北、河南、山东以及内蒙古等地,并成为中国麦田中最难防除的恶性杂草之一[12-13]。据报道,仅2003年河北南部地区由于节节麦危害导致的小麦损失就达10%~25%[14]。因严重威胁粮食的生产与安全,早在2007年节节麦已被列入中国首次发布的《中华人民共和国进境植物检疫性有害生物名录》[15]。生产上仅有甲基二磺隆和异丙隆对节节麦具有一定防除效果,但由于不同小麦品种及节节麦种群对上述2种除草剂的敏感性存在差异[16-17],导致它们在实际生产中的推广应用受到一定限制。这也是节节麦入侵范围愈来愈广泛的重要原因之一。为此,本研究以入侵植物节节麦为试材,探讨香豆素对其种子萌发及幼苗生长的影响,以期为节节麦的生物防控提供参考。
-
2017年5月,节节麦种子来源于河南省新乡市农业科学院试验田(35°18′N,113°52′E)。研究在河南科技大学开元校区林学院园林植物实验室进行。固体香豆素来源于上海试剂厂。
-
香豆素溶液配置:准确称取(万分之一天平)75 mg香豆素,蒸馏水定容至500 mL,充分溶解后得150 mg·L-1香豆素溶液,分别稀释得到50,100 mg·L-1的溶液,置4 ℃冰箱保存。使用时提前取出,室内放置3~5 h,避免低温溶液对种子萌发造成影响。
种子萌发试验:采用培养皿沙培法[18]。选取籽粒饱满、大小基本一致的节节麦种子,先用体积分数为1%的次氯酸钠溶液浸泡10 min,之后蒸馏水反复冲洗,室内自然晾干备用。将种子置于铺有经高温消毒处理石英砂的培养皿(直径12 cm)内,加入10 mL(经预试验确定)处理液,50粒·皿-1,利用透明保鲜膜覆盖包裹培养皿(保证培养皿口保鲜膜平展)以减少试验过程中水分的散失,并称量记录每皿的质量(试验期间根据石英砂颜色,加入适量处理液保持每皿质量不变)。各处理5次重复,各重复3皿。以加等量蒸馏水的处理为对照(ck)。置于昼夜25 ℃/15 ℃,光暗各12 h的光照培养箱内培养。每天观察统计1次种子萌发情况(以胚根长度大于等于种子长度为萌发标准),培养10 d后各处理随机选取10株幼苗分别测量根长及苗高。其中,发芽率RG=(10 d内供试种子发芽数/供试种子数)×100%,发芽指数IG= ∑(Gt/Dt),其中:Dt为相应发芽天数,Gt为逐日发芽种子数。抑制率[4, 19]RI=(T0-T1)/T0。其中:T1为处理值,T0为对照值。RI<0为促进作用,RI>0,为抑制作用,RI绝对值大小代表抑制的强弱。
幼苗生长试验:首先对节节麦种子进行催芽处理(水浸泡24 h,25 ℃培养箱内),选出露白基本一致的种子播种于装有适量石英砂的塑料杯(开口直径10 cm,高15 cm)内,10粒·杯-1,随后加入15 mL(经预备实验确定)不同质量浓度的处理液,以加等量蒸馏水的为对照(ck),各处理5次重复,各重复3杯。置于昼夜25 ℃/15 ℃,光暗各12 h的光照培养箱内培养。并分别在处理的5,10和15 d采样,取全株进行各项生理指标的测定。电导法测定相对电导率(REC)[20];酸性茚三酮法测定脯氨酸质量分数,氮蓝四唑光还原法测超氧化物歧化酶活性,硫代巴比妥酸法测定丙二醛质量摩尔浓度[21]。
-
用Excel对数据进行初步处理,利用SPSS 18.0进行单因素方差分析,数据为平均值±标准差。
-
随香豆素溶液质量浓度的增加,节节麦种子发芽率、发芽指数均明显下降(表 1)。不同质量浓度处理的发芽率、发芽指数与对照差异均达到显著水平(P<0.05)。150 mg·L-1处理中,发芽率仅有31.60%,较对照下降66.38%。此外,从抑制率可看出,发芽率及发芽指数的抑制率均大于0,且呈质量浓度效应的变化。由此表明,香豆素溶液对节节麦种子萌发存在较强的抑制作用。
表 1 香豆素溶液对节节麦种子萌发和幼苗生长过程中生长指标的影响
Table 1. Coumarin solution influence of Aegilops tauschii growth indexes on seed germination and growth of seedlings
ρ/(mg·L-1) 发芽率/% 发芽率抑制率 发芽指数 发芽指数抑制率 苗高/cm 苗高抑制率 根长/cm 根长抑制率 0(对照) 94.00 ± 0.11 a 0.00 33.73 ± 0.87 a 0.00 16.52 ± 0.58 a 0.00 13.62 ± 0.76 a 0.00 50 71.60 ± 2.19 b 0.24 22.84 ± 1.10 b 0.32 14.05 ± 0.41 ab 0.14 9.87 ± 0.14 b 0.49 100 45.60 ± 1.95 c 0.51 14.38 ± 1.42 c 0.57 10.15 ± 0.65 c 0.38 7.44 ± 0.31 c 0.60 150 31.60 ± 0.99 d 0.66 7.59 ± 1.19 d 0.77 7.16 ± 0.16 d 0.57 4.81 ± 0.11 d 0.65 说明:同列不同小写字母表示不同处理间差异显著 随香豆素质量浓度的增加,节节麦幼苗的苗高及根长同样呈明显的减少趋势。其中,100和150 mg·L-1的处理苗高均显著低于对照(P<0.05)。此外,不同质量浓度处理的根长均显著低于对照(P<0.05)。香豆素对苗高及根长的抑制率均大于0,且也随溶液质量浓度的增加而增大。由此表明,香豆素溶液对节节麦幼苗的生长也存在明显抑制作用。
-
由图 1A可知:相同质量浓度处理中,节节麦幼苗的相对电导率随时间的延长呈快速上升的变化。其中,50 mg·L-1处理中,处理后第5天时,相对电导率略低于对照,之后则均高于对照,至第15天时较第5天时增加显著(P<0.05)。100和150 mg·L-1处理中,相对电导率均高于对照,至第10天时较第5天时增加均已达到显著水平(P<0.05)。

图 1 不同质量浓度香豆素对节节麦幼苗相对电导率及丙二醛质量摩尔浓度的影响
Figure 1. Relative electric conductivity and MDA content of Aegilops tauschii seedling under different concentrations of coumarin solution
相同质量浓度处理中,丙二醛质量摩尔浓度随时间的延长呈逐渐上升的变化(图 1B)。其中,50 mg·L-1处理中,至第15天时增加达到显著水平(P<0.05)。100和150 mg·L-1处理中,至第10天时丙二醛质量摩尔浓度较第5天时增加已达到显著水平(P<0.05)。同样,相同时间下,丙二醛质量摩尔浓度与香豆素质量浓度呈正相关。
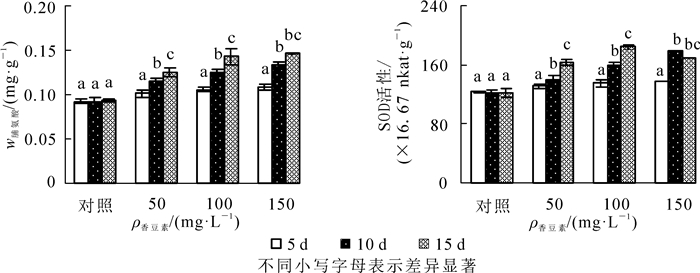
相同质量浓度处理中,节节麦幼苗的脯氨酸质量分数随时间延长呈逐渐增加的变化(图 2A)。其中,不同质量浓度香豆素溶液处理中,至处理后第10天时脯氨酸质量分数较第5天时增加均已达到显著水平(P<0.05)。150 mg·L-1处理中,至第15天时较第10天时增加未达到显著水平。且相同时间下,脯氨酸质量分数与香豆素质量浓度呈正相关。

图 2 不同质量浓度香豆素对节节麦幼苗脯氨酸质量分数及超氧化物歧化酶活性的影响
Figure 2. Proline content and SOD activity of Aegilops tauschii seedling under different concentrations of coumarin solution
50和100 mg·L-1处理中,超氧化物歧化酶活性随时间的延长呈持续增加的变化,而150 mg·L-1处理中,超过氧化物歧化酶活性呈先升后降的变化(图 2B)。其中,50及100 mg·L-1处理中,至处理后第10天时超氧化物歧化酶活性较第5天时增加已达到显著水平(P<0.05)。150 mg·L-1处理中,至第10天时超氧化物歧化酶活性较第5天时增加显著,至第15天时较第10天时略有下降,但差异不显著。且相同时间下,超氧化物歧化酶活性与香豆素质量浓度呈正相关。
-
种子能否正常萌发决定了植物的繁衍及生存,对该物种的更新至关重要。而发芽率的增加及下降则会降低植物在群落中的竞争力及多度[22]。幼苗阶段在植物的生命周期中较为关键,影响着植物的形态建成,且是植物对外界环境较为敏感的时期[23]。本试验结果表明:香豆素溶液对节节麦种子萌发产生明显的抑制作用,且抑制作用随香豆素溶液质量浓度的增加而增大。在王婧怡等[6]对苏丹草,姚丹丹等[7]对多花黑麦草的研究中均发现:香豆素溶液对上述2种植物种子萌发存在一定的化感抑制作用。香豆素溶液处理造成节节麦种子发芽率下降的原因可能有:一是香豆素可能影响了种子萌发所需关键性酶等物质,造成萌发过程中萌发所需能量物质的缺乏,从而导致种子活力的下降;二是香豆素可能改变了节节麦种皮透性,进而影响种子的吸水作用,造成种子内部物质的外渗,而此类物质造成了真菌病原体的生长,导致种子腐烂而无法萌发。本研究的种子萌发后期,也确有种子腐烂现象的发生。此外,香豆素溶液处理对节节麦幼苗的苗高及根长生长也存在质量浓度效应的化感抑制作用。其中,不同质量浓度处理中根长的抑制率均明显大于苗高。由此也表明:香豆素对节节麦幼苗根长生长的化感抑制作用大于苗高,这可能与根系最先直接接触并吸收到处理物质有关,只有在根系被抑制达到一定程度后,幼苗的地上部分才会逐渐受到影响。这与宋亮等[24]利用酚酸类物质对苜蓿Medicago sativa的研究结论相符。
相对电导率和丙二醛是植物质膜受伤害程度的重要反映指标。本研究中,相对电导率及丙二醛质量摩尔浓度均随处理时间的延长呈逐渐增加的变化。其中,相对电导率的增加表明香豆素对节节麦幼苗膜结构造成一定破坏,导致电解质外渗的增大;而丙二醛质量摩尔浓度的增加则表明膜脂过氧化作用的逐渐增大。此外,50 mg·L-1溶液处理中,至第10天时相对电导率及丙二醛质量摩尔浓度较第5天时增加均不显著。这可能是香豆素质量浓度低且处理时间短,节节麦通过自身的调节机制,对香豆素造成膜系统损伤进行了积极修复的结果。
渗透调节物质含量的增加是植物对逆境胁迫的重要生理反映,而游离脯氨酸是植物体内重要的渗透调节物质之一。本研究中,随时间的延长,不同溶液处理中的脯氨酸质量分数均显著增加,这是节节麦通过脯氨酸质量分数增加以适应香豆素胁迫的结果,这也与香豆素处理苏丹草[6]、多花黑麦草[7]可溶性糖含量增加的结论相符。而150 mg·L-1处理中,至第15天时较第10天时增加不显著,这可能与香豆素溶液的过度胁迫所致。超氧化物歧化酶是植物体内重要的保护酶,其活性大小是植物抗逆性强弱的重要体现。本研究中,随时间的延长,超氧化物歧化酶活性呈持续增加的变化,表明节节麦启动抗氧化系统以应对香豆素处理造成的其体内活性氧自由基的逐渐增加,这也是节节麦为适应香豆素胁迫而产生的应激反应。150 mg·L-1处理中,至第15天时较第5天时增加显著,但较第10天时略有下降。这表明随着香豆素胁迫的进一步加剧,幼苗体内活性氧自由基的增加速率逐渐超过了超氧化物歧化酶的清除能力,造成自由基不断积累,最终导致超氧化物歧化酶活性的下降,这也与SCANDALIOS[25]及YE等[26]研究结论一致。
Seed germination and seedling growth of invasive Aegilops tauschii with application of coumarin
-
摘要: 为探讨利用化感物质香豆素对入侵植物节节麦Aegilops tauschii进行生物防控的可行性,采用培养皿生物检测法,研究了50,100,150 mg·L-1等3种质量浓度香豆素溶液对其种子萌发及幼苗生长和生理生化指标的影响。结果表明:随质量浓度的增加,香豆素溶液显著降低了节节麦种子发芽率、发芽指数,以及幼苗苗高和根长的生长。从抑制率(RI)可知,香豆素溶液对节节麦种子萌发及幼苗生长存在质量浓度效应的抑制作用。此外,不同质量浓度香豆素溶液处理对根长的RI值均明显大于苗高,表明香豆素对节节麦幼苗根长生长的抑制作用大于苗高。幼苗生长试验中,仅50 mg·L-1溶液处理中,至第10天时相对电导率及丙二醛质量摩尔浓度较第5天时增加未达到显著水平,这可能是节节麦通过自身调节机制对香豆素造成的损伤进行积极修复的结果。而150 mg·L-1溶液处理中,至第15天时脯氨酸质量分数较第10天时增加不显著,而超氧化物歧化酶活性则呈下降趋势。表明香豆素150 mg·L-1溶液处理第15天已超出节节麦的适应承受范围。以上结果显示:150 mg·L-1香豆素溶液对节节麦种子萌发具有较强的化感抑制作用,并通过影响植株抗氧化酶系统、细胞膜透性及膜脂过氧化作用等途径调节节节麦的生长。Abstract: To determine the allelopathic inhibition of coumarin on Aegilops tauschii and the feasibility of using coumarin as a biological method for invasive plant control, the effects of coumarin with different concentrations (50, 100, 150 mg·L-1) on A. tauschii seed germination and seedling growth were evaluated in a sand-culture experiment. Results showed that compared with a control, the germination rate, germination index, growth of seedling height, and root length were inhibited with increase of coumarin solution treatment. The inhibitory effect of coumarin on the growth of root length was greater than that of seedling height based on the value of the inhibition rate. Only with the 50 mg·L-1 coumarin solution treatment did the relative conductivity and MDA content not increase significantly at 10 d compared with at 5 d (P < 0.05). With the 150 mg·L-1 coumarin solution treatment, the proline content did not increase significantly, and the superoxide dismutase (SOD) activity decreased at 15 d compared with at 10 d. The above results indicated that 150 mg·L-1 coumarin solution had a strong allelopathic and inhibitory effect on seed germination of A. tauschii and could regulate seedling growth of A. tauschii by changing antioxidant enzyme systems, cell membrane permeability, and membrane lipid peroxidation.
-
Key words:
- botany /
- coumarin /
- Aegilops tauschii /
- germination rate /
- SOD activity
-
图 1 不同质量浓度香豆素对节节麦幼苗相对电导率及丙二醛质量摩尔浓度的影响
Figure 1 Relative electric conductivity and MDA content of Aegilops tauschii seedling under different concentrations of coumarin solution
图 2 不同质量浓度香豆素对节节麦幼苗脯氨酸质量分数及超氧化物歧化酶活性的影响
Figure 2 Proline content and SOD activity of Aegilops tauschii seedling under different concentrations of coumarin solution
表 1 香豆素溶液对节节麦种子萌发和幼苗生长过程中生长指标的影响
Table 1. Coumarin solution influence of Aegilops tauschii growth indexes on seed germination and growth of seedlings
ρ/(mg·L-1) 发芽率/% 发芽率抑制率 发芽指数 发芽指数抑制率 苗高/cm 苗高抑制率 根长/cm 根长抑制率 0(对照) 94.00 ± 0.11 a 0.00 33.73 ± 0.87 a 0.00 16.52 ± 0.58 a 0.00 13.62 ± 0.76 a 0.00 50 71.60 ± 2.19 b 0.24 22.84 ± 1.10 b 0.32 14.05 ± 0.41 ab 0.14 9.87 ± 0.14 b 0.49 100 45.60 ± 1.95 c 0.51 14.38 ± 1.42 c 0.57 10.15 ± 0.65 c 0.38 7.44 ± 0.31 c 0.60 150 31.60 ± 0.99 d 0.66 7.59 ± 1.19 d 0.77 7.16 ± 0.16 d 0.57 4.81 ± 0.11 d 0.65 说明:同列不同小写字母表示不同处理间差异显著  下载: 导出CSV
下载: 导出CSV
-
[1] 王建花, 陈婷, 林文雄.植物化感作用类型及其在农业中的应用[J].中国生态农业学报, 2013, 21(10):1173-1183. WANG Jianhua, CHEN Ting, LIN Wenxiong. Plant allelopathy types and their application in agriculture[J]. Chin J Eco-Agric, 2013, 21(10):1173-1183. [2] DUKE S O. Allelopathy:current status of research and future of the discipline:a commentary[J]. Allelopathy J, 2010, 25(1):17-30. [3] 邬彩霞, 刘苏娇, 赵国琦.黄花草木樨水浸提液中潜在化感物质的分离、鉴定[J].草业学报, 2014, 23(5):184-192. WU Caixia, LIU Sujiao, ZHAO Guoqi. Isolation and identification of the potential allelochemicals in the aqueous extract of yellow sweet clover (Melilotus officinalis)[J]. Acta Pratac Sin, 2014, 23(5):184-192. [4] 邬彩霞, 刘苏娇, 赵国琦, 等.黄花草木樨对杂草的化感作用研究[J].草地学报, 2015, 23(1):82-88. WU Caixia, LIU Sujiao, ZHAO Guoqi, et al. The allelopathy of yellow sweet of clover on weeds[J]. Acta Agrestia Sin, 23(1):82-88. [5] 邬彩霞, 赵国琦, 刘苏娇, 等.黄花草木樨水浸液中香豆素的含量及其对7种植物种子萌发和幼苗生长的影响[J].草业科学, 2014, 31(12):2262-2269. WU Caixia, ZHAO Guoqi, LIU Sujiao, et al. The content and effects of coumarin in the aqueous extract of yellow sweet clover[J]. Pratac Sci, 2014, 31(12):2262-2269. [6] 王婧怡, 姚丹丹, 徐军, 等.香豆素对苏丹草种子萌发和幼苗生长的影响[J].草业科学, 2017, 34(11):2279-2288. WANG Jingyi, YAO Dandan, XU Jun, et al. Effect of coumarin on Soughum sudanense seed germination and seedling growth[J]. Pratac Sci, 2017, 34(11):2279-2288. [7] 姚丹丹, 王婧怡, 周倩, 等.香豆素对多花黑麦草种子萌发和幼苗生长化感作用的机理研究[J].草业学报, 2017, 26(2):136-145. YAO Dandan, WANG Jingyi, ZHOU Qian, et al. Effect of coumarin on Italian ryegrass seed germination and seedling growth[J]. Acta Pratac Sin, 2017, 26(2):136-145. [8] 覃逸明, 聂刘旺, 黄雨清, 等.凤丹(Paeonia ostii T.)自毒物质的检测及其作用机制[J].生态学报, 2009, 29(3):1153-1161. QIN Yiming, NIE Liuwang, HUANG Yuqing, et al. Detection of Paeonia ostii autotoxins and their mechanism[J]. Acta Ecol Sin, 2009, 29(3):1153-1161. [9] CHON S U, CHOI S K, JUNG S, et al. Effects of alfalfa leaf extracts and phenolic allelochemicals on early seedling growth and root morphology of alfalfa and barnyard grass[J]. Crop Prot, 2002, 21(10):1077-1082. [10] RAZAVI S M, ZARRINI G, ZAHRI S. Biological activity of Prangos uloptera DC. roots, a medicinal plant from Iran[J]. Nat Prod Res, 2010, 24(9):797-803. [11] 王晓阳.节节麦(Aegilops tauschii Coss)生物学特性和遗传多样性[D].北京: 中国农业科学院, 2017. WANG Xiaoyang. The Biological Characteristics and Genetic Diversity of Aegilops tauschii Coss.[D]. Beijing: Chinese Academy of Agricultural Sciences, 2017. [12] 房锋, 张朝贤, 黄红娟, 等.麦田节节麦发生动态及其对小麦产量的影响[J].生态学报, 2014, 34(14):3917-3923. FANG Feng, ZHANG Chaoxian, HUANG Hongjuan, et al. The occurrence of Tausch'sgoatgrass(Aegilops tauschii Coss.)in wheat fields and its effect on wheat yield[J]. Acta Ecol Sin, 2014, 34(14):3917-3923. [13] 房锋, 高兴祥, 魏守辉, 等.麦田恶性杂草节节麦在中国的发生发展[J].草业学报, 2015, 24(2):194-201. FANG Feng, GAO Xingxiang, WEI Shouhui, et al. Occurrence and effects of Aegilops tauschii in China[J]. Acta Pratac Sin, 2015, 24(2):194-201. [14] 段美生, 杨宽林, 李香菊, 等.河北省南部小麦田节节麦发生特点及综合防除措施研究[J].河北农业科学, 2005, 9(1):72-74. DUAN Meisheng, YANG Kuanlin, LI Xiangju, et al. Studies on characteristic of Aegilops squarrosa occurrence and integrated control approaches in winter wheat in the South of Hebei Province[J]. J Hebei Agric Sci, 2005, 9(1):72-74. [15] 王宁, 袁美丽, 陈浩.小麦水浸提液对节节麦种子萌发和幼苗生长的影响[J].浙江农林大学学报, 2018, 35(1):112-120. WANG Ning, YUAN Meili, CHEN Hao. Seed germination and seedling growth of Aegilops tauschii with wheat extracts[J]. J Zhejiang A&F Univ, 2018, 35(1):112-120. [16] 隋标峰.节节麦(Aegilops tauschii Coss.)不同种群对甲基二磺隆的敏感性差异研究[D].北京: 中国农业科学院, 2010. SUI Biaofeng. Basis of Variable Sensitivity to Mesosulfuron-methyl in Different Tausch's goatgrass (Aegilops tauschii Coss.)Population[D]. Beijing: Chinese Academy of Agricultural Sciences, 2010. [17] 谢艳红.不同小麦对甲基二磺隆耐药性及安全剂对耐药性的影响[D].北京: 中国农业大学, 2004. XIE Yanhong. Effects of Different Wheat on Mesosulfuron-resistance and Safenerson Drug Resistance[D]. Beijing: China Agricultural University, 2004. [18] 李美娜, 李志华.紫花苜蓿茎叶浸提液对波斯婆婆纳化感效应的研究[J].中国草地学报, 2010, 32(1):69-74. LI Meina, LI Zhihua. Allelopathic effect of aqueous extracts from stems and leaves of alfalfa on persian speedwell[J]. Chin J Grassl, 2010, 32(1):69-74. [19] WILLIAMSON G B, RICHARDSON D. Bioassays for allelopathy:measuring treatment responses with independent controls[J]. J Chem Ecol, 1988, 14(1):181-187. [20] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社, 2000. [21] 邹琦.植物生理生化实验指导[M].北京:中国农业出版社, 2000. [22] 黄洪武, 李俊, 董立尧, 等.加拿大一枝黄花对植物化感作用的研究[J].南京农业大学学报, 2009, 32(1):48-54. HUANG Hongwu, LI Jun, DONG Liyao, et al. The study on allelopathy of Solidago canadensis L. to several plants[J]. J Nanjing Agric Univ, 2009, 32(1):48-54. [23] 皇甫超河, 陈冬青, 王楠楠, 等.外来入侵植物黄顶菊与四种牧草间化感互作[J].草业学报, 2010, 19(4):22-32. HUANGPU Chaohe, CHEN Dongqing, WANG Nannan, et al. The mutual allelopathic effect between invasive plant Flaveria bidentis and four forage species[J]. Acta Pratac Sin, 2010, 19(4):22-32. [24] 宋亮, 潘开文, 王进闯, 等.酚酸类物质对苜蓿种子萌发及抗氧化物酶活性的影响[J].生态学报, 2006, 26(10):3393-3403. SONG Liang, PAN Kaiwen, WANG Jinchuang, et al. Effect of phenolic acids on seed germination and seedling antioxidant enzyme activity of alfalfa[J]. Acta Ecol Sin, 2006, 26(10):3393-3403. [25] SCANDALIOS L G. Oxygen stress and superoxide dismutase[J]. Plant Physiol, 1993, 101(1):7-12. [26] YE Shufeng, ZHOU Yanhong, SUN Yan, et al. Cinnamic acid causes oxidative stress in cucumber roots, and promotes incidence of Fusarium wilt[J]. Environ Exp Bot, 2006, 56(3):255-262. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.04.025
 点击查看大图
点击查看大图

计量
- 文章访问数: 3248
- HTML全文浏览量: 743
- PDF下载量: 42
- 被引次数: 0
