






-
常绿阔叶林是亚热带地区的地带性植被,在调节气候、涵养水源和保持水土等发挥重要作用[1]。树木对太阳辐射和降水的利用受限于林冠的结构和空间分布特征[2]。林冠结构受生物环境和非生物环境的综合影响[3]。林冠层是次生常绿阔叶林与外界联系最直接与最活跃的界面[4]。林冠结构、物种组成、立地条件、季节变化[5]以及间伐[6]等都会改变林下光环境。林冠通过对光照的吸收、反射和散射改变林下光照条件[7],进而影响林下植被的种子萌发[8]、林下更新[9]、物种组成和分布[10-11]、种群特征[12]以及多样性[13]等。研究者根据森林群落类型[13]、林冠开度[14]、光照总立地因子[15]等单一因子划分森林群落类群。综合采用林冠结构多个指标,通过聚类分析划分类群开展次生常绿阔叶林林冠结构与林下植被的关系还相对缺乏研究。珠江三角洲城市如广州、东莞、佛山等9市已建成国家森林城市,粤港澳大湾区为国家森林城市群的建设和发展带来新的发展机遇。本研究以珠江三角洲城郊次生常绿阔叶林为研究对象,试图回答以下问题:林冠结构、林下光环境以及林下植被的数量特征沿林冠梯度变化是否一致?不同林冠结构及林下光环境对林下植被物种分布的影响是否有差异?通过对以上问题的探讨,为珠江三角洲国家森林城市群群落构建及林下植被物种选择提供科学依据。
-
试验监测样地位于广州市和东莞市城郊。该区域位于广东省东南部珠江三角洲核心地带。地理位置22°51′29″~23°38′21″N,113°27′36″~113°56′59″E,属南亚热带季风气候区,气候温和,雨量充沛,年平均气温20.5~22.5 ℃,年平均降水量1 100.0~2 200.0 mm,年平均相对湿度71%~79%。地带性土壤为赤红壤,主要土壤类型还有山地赤红壤、山地黄壤和耕作土等。地带性植被为常绿阔叶林,但该区南亚热带原生常绿阔叶林消失殆尽,城郊仅存少量次生常绿阔叶林,人工林较常见。
-
在线路踏查的基础上,选取广州帽峰山森林公园、广州石门国家森林公园和东莞大岭山森林公园典型次生常绿阔叶林,作为珠江三角洲城郊次生常绿阔叶林群落调查样地。3个样地的土壤、地形等因素尽量保持一致。每个样地分别设置10个20 m × 20 m的样方开展植被调查,调查面积共计1.2 hm2。对每个样方进行每木调查,测定胸径(DBH)≥ 3 cm的所有立木的种名、胸径、树高和冠幅。在每个样方内按顺时针方向选取4个面积为5 m × 5 m的小样方,记录灌木的种名、高度与盖度;在每个样方的四角和对角线中心各设置1个面积为1 m × 1 m的小样方,记录草本植物的种名、高度与盖度。
-
选择阴天或晴好天气的日出或日落时刻,将Nikon CoolPix D750数码相机外接Nikkor AF-S 8-15/3.5-4.5 E鱼眼镜头转换器用三脚架水平放置于离地面1.65 m处,用指南针确定方向使记录的照片顶部与磁北方向重合,在每个20 m × 20 m样方四角和对角线四分位处镜头朝上拍摄半球面林冠影像,共计拍摄150张半球面林冠影像。采用Gap Light Analyzer 2.0(GLA)图像处理软件分析林冠影像,输出林冠开度(canopy openness),叶面积指数(leaf area index),林下直射光(transmitted direct light),林下散射光(transmitted diffused light)和林下总光照(transmitted total light)。
-
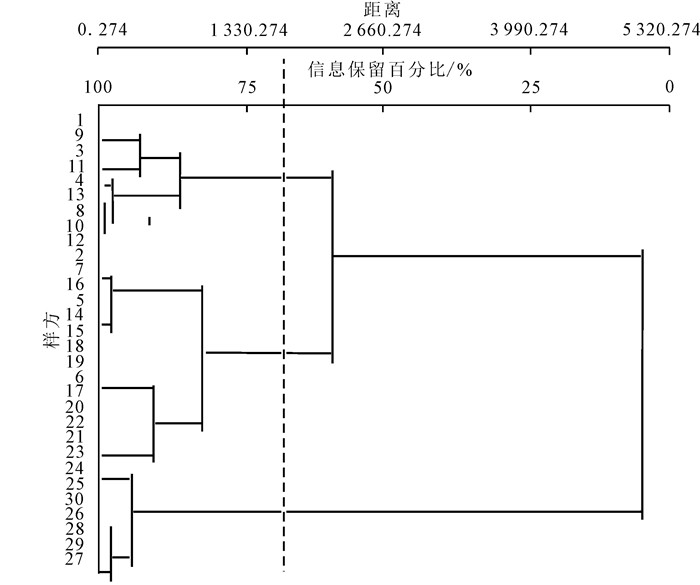
综合采用次生常绿阔叶林群落样方立木多度、胸径、树高、林冠开度和叶面积指数矩阵信息,采用Euclidean距离系数和组平均聚类法进行林冠梯度聚类分析。当信息保留70%左右时,组平均聚类分析将次生常绿阔叶林30个样方分为3个林冠梯度(图 1)。林冠梯度1的12个样方主要分布在广州帽峰山森林公园,林冠梯度2的10个样方主要分布在广州石门国家森林公园,林冠梯度3的8个样方主要分布在东莞大岭山森林公园。

图 1 次生常绿阔叶林群落组平均聚类图
Figure 1. Group average clustering diagram of secondary evergreen broadleaved forest communities
-
对次生常绿阔叶林群落林冠结构与林下植被数量特征分别进行Kruskal-Wallis test(非参数检验),并对差异显著的结果进一步做多重比较。
-
针对次生常绿阔叶林群落的林冠结构、林下光环境与林下植被盖度的关系开展指示种分析,将P<0.05的物种作为该林冠结构和林下光环境参数的指示种。
-
利用林下植被样方物种盖度与林冠结构矩阵进行非度量的多维标定法,分析次生常绿阔叶林群落林下物种与林冠结构的关系。
以上数据分析方法在软件PC-ORD(6.0),Statistica(8.0)和Canoco(4.5)中完成。
-
基于1.2 hm2调查面积,南亚热带城郊次生常绿阔叶林群落林冠层立木共有立木3 198株,胸径10 cm左右,树高约7 m。
非参数检验(Kruskal-Wallis test)揭示,次生常绿阔叶林林冠结构和林下光环境各参数沿林冠梯度变化表现不一。平均树高、林冠开度和叶面积指数沿林冠梯度差异不显著(P>0.05),立木密度和平均胸径沿林冠梯度差异极显著(P<0.001),林下直射光、林下散射光和林下总光照沿林冠梯度差异显著(P<0.05)。多重比较进一步表明,3个林冠梯度的立木密度差异显著。平均胸径、林下直射光、林下散射光和林下总光照在林冠梯度1和林冠梯度2间差异显著,但在林冠梯度2和林冠梯度3之间差异不显著(表 1)。
表 1 次生常绿阔叶林林冠结构和林下光环境特征
Table 1. Canopy structure andunderstory light conditions of secondary evergreen broadleaved forest communities
林冠梯度 立木密度/(株·hm-2) 平均胸径/cm 平均树高/m 林冠开度 叶面积指数 林下直射光/% 林下散射光/% 林下总光照/% 1 1 615 ± 122 c 12.7 ± 1.1 a 7.6 ± 0.4 a 12.16 ± 0.43 a 2.37 ± 0.04 a 15.88 ± 0.77 b 13.84 ± 0.37 b 14.86 ± 0.53 b 2 2 683 ± 111 b 9.7 ± 0.4 b 7.4 ± 0.3 a 12.98 ± 0.75 a 2.24 ± 0.07 a 18.99 ± 0.81 a 15.82 ± 0.60 a 17.40 ± 0.67 a 3 3 950 ± 62 a 7.5 ± 0.3 b 6.6 ± 0.3 a 11.57 ± 0.28 a 2.30 ± 0.04 a 17.26 ± 0.66 ab 14.87 ± 0.44 ab 16.07 ± 0.51 ab -
次生常绿阔叶林林下植被物种丰富,物种数、平均树高和总盖度沿林冠梯度差异显著(P<0.05)。基于1.2 hm2样地,群落整体共有林下植被224种,分属于140属82科,群落平均高度85 cm左右。林冠梯度2的林下植被物种最多。但群落平均高度和总盖度皆以林冠梯度3最大,其次为林冠梯度2,最小的是林冠梯度3。非参数检验(Kruskal-Wallis test)揭示,林下植被数量特征沿林冠梯度差异显著(P<0.05)。多重比较进一步揭示,林冠梯度3的物种数和平均树高与林冠梯度1及林冠梯度2有显著差异,但林冠梯度3的总盖度只与林冠梯度2有差异,与林冠梯度1差异不显著(图 2)。芒萁Dicranopteris dichotoma,粗叶榕Ficus hirta和九节Psychotria rubra等是林冠梯度1的优势种,鸭公树Neolitsea chuii,大叶鼠刺Itea macrophylla和扇叶铁线蕨Adiantum flabellulatum等是林冠梯度2的优势种,华山姜Alpinia oblongifolia,密花树Rapanea neriifolia和九节等是林冠梯度3的优势种。

图 2 林下植被数量特征沿林冠梯度的分布
Figure 2. Quantitative features of understory vegetation along the canopy gradient
-
次生常绿阔叶林群落林下植被对不同林冠结构参数及林下光环境的响应不一。立木多度、胸径、林冠开度和叶面积指数的林下植被指示种分析P皆大于0.05,未能筛选出指示种。而立木平均高度、林下直射光、林下散射光和林下总光照分别有2~5种指示种(表 2)。
表 2 次生常绿阔叶林林冠结构及林下光照的指示种
Table 2. Indicator species of canopy structure and transmitted lightof secondary evergreen broadleaved forest communities
林冠结构 指示种 总盖度/% 出现样方数 生长型 指示值 P 立木平均高度 黄杞Engelhardtia roxburghiana 26 6 乔木 75.0 0.012 8 竹叶木姜子Litsea pseudoelongata 6 1 乔木 46.2 0.044 6 林下直射光 水锦树Wendlandia uvariifolia 25 2 乔木 66.7 0.024 4 绿冬青Ilex viridis 42 5 灌木 57.6 0.035 8 异形南五味子Kadsura heteroclite 11 3 藤本 58.8 0.041 6 林下散射光 冬青Ilex chinensis 48 4 灌木 62.6 0.011 0 虎舌红Ardisia mamillata 4 2 灌木 50 0.029 4 谷木Memecylon ligustrifolium 2 2 灌木 50 0.029 4 友水龙骨Polypodiodes amoena 10 2 蕨类 50 0.029 4 林下总光照 绿冬青Ilex viridis 42 5 灌木 94.1 0.005 6 水锦树Wendlandia uvariifolia 25 2 乔木 100 0.009 6 华南紫萁Osmunda vachellii 13 3 蕨类 82.8 0.014 0 金樱子Rosa laevigata 14 3 乔木 71.4 0.024 8 土沉香Aquilaria sinensis 21 4 乔木 59.3 0.048 6 立木平均高度的指示种为黄杞(乔木)和竹叶木姜子(乔木)。其中,黄杞出现的样方数较多,指示性较强。林下直射光的指示种为水锦树(乔木)、绿冬青(灌木)和异形南五味子(木质藤本)。林下散射光的指示种为灌木类的冬青、虎舌红和谷木,还有蕨类植物友水龙骨。林下总光照的指示种较多,除了同为林下直射光指示种的水锦树和绿冬青外,还有华南紫萁(蕨类)、金樱子(乔木)和土沉香(乔木)。其中,水锦树的指示性最强。
-
采用典范对应分析对林下植被盖度与林冠结构及林下光环境的关系进行分析(表 3),4个排序轴的特征值分别为0.434,0.214,0.055和0.041。前3个排序轴的累积贡献率达88.5%,特别是第1排序轴已经解释了林下植被物种与环境之间关系的54.6%,可以比较好地分析出林冠结构中物种分布起主要作用的指标。同时,蒙特卡罗检验结果P<0.05,说明排序结果可信。
表 3 各排序轴的加权相关矩阵
Table 3. Weighted correlation matrix of CCA ordination axles
参数 特征值 物种-环境关系 物种数据变量累积百分比 物种-环境关系变量累积百分比 所有特征值之和 所有典范特征值之和 第一典范轴P值 排序轴1 0.434 0.904 24.0 54.6 1.804 0.794 0.001 排序轴2 0.214 0.702 35.9 81.5 排序轴3 0.055 0.738 38.9 88.5 排序轴4 0.041 0.689 41.2 93.6 分析结果表明:林冠结构各项指标与物种轴和环境轴的第一、二排序轴均表现出较大相关性,其中立木多度的相关性最大,其次是平均树高和平均胸径,林下光环境各参数与轴1的相关性也较强。不同林冠梯度的样方集中分布在不同象限(图 3)。林冠梯度1的样方与林下光环境和林冠开度关系较密切,林冠梯度2的样方与平均树高和平均胸径相关性较强,林冠梯度3的样方与立木多度相关性较强。扇叶铁线蕨Adiantum flabellulatum,乌毛蕨Blechnum orientale,芒萁Dicranopteris dichotoma,狗脊Woodwardia japonica等蕨类植物均分布在林下光环境较弱的区域,油茶Camellia oleifera和锡叶藤Tetracera sarmentosa等喜阳植物则分布在林下光环境较强的区域,而木荷Schima superba和腺点紫金牛Ardisia lindleyana等中性植物受林冠层立木的高度和胸径的影响更大。

图 3 林下植被与林冠结构各参数的典范对应分析
Figure 3. Canonical correspondence analysis of under story vegetation and canopy structure parameters
-
不同季节林下光环境差异明显[16],林冠不同建群种对林下草本植物多样性影响不同[17],中、下层林冠对林下更新植物多样性的作用更明显[9]。不同人工林林分的冠层结构和林下光照指标间差异显著[18]。从落叶阔叶林到常绿落叶阔叶混交林再到常绿阔叶林,不同森林群落类型的林冠结构发生了显著变化,林冠开度降低,叶面积指数增加,平均叶倾角变小,林冠对光的截获能力和消光能力增强,林下光照强度降低[12]。本研究组平均聚类分析将次生常绿阔叶林30个样方分为3个林冠梯度,分别分布在不同的3个地点。究其原因,可能是由于珠江三角洲城郊这3个地点的次生常绿阔叶林立木密度和平均胸径差异极显著(P<0.001),林下光环境差异也显著(P<0.05),因而可明确划分出不同的林冠梯度。这还表明,除了林冠开度和叶面积指数,立木密度和平均胸径也是较好的林冠结构指标。
-
林冠结构影响林下草本层的结构和多样性,混交林的冠层斑块对于保护林下植被的自然分布格局很重要[13],常绿林冠和落叶林冠对林下灌木的影响不一致[11]。本研究也表明:次生常绿阔叶林群落不同林冠结构参数对林下植被的指示作用不同。林下植被物种分布与立木多度、平均树高和平均胸径相关性较强,不同林冠梯度林下植被物种组成和分布都有明显差异。无论是资源数量还是环境异质性都不能单独影响物种多样性,但它们通过森林生态系统林分发展和干扰影响林下植被多样性[19]。冠层结构所形成的光环境强烈影响着林下植被的分布。林冠开度和林下总光照对木本植物的影响最大[20]。草本植物覆盖度与漫射光和基质显著相关[21]。本研究对次生常绿阔叶林林冠结构的指示种分析也揭示,14个指示种中有5种灌木、6种乔木、2种蕨类和1种藤本。不同林冠梯度下的林下植被物种组成因林下光照强弱而不同。林冠结构以及林下光环境共同影响林下植被的物种分布,但对次生常绿阔叶林木本植物影响最大的林下光环境指标是林下总光照和林下散射光。
Influence of canopy structure on understory vegetation of secondary evergreen broadleaf forest communities
-
摘要: 采用经典群落学调查方法和半球面影像技术,以珠江三角洲城郊次生常绿阔叶林群落为研究对象,运用聚类分析、非参数检验(Kruskal-Wallis test),指示种分析(ISA)和典范对应分析(CCA)等方法,定量研究次生常绿阔叶林群落林冠结构对林下植被的影响。主要结果如下:①次生常绿阔叶林林冠结构和林下光环境各参数沿林冠梯度变化表现不一。立木密度和平均胸径沿林冠梯度差异极显著(P < 0.001),林下直射光、林下散射光和林下总光照沿林冠梯度差异显著(P < 0.05)。②林下植被物种丰富,物种数、平均树高和总盖度沿林冠梯度差异显著(P < 0.05)。林冠梯度2的林下植被物种最多,但群落平均高度和总盖度皆以林冠梯度3最大。③林下植物对不同林冠结构和林下光照参数的响应不一。水锦树Wendlandia uvariifolia和绿冬青Ilex viridis同为林下直射光和林下总光照的指示种。④典范对应分析(CCA)揭示,林冠结构以及林下光环境共同影响林下植被的物种分布,扇叶铁线蕨Adiantum flabellulatum,乌毛蕨Blechnum orientale,芒萁Dicranopteris dichotoma和狗脊Woodwardia japonica等蕨类植物均分布在林下光环境较弱的区域,油茶Camellia oleifera和锡叶藤Tetracera sarmentosa等喜阳植物则分布在林下光环境较强的区域,而木荷Schima superba和腺点紫金牛Ardisia lindleyana等中性植物受林冠层立木的高度和胸径的影响更大。不同林冠梯度下的林下植被物种组成因林下光照强弱而不同,对次生常绿阔叶林木本植物影响最大的林下光环境指标是林下总光照和林下散射光。Abstract: Understory vegetation, representing the main component of biodiversity in a forest community and playing a key role in forest functioning and canopy structure, are important characteristics of a forest community. To evaluate the effects of different gradients of canopy structure on understory vegetation, secondary evergreen broadleaf forest communities in the suburbs of the Pearl River Delta were studied. The effect on understory plants was tested using classical community survey methods and hemispherical photography. Analysis included use of Cluster Analysis, Non-parametric Test (Kruskal-Wallis test), Indicator Species Analysis (ISA), and Canonical Correspondence Analysis (CCA). Results showed that (1) Canopy structure and understory light conditions varied along the canopy gradient. The standing density and average diameter at breast height (DBH) showed a highly significant difference (P < 0.001); whereas, the transmitted direct light, transmitted diffused light, and transmitted total light significantly differed along the canopy gradient (P < 0.05). (2) The understory vegetation was rich; and the abundance and average height as well as total coverage, significantly differed with canopy gradient (P < 0.05). Species were richest in canopy gradient 2, but the average height and total coverage were both highest in canopy gradient 3. (3) The responses of understory vegetation to canopy structure and transmitted light varied. Wendlandia uvariifolia and Ilex viridis were both indicator species of transmitted direct light and transmitted total light. (4) The CCA showed that canopy structure and understory light conditions affected the distribution of understory vegetation. Ferns such as Adiantum flabellulatum, Blechnum orientale, Dicranopteris dichotoma, and Woodwardia japonica resided in areas of weak sunlight, but heliophytes like Camellia oleifera and Tetracera sarmentosa inhabited places where they received better light. Moreover, distribution of neutral plants like Schima superba and Ardisia lindleyana were more affected by the height and DBH of standing trees. This study demonstrated that understory vegetation varied according to understory light conditions along the canopy gradient with transmitted total light and transmitted diffused light being the main factors influencing the understory light environment of the secondary evergreen broadleaf forest.
-
图 1 次生常绿阔叶林群落组平均聚类图
Figure 1 Group average clustering diagram of secondary evergreen broadleaved forest communities
图 2 林下植被数量特征沿林冠梯度的分布
Figure 2 Quantitative features of understory vegetation along the canopy gradient
图 3 林下植被与林冠结构各参数的典范对应分析
Figure 3 Canonical correspondence analysis of under story vegetation and canopy structure parameters
表 1 次生常绿阔叶林林冠结构和林下光环境特征
Table 1. Canopy structure andunderstory light conditions of secondary evergreen broadleaved forest communities
林冠梯度 立木密度/(株·hm-2) 平均胸径/cm 平均树高/m 林冠开度 叶面积指数 林下直射光/% 林下散射光/% 林下总光照/% 1 1 615 ± 122 c 12.7 ± 1.1 a 7.6 ± 0.4 a 12.16 ± 0.43 a 2.37 ± 0.04 a 15.88 ± 0.77 b 13.84 ± 0.37 b 14.86 ± 0.53 b 2 2 683 ± 111 b 9.7 ± 0.4 b 7.4 ± 0.3 a 12.98 ± 0.75 a 2.24 ± 0.07 a 18.99 ± 0.81 a 15.82 ± 0.60 a 17.40 ± 0.67 a 3 3 950 ± 62 a 7.5 ± 0.3 b 6.6 ± 0.3 a 11.57 ± 0.28 a 2.30 ± 0.04 a 17.26 ± 0.66 ab 14.87 ± 0.44 ab 16.07 ± 0.51 ab  下载: 导出CSV
下载: 导出CSV
表 2 次生常绿阔叶林林冠结构及林下光照的指示种
Table 2. Indicator species of canopy structure and transmitted lightof secondary evergreen broadleaved forest communities
林冠结构 指示种 总盖度/% 出现样方数 生长型 指示值 P 立木平均高度 黄杞Engelhardtia roxburghiana 26 6 乔木 75.0 0.012 8 竹叶木姜子Litsea pseudoelongata 6 1 乔木 46.2 0.044 6 林下直射光 水锦树Wendlandia uvariifolia 25 2 乔木 66.7 0.024 4 绿冬青Ilex viridis 42 5 灌木 57.6 0.035 8 异形南五味子Kadsura heteroclite 11 3 藤本 58.8 0.041 6 林下散射光 冬青Ilex chinensis 48 4 灌木 62.6 0.011 0 虎舌红Ardisia mamillata 4 2 灌木 50 0.029 4 谷木Memecylon ligustrifolium 2 2 灌木 50 0.029 4 友水龙骨Polypodiodes amoena 10 2 蕨类 50 0.029 4 林下总光照 绿冬青Ilex viridis 42 5 灌木 94.1 0.005 6 水锦树Wendlandia uvariifolia 25 2 乔木 100 0.009 6 华南紫萁Osmunda vachellii 13 3 蕨类 82.8 0.014 0 金樱子Rosa laevigata 14 3 乔木 71.4 0.024 8 土沉香Aquilaria sinensis 21 4 乔木 59.3 0.048 6
下载: 导出CSV
表 3 各排序轴的加权相关矩阵
Table 3. Weighted correlation matrix of CCA ordination axles
参数 特征值 物种-环境关系 物种数据变量累积百分比 物种-环境关系变量累积百分比 所有特征值之和 所有典范特征值之和 第一典范轴P值 排序轴1 0.434 0.904 24.0 54.6 1.804 0.794 0.001 排序轴2 0.214 0.702 35.9 81.5 排序轴3 0.055 0.738 38.9 88.5 排序轴4 0.041 0.689 41.2 93.6
下载: 导出CSV
-
[1] 宋永昌, 王希华, 阎恩荣.中国常绿阔叶林:分类·生态·保育[M].北京:科学出版社, 2013. [2] 郝建, 潘丽琴, 潘启龙, 等.格木人工林不同冠层光合特征[J].浙江农林大学学报, 2017, 34(5):871-877. HAO Jian, PAN Liqin, PAN Qilong, et al. Photosynthetic characteristics in different canopy positions of an Erythrophleum fordii plantation[J]. J Zhejiang A&F Univ, 2017, 34(5):871-877. [3] FOTIS A T, MORIN T H, FAHEY R T, et al. Forest structure in space and time:biotic and abiotic determinants of canopy complexity and their effects on net primary productivity[J]. Agric For Meteorol, 2018, 250/251:181-191. [4] 敬小丽, 张璐, 杜伟静, 等.南岭山地森林群落冠层结构对林下野生花卉的影响[J].西南农业学报, 2015, 28(2):833-838. JING Xiaoli, ZHANG Lu, DU Weijing, et al. Effect of canopy structure on understory wild flower in Nanling mountain forest community of South China[J]. Southwest Chin J Agric Sci, 2015, 28(2):833-838. [5] QUEVEDO A, SCHWARZKOPF T, GARCIA C, et al. Light environment in the understory of an Andean cloud forest:canopy structure and climatic seasonality[J]. Rev Biol Trop, 2016, 64(4):1699-1707. [6] LETTOW M C, BRUDVIG L A, BAHLAI C A, et al. Oak savanna management strategies and their differential effects on vegetative structure, understory light, and flowering forbs[J]. For Ecol Manage, 2014, 329:89-98. [7] BEAUDET M, HARVEY B D, MESSIER C, et al. Managing understory light conditions in boreal mixedwoods through variation in the intensity and spatial pattern of harvest:a modelling approach[J]. For Ecol Manage, 2011, 261(1):84-94. [8] WILLIS J L, WALTERS M B, GOTTSCHALK K W. Scarification and gap size have interacting effects on northern temperate seedling establishment[J]. For Ecol Manage, 2015, 347:237-246. [9] 周晓果, 温远光, 朱宏光, 等.大明山常绿阔叶林冠层垂直结构与林下植物更新[J].应用生态学报, 2017, 28(2):367-374. ZHOU Xiaoguo, WEN Yuanguang, ZHU Hongguang, et al. Canopy vertical structure and understory plant regeneration of an evergreen broadleaved forest in Damingshan, Guangxi, China[J]. Chin J Appl Ecol, 2017, 28(2):367-374. [10] 王永强, 蔡燕茹, 曾焕忱, 等.不同林冠开度下亚热带林下植物的组成和多样性[J].西北农林科技大学学报(自然科学版), 2016, 44(5):64-72. WANG Yongqiang, CAI Yanru, ZENG Huanchen, et al. Composition and diversity of understory plant species in subtropical forests under different canopy openness[J]. J Northwest A&F Univ Nat Sci Ed, 2016, 44(5):64-72. [11] DENNY C K, NIELSEN S E. Spatial heterogeneity of the forest canopy scales with the heterogeneity of an understory shrub based on fractal analysis[J]. Forests, 2017, 8(5):1-18. [12] 黄慧敏, 董蓉, 何丹妮, 等.冠层结构和光环境的时空变化对紫耳箭竹种群特征的影响[J].应用生态学报, 2018, 29(7):2129-2138. HUANG Huimin, DONG Rong, HE Danni, et al. Effects of temporal and spatial variation of canopy structures and light conditions on population characteristics of Fargesia decurvata[J]. Chin J Appl Ecol, 2018, 29(7):2129-2138. [13] MESTRE L, TORO-MANRÍQUEZ M, SOLER R, et al. The influence of canopy-layer composition on understory plant diversity in southern temperate forests[J]. For Ecosyst, 2017, 4(6):1-13. [14] AMADEU L S N, SAMPAIO M B, DOS SANTOS F A M. Influence of light and plant size on the reproduction and growth of small palm tree species:comparing two methods for measuring canopy openness[J]. Am J Bot, 2016, 103(9):1678-1686. [15] 胡砚秋, 柯娴氡, 徐明锋, 等.亚热带森林群落草本植物对林下光环境的响应[J].中南林业科技大学学报, 2016, 36(8):72-76. HU Yanqiu, KE Xiandong, XU Mingfeng, et al. Herbaceous plant in response to understory light regimes ina subtropical forest community[J]. J Cent South Univ For Technol, 2016, 36(8):72-76. [16] CONSTABEL A J, LIEFFERS V J. Seasonal patterns of light transmission through boreal mixedwood canopies[J]. Can J For Res, l996, 26:1008-1014. [17] 刘晓瞳, 戴兴安, 胡婷, 等.基于1公顷样地的上海崇明岛人工林草本植物多样性及其对林冠结构的响应[J].生态学杂志, 2017, 36(6):1564-1569. LIU Xiaotong, DAI Xing'an, HU Ting, et al. Herbaceous plant diversity and its response to canopy structure of tree plantation based on 1 hm2 plot at Chongming Island, Shanghai[J]. Chin J Ecol, 2017, 36(6):1564-1569. [18] 谭一波, 何琴飞, 郑威, 等.珠江流域中上游防护林冠层结构对林下植被的影响[J].生态学杂志, 2016, 35(12):3148-3156. TAN Yibo, HE Qinfei, ZHENG Wei, et al. Effects of canopy structure on understory vegetation in shelterbelt forests along the middle and upper reaches of Pearl River[J]. Chin J Ecol, 2016, 35(12):3148-3156. [19] BARTELS S F, CHEN H Y H. Is understory plant species diversity driven by resource quantity or resource heterogeneity?[J]. Ecology, 2010, 91(7):1931-1938. [20] 孙东, 朱纯, 熊咏梅, 等.林隙光照对广州市风景林林下植物分布的影响[J].华南农业大学学报, 2011, 32(4):63-66. SUN Dong, ZHU Chun, XIONG Yongmei, et al. Effect of gap light factors on understory plant species distribution in urban scenic forests of Guangzhou[J]. J South Chin Agric Univ, 2011, 32(4):63-66. [21] 段文标, 王丽霞, 陈立新, 等.红松阔叶混交林林隙大小及光照对草本植物的影响[J].应用生态学报, 2013, 24(3):614-620. DUAN Wenbiao, WANG Lixia, CHEN Lixin, et al. Effects of forest gap size and light intensity on herbaceous plants in Pinus koraiensis-dominated broadleaved mixed forest[J]. Chin J Appl Ecol, 2013, 24(3):614-620. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.06.012
 点击查看大图
点击查看大图
计量
- 文章访问数: 3497
- HTML全文浏览量: 988
- PDF下载量: 78
- 被引次数: 0
