





-
水稻Oryza sativa是中国主要的粮食作物之一,2017年水稻的栽植面积为3 074.7 万hm2,约占主要粮食作物总面积的26.1%[1],并广泛分布于秦岭淮河以南地区[2]。水稻栽植所处地区常年多云多雨,传统光学被动遥感技术难以获取清晰且连续的时间序列影像。合成孔径雷达(synthetic aperture radar, SAR)是一种微波波段的主动遥感技术,可以穿透云雨,不依赖太阳辐射,全天候全天时成像,可在一定程度上弥补光学遥感的不足,利用SAR遥感进行农作物监测具有较大的潜力[3-8]。目前对作物的非全极化SAR时序监测研究中,普遍依赖于后向散射系数这一极化特征,以及在时间序列上的变化曲线。但是微波对地物含水量的变化十分敏感,降雨会使SAR后向散射系数在时间序列上出现较大波动,导致在对水稻等南方作物的长势进行判断时容易出现较大偏差,而现有研究更多关注植株结构变化对后向散射系数的作用,较少考虑降雨的影响。针对降雨对水稻长势极化特征的影响,研究相应的消减方法,及时、有效、准确地获取多云雨地区水稻的长势信息,对重国粮食生产和粮食安全都具有重要意义。本研究在已有Sentinel-1 SAR影像所具备的同向极化(VV)和交叉极化(VH)后向散射系数基础上,提出归一化差分后向散射指数(normalize difference backscattering index,NDBI)。并对VH、VV、NDBI分别建立后向散射系数时序特征曲线,利用同期降雨数据和光学影像作为辅助资料,探讨水稻在幼苗期、分蘖拔节期、长穗期和结实期4个关键生长期中,植株形态特点,相应的极化特征变化趋势,以及降雨对后向散射系数的影响。
-
研究区位于广西桂平市以东约6.5 km,寻旺乡复兴村正南方(23°24′0″~23°24′40″N,110°7′40″~110°8′30″E)的水稻田地。桂平市地处广西东南部,属南亚热带季风气候,气候温暖,雨量充沛,雨热同期,适合水稻生长。2017年桂平市水稻产量达47.9 万t,并连续5 a居广西各县(区)第1位[9],是广西水稻栽植大县。研究区内所栽植的水稻品种为‘百香139’‘Bailixiang 139’,属于感温型常规水稻,适宜在桂南、桂中稻作区作早、晚稻栽植。早稻全生育期约124 d;晚稻全生育期约104 d[10]。本研究将对该研究区内2018年的早稻和晚稻的长势进行监测。
-
选用SAR影像数据来源于Sentinel-1 C波段SAR卫星,该卫星在默认TOPS(Terrain Observation with Progressive Scan)成像模式下的重访周期为12 d,地理编码后的影像空间分辨率为10 m,是具有较高空间和时间分辨率的极轨卫星[11]。所采用的影像数据产品为单视复数影像(single look complex,SLC),具备VV和VH双极化方式。水稻的生长周期较短,一般为3~4个月,在此期间该卫星可拍摄8~12期影像数据,相较于常规光学卫星影像数据,能够形成更为连续且密集的影像序列。本研究使用2018年3−11月共23期SAR影像对研究区内早、晚2季水稻的长势极化特征进行研究。
选用的辅助验证数据为Planet光学卫星在线高分辨率影像数据集,空间分辨率为3~5 m,可清晰分辨研究区内稻田的轮廓和内部作物的长势。Planet卫星数据由一系列微小卫星群组成,重访周期可缩短到1 d[12]。但是卫星群在过境研究区对其进行拍摄时,多数情况下仍会受云雨遮挡,成像质量较差或无法成像,因此采用1个月合成影像数据对水稻的长势做验证。
降雨数据采用日本宇宙航空研究开发机构(Japan Aerospace Exploration Agency,JAXA)提供的Global Rainfall Watch观测数据。该数据由全球降水测量任务(Global Precipitation Measurement Mission,GPM)中的核心卫星和星座卫星,3 h观测1次全球降水产生。表1记录了Sentinel-1卫星过境当天、前1日、前2日研究区最大降雨量。
表 1 Sentinel-1卫星成像时刻雨量
Table 1. Rainfall of Sentinel-1 satellite imaging day
成像日期(年-月-日) 降雨量/mm 当天 前1日 前2日 2018-05-16 1.34 2018-05-28 0.08 5.51 2018-06-21 5.91 2018-07-15 1.33 2018-07-27 3.76 2018-08-20 5.87 2018-09-01 3.02 -
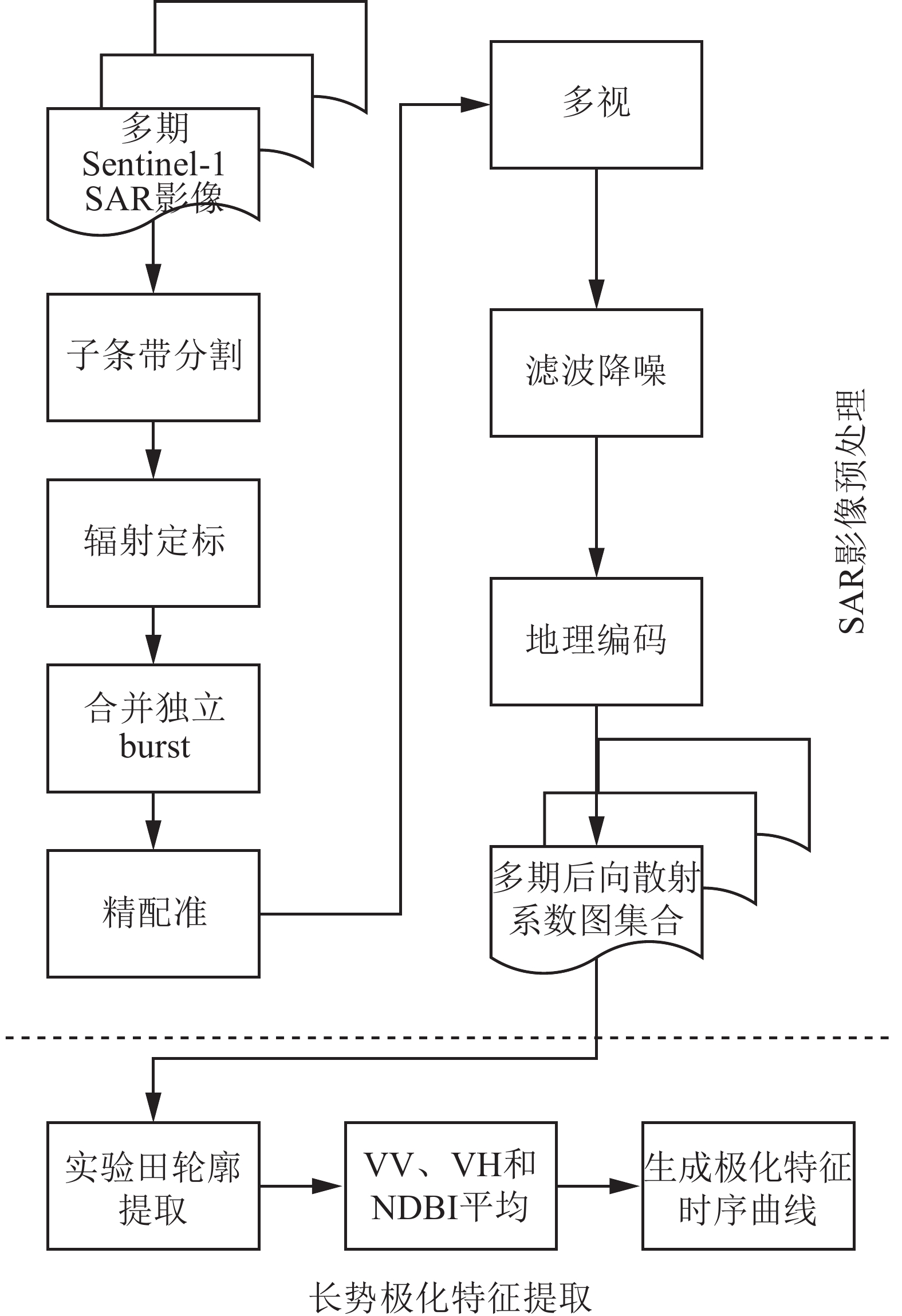
对Sentinel-1 SAR影像数据经过一系列的预处理,才能获取时间序列上的水稻长势极化特征。具体处理流程如图1所示,分为SAR影像预处理和长势极化特征提取两部分。
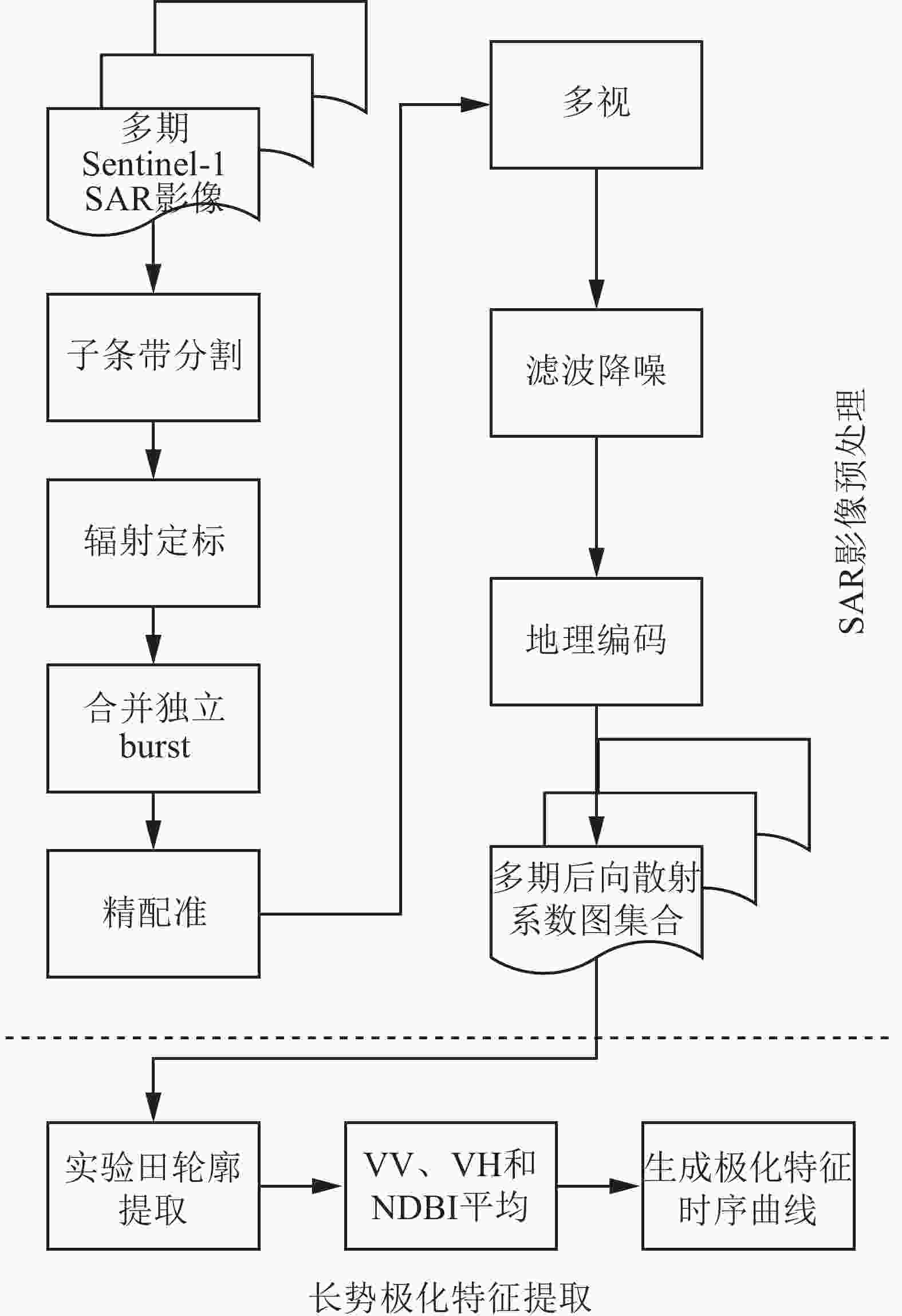
图 1 数据处理流程
Figure 1. Data processing flow
-
影像预处理主要有2个目的:①对影像进行辐射定标,从而确定影像的灰度与标准雷达散射截面的关系。影像经辐射定标后,每个像素都能表示唯一确定的地物后向散射信息[13],这也是本研究开展对不同极化方式、不同时相下记录的水稻极化信息进行定量分析和计算的前提。②对SAR影像进行地理编码,赋予地理坐标。SAR影像的X轴和Y轴分别与成像时刻传感器到地物的斜距距离以及传感器运动矢量相关,并按照距离-方位坐标系(也称SAR坐标系)而非真实地理坐标记录地物位置。因此,需要进行地理编码,将SAR坐标系转为地理坐标系,使处理的结果影像能与其他空间数据进行叠加分析。影像预处理后具有地理坐标,并获得经过精确配准后的多时期后向散射系数图集合。在该图集合中,每个坐标点都包含该处地物的后向散射系数随时间变化形成的序列。将基于该图集合开展对水稻长势的分析。
-
将针对VV和VH 2种极化成像方式在水稻不同生长阶段下产生的后向散射系数进行研究,并认为在同一拍摄时间中,同一稻田内部作物的分布和长势是一致的。因此,对于水稻长势极化特征的提取,是对每期影像中的单块稻田内部所有像素的后向散射系数取平均值,并根据时间顺序排列形成序列。
-
已有研究成果表明[5,8]:降雨会对各极化方式下的后向散射系数均产生较大影响,反映在时序曲线上则是形成较大波动,且上升和下降趋势基本相同。根据这一规律,在研究了归一化植被指数(NDVI)和归一化水指数(NDWI)[14]的基础上,提出归一化差分后向散射指数(NDBI)计算方法,并基于NDBI建立极化特征曲线。
$${I_{{\rm{NDB}}}} = \frac{{{C_{{\rm{VH}}}} - {C_{{\rm{VV}}}}}}{{{C_{{\rm{VH}}}} + {C_{{\rm{VV}}}}}}\text{。}$$ 式(1)中:INDB表示归一化差分后向散射指数,为无量纲量,CVH和CVV分别表示Sentinel-1影像数据中交叉极化和同化极化方式的后向散射系数。引入NDBI在于强化VH和VV的差异,弱化两者的共有趋势,从而在一定程度上抵消降雨对单一后向散射系数时序曲线产生的波动影响。
-
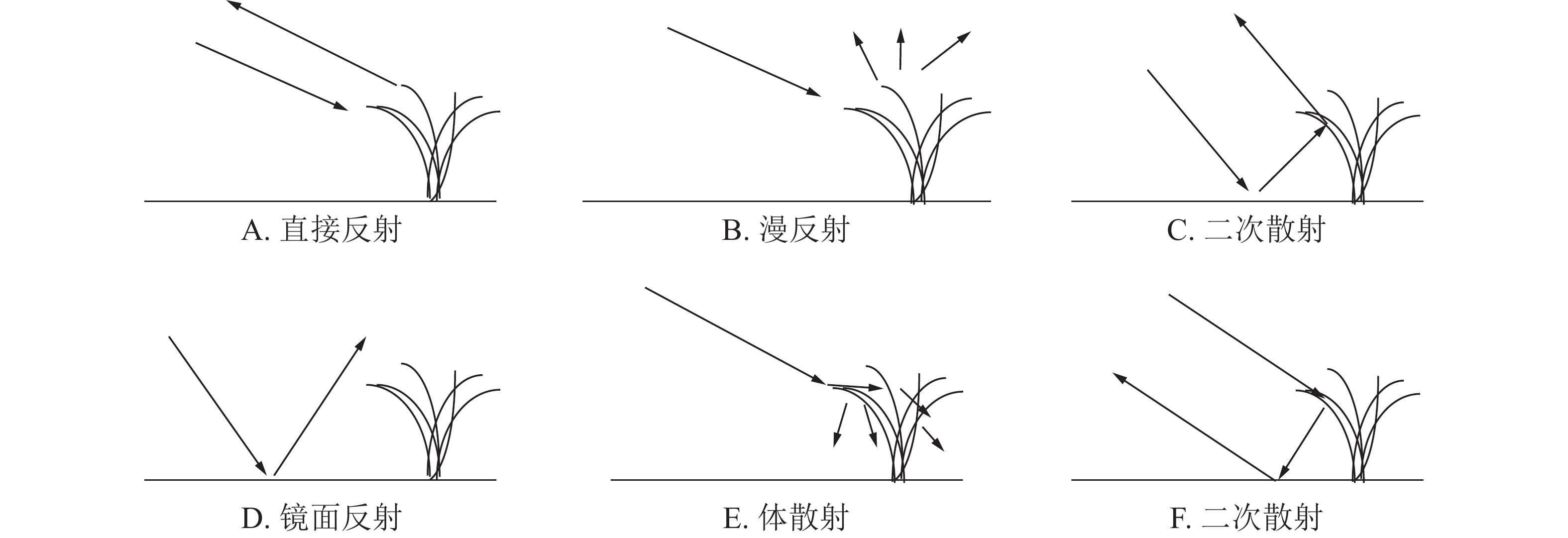
水稻的散射类型一般可分为上部冠层的直接后向散射(图2A)、漫反射(图2B),水面和主茎之间的二次散射(图2C和图2F),水稻田水面的镜面反射(图2D)、水稻植株内部的体散射(图2C)为主[15-16]。
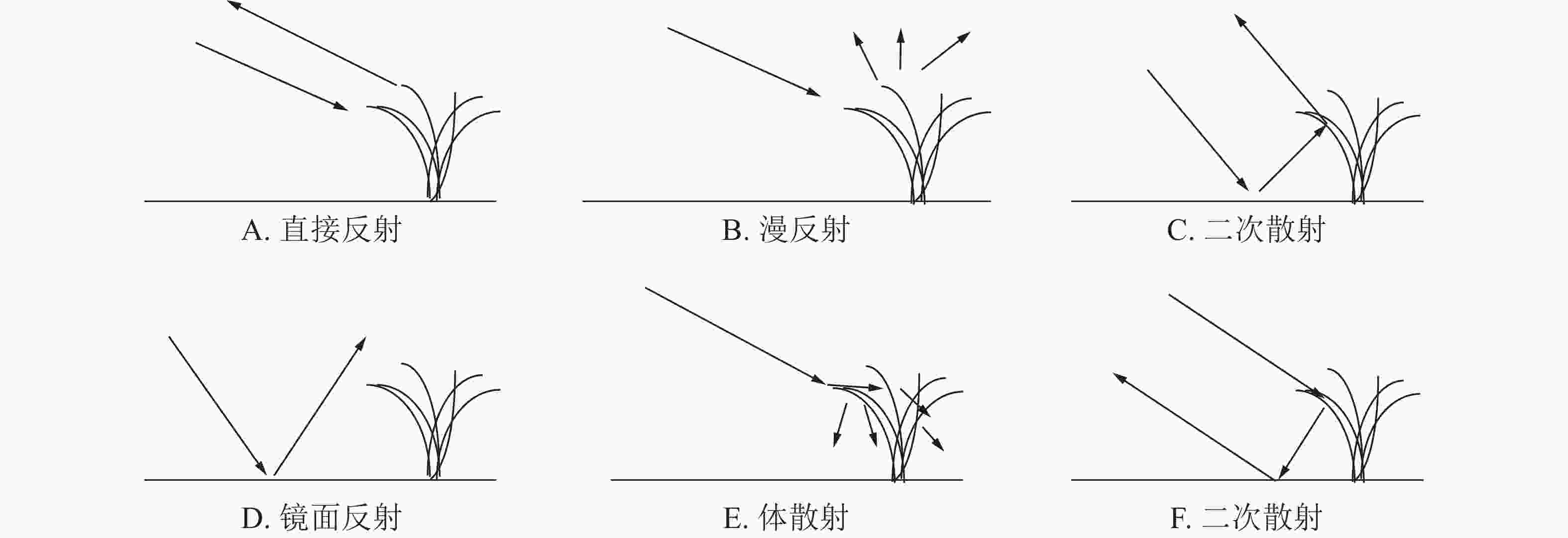
图 2 水稻后向散射类型
Figure 2. Rice backscattering types
-
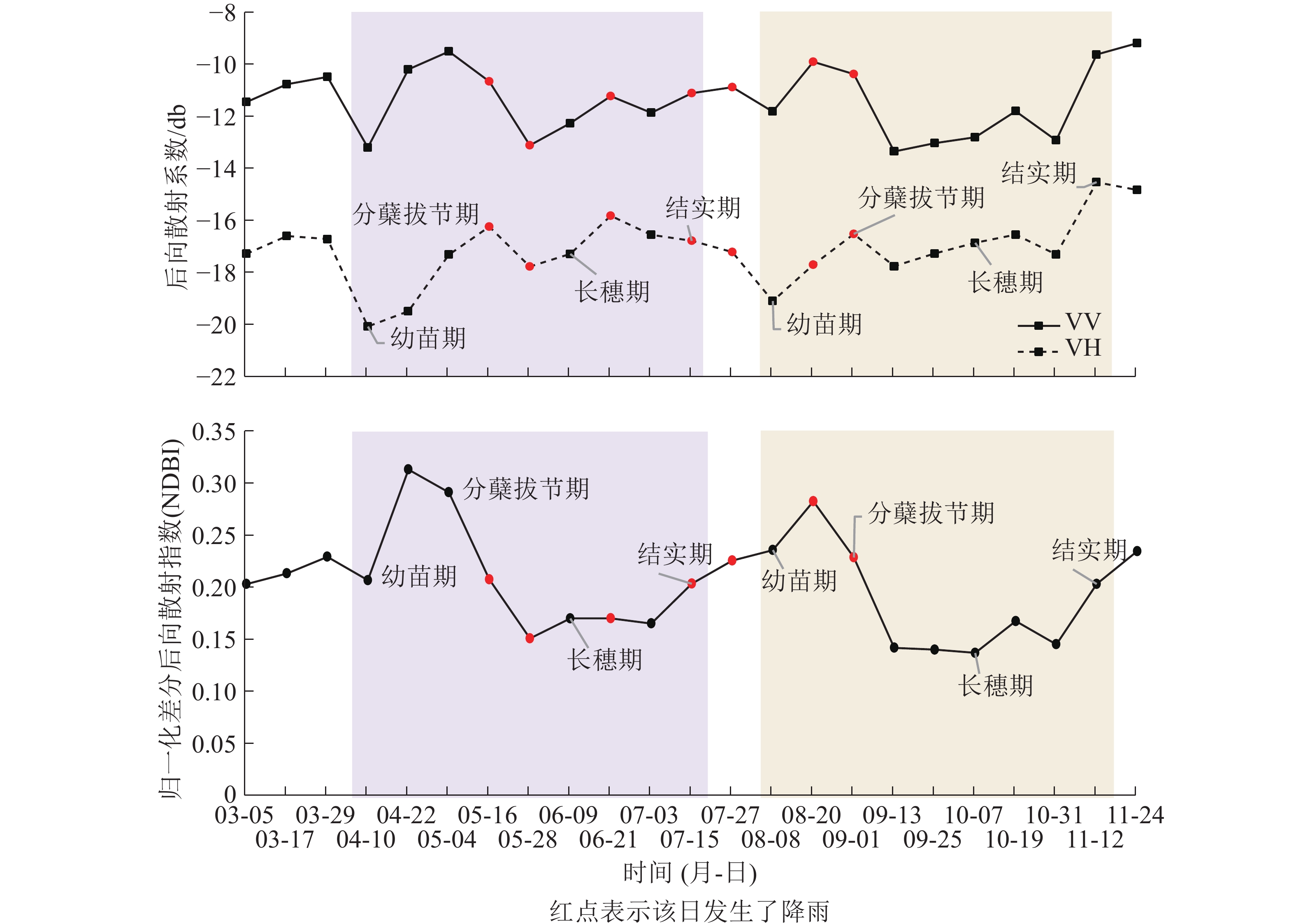
已有研究表明:VV极化具有更高的穿透性,能够穿透植被层到达下垫土壤并与之发生相互作用;VH极化对植被更为敏感[14];本研究的NDBI指数则同时考虑了VH和VV极化之间的差异性。结合图3进行分析发现:VV后向散射系数值整体高于VH,说明VV极化方式更容易穿透水稻植株,在其后向散射系数中包含了更多的来自稻田表面的回波能量,而VH极化的穿透性稍弱,其回波能量主要来自于植株。在幼苗期至分蘖初期VV和VH曲线都呈上升趋势,其原因在于此阶段水稻叶片细少,植株矮小且分布稀疏,散射类型以二次散射(图2C或图2F)和镜面散射(图2D)为主,只夹杂了少量的由水稻幼苗至分蘖初期贡献的体散射(图2E)能量。期间未出现降雨,VV和VH曲线呈单调上升趋势。NDBI曲线也呈上升趋势,并在幼苗期至分蘖期过程中到达整个生长期的最高值,这是因为VV的上升速度较VH更快,二者的差异经NDBI方法放大形成的。在分蘖中期至长穗期,出现了2次降雨,造成了VV和VH曲线不同程度的波动。其中5月16日的降雨使VV曲线由上升转为下降,5月28日前1日出现的5.5 mm降雨,使VV和VH曲线都出现了大幅下降,原因可能由于稻田内部形成较厚积水层,部分微波信号被积水层吸收[15]。NDBI曲线在上述降雨时间点上的变化较为平缓,表现为分蘖中后期单调下降后平稳进入长穗期,其原因在于水稻不断分蘖拔节,植株不断伸长,冠层叶片趋于密集,主要由VH贡献的漫反射(图2B)和体散射能量增加,VV穿透植株经水面反射的回波能量被植株消减降低,二者的差异逐渐缩小,相应的NDBI也有所下降。从长穗期到结实期直至收割期间,出现了3次降雨,对VV和VH曲线的趋势产生了或同向或反向的无规律波动影响,影响程度相较于分蘖期略轻微。NDBI曲线则保持较平稳上升的趋势,几乎不受降雨影响。其原因在于长穗期的叶面积指数和植株高度均达到最大,此时的水稻植株粗壮,冠层茂密,水面二次散射和镜面散射的回波信号受到冠层阻挡,而接收到的回波信号多为植株表面散射,以漫反射和体反射为主,因降雨引起的稻田水面二次散射和镜面散射的回波能量变化受到冠层较多衰减,在曲线的趋势上并不能明显体现。水稻在结实期不断成熟过程中,稻粒体积和质量不断增加,逐渐压弯茎秆,造成植株高度小幅降低,使得VV回波能量小幅提升,VH回波能量则小幅下降,二者在结实期的差异经NDBI放大后得到曲线小幅上升的结果。NDBI曲线在早晚2季水稻的生长周期中都体现出相近的变化趋势,即在幼苗期和分蘖期为快速上升至最高值,在分蘖中后期到长穗初期开始下降,在长穗期到结实期为平稳小幅上升。
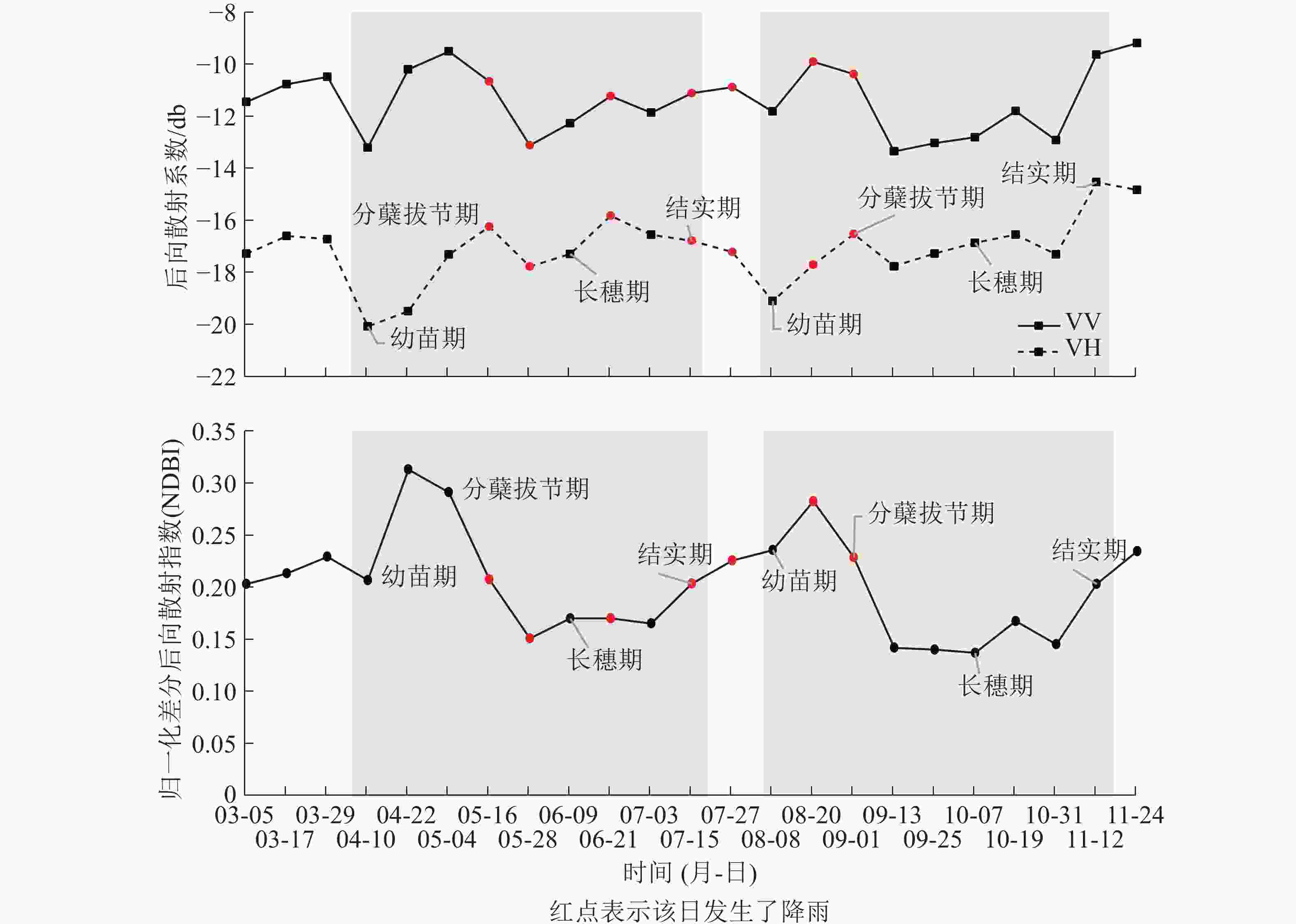
图 3 极化特征时序曲线
Figure 3. Time series curves of polarization characteristics
对时序曲线的波动程度评价,采用二次多项式分段拟合并求平均曲率的方法,平均曲率越低,表明该曲线波动幅度越小,越趋于平稳。
$$ \overline K = \frac{1}{n}\mathop \sum \limits_{i = 2}^{n - 1} \frac{{\left| {P_i^{''}} \right|}}{{{{\left( {1 + P{{_i^{'}}^2}} \right)}^{\frac{3}{2}}}}}\text{。} $$ (2) 式(2)中:
$\overline K$ 为平均曲率,$n = 1,\;2,\; \cdots ,\;23$ 为时间序号,$ i\in n $ ,$ {P}_{i} $ 是对第i−1、i、i+1期极化特征值组成的曲线进行二次多项式拟合得到的函数,$P_i^{'}$ 和$P_i^{''}$ 分别是$ {P}_{i} $ 函数的一阶导数和二阶导数。经计算,NDBI曲线的平均曲率为0.86,VV和VH曲线的平均曲率分别为2.90和2.31。说明NDBI曲线的变化趋势更为平稳,受降雨不规则波动的影响更小,能够更清晰地反映水稻的长势变化。
-
引入Planet光学卫星在线高分辨率影像数据对极化特征时序曲线进行验证,从可见光的角度直观的观测目标实验田的水稻长势情况。通过目视解译并结合已有研究,可大致推断水稻在当月所处的生长阶段。由图4可见:3和4月影像(图4A和图4B)的色调以当地黄土壤背景色为主,夹杂少量淡绿色,可判断此时水稻正值幼苗期,植株矮小叶片稀疏;5月影像(图4C)的色调为浓绿色,可判断此时水稻为分蘖拔节期,叶片生长旺盛;6月影像(图5D)的色调相比5月略淡,可判断此时水稻进入长穗期和结实期;7月影像(图4E)的色调重新回到土壤背景色,表明水稻经过结实期后已完成收割。晚稻的光学影像(图4F~I)在色调也具有与早稻相似的变化特征,其共同在时序上反映出的生长阶段,可与2.1小节中对NDBI时序曲线的分析结果相对应[12]。

图 4 2018年3−11月的Planet卫星影像示意图
Figure 4. Satellite images of Planet from March to November, 2018
-
降雨对后向散射系数的影响普遍存在,会在水稻种植期间后向散射系数时间序列上造成不同程度的波动。在本研究区中,5−7月处于早稻的长穗期,该阶段植株结构变化趋于平缓,但由于期间多次降雨,导致VH和VV时序曲线分别在4.0和3.5 db的范围内出现了波动,这一观测结果与梁瀚月[8]研究结果十分接近。后者所处江西省南昌市的水稻研究区,在5−6月受地当地降雨影响,使得VH和VV时序曲线分别在4.7和3.3 db内出现了波动,且曲线形态也呈现与本研究相似的先快速上升后快速下降的趋势。而NDBI时序曲线,相较于上述单一极化方式在这一时间段中则更趋于平稳。
在水稻生长的中后期,水稻的体散射特征逐渐显现,散射强度由弱转强,NDBI方法可较清晰地展现这一特征,并且与韩宇[5]利用全极化数据获得的水稻生长中后期的体散射变化趋势一致。体散射的强弱通常与植被茎叶密集程度,以及水稻的成熟度密切相关,因此,用NDBI方法可以更直观地表现出水稻长势阶段的规律特征。
-
本研究在常规的VV和VH后向散射系数时序分析的基础上提出了NDBI方法,并对比分析了三者在水稻生长期中的极化特征表现,尤其是受降雨影响的程度。研究发现:降雨发生后,短时间内会使VH和VV的后向散射系数出现较大波动,从而对水稻当前生长阶段的判断产生干扰;而同期的NDBI曲线则能有效地消减这一影响,更准确地反映水稻长势规律。通过这一手段,得到了水稻生长期中极化特征变化规律:幼苗期主要以二次反射为主,而从分蘖拔节时期开始到长穗期再到结实期直至收割,体反射和漫反射不断增强,二次散射逐渐减小。
Extraction of polarization characteristics of Oryza sativa growth under the rainfall fluctuation
-
摘要:
目的 直接利用合成孔径雷达(SAR)后向散射系数监测水稻Oryza sativa等南方作物长势的方法易受降雨影响,在时序曲线上易产生大幅波动。研究降低这一影响的方法,以期获得更准确反映水稻长势规律的结果。 方法 以广西省桂平市水稻栽植区为研究对象,采用2018年3−11月共23期Sentinel-1双极化SAR数据,以及同期降雨数据,分析水稻长势在降雨影响下的同向极化(VV)和交叉极化(VH)后向散射系数时序特征,在此基础上提出基于VV和VH后向散射系数组合的改进方法——归一化差分后向散射指数(NDBI),并结合光学卫星影像数据对结果加以验证。 结果 NDBI时序曲线的平均曲率为0.86,低于VV和VH时序曲线的平均曲率(分别为2.90和2.31),表明NDBI时序曲线的变化趋势更为平稳,受降雨不规则波动的影响更小。早、晚2季水稻的NDBI时序曲线具有相似的变化趋势,能够更准确地反映水稻幼苗期、分蘖拔节期、长穗期和结实期各阶段的极化特征,即幼苗期主要以二次反射为主,而从分蘖拔节时期开始到长穗期再到结实期直至收割,体反射和漫反射不断增强,二次散射逐渐减小。 结论 利用NDBI方法降低了降雨对极化特征时序曲线扰动的影响,获得更准确的水稻长势规律。图4表1参16 -
关键词:
- Sentinel-1 /
- 降雨 /
- 水稻 /
- 极化特征
Abstract:Objective The method of directly using synthetic aperture radar (SAR) backscatter coefficient to monitor the growth trend of Oryza sativa and other crops in south China is susceptible to rainfall and prone to produce large fluctuations on the time series curve. The objective of this study is to explore methods to reduce this impact and obtain more accurate results reflecting the law of O. sativa growth. Method Taking the O. sativa planting area in Guiping City of Guangxi as sample area, the temporal characteristics of VV and VH-polarized backscattering coefficient of O. sativa growth under the influence of rainfall were analyzed using Sentinel-1 dual-polarization SAR data of 23 periods from March to November 2018 and the corresponding rainfall data. On this basis, an improved method named “Normalized Differential Backward Scattering Index (NDBI)” based on the combination of VV and VH-polarized backscatter coefficients was proposed and the results were verified by optical satellite image data. Result The mean curvature of the NDBI curve was 0.86, which was lower than that of VV and VH curves (2.90 and 2.31, respectively), indicating that the NDBI curve was more stable and less affected by irregular rainfall fluctuations. The NDBI curves of O. sativa in the early and late seasons had similar trends, which could more accurately reflect the polarization characteristics of O. sativa at the seedling stage, tiller jointing stage, long spike stage and fruiting stage. The secondary reflection was the main part in seedling stage, and from tiller jointing stage to long spike stage to fruiting stage to harvest stage, the body reflection and diffuse reflection increased continuously and the secondary scattering decreased gradually. Conclusion It is feasible and applicable to use the NDBI method to reduce the influence of rainfall on the disturbance of polarization characteristic time series curve and to obtain more accurate O. sativa growth law. [Ch, 4 fig. 1 tab. 16 ref.] -
Key words:
- Sentinel-1 /
- rainfall /
- Oryza sativa /
- polarization characteristics
-
图 4 2018年3−11月的Planet卫星影像示意图
Figure 4 Satellite images of Planet from March to November, 2018
表 1 Sentinel-1卫星成像时刻雨量
Table 1. Rainfall of Sentinel-1 satellite imaging day
成像日期(年-月-日) 降雨量/mm 当天 前1日 前2日 2018-05-16 1.34 2018-05-28 0.08 5.51 2018-06-21 5.91 2018-07-15 1.33 2018-07-27 3.76 2018-08-20 5.87 2018-09-01 3.02  下载: 导出CSV
下载: 导出CSV
-
[1] 国家统计局. 中国统计年鉴2018[M]. 北京: 中国统计出版社, 2018. [2] 杨文钰, 屠乃美. 作物栽培学各论: 南方本[M]. 2版. 北京: 中国农业出版社, 2011. [3] SHUAI G Y, ZHANG J S, BASSO B, et al. Multi-temporal RADARSAT-2 polarimetric SAR for maize mapping supported by segmentations from high-resolution optical image [J]. Int J Appl Earth Obs Geoinf, 2019, 74: 1 − 15. [4] LI M M, BIJKER W. Vegetable classification in Indonesia using Dynamic Time Warping of Sentinel-1A dual polarization SAR time series [J]. Int J Appl Earth Obs Geoinf, 2019, 78: 268 − 280. [5] 韩宇. 多时相C波段全极化SAR农作物识别方法研究[D]. 呼和浩特: 内蒙古师范大学, 2015. HAN Yu. Research of Crop Recognition Method Based on Multi-temporal C-Band Quad-Pol SAR[D]. Hohhot: Inner Mongolia Normal University, 2015. [6] 张晓倩. 基于全极化SAR数据的水稻识别与作物参数估算模型[D]. 北京: 中国地质大学, 2012. ZHANG Xiaoqian. Rice Extrsction Reseach and Rice Crop Parameter Estimation Based on Full-polarization SAR Data[D]. Beijing: China University of Geoscience, 2012. [7] LOPEZSANCHEZ J M, Ballesterberman J D, Cloude S R. Monitoring and retrieving rice phenology by means of satellite SAR polarimetry at X-band [J]. Geosci Remote Sensing Symp IEEE Int, 2011, 24(8): 2741 − 2744. [8] 梁翰月. 基于Sentinel-1数据的江西省南昌县早稻信息提取研究[D]. 成都: 成都理工大学, 2017. LIANG Hanyue. Extraction of Early Paddy Rice Information in Nanchang County of Jiangxi Province Based on Sentinel-1 Data[D]. Chengdu: Chengdu University of Technology, 2017. [9] 广西壮族自治区统计局. 广西统计年鉴2013−2018. [M]. 北京: 中国统计出版社, 2014−2019. [10] 付盛贤, 莫永生. 优质籼稻百香139的特征特性及栽培技术[J]. 中国稻米, 2011(3): 76 − 77. FU Shengxian, MO Yongsheng. Characteristics and cultivation techniques of high quality indica “Bai xiang 139” [J]. China Rice, 2011(3): 76 − 77. [11] 欧阳伦曦, 李新情, 惠凤鸣, 等. 哨兵卫星Sentinel-1A数据特性及应用潜力分析[J]. 极地研究, 2017(2): 286 − 295. OUYANG Lunxi, LI Xinqing, HUI Fengming, et al. Sentinel-1A data products’ characteristics and the potential applications [J]. Chin J Polar Res, 2017(2): 286 − 295. [12] Planet Labs Inc. Planet imagery product specifications[EB/OL]. 2019. https://assets.planet.com/docs/combined-imagery-product-spec-final-may-2019.pdf [13] HENRI M. 合成孔径雷达图像处理[M]. 孙洪, 译. 北京: 电子工业出版社, 2013. [14] 李新尧. 基于Sentinel-1 SAR数据的表层土壤水分遥感反演[D]. 西安: 西北大学, 2019. LI Xinyao. Study of Retrieving Surface Soil Moisture Based on Sentinel-1 SAR Data[D]. Xi’an: Northwest University, 2019. [15] 王君毅, 杨彬, 张远. 水稻冠层参数微波雷达反演系统设计与实现[J]. 遥感信息, 2016, 31(2): 30 − 35. WANG Junyi, YANG Bin, ZHANG Yuan. Design and implementation of inversion system for rice canopy parameters based on microwave remote sensing technology [J]. Remote Sensing Inf, 2016, 31(2): 30 − 35. [16] 邵芸, 廖静娟, 范湘涛, 等. 水稻时域后向散射特性分析: 雷达卫星观测与模型模拟结果对比[J]. 遥感学报, 2002, 6(6): 440 − 450. SHAO Yun, LIAO Jingjuan, FAN Xiangtao, et al. Analysis of backscattering characteristics of rice in time domain: comparison between radar satellite observation and model simulation [J]. J Remote Sensing, 2002, 6(6): 440 − 450. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190605
 点击查看大图
点击查看大图
图(4) / 表(1)
计量
- 文章访问数: 2075
- HTML全文浏览量: 365
- PDF下载量: 45
- 被引次数: 0









