





-
森林对大气降水产生影响的第1个层次就是林冠[1],林冠层对大气降水的再分配作用在森林生态系统水文循环过程中占据重要的地位。再分配作用增加了降水在空间分布上的多样性,形成了3个部分:林冠截留、穿透雨以及树干径流,这3个部分对整个森林生态系统均有重要的影响。目前已有大量文献报道了中国主要森林的林冠截留特征,国内主要森林生态系统的林冠截留率为11%~37%[2],林冠截留中一部分水分被叶片、枝干吸收,一部分通过蒸发直接返回大气[3]。在亚马逊地区水循环过程中蒸发的水分25%~50%来自于降水,而其中的1/3来源于林冠截留[4]。降雨经过林冠层形成具有较小动能的穿透雨,减小降雨对林地的侵蚀,穿透雨中含有较为丰富的养分,这部分养分构成了森林生态系统物质循环和能量流动的重要组成[5],部分研究显示穿透雨具有汇集效应[1, 6]。树干径流所占比例较小,在研究中通常被忽视,但部分学者研究证明,某些地区忽视树干径流会造成很大的误差[7-8],树干径流通过淋洗林冠及树干而获得的养分数倍于降雨以及穿透雨,这部分水分和养分可直达植物的根系,对干旱或半干旱地区的植被具有重要的生态意义[9]。杉木 Cunninghamia lanceolata是中国特有的速生用材树种,分布广泛,有关杉木林水文效应研究的文献较多[10-14],但这些研究存在研究地域较为分散和研究方法精确度不高等问题。长三角地区经济发达、人口密集,对水资源的需求大,杉木是该地区典型的常绿针叶树之一,是重要的的造林树种。目前,有关长三角地区杉木林降雨再分配关系的研究较少,杉木林对该区域净化水质、水源涵养及水土保持具有重要的作用,对维持该地区的区域生态安全有着重要意义,对其丰水季的降雨再分配格局进行深入分析,以期加深杉木林水文效应机理的认识,为本地区水源涵养林树种的选择以及经营、管理提供理论依据。
-
研究点设在南京市东善桥林场铜山分场(31°35′~31°39′N,118°50′~118°52′E),试验区属北亚热带季风气候区,年平均气温为15.1 ℃,无霜期229 d,年日照时数2 199 h,年平均降水量1 100 mm,时间分布上多存在2个多雨期:一是春夏之交的梅雨,二是夏季的台风雨,为每年的4月至9月。地形为苏南丘陵,土壤类型为黄棕壤。海拔在38~388 m,林分类型以杉木Cunninghamia lanceolata,马尾松Pinus massoniana,麻栎Quercus acutissima,毛竹Phyllostachys edulis林和茶Camallia sinensis园为主。调查样地位于阴坡中下部,杉木林平均坡度为22°,平均林龄45 a,平均郁闭度0.61,平均树高10.8 m,平均胸径13.2 cm,密度850株·hm-2。
-
在试验样地外约20 m处的空旷地设置1台翻斗式自记雨量计,持续测定(隔5 min自动采集1次数据)林外降雨量和降雨过程。利用Decagon自动气象站(美国产)对空气温度、湿度、风速以及风向进行测定(隔15 min自动采集1次数据)。
选择杉木林分中生长条件良好的典型坡面设置林冠水文野外观测点,选取1个样地(20 m×20 m)设置3个集水槽(200 cm×20 cm×20 cm)收集穿透雨,布设集水槽时需要除去高于集水槽的植被,将降雨导入称量式自动排液式测定系统,最后根据集水槽的面积以及3个集水槽收集到的穿透雨量换算出林内穿透雨量(mm)。
试验采用自制试验装置—树干径流仪。该装置采用喷塑铁皮由模具制成半圆形容器,2个半圆形容器围绕同等直径树干合围,形成树干径流收集装置。铁皮与树干直接用玻璃胶消除缝隙,装置下方有出水口,连接皮管将收集到的雨水导入下方放置的翻斗式雨量计中,通过翻斗式雨量计来计算收集到的树干径流量。按4 cm 为1个径级划分分布,选取样地中3株接近各径级平均胸径的样木(共9株),装置仪器进行观测,然后将径流量按林木径级及其权重进行统计,利用加权平均法推算出林分的树干径流量。
利用水量平衡方程I=P-S-T计算林冠截留量,其中:I为林冠截留量(mm),P为大气降雨量(mm),S为树干径流量(mm),T为穿透雨量(mm)。
-
在观测期2012年4-9月内,累计47场降雨。为方便研究该地区降雨特征,依据降雨量的大小依次分为:0~1,1~5,5~10,10~15,15~20,20~30,>30 mm共7个降雨等级(表 1)。对检测到的大气降雨数据进行分析表明:从降雨量级的分布来看,降雨量<10.0 mm的小雨占总降雨场数的比例最大,总计70.21%。从降雨量来看,占降雨场数最少的>30.0 mm的降雨占总降雨量的比例最大,为41.56%,而占据降雨场数比例优势的小雨仅占降雨量的19.86%。研究区内降雨强度较小的降雨事件占据优势,<1.5 mm·h-1的降雨占到降雨次数的68.1%。
表 1 降雨特征
Table 1. Rainfall characteristics
降雨量等级/(mm • d-1) 降雨天数/d 降雨天数百分比/% 降雨量/mm 降雨量百分比/% 0~1 12 25.33 6.6 1.40 1~5 12 25.53 29 6.16 5~10 9 19.15 57.9 12.30 10~15 4 8.51 49.8 10.58 15~20 3 6.38 51 10.83 20~30 3 6.38 80.8 17.17 >30 4 8.51 195.6 41.56 -
林分特征与气象特征影响林冠层降雨再分配格局。本研究观测的气象因子包括降雨量、降雨强度、空气相对湿度、空气温度、风速、风向,利用SPSS 19对气象因子与穿透雨、树干径流以及林冠截留进行相关性分析发现,降雨量、降雨强度与穿透雨、树干径流以及林冠截留相关性极为显著,风速与穿透雨和树干径流呈显著正相关,空气相对湿度、空气温度以及风向与穿透雨、树干径流以及林冠截留并无显著相关性(表 2)。
表 2 穿透雨、树干径流、林冠截留与气象因子的Pearson相关系数
Table 2. Pearson correlation between throughfall, stemflow and canopy interception and environmental factors
项目 降雨量 降雨强度 空气相对湿度 空气温度 风速 风向 穿透雨T 0.994** 0.525** 0.209 0.120 0.346* -0.002 树干径流S 0.879** 0.311* 0.142 0.160 0.601** 0.13 林冠截留I 0.786** 0.578** 0.238 0.029 -0.006 -0.099 说明:** 表示在0.01 水平(双侧)上显著相关;*表示在0.05水平(双侧)上显著相关。 -
基于2012年4月至9月观测到的降雨与穿透雨的数据,杉木林内累计穿透雨量344.1 mm,占降雨量比例的73.1%。影响穿透雨的影响因子包括气象因子和林分因子。本次研究探讨了穿透雨量、穿透雨率与降雨量及降雨强度的关系。图 1显示了长三角地区杉木林的穿透雨量与降雨量具有显著的线性正相关,穿透雨量随林外降雨量的增加而变大。在降雨量较小时穿透雨会被林冠全部截持,当穿透雨量为0 mm时X轴的截距就为形成穿透雨的最小雨量,而这个值可以根据拟合出的方程计算得到,长三角地区杉木林形成穿透雨的最小雨量为0.9 mm。降雨量与穿透雨率的回归关系为对数函数关系,降雨量<20.0 mm时,随降雨量增大穿透雨率增加速度较快,之后增加较为缓慢并趋于稳定。根据最大R2值来判断,

图 1 穿透雨量、穿透雨率与降雨量的关系
Figure 1. Relationship between throughfall, throughfall rate and rainfall
穿透雨量、穿透雨率与降雨强度的关系用对数函数拟合效果较好,降雨强度<2.5 mm·h-1时,穿透雨量增长较快,之后穿透雨增长速度减缓,穿透雨率在降雨强度<5.0 mm·h-1时增长速度最快,随后增长速度变缓(图 2)。

图 2 穿透雨量、穿透雨率与降雨强度的关系
Figure 2. Relationship between throughfall, throughfall rate and rainfall intensity
-
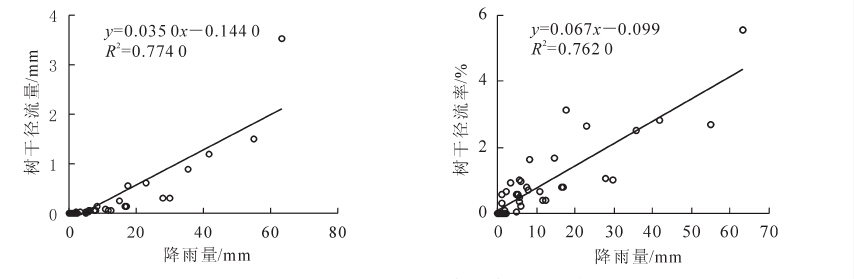
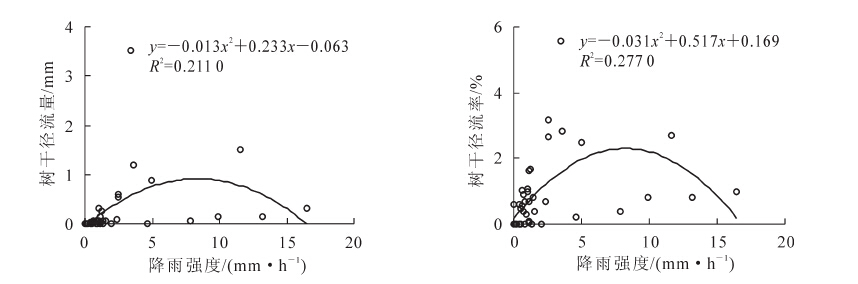
树干径流量所占比例小,在研究过程中通常会被很多研究者忽略,但是这部分水分中包含的养分浓度数倍于大气降水和穿透雨,并且可以直达植物根系地区便于植物的吸收和利用,在干旱区这部分水分对植被尤为重要。观测期内,杉木林累计树干径流量10.3 mm,占降雨量的2.2%。长三角地区杉木林树干径流量与降雨量呈显著的线性关系,树干径流量随降雨量的增大而增加,雨量较小的降雨形成不了树干径流。形成树干径流的最小雨量为拟合方程与X轴的截距,经计算杉木林树干径流形成的最小雨量为4.1 mm。树干径流率与降雨量也有显著的线性正相关关系(图 3)。树干径流量、树干径流率与降雨强度的相关关系用二次函数可得到较为满意的拟合结果,树干径流量、树干径流率随降雨强度的变化规律都是先增大后减小呈抛物线形,在5 mm·h-1左右达到最大值随后开始减小,推测原因降雨强度超过一定范围,雨滴的冲击力过强导致叶片、枝干截留降水减少,因此树干径流量、树干径流率也随之变小(图 4)。

图 3 树干径流量、树干径流率与降雨量的关系
Figure 3. Relationship between stemflow, stemflow rate and rainfall

图 4 树干径流量、树干径流率与降雨强度的关系
Figure 4. Relationship between stemflow, stemflow rate and rainfall intensity
-
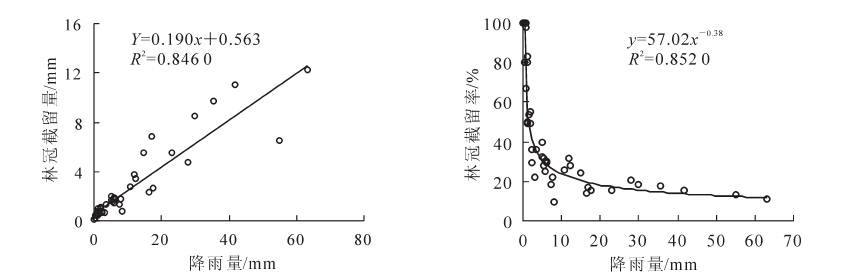
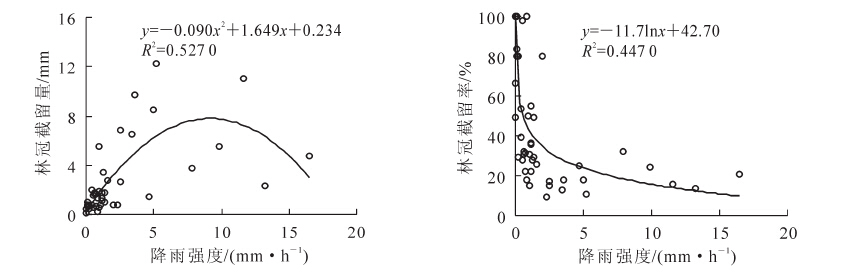
由图 5可知:杉木林林冠截留量与降雨量呈线性关系,林冠截留率与降雨量的关系可以用幂函数曲线来拟合,拟合效果较好。这与多数学者在不同地区的研究相符合。林冠截留量随降雨的增大而增大,但增长速率渐缓,林冠截留率在<10 mm阶段下降速度快,之后随降雨量的增加而减少并趋于稳定。林冠截留量与降雨强度的相关关系用二次函数拟合可得到较满意的效果,林冠截留量并不是一直随降雨强度的增大而增大,而是随降雨强度的增大先增大后减小呈抛物线形。推测原因,是由于降雨强度较小时林冠层可以截留大部分的降水,随着降雨强度的增大截留量也紧跟着增大,但当降雨强度超过一定值之后,雨滴的动能过大造成林冠层机械晃动剧烈导致冠层截持不住降水,导致截留量反而下降。林冠截留率与降雨强度呈显著的对数函数关系(P<0.01),在0~5 mm·h-1阶段下降十分迅速,之后随降雨强度的变大而减小,但趋势较缓和(图 6)。

图 5 林冠截留量、林冠截留率与降雨量的关系
Figure 5. Relationship between canopy interception, canopy interception rate and rainfall

图 6 林冠截留量、林冠截留率与降雨强度的关系
Figure 6. Relationship between canopy interception, canopy interception rate and rainfall intensity
-
本研究采用定位观测的方法对南京市东善桥林场铜山分场杉木林的降雨再分配进行了测定。通过相关性分析,筛选出的影响降雨再分配的主要气象因子为降雨量、降雨强度以及风速,并详细分析了降雨量、降雨强度与穿透雨、树干径流以及林冠截留的关系,分析结果如下:①2012年4月至2012年9月之间,长三角地区杉木林内穿透雨占降雨量的73.1%。杉木林的穿透雨量与降雨量具有极显著的线性正相关,形成穿透雨的最小雨量为0.9 mm,在某些降雨量很小但降雨强度较大的降雨事件中,降雨量未达到0.9 mm穿透雨量也会出现,但0.9 mm这个结果与观测到的绝大多数降雨数据吻合。穿透雨率与降雨量的回归关系为对数函数关系。穿透雨量、穿透雨率与降雨强度的关系用对数函数拟合效果较好,曲线变化趋势均是前期增长较快,随后增加速度变缓并趋于稳定。②调查期内,杉木林累计树干径流量10.3 mm,占据降雨量比例的2.2%,低于于小军等[11]在会同生态试验站测定的杉木林树干径流率(4.45%),推测原因本研究区年平均降水量较低,且小雨量低强度的降雨占据优势,导致了树干径流率较低。杉木林树干径流量、树干径流率与降雨量呈极显著的线性正相关关系,均随林外降雨量的增加而增大。杉木林树干径流量、树干径流率与降雨强度的关系用二次函数拟合可得到较满意的结果,两者随降雨强度的变化趋势都呈抛物线形,先增大后减小,这可能是雨强较小时林冠层截持的降雨顺着枝干到达根部底端转化为树干径流,降雨强度变大,降雨较强的冲击力导致林冠层可截留的降雨减少,最终到达根部的树干径流量也变少。③研究林分杉木林冠截留量(116.3 mm)占降雨量的24.7%,杉木林冠截留量与降雨量的关系用幂函数拟合效果最好,与巩合德等[13]和张志永等[14]研究得出林冠截留量与降雨量呈幂函数(I=aP b)的关系结论一致,林冠截留率随降雨量的增大而减小,体现出林冠层蓄水能力的有限性,用幂函数方程最能反映林冠截留率与降雨量的关系。贾永正等[15]对长三角地区毛竹林冠截留的研究也得出相同结果。林冠截留量与降雨强度用二次函数拟合效果较好,随降雨强度的增大林冠截留先增大后减小,林冠截留量曲线呈抛物线形,当雨强超过10 mm·h-1左右时,林冠层受到雨滴的冲击过大导致截留的降水变少。随降雨强度的增大,截留率持续降低,前期下降较快,之后下降速率变缓。
John等[16]研究得出风速风向、空气温湿度等气象因子也会对树干径流、穿透雨以及林冠截留产生影响,此次研究中降雨再分配与空气相对湿度、气温以及风向等气象因子并无显著的相关性,仅仅与风速有一定的相关性,在今后的观测中需要进行连续长期的定位观测,以获得更加准确的结论。
Rainfall redistribution in a Cunninghamia lanceolata forest of the Yangtze River Delta
-
摘要: 对长三角地区丰水季杉木Cunninghamia lanceolata林大气降雨量、穿透雨量以及树干径流量进行了定位观测,利用水量平衡方程计算得出林冠截留量。通过相关性分析筛选出影响降雨再分配的主要因子为降雨量和降雨强度,分别分析了降雨量与穿透雨、树干径流、林冠截留之间的关系,以及降雨强度与三者之间的关系。结果显示:2012年4月至9月长三角地区杉木林外降雨累计470.7 mm,研究区以小雨量、低强度的降雨事件为主,杉木林内累计穿透雨量344.1 mm,占降雨量的73.1%,树干径流总量10.3 mm,占降雨量的2.2%,林冠截留量达到116.3 mm,占降雨量的24.7%。建立了降雨量、降雨强度与杉木林穿透雨量、树干径流量、林冠截留量之间的回归模型,利用拟合出的方程,可得出杉木林形成穿透雨的最小雨量为0.9 mm,形成树干径流的最小雨量为4.1 mm。Abstract: Water transport through the canopy layer, important in forest hydrology and soil and water conservation, was studied to determine characteristics of rainfall redistribution and the eco-hydrological functions of forests.Using the fixed point observation method, rainfall, throughfall, and stemflow of Cunninghamia lanceolata in the Yangtze River Delta were studied during the high flow season.A water balance method was adopted to calculate canopy interception, and a regression analysis was employed to select the main factors affecting rainfall redistribution and to establish the relationship among throughfall, stemflow, canopy interception, rainfall, and rainfall intensity.Results indicated a cumulative rainfall of 470.7 mm with most rainfall events from April to September 2012 featuring light rain with low intensity.Cumulative throughfall was 344.1 mm; total stemflow was 10.3 mm; and total canopy interception was 116.3 mm.The fitted regression equation showed that the minimum rainfall necessary for throughfall was 0.9 mm, and the minimum rainfall needed to start stemflow was 4.1 mm.
-
Key words:
- forest hydrology /
- the Yangtze River Delta /
- Cunninghamia lanceolata /
- throughfall /
- stemflow /
- canopy interception
-
图 1 穿透雨量、穿透雨率与降雨量的关系
Figure 1 Relationship between throughfall, throughfall rate and rainfall
图 2 穿透雨量、穿透雨率与降雨强度的关系
Figure 2 Relationship between throughfall, throughfall rate and rainfall intensity
图 3 树干径流量、树干径流率与降雨量的关系
Figure 3 Relationship between stemflow, stemflow rate and rainfall
图 4 树干径流量、树干径流率与降雨强度的关系
Figure 4 Relationship between stemflow, stemflow rate and rainfall intensity
图 5 林冠截留量、林冠截留率与降雨量的关系
Figure 5 Relationship between canopy interception, canopy interception rate and rainfall
图 6 林冠截留量、林冠截留率与降雨强度的关系
Figure 6 Relationship between canopy interception, canopy interception rate and rainfall intensity
表 1 降雨特征
Table 1. Rainfall characteristics
降雨量等级/(mm • d-1) 降雨天数/d 降雨天数百分比/% 降雨量/mm 降雨量百分比/% 0~1 12 25.33 6.6 1.40 1~5 12 25.53 29 6.16 5~10 9 19.15 57.9 12.30 10~15 4 8.51 49.8 10.58 15~20 3 6.38 51 10.83 20~30 3 6.38 80.8 17.17 >30 4 8.51 195.6 41.56  下载: 导出CSV
下载: 导出CSV
表 2 穿透雨、树干径流、林冠截留与气象因子的Pearson相关系数
Table 2. Pearson correlation between throughfall, stemflow and canopy interception and environmental factors
项目 降雨量 降雨强度 空气相对湿度 空气温度 风速 风向 穿透雨T 0.994** 0.525** 0.209 0.120 0.346* -0.002 树干径流S 0.879** 0.311* 0.142 0.160 0.601** 0.13 林冠截留I 0.786** 0.578** 0.238 0.029 -0.006 -0.099 说明:** 表示在0.01 水平(双侧)上显著相关;*表示在0.05水平(双侧)上显著相关。
下载: 导出CSV
-
[1] 李振新, 郑华, 欧阳志云, 等.岷江冷杉针叶林下穿透雨空间分布特征[J].生态学报, 2004, 24(5):1015-1021. LI Zhenxin, ZHENG Hua, OUYANG Zhiyun, et al.The spatial distribution characteristics of throughfall under Abies faxoniana forest in the Wolong Nature Reserve[J].Acta Ecol Sin, 2004, 24(5):1015-1021. [2] 刘世荣, 孙鹏森, 温远光.中国主要森林生态系统水文功能的比较研究[J].植物生态学报, 2003, 27(1):16-22. LIU Shirong, SUN Pengsen, WEN Yuanguang.Comparative analysis of hydrological functions of major forest ecosystems in China[J].Acta Phytoecol Sin, 2003, 27(1):16-22. [3] 张娜, 乔玉娜, 刘兴诏, 等.鼎湖山季风常绿阔叶林大气降雨、穿透雨和树干径流的养分特征[J].热带亚热带植物学报, 2010, 18(5):502-510. ZHANG Na, QIAO Yuna, LIU Xingzhao, et al.Nutrient characteristics in incident rainfall, throughfall,and stemflow in monsoon evergreen broad-leaved forest at Dinghushan[J].J Trop Subtrop Bot, 2010, 18(5):502-510. [4] BÄSE F, HELMUT E, NEILL C, et al.Differences in throughfall and net precipitation between soybean and transitional tropical forest in the southern Amazon,Brazil[J].Agric Ecosys Environ, 2012, 159:19-28. [5] HOFHANSL F, WANEK W, DRAGE S, et al.Controls of hydrochemical fluxes via stemflow in tropical lowland rainforests:effects of meteorology and vegetation characteristics[J].J Hydrol,2012,452/453:247-258. [6] 刘建立, 王彦辉, 于澎涛, 等.六盘山叠叠沟小流域华北落叶松人工林的冠层降水再分配特征[J].水土保持学报, 2009, 23(4):76-81. LIU Jianli,WANG Yanhui,YU Pengtao,et al.Characteristics of rainfall redistribution under the canopy of Larix principis-rupprechtii forest in Diediegou watershed of Liupanshan Mountains[J].J Soil Water Conserv, 2009, 23(4):76-81. [7] 杨志鹏, 李小雁, 伊万娟, 等.荒漠灌木树干茎流及其生态水文效应研究进展[J].中国沙漠, 2010, 30(2):304-311. YANG Zhipeng, LI Xiaoyan, YI Wanjuan, et al.Review on stemflow of desert shrubs-research methods and eco-hydrological effects[J].J Desert Res, 2010, 30(2):304-311. [8] LIANG Weili, KOSUGI K, MIZUYAMA T, et al.A three-dimensional model of the effect of stemflow on soil water dynamics around a tree on a hillslope[J].J Hydrol,2009,366(1-4):62-75. [9] ABRAHAMS A D, PARSONS A J, WAINWRIGHT J.Disposition of rainwater under creosotebush[J].Hydrol Proc, 2003,17:2555-2566. [10] 杨茂瑞.亚热带杉木马尾松人工林的林内降雨林冠截留和树干茎流[J].林业科学研究, 1992, 5(2):158-162. YANG Maorui.Interrelations of forest precipitation,crown interception and trunk stemflow in sub-tropical plantations of cunninghamia lanceolata and Pinus massoniana[J].For Res, 1992, 5(2):158-162. [11] 于小军,汪思龙,邓仕坚,等.亚热带常绿阔叶林和杉木人工林茎流与穿透雨的养分特征[J].生态学杂志, 2003, 22(6):7-11. YU Xiaojun, WANG Silong, DENG Shijian, et al.Nutrient characteristics of stemflow and throughfall in evergreen broad-leaved forest and Cunninghamia lanceolata plantation forest[J].Chin J Ecol, 2003, 22(6):7-11. [12] 杜紫贤, 韩永刚, 谢锦升, 等.木荷与杉木人工林林冠截留初步探讨[J].亚热带水土保持, 2007, 19(2):5-9. DU Zixian, HAN Yonggang, XIE Jinsheng, et al.Initial probe into the crown intereeption of artificial Schina superba&fir forest[J].Subtrop Soil Water Conserv,2007,19(2):5-9. [13] 巩合德, 王开运,杨万勤.川西亚高山3种森林群落穿透雨和茎流养分特征研究[J].林业科学,2005,41(5):14-20. GONG Hede,WANG Kaiyun,YANG Wanqin,et al.Nutrient characteristics of throughfall and stemflow in three forests at the subalpine of western Sichuan[J].Sin Sliv Sci,2005,41(5):14-20. [14] 张志永, 张卓文, 陈玉生, 等.5种主要森林类型涵养水源能力比较研究[J].福建林学院学报, 2005, 25(2):171-175. ZHANG Zhiyong, ZHANG Zhuowen, CHEN Yusheng, et al.A comparative study on water conservation capacity of five main forest types[J].J Fujian Coll For,2005,25(2):171-175. [15] 贾永正, 胡海波, 张家洋, 等.苏南丘陵区降雨分布格局[J].生态学报, 2011, 31(12):3537-3542. JIA Yongzheng, HU Haibo, ZHANG Jiayang, et al.Canopy interception of rainfall by bamboo plantations growing in the hill areas of southern Jiangsu Province[J].Acta Ecol Sin,2011,31(12):3537-3542. [16] van STAN Ⅱ,SIEGERT C M,LEVIA D F Jr, et al.Effects of wind-driven rainfall on stemflow generation between codominant tree species with differing crown characteristics[J].Agric For Meteorol, 2011, 151(9):1277-1286. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.03.015
 点击查看大图
点击查看大图

计量
- 文章访问数: 3635
- HTML全文浏览量: 782
- PDF下载量: 645
- 被引次数: 0
