





-
植被数量分类和排序是植被生态学研究的热点问题之一,被广泛应用于森林、草原、湿地等各种生态类型的群落研究中[1-5]。数量分类是植被分类的分支学科,是基于样方及物种间相似关系之上的一种分类方法,通过计算样方或物种间的相似系数,把样方或物种归并为组,使得组内成员尽量相似,而不同组的成员尽量相异。双向指示种分析(twinspan)是常用的数量分类方法之一,它能同时完成样方和物种的分类,并反映一定的生态特征。排序是将样方或物种排列在一定的空间,使得排序轴能够反映一定的生态梯度,从而能够解释植被或物种的分布与环境因子间的关系。只使用物种组成数据的排序称为间接排序,同时使用物种数据和环境因子数据的排序称为直接排序。去趋势对应分析(DCA)和典范对应分析(CCA)是应用最为广泛的间接排序和直接排序方法[6]。应用植被数量分类和排序不仅可以把植物群落的分布格局与环境因子进行比较,还可以更好地解释植被或物种的分布与环境因子之间的关系,具有重要的生态学意义[4, 7-8]。浙江省庆元县巾子峰地处亚热带常绿阔叶林植被带,区内生物多样性丰富,特有珍稀物种多。浙江省正按照“一园两区”思路建设钱江源—百山祖国家公园,其中,庆元巾子峰国家森林公园作为中亚热带常绿阔叶林生态系统的典型代表,将与浙江凤阳山—百山祖国家级自然保护区、浙江庆元大鲵国家级水产种质资源保护区等自然保护地整合为百山祖国家公园。范良敏等[9-10]对该公园的木本植物区系及生态旅游开发进行了研究,但关于其森林群落的数量分类、排序、环境生态因子之间的关系还未见报道。本研究以巾子峰国家森林公园森林群落为研究对象,运用双向指示种分析分类和去趋势对应分析、典范对应分析排序方法,分析其植被群落类型及特征,揭示森林群落及优势种与环境因子之间的关系,以期为巾子峰国家森林公园的科学管理、有效保护和可持续利用提供基本资料和科学依据。
-
巾子峰国家森林公园位于浙江省庆元县西部,面积5 752 hm2,27°27′~27°39′N,118°50′~119°06′E,最高峰巾子峰海拔1 563 m。该区气候属中亚热带季风气候区,温暖潮湿,雨水充沛,四季分明。年平均气温为17.2 ℃,最高月平均气温26.9 ℃,最低月平均气温7.0 ℃,年平均降水量1 689.0 mm,平均相对湿度76%;年平均日照时数1 828.2 h,日照率41%;年平均无霜期256.0 d。土壤类型为红壤和黄壤2个土类,均为地带性土壤。红壤广泛分布于海拔800 m以下的低山丘陵,黄壤分布于800 m以上的中低山。主要植被类型有常绿阔叶林、常绿-落叶阔叶混交林、针阔叶混交林、针叶林、毛竹Phyllostachys edulis林等,地带性典型植被为常绿阔叶林。
-
2019年4−5月,在巾子峰国家森林公园海拔500~1 500 m内,隔100 m设置1个取样区间,每个取样区间布设2~8个20 m×20 m样方,共布设10个取样区间48个样方。记录样方内胸径(DBH)≥2 cm的木本植物个体种名、高度、胸径、盖度,每种灌木及草本的种名、高度、盖度、株数或丛数,同时记录每个样方的海拔、坡向、坡度、土壤类型、土壤湿度、土壤温度。其中,海拔高度用全球定位系统测量,坡向和坡度用罗盘仪测定,土壤温度和土壤湿度用Veinasa-TS土壤温湿速测仪测量。48个样方的各个生境因子如表1所示。
表 1 浙江庆元巾子峰国家森林公园样方环境因子
Table 1. Environmental factors of plots in Jinzifeng Forest Park, Qingyuan, Zhejiang
样方号 群落郁闭度 海拔/m 坡位 坡度/(°) 坡向 北纬(N) 东经(E) 土壤类型 1 0.85 554 下坡位 20 西南 27°41'04.026" 119°00'42.080" 红壤 2 0.90 550 下坡位 30 东北 27°41'07.635" 119°00'41.897" 红壤 3 0.85 540 中坡位 30 东 27°41'16.329" 119°00'40.030" 红壤 4 0.80 572 中坡位 30 东 27°41'00.230" 119°00'36.095" 红壤 5 0.85 657 中坡位 30 东 27°40'40.769" 119°00'26.647" 红壤 6 0.90 565 下坡位 35 东南 27°41'12.993" 119°00'40.357" 红壤 7 0.70 694 中坡位 30 西北 27°41'40.270" 119°01'06.858" 红壤 8 0.80 727 中坡位 30 东北 27°40'37.443" 119°00'23.501" 红壤 9 0.90 1 052 山脊 35 东 27°39'59.132" 118°59'59.641" 黄壤 10 0.50 628 中坡位 25 东南 27°39'51.851" 119°00'37.342" 黄壤 11 0.90 571 下坡位 25 东 27°41'00.880" 119°00'41.821" 红壤 12 0.90 538 下坡位 25 北 27°41'18.531" 119°00'39.928" 红壤 13 0.85 791 中坡位 30 西 27°41'27.328" 119°01'02.480" 红壤 14 0.80 780 上坡位 10 西 27°41'13.857" 119°00'57.785" 红壤 15 0.85 767 中坡位 25 东 27°41'33.945" 119°01'09.110" 红壤 16 0.60 750 下坡位 45 东北 27°41'37.304" 119°01'09.533" 红壤 17 0.70 832 中坡位 25 南 27°41'11.570" 119°01'07.716" 红壤 18 0.80 797 下坡位 25 西南 27°41'14.264" 119°00'58.968" 红壤 19 0.80 734 下坡位 30 北 27°40'32.842" 119°00'29.121" 红壤 20 0.80 821 谷底 30 东南 27°40'23.970" 119°00'33.259" 红壤 21 0.80 1 108 上坡位 35 东 27°39'59.474" 118°59'54.098" 黄壤 22 0.60 1 382 上坡位 35 西北 27°39'44.438" 118°59'42.841" 黄壤 23 0.80 776 下坡位 25 东北 27°40'33.656" 119°00'48.684" 红壤 24 0.95 731 中坡位 25 西南 27°42'06.523" 119°00'48.359" 红壤 25 0.80 518 中坡位 25 西 27°39'37.175" 119°00'54.164" 黄壤 26 0.70 613 中坡位 5 南 27°39'44.363" 119°00'35.918" 黄壤 27 0.80 562 中坡位 30 西 27°41'13.205" 119°00'41.127" 红壤 28 0.85 736 山脊 10 北 27°40'43.434" 119°00'39.118" 红壤 29 0.85 681 下坡位 25 东北 27°40'43.766" 119°00'48.120" 红壤 30 0.85 820 下坡位 30 西 27°40'25.046" 119°00'33.625" 红壤 31 0.85 1 250 山顶 30 西南 27°39'53.117" 118°59'49.230" 黄壤 32 0.60 1 317 上坡位 35 西北 27°39'46.784" 118°59'44.727" 黄壤 33 0.80 680 下坡位 30 东北 27°40'36.989" 119°00'52.495" 红壤 34 0.80 590 下坡位 25 东 27°40'55.924" 119°00'46.219" 红壤 35 0.80 747 下坡位 30 东南 27°40'31.064" 119°00'51.985" 红壤 36 0.70 720 下坡位 30 东 27°40'35.337" 119°00'55.786" 红壤 37 0.75 925 山脊 10 东 27°40'18.521" 119°00'25.013" 红壤 38 0.60 784 中坡位 25 西北 27°40'44.187" 119°00'56.161" 红壤 39 0.95 767 中坡位 25 西 27°41'39.635" 119°01'10.765" 红壤 40 0.70 1 148 山脊 25 东北 27°40'13.334" 118°59'41.175" 黄壤 41 0.85 1 178 上坡位 30 东 27°40'07.905" 118°59'34.997" 黄壤 42 0.85 1 197 上坡位 25 东 27°40'08.949" 118°59'29.214" 黄壤 43 0.95 1 203 上坡位 30 西 27°40'11.715" 118°59'22.209" 黄壤 44 0.95 1 194 上坡位 10 南 27°40'08.334" 118°59'37.403" 黄壤 45 0.85 1 198 上坡位 30 北 27°39'56.373" 118°59'33.218" 黄壤 46 0.85 1 231 山脊 25 南 27°39'54.912" 118°59'46.670" 黄壤 47 0.60 1 271 中坡位 25 东北 27°39'49.007" 118°59'47.710" 黄壤 48 0.60 1 413 上坡位 10 北 27°39'41.018" 118°59'42.226" 黄壤 -
用重要值表示每个物种在森林群落中的优势程度,按照乔木树种、灌木及草本植物不同的生长特征,选择不同的重要值计算方法。计算公式如下:乔木树种重要值=(相对密度+相对优势度+相对频度)/3。灌木及草本植物重要值=(相对盖度+相对高度)/2。
-
海拔高度、坡度、土壤湿度、土壤温度直接测量数值。用数字表示不同的土壤类型,其中:0代表红壤,1代表黄壤。为了分析森林群落的向阳程度,将坡向因子进行转换,以数字表示不同的坡向等级,其中:1表示北坡(0°~22.5°和337.5°~360°),2表示东北坡(22.5°~67.5°),3表示西北坡(292.5°~337.5°),4表示东坡(67.5°~112.5°),5表示西坡(247.5°~292.5°),6表示东南坡(112.5°~157.5°),7表示西南坡(202.5°~247.5°),8表示南坡(157.5°~202.5°)。数值越大说明越向阳。
-
使用Wintwins 2.3软件,采用双向指示种分析分类方法,对由48个样方及样方内194个种的重要值组成的48×194维矩阵进行分析,得出数量分类结果。
-
使用CANOCO 4.5软件[11],采用去趋势对应分析排序方法,对由48个样方及样方内194个种的重要值组成的48×194维矩阵进行分析,得出排序结果。采用典范对应分析对由48个样方及样方内194个种的重要值组成的48×194维矩阵和48个样方及6个环境因子组成的48×6维矩阵进行分析,得出排序结果。
-
野外调查共记录种子植物193种,隶属66科129属。科物种组成占总物种数比例分别为壳斗科Fagaceae 28.79% (19种)、樟科Lauraceae 27.27% (18种)、山茶科 Theaceae 13.64% (9种)、杜鹃花科Ericaceae 13.64% (9种)、蔷薇科Rosaceae 13.64% (9种)、冬青科Aquifoliaceae 9.09% (6种)、禾本科Gramineae 9.09% (6种)、茜草科Rubiaceae 9.09% (6种)、金粟兰科Chloranthaceae 7.58% (5种)、百合科 Liliaceae 7.58% (5种)。根据表2中群落不同层次物种重要值的排序可知(排名前20),乔木层优势物种主要为杉木、木荷、黄山松、青冈、甜槠、红楠、少叶黄杞和拟赤杨。灌木层优势种为杜茎山、草珊瑚、隔药柃和映山红。草本层优势种为蕨、芒萁、黑足鳞毛蕨、中华薹草、白茅、江南卷柏和荩草。
表 2 浙江庆元巾子峰国家森林公园植被群落优势种数量特征
Table 2. Quantitative characteristics of dominant species of the plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
种名 科名 生活型 重要值/% 蕨 Pteridum aquilinum var. latiusculum 蕨科Pteridiaceae 草本 1 130.85 芒萁 Dicranopteris dichotoma 里白科Gleicheniaceae 草本 1 061.41 杉木 Cunninghamia lanceolata 杉科Taxodiaceae 乔木 925.88 木荷 Schima superba 山茶科Theaceae 乔木 621.38 黄山松 Pinus taiwanensis 松科Pinaceae 乔木 515.13 杜茎山 Maesa japonica 紫金牛科Myrsinaceae 灌木 435.24 黑足鳞毛蕨 Dryopteris fuscipes 鳞毛蕨科Dryopteridaceae 草本 431.89 中华薹草 Carex chinensis 莎草科Cyperaceae 草本 407.44 毛竹 Phyllostachys edulis 禾本科Gramineae 乔木状禾本科植物 343.66 青冈 Cyclobalanopsis glauca 壳斗科Fagaceae 乔木 334.72 甜槠 Castanopsis eyrei 壳斗科 乔木 314.95 白茅 Imperata cylindrica 禾本科 草本 269.49 红楠 Machilus thunbergii 樟科Lauraceae 乔木 247.42 草珊瑚 Sarcandra glabra 金粟兰科Chloranthaceae 灌木 240.64 少叶黄杞 Engelhardtia fenzlii 胡桃科Juglandaceae 乔木 230.03 隔药柃 Eurya muricata 山茶科 灌木 221.27 江南卷柏 Selaginella moellendorffii 卷柏科Selaginellaceae 草本 213.64 拟赤杨 Alniphyllum fortunei 安息香科Styracaceae 乔木 200.82 映山红 Rhododendron simsii 杜鹃花科Ericaceae 灌木 195.90 荩草 Arthraxon hispidus 禾本科 草本 183.05 树参 Dendropanax dentiger 五加科Araliaceae 灌木 167.01 石栎 Lithocarpus glaber 壳斗科 乔木 158.91 马尾松 Pinus massoniana 松科 乔木 158.34 山莓 Rubus corchorifolius 蔷薇科Rosaceae 灌木 142.24 矩形叶鼠刺 Itea chinensis var. oblonga 虎耳草科Saxifragaceae 灌木 141.90 虎皮楠 Daphniphyllum oldhami 虎皮楠科Daphniphyllaceae 乔木 140.93 薯豆 Elacocarpus japonicus 杜英科Elaeocarpaceae 乔木 138.80 朱砂根 Ardisia crenata 紫金牛科 灌木 135.49 马银花 Rhododendron ovatum 杜鹃花科 灌木 134.90 毛花连蕊茶 Camellia fraterna 山茶科 灌木 134.69 少花万寿竹 Disporum uniflorum 秋水仙科Colchicaceae 草本 132.29 冬青 Ilex chinensis 冬青科Aquifoliaceae 乔木 125.19 宜昌荚蒾 Viburnum erosum 忍冬科Caprifoliaceae 灌木 123.84 乌药 Lindera aggregata 樟科 灌木 116.19 多花黄精 Polygonatum cyrtonema 百合科Liliaceae 草本 109.51 中华野海棠 Bredia sinensis 野牡丹科Melastomataceae 灌木 108.62 里白 Hicriopteris glauca 里白科Gleicheniaceae 草本 107.89 藜芦 Veratrum nigrum 百合科 草本 106.03 刨花楠 Machilus pauhoi 樟科 乔木 103.29 细枝柃 Eurya loquaiana 山茶科 灌木 99.45 栲 Castanopsis fargesii 壳斗科 乔木 98.24 细叶青冈 Cyclobalanopsis gracilis 壳斗科 乔木 94.52 苦茶槭 Acer ginnala subsp. theiferum 槭树科Aceraceae 灌木 93.11 锥栗 Castanea henryi 壳斗科 乔木 86.52 黄绒润楠 Machilus grijsii 山矾科Symplocaceae 乔木 84.86 米槠 Castanopsis carlesii 樟科 乔木 77.54 老鼠矢 Symplocos stellaris 壳斗科 乔木 74.62 刺毛杜鹃 Rhododendron championiae 杜鹃花科 灌木 74.18 蓝果树 Nyssa sinensis 蓝果树科Nyssaceae 乔木 68.94 江南越橘 Vaccinium mandarinorum 杜鹃花科 灌木 68.88 赤楠 Syzygium buxifolium 桃金娘科Ericaceae 灌木 63.31 五针松 Pinus parviflora 松科 乔木 62.94 檵木 Loropetalum chinensis 金缕梅科Hamamelidaceae 灌木 61.97 光皮桦 Betula luminifera 桦木科Betulaceae 乔木 59.91 地菍 Melastoma dodecandrum 野牡丹科 草本 58.65 贯众 Dryopteris setosa 鳞毛蕨科 草本 57.01 日本蛇根草 Ophiorrhiza japonica 茜草科Rubiaceae 草本 56.71 豹皮樟 Litsea coreana var. sinensis 樟科 灌木 56.41 宽叶金粟兰 Chloranthus henryi 金粟兰科 草本 53.94 大叶青冈 Cyclobalanopsis jenseniana 壳斗科 乔木 53.88 说明:物种重要值为48个样方的累加,包括乔木层、灌木层和草本层重要值排序为前60的物种[5] -
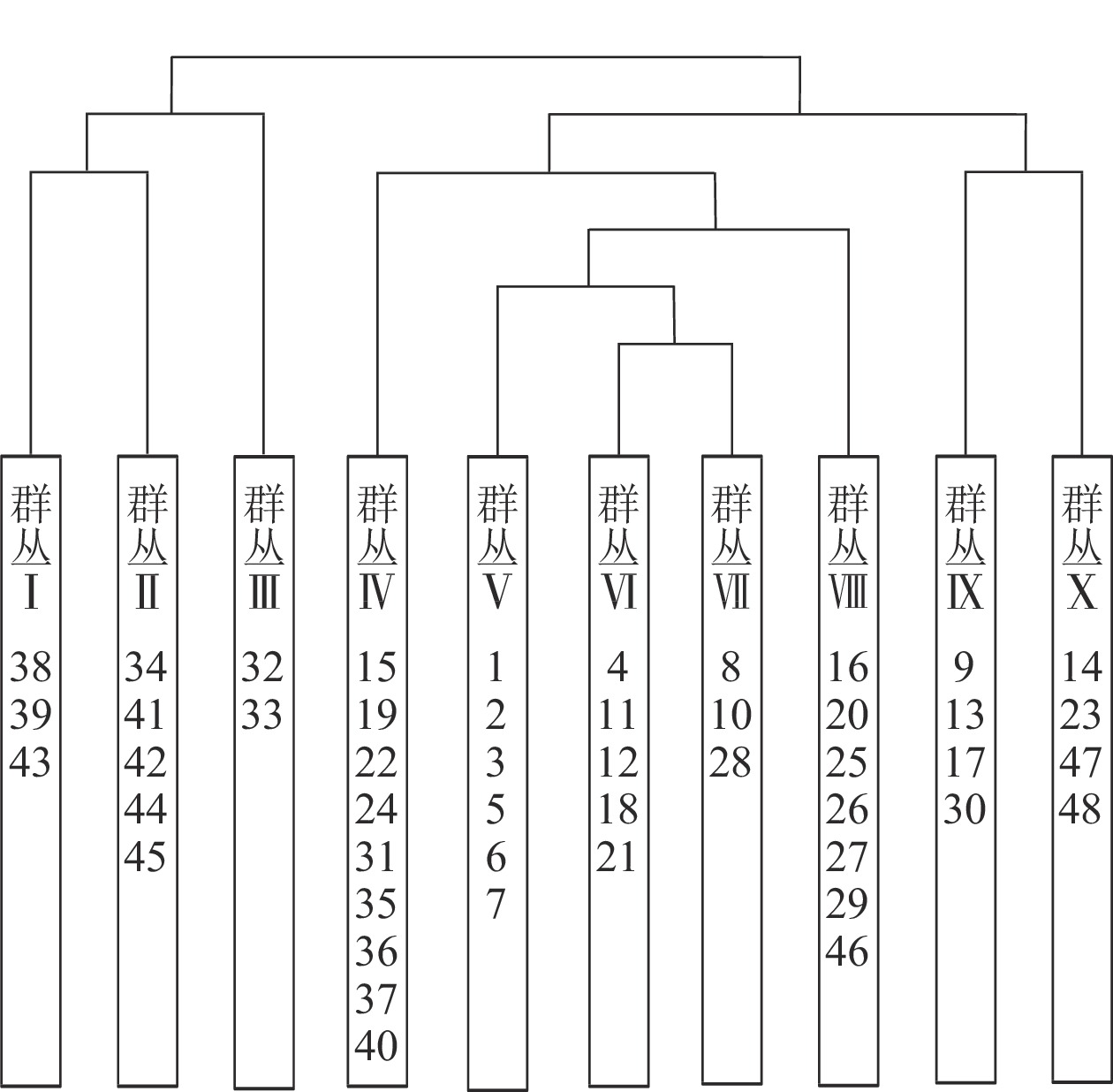
根据中国植被群落的分类原则[12]和双向指示种分析分类结果,将48个样方分成10个群丛(图1),采用生境指示种及群丛优势种的方式对群丛进行命名。各群丛环境因子描述见表3。
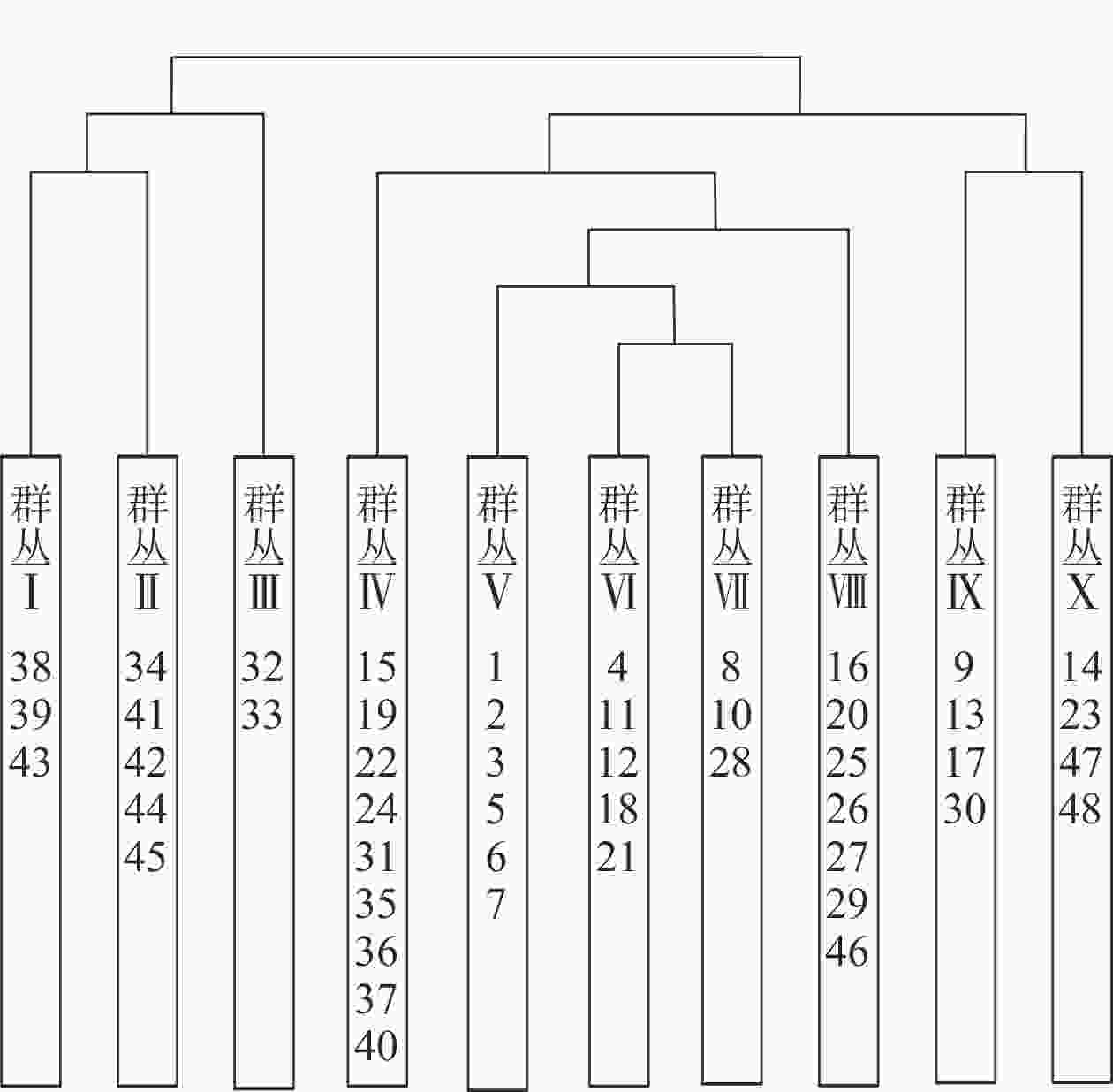
图 1 浙江庆元巾子峰国家森林公园植被群落的双向指示种分析分类结果树状图
Figure 1. Dendrogram of TWINSPAN classified result in Jinzifeng Forest Park of Qingyuan, Zhejiang
群丛Ⅰ:黄山松-映山红-东方古柯-少花万寿竹群丛Association Pinus taiwanensis-Rhododendron simsii-Erythroxylum sinense-Disporum uniflorum,包括38、39、43号样方。乔木层优势种为黄山松,主要伴生种为木荷。灌木层优势种为映山红、东方古柯,主要伴生种为宜昌荚蒾、麂角杜鹃Rhododendron latoucheae。草本层优势种为少花万寿竹、藜芦。
群丛Ⅱ:黄山松-木荷-映山红-多花黄精群丛Association Pinus taiwanensis-Schima superba-Rhododendron simsii-Polygonatum cyrtonema,包括34、41、42、44、45号样方。乔木层优势种为黄山松、木荷、青冈、甜槠,主要伴生种为浙闽樱Prunus schneideriana、云和新木姜子Neolitsea aurata var. paraciculata、苦茶槭、湖北鹅耳枥Carpinus hupeana、多脉鹅耳枥Carpinus polyneura。灌木层优势种为映山红、麂角杜鹃,主要伴生种为映山红、江南越橘、中华野海棠。草本层优势种为多花黄精、少花万寿竹。
群丛Ⅲ:甜槠-木荷-黄丹木姜子-少花万寿竹群丛Association Castanopsis eyrei-Schima superba-Litsea elongata-Disporum uniflorum,包括32、33号样方。乔木层优势种为甜槠、木荷、青冈,主要伴生种为马银花、多脉鹅耳枥、江南油杉Keteleeria fortunei var. cyclolepis、浙江樟Cinnamomum chekiangense、红楠、四照花Cornus kousa subsp. chinensis。灌木层优势种为黄丹木姜子Litsea elongata,主要伴生种为毛花连蕊茶、隔药柃、江南越橘。草本层优势种为少花万寿竹、藜芦、蕨。
群丛Ⅳ:杉木-毛花连蕊茶-江南卷柏群丛Association Cunninghamia lanceolata-Camellia fraterna-Selaginella moellendorffii,包括15、19、22、24、31、35、36、37、40号样方。乔木层优势种为杉木,主要伴生种为木荷、拟赤杨、虎皮楠、刨花楠、光皮桦、青冈、薯豆、少叶黄杞。灌木层优势种为毛花连蕊茶,主要伴生种为细枝柃、杜茎山、隔药柃、矩形叶鼠刺、毛冬青Ilex pubescens、黄绒润楠、乌药、赤楠、山鸡椒Litsea cubeba、刺毛杜鹃、豹皮樟、山莓、映山红。草本层优势种为江南卷柏、蕨、赤车Pellionia radicans,主要伴生种为贯众、黑足鳞毛蕨、芒萁、日本蛇根草。
群丛Ⅴ:杉木-木荷-杜茎山-芒萁群丛Association Cunninghamia lanceolata-Schima superba-Maesa japonica-Dicranopteris pedata,包括1、2、3、5、6、7号样方。乔木层优势种为杉木、木荷、薯豆、拟赤杨,主要伴生种为少叶黄杞、细叶青冈、树参、青冈、刨花楠、米槠、红楠。灌木层优势种为杜茎山、刺毛杜鹃,主要伴生种为江南越橘、草珊瑚、矩形叶鼠刺、檵木、黄绒润楠。草本层优势种为芒萁、黑足鳞毛蕨,主要伴生种为蕨、赤车、江南卷柏、中华薹草。
群丛Ⅵ:青冈-木荷-细枝柃-黑足鳞毛蕨群丛Association Cyclobalanopsis glauca-Schima superba-Eurya loquaiana-Dryopteris fuscipes,包括4、11、12、18、21号样方。乔木层优势种为青冈、木荷、锥栗,主要伴生种为冬青、虎皮楠、少叶黄杞、米槠、马银花。灌木层优势种为细枝柃,主要伴生种为杜茎山、石楠 Photinia serrulata、草珊瑚、乌药、狗骨柴Diplospora dubia、赤楠、矩形叶鼠刺、刺毛杜鹃、江南越橘。草本层优势种为黑足鳞毛蕨、芒萁、中华薹草。
群丛Ⅶ:拟赤杨-虎皮楠-山矾-日本蛇根草群丛Association Alniphyllum fortunei-Daphniphyllum oldhami-Symplocos sumuntia-Ophiorrhiza japonica,包括8、10、28号样方。乔木层优势种为拟赤杨、虎皮楠,主要伴生种为米槠、树参、光皮桦、少叶黄杞、木荷、马银花。灌木层优势种为山矾Symplocos sumuntia,主要伴生种为杜茎山、矩形叶鼠刺、黄绒润楠、草珊瑚、乌药、隔药柃。草本层优势种为日本蛇根草、黑足鳞毛蕨,主要伴生种为中华薹草、蕨、江南卷柏。
群丛Ⅷ:刨花楠-隔药柃-蕨群丛Association Machilus pauhoi-Eurya muricata-Pteridium aquilinum var. latiusculum,包括16、20、25、26、27、29、46号样方。乔木层优势种为刨花楠、甜槠、栲,主要伴生种为少叶黄杞、深山含笑Michelia maudiae、青冈、大叶青冈、冬青、虎皮楠、锥栗。灌木层优势种为隔药柃,主要伴生种为毛花连蕊茶、细枝柃、山矾、乌药、黄绒润楠、石楠、矩形叶鼠刺、赤楠、豹皮樟、乌冈栎Quercus phillyreoides。草本层优势种为蕨,主要伴生种为中华薹草、江南卷柏、贯众、地菍。
群丛Ⅸ:马尾松-木荷-草珊瑚-中华薹草群丛Association Pinus massoniana-Schima superba-Sarcandra glabra-Carex chinensis,包括9、13、17、30号样方。乔木层优势种为马尾松、木荷、青冈,主要伴生种为少叶黄杞、拟赤杨、虎皮楠、锥栗、细叶青冈、红楠、蓝果树。灌木层优势种为草珊瑚,主要伴生种为檵木、黄绒润楠、朱砂根、映山红、江南越橘、隔药柃、山莓、老鼠矢。草本层优势种为中华薹草、芒萁、日本蛇根草。
群丛Ⅹ:毛竹-山鸡椒-地菍群丛Association Phyllostachys edulis-Litsea cubeba-Melastoma dodecandrum,包括14、23、47、48号样方。乔木层优势种为毛竹,主要伴生种为杉木、拟赤杨、马尾松。灌木层优势种为山鸡椒,主要伴生种为隔药柃、乌药、黄绒润楠、檵木、胡枝子Lespedeza bicolor、老鼠矢。草本层优势种为地菍,主要伴生种为芒萁、杏香兔耳风Ainsliaea fragrans、蕨、中华薹草。
表 3 浙江庆元巾子峰国家森林公园植被群落各群丛环境因子描述
Table 3. Description of environmental factors for each association in Jinzifeng Forest Park of Qingyuan, Zhejiang
群丛 群丛名称 海拔/m 坡向 坡度/(°) 土壤类型 土壤温度/℃ 土壤湿度/% Ⅰ 黄山松-映山红-东方古柯-少花万寿竹群丛 1 200~1 500 北 10~30 黄壤 15~20 16~19 Ⅱ 黄山松-木荷-映山红-多花黄精群丛 1 200~1 400 西北、东北 25~35 黄壤 15~22 10~32 Ⅲ 甜槠-木荷-黄丹木姜子-少花万寿竹群丛 1 000~1 200 东 30~35 黄壤 24~25 12~16 Ⅳ 杉木-毛花连蕊茶-江南卷柏群丛 600~1 300 东、西 10~30 红壤、黄壤 17~25 25~52 Ⅴ 杉木-木荷-杜茎山-芒萁群丛 500~800 东北、东南 20~30 红壤 17~22 18~61 Ⅵ 青冈-木荷-细枝柃-黑足鳞毛蕨群丛 500~800 东、东南 10~30 红壤 18~26 18~28 Ⅶ 拟赤杨-虎皮楠-山矾-日本蛇根草群丛 500~800 东 25~30 红壤 19~23 19~40 Ⅷ 刨花楠-隔药柃-蕨群丛 500~900 东南、南 25~45 红壤 20~23 15~52 Ⅸ 马尾松-木荷-草珊瑚-中华薹草群丛 500~900 西 10~30 红壤 20~24 16~39 Ⅹ 毛竹-山鸡椒-地菍群丛 500~800 西南、西 5~25 红壤 21~25 23~42 -
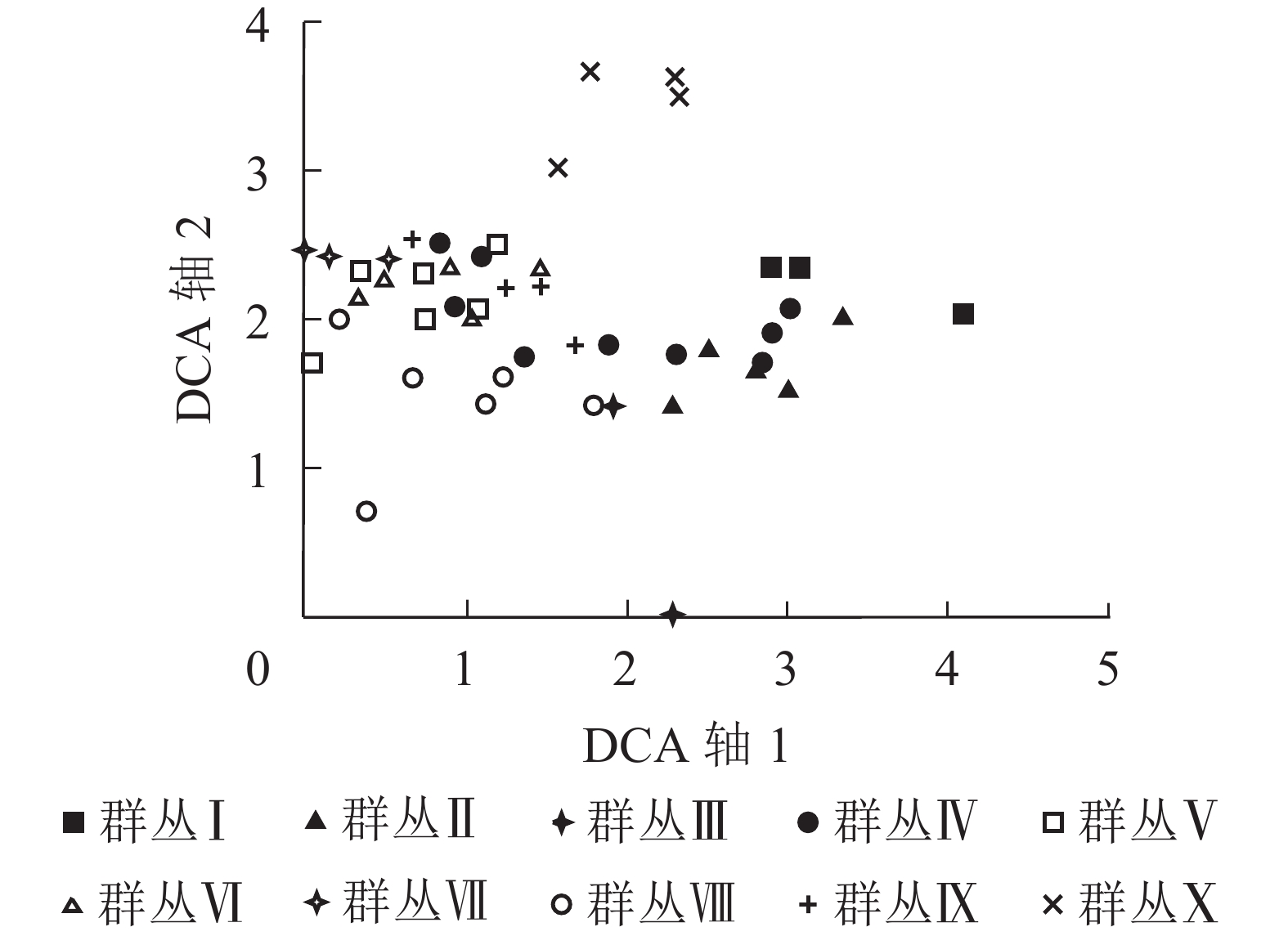
48个样方的去趋势对应分析结果表明:4个排序轴的特征值分别为0.549、0.400、0.295、0.225,因前2个排序轴的特征值比较大,能显示出一定的生态意义,所以采用前2个排序轴的数据分别作二维排序图(图2)。从图2可以看出:去趋势对应分析排序的横轴和纵轴基本反映了不同的生态意义。第1排序轴(横轴)反映了各群丛所在环境的海拔和土壤类型梯度,即沿第1排序轴从左往右,随着海拔高度逐渐升高,土壤类型由红壤变为黄壤;第2排序轴(纵轴)反映了各群丛所在的坡度和土壤情况变化,沿第2排序轴从下往上,坡度由陡坡转为缓坡,土壤温度逐渐降低,土壤湿度逐渐升高。
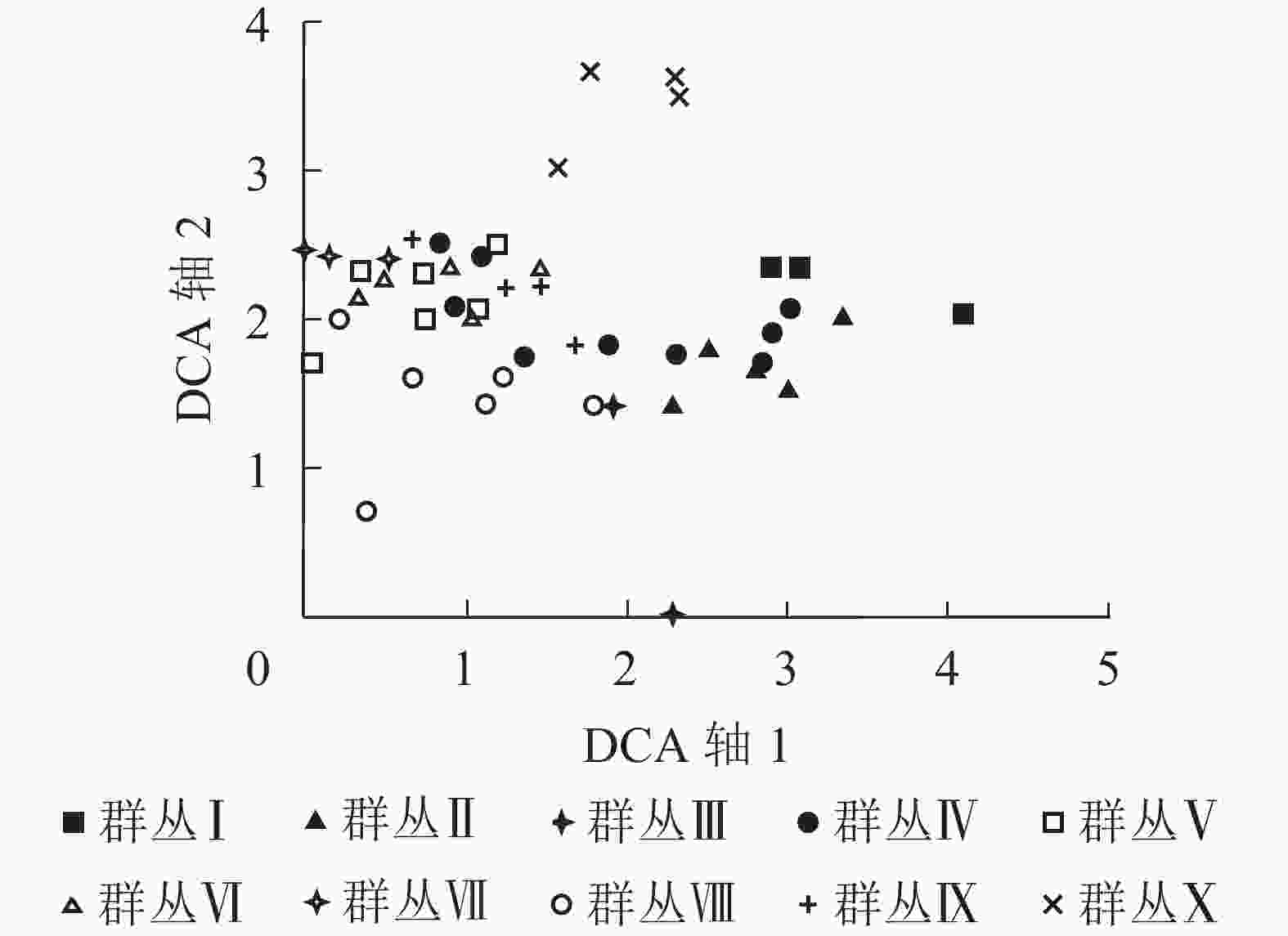
图 2 浙江庆元巾子峰国家森林公园植被群落样方的去趋势对应分析排序图
Figure 2. DCA two-dimensional ordination diagram for samples of plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
结合双向指标种分析的分类结果对排序图进行分析,发现不同的森林群落类型在排序图中呈现出一定规律的分布。黄山松、黄山松-木荷群丛分布在海拔较高的中山带阴坡、半阴坡,土壤类型为黄壤,土壤温度和湿度均较低,位于排序图的右下方。甜槠-木荷群丛分布在中山半阳坡、半阴坡,坡度较陡,土壤类型为黄壤,土壤温度较高,土壤湿度较小,位于排序图的左下方。杉木-毛花连蕊茶群丛分布在中山带半阳坡、半阴坡,土壤类型为红壤或黄壤,位于排序图的中心。杉木-木荷、马尾松-木荷群丛分布在低山半阳坡、半阴坡,土壤类型为红壤,位于排序图的中心靠左位置。青冈-木荷群丛分布在低山半阳坡、半阴坡,土壤类型为红壤,位于排序图的左方。拟赤杨-虎皮楠群丛分布在低山半阳坡,土壤类型为红壤,位于排序图的左上方。刨花楠群丛分布在低山阳坡、半阳坡,土壤类型为红壤,位于排序图的左下方。毛竹群丛分布在低山半阳坡,土壤类型为红壤,位于排序图的上方。
-
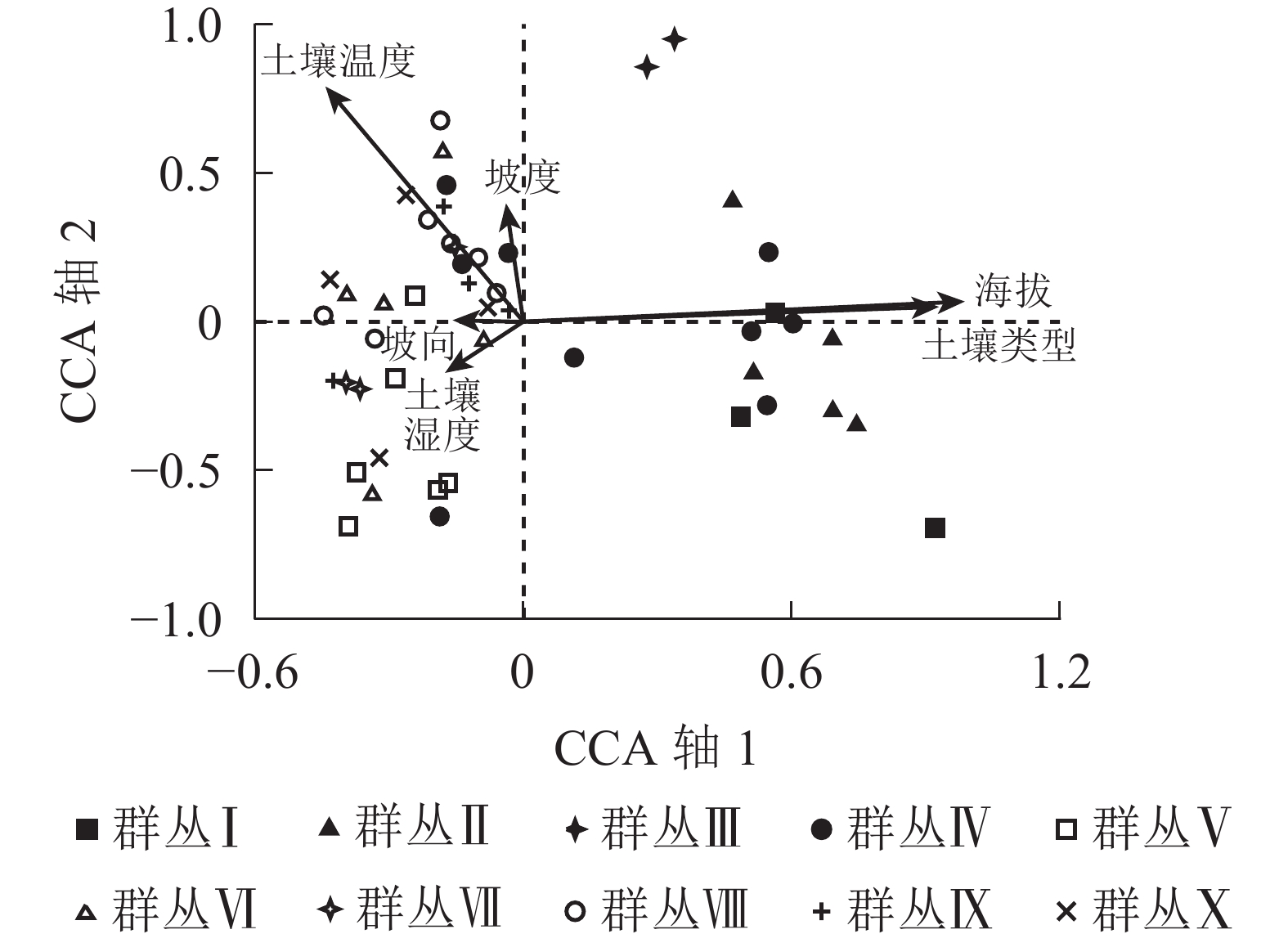
浙江庆元巾子峰国家森林公园植被群落物种重要值和样方矩阵的典范对应分析排序所有轴的梯度长度最大为4.115,所以选择单峰模型。蒙特卡罗检验结果显示:所有典范排序轴通过统计检验(F=1.629, P=0.002),说明所选择的环境因子对植被群落物种分布具有显著影响。由表4可知:典范对应分析排序轴中前2轴累计解释了物种-环境因子关系的55.2%,说明典范对应分析前2轴能较好地反映森林植被与环境因子之间的关系。根据环境因子与排序轴的相关性大小可知,海拔、土壤类型与第1排序轴呈极显著正相关(P<0.01),土壤温度与第1排序轴呈显著负相关(P<0.05),相关性从大到小依次为海拔、土壤类型、土壤温度,表明第1排序轴反映的是海拔和土壤类型的梯度变化。土壤温度与第2排序轴呈极显著负相关(P<0.01),与坡度呈显著正相关(P<0.05),表明第2排序轴反映了土壤温度的梯度变化。
表 4 浙江庆元巾子峰国家森林公园植被群落的典范对应分析排序
Table 4. The CCA analysis of characteristics of the plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
排序轴 土壤湿度 土壤温度 坡度 坡向 海拔 土壤类型 特征值 物种-环境
相关性物种数据累计
贡献率/%物种-环境因子关系
数据累计贡献率/%第1排序轴 −0.171 −0.432* −0.038 −0.154 0.962** 0.906** 0.497 0.975 7.3 38.0 第2排序轴 −0.152 −0.699** 0.352* 0.003 0.059 0.044 0.224 0.885 10.6 55.2 第3排序轴 −0.068 0.226 −0.506 0.347* −0.015 0.152 0.184 0.818 13.3 69.2 第4排序轴 0.661** 0.140 −0.304* 0.240 −0.022 −0.163 0.160 0.839 15.7 81.5 说明:**P<0.01, *P<0.05。所有典范轴的显著性监测F=1.629,P=0.002 样方-环境因子的典范对应分析排序图较为直观地呈现了植被群落各群丛类型分布与环境因子的关系(图3),带箭头射线代表环境因子,射线长度表示植被群落与环境因子相关性的大小,射线所在象限表示环境因子与排序轴相关性的正负。典范对应分析排序第1轴代表了海拔和土壤类型,从左往右海拔逐渐升高,土壤由红壤转变为黄壤;第2排序轴代表了土壤温度,从下往上土壤温度逐渐增加。群丛Ⅰ和群丛Ⅱ的分布与海拔和土壤类型关系比较密切,分布在海拔最高的区域,且均是黄壤。群丛Ⅲ分布在较高海拔。群丛Ⅳ分布比较宽泛,在海拔600~1 300 m均有分布,土壤有红壤还有黄壤。群丛Ⅴ和群丛Ⅶ与土壤湿度关系较密切。群丛Ⅷ、群丛Ⅸ和群丛Ⅹ则与土壤温度和土壤湿度关系密切。
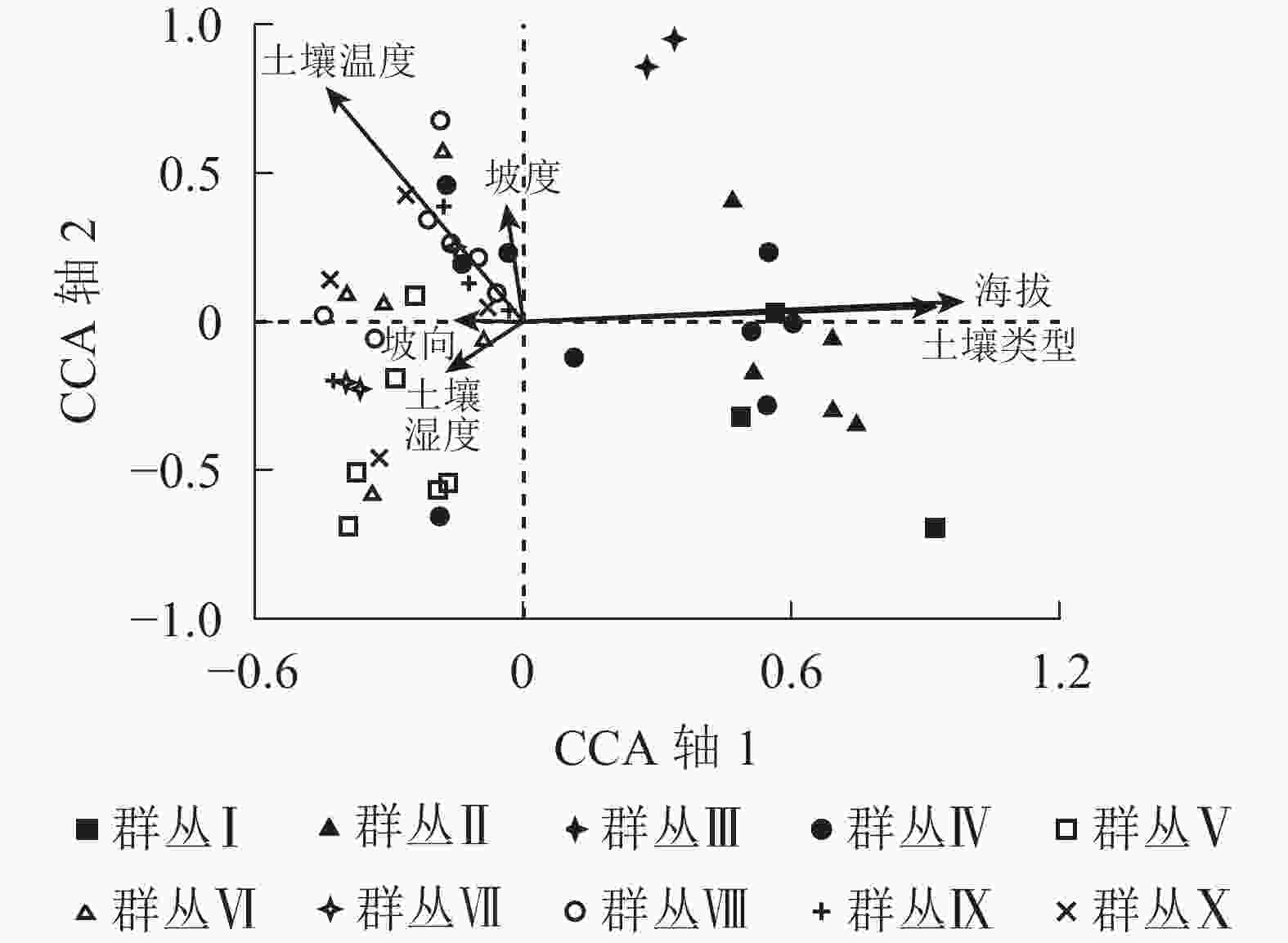
图 3 浙江庆元巾子峰国家森林公园植被群落样方与环境因子的典范对应分析排序图
Figure 3. CCA diagram for samples and environmental variables of the plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
优势乔木树种对群落结构和群落环境的形成有明显控制作用,个体数量不一定很多,但却能决定群落结构和内部环境条件,是植被群落的建造者。对森林群落中20个优势乔木树种进行典范对应分析排序,排序结果如图4所示。海拔、土壤类型、土壤湿度和坡度对物种分布起决定作用。黄山松分布在中山带阴坡,土壤类型为黄壤,位于排序图的右方。甜槠、锥栗分布在低山半阳坡,土壤类型为黄壤,位于排序图的上方。杉木分布在低山、中山带半阳坡、半阴坡,土壤类型为红壤或黄壤,位于排序图的中间靠上位置。青冈、木荷分布在低山、中山带半阳坡、半阴坡,土壤类型为红壤或黄壤,位于排序图的中间靠下位置。拟赤杨、石栎分布在低山半阳坡,土壤类型为红壤,位于排序图的中间靠左位置。马尾松分布在低山半阳坡,土壤类型为红壤,位于排序图的左方。刨花楠、虎皮楠、栲、米槠、红楠、冬青、薯豆、少叶黄杞、细叶青冈、黄绒润楠分布在低山半阳坡、半阴坡,土壤类型为红壤,位于排序图的左下方。毛竹分布在低山阳坡,土壤类型为红壤,位于排序图的左上方。
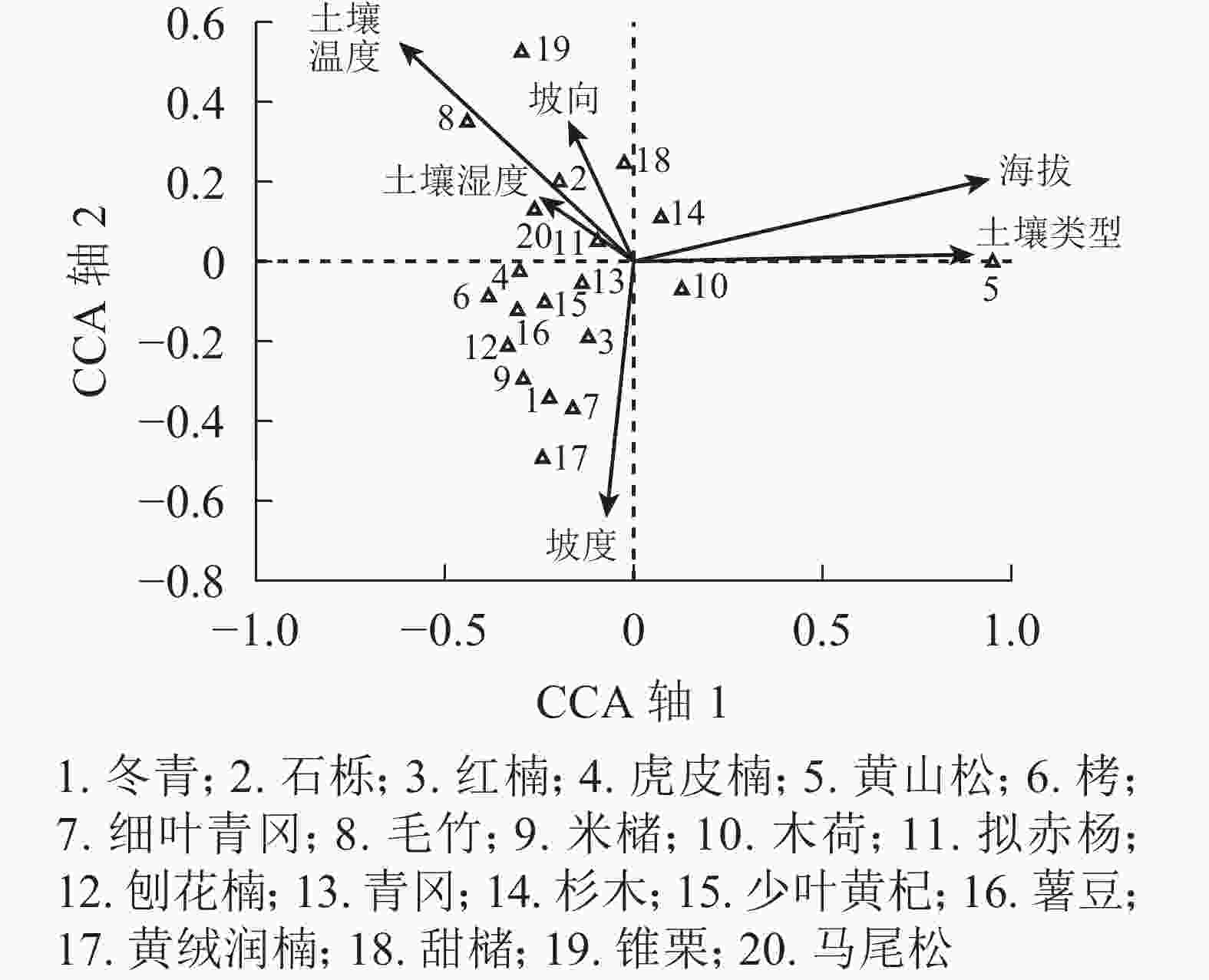
图 4 浙江庆元巾子峰国家森林公园20个优势树种的典范对应分析排序图
Figure 4. CCA diagram for trees and environmental variables in Jinzifeng Forest Park of Qingyuan, Zhejiang
-
本研究根据中国植被群落的分类原则和双向指标种分析方法,将浙江庆元巾子峰国家森林公园内48个样方分成10个群丛。对10个群丛及其相应样方进行去趋势对应分析排序。结果表明:各群丛在排序图中的分布均较为集中,说明去趋势对应分析排序结果与双向指标种分析分类结果较为一致,综合应用双向指标种分析分类和去趋势对应分析排序能够更好地解释森林群落之间的差异性和连续性,这与以往的研究结果相同[13-17]。双向指标种分析数量分类是依据指示种和地区分布差异划分的,而去趋势对应分析是依据排序轴的综合信息,将样方或植物物种排列在一定的空间[2]。去趋势对应分析排序轴能够反映一定的生态梯度。本研究横轴主要反映了样方的海拔高度和土壤类型,纵轴主要反映了样方的土壤温度和坡度。各群丛在排序图中的位置能有效反映出各群丛的环境特征及群丛间的连续分布关系,可以看出植被类型间出现了交错现象,表明植被类型与环境因子间关系比较复杂,植被类型的分布受海拔、土壤环境、坡度、坡向等多种环境因子的共同影响。
典范对应分析排序与去趋势对应分析排序相比,去趋势对应分析排序仅使用物种数据,在反映样方之间或物种之间的关系上具有较大优势,但不能较好地反映与环境因子之间的相关性;典范对应分析同时使用了物种数据和环境因子数据,能较好地表达群落的环境梯度,排序轴能更好地对环境因子进行解释[2, 4, 18]。本研究划分的10种群丛类型与样方在典范对应分析排序图上的分布格局基本吻合,能较好地揭示植被类型与环境因子之间的关系,反映特定的植被类型对不同的环境因子差异具有指示作用。从典范对应分析排序图上可以看出:海拔、土壤类型和土壤温度这3个主要环境因子共同影响了植被的分布。物种丰富度海拔梯度格局及其形成机制一直是宏观生态学研究的热点。研究表明:海拔梯度是影响植被群落物种分布格局的主要控制因子[4,19]。因为通过海拔梯度的变化可以改变局部水分状况、土壤理化性质、干扰程度等其他生态环境,直接或间接地对群落分布格局产生影响[20-22]。本研究也证明海拔对山地植被分布具有决定性作用,海拔是影响巾子峰国家森林公园植被群落分布的最重要环境因子。
综上所述,本研究中影响植被群落分布最重要的环境因子是海拔,其次为土壤类型、土壤温度和坡度,土壤湿度和坡向对植被群落分布的影响相对较小。对20个优势树种进行的典范对应分析排序发现,优势树种的分布特征与各群丛及相应样方的分布特征具有较好的相似性,影响各优势树种分布最重要的环境因子也是海拔,其次为土壤类型、土壤温度和坡度。这与各群丛及相应样方典范对应分析排序结果一致,说明各优势树种的分布在一定程度上决定了各群丛类型的分布。
Numerical classification and ordination of vegetation communities in Jinzifeng National Forest Park, Qingyuan, Zhejiang
-
摘要:
目的 了解浙江省庆元县巾子峰国家森林公园中植被群落与环境因子之间的关系。 方法 采用样方调查法调查庆元巾子峰国家森林公园中48个样方的植被群落,并进行数量分类和排序,分析群落类型、分布特征及其与环境因子的相关关系。 结果 ①共调查记录植物66科129属193种。植物物种数较多的科分别为壳斗科Fagaceae(19种)、樟科 Lauraceae(18种)、山茶科 Theaceae(9种)、杜鹃花科Ericaceae(9种)、蔷薇科Rosaceae(9种)、冬青科Aquifoliaceae(6种)、禾本科Gramineae(6种)、茜草科Rubiaceae(6种)、金粟兰科Chloranthaceae(5种)和百合科 Liliaceae(5种)。乔木层优势物种主要为杉木Cunninghamia lanceolata、木荷Schima superba、黄山松Pinus taiwanensis、青冈Cyclobalanopsis glauca、甜槠Castanopsis eyrei、红楠Machilus thunbergii、少叶黄杞Engelhardia fenzelii和拟赤杨Alniphyllum fortunei。灌木层优势种为杜茎山Maesa japonica、草珊瑚Sarcandra glabra、隔药柃Eurya muricata和映山红Rhododendron simsii。草本层优势种为蕨Pteridum aquilinum var. latiusculum、芒萁Dicranopteris dichotoma、黑足鳞毛蕨Dryopteris fuscipes、中华薹草Carex chinensis、白茅Imperata cylindrica、江南卷柏Selaginella moellendorffii和荩草Arthraxon hispidus。②双向指示种分析将植被群落分成10个群丛类型。③10个群丛在去趋势对应分析排序图中呈现有规律的分布,反映了较好的环境梯度。④典范对应分析排序结果表明:影响各群丛分布的最重要环境因子是海拔,其次为土壤类型、土壤温度和坡度;各优势树种的分布特征与各群丛的分布特征具有较好的相似性。 结论 海拔是影响庆元巾子峰国家森林公园植被群落分布的最重要环境因子,海拔对山地植被群落分布具有决定性作用。图4表4参22 Abstract:Objective The objective is to analyze the relationship between vegetation community and environmental factors in Jingzifeng National Forest Park of Qingyuan County, Zhejiang Province. Method Data of 48 quadrats in park were classified and sorted, and the relationship between forest community types, distribution characteristics and environmental factors was analyzed based on these data. Result (1)A total of 193 species belonging to 129 genera and 66 families were recorded in 48 plots. The families with abundant plant species were Fagaceae (19 species), Lauraceae (18 species), Theaceae (9 species), Ericaceae (9 species), Rosaceae (9 species), Aquifoliaceae (6 species), Gramineae (6 species), Rubiaceae (6 species), Chloranthaceae (5 species) and Liliaceae (5 species). The dominant species in tree layer included Cunninghamia lanceolata, Schima superba, Pinus taiwanensis, Cyclobalanopsis glauca, Castanopsis eyrei, Machilus thunbergii, Engelhardia fenzelii and Alniphyllum fortunei. The dominant species in shrub layer were Maesa japonica, Sarcandra glabra, Eurya muricata, and Rhododendron simsii. The dominant species in herbaceous layer were Pteridum aquilinum var. latiusculum, Dicranopteris dichotoma, Dryopteris fuscipes, Carex chinensis, Imperata cylindrica, Selaginella moellendorfii and Arthraxon hispidus. (2) TWINSPAN divided the vegetation community into 10 cluster types. (3) The distribution of the 10 clusters in DCA ordination diagram was regular, which reflected a good environmental gradient. (4) CCA ordination results showed that the most important environmental factor affecting the distribution of each cluster was altitude, followed by soil type, soil temperature and slope. The distribution characteristics of the dominant tree species were similar to those of each cluster. Conclusion Altitude is the most important environmental factor affecting the distribution of vegetation community in Jinzifeng National Park, and the altitude plays a decisive role in the distribution of mountain vegetation community. [Ch, 4 fig. 4 tab. 22 ref.] -
图 1 浙江庆元巾子峰国家森林公园植被群落的双向指示种分析分类结果树状图
图中数字为样方号
Figure 1 Dendrogram of TWINSPAN classified result in Jinzifeng Forest Park of Qingyuan, Zhejiang
图 2 浙江庆元巾子峰国家森林公园植被群落样方的去趋势对应分析排序图
Figure 2 DCA two-dimensional ordination diagram for samples of plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
图 3 浙江庆元巾子峰国家森林公园植被群落样方与环境因子的典范对应分析排序图
Figure 3 CCA diagram for samples and environmental variables of the plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
图 4 浙江庆元巾子峰国家森林公园20个优势树种的典范对应分析排序图
Figure 4 CCA diagram for trees and environmental variables in Jinzifeng Forest Park of Qingyuan, Zhejiang
表 1 浙江庆元巾子峰国家森林公园样方环境因子
Table 1. Environmental factors of plots in Jinzifeng Forest Park, Qingyuan, Zhejiang
样方号 群落郁闭度 海拔/m 坡位 坡度/(°) 坡向 北纬(N) 东经(E) 土壤类型 1 0.85 554 下坡位 20 西南 27°41'04.026" 119°00'42.080" 红壤 2 0.90 550 下坡位 30 东北 27°41'07.635" 119°00'41.897" 红壤 3 0.85 540 中坡位 30 东 27°41'16.329" 119°00'40.030" 红壤 4 0.80 572 中坡位 30 东 27°41'00.230" 119°00'36.095" 红壤 5 0.85 657 中坡位 30 东 27°40'40.769" 119°00'26.647" 红壤 6 0.90 565 下坡位 35 东南 27°41'12.993" 119°00'40.357" 红壤 7 0.70 694 中坡位 30 西北 27°41'40.270" 119°01'06.858" 红壤 8 0.80 727 中坡位 30 东北 27°40'37.443" 119°00'23.501" 红壤 9 0.90 1 052 山脊 35 东 27°39'59.132" 118°59'59.641" 黄壤 10 0.50 628 中坡位 25 东南 27°39'51.851" 119°00'37.342" 黄壤 11 0.90 571 下坡位 25 东 27°41'00.880" 119°00'41.821" 红壤 12 0.90 538 下坡位 25 北 27°41'18.531" 119°00'39.928" 红壤 13 0.85 791 中坡位 30 西 27°41'27.328" 119°01'02.480" 红壤 14 0.80 780 上坡位 10 西 27°41'13.857" 119°00'57.785" 红壤 15 0.85 767 中坡位 25 东 27°41'33.945" 119°01'09.110" 红壤 16 0.60 750 下坡位 45 东北 27°41'37.304" 119°01'09.533" 红壤 17 0.70 832 中坡位 25 南 27°41'11.570" 119°01'07.716" 红壤 18 0.80 797 下坡位 25 西南 27°41'14.264" 119°00'58.968" 红壤 19 0.80 734 下坡位 30 北 27°40'32.842" 119°00'29.121" 红壤 20 0.80 821 谷底 30 东南 27°40'23.970" 119°00'33.259" 红壤 21 0.80 1 108 上坡位 35 东 27°39'59.474" 118°59'54.098" 黄壤 22 0.60 1 382 上坡位 35 西北 27°39'44.438" 118°59'42.841" 黄壤 23 0.80 776 下坡位 25 东北 27°40'33.656" 119°00'48.684" 红壤 24 0.95 731 中坡位 25 西南 27°42'06.523" 119°00'48.359" 红壤 25 0.80 518 中坡位 25 西 27°39'37.175" 119°00'54.164" 黄壤 26 0.70 613 中坡位 5 南 27°39'44.363" 119°00'35.918" 黄壤 27 0.80 562 中坡位 30 西 27°41'13.205" 119°00'41.127" 红壤 28 0.85 736 山脊 10 北 27°40'43.434" 119°00'39.118" 红壤 29 0.85 681 下坡位 25 东北 27°40'43.766" 119°00'48.120" 红壤 30 0.85 820 下坡位 30 西 27°40'25.046" 119°00'33.625" 红壤 31 0.85 1 250 山顶 30 西南 27°39'53.117" 118°59'49.230" 黄壤 32 0.60 1 317 上坡位 35 西北 27°39'46.784" 118°59'44.727" 黄壤 33 0.80 680 下坡位 30 东北 27°40'36.989" 119°00'52.495" 红壤 34 0.80 590 下坡位 25 东 27°40'55.924" 119°00'46.219" 红壤 35 0.80 747 下坡位 30 东南 27°40'31.064" 119°00'51.985" 红壤 36 0.70 720 下坡位 30 东 27°40'35.337" 119°00'55.786" 红壤 37 0.75 925 山脊 10 东 27°40'18.521" 119°00'25.013" 红壤 38 0.60 784 中坡位 25 西北 27°40'44.187" 119°00'56.161" 红壤 39 0.95 767 中坡位 25 西 27°41'39.635" 119°01'10.765" 红壤 40 0.70 1 148 山脊 25 东北 27°40'13.334" 118°59'41.175" 黄壤 41 0.85 1 178 上坡位 30 东 27°40'07.905" 118°59'34.997" 黄壤 42 0.85 1 197 上坡位 25 东 27°40'08.949" 118°59'29.214" 黄壤 43 0.95 1 203 上坡位 30 西 27°40'11.715" 118°59'22.209" 黄壤 44 0.95 1 194 上坡位 10 南 27°40'08.334" 118°59'37.403" 黄壤 45 0.85 1 198 上坡位 30 北 27°39'56.373" 118°59'33.218" 黄壤 46 0.85 1 231 山脊 25 南 27°39'54.912" 118°59'46.670" 黄壤 47 0.60 1 271 中坡位 25 东北 27°39'49.007" 118°59'47.710" 黄壤 48 0.60 1 413 上坡位 10 北 27°39'41.018" 118°59'42.226" 黄壤  下载: 导出CSV
下载: 导出CSV
表 2 浙江庆元巾子峰国家森林公园植被群落优势种数量特征
Table 2. Quantitative characteristics of dominant species of the plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
种名 科名 生活型 重要值/% 蕨 Pteridum aquilinum var. latiusculum 蕨科Pteridiaceae 草本 1 130.85 芒萁 Dicranopteris dichotoma 里白科Gleicheniaceae 草本 1 061.41 杉木 Cunninghamia lanceolata 杉科Taxodiaceae 乔木 925.88 木荷 Schima superba 山茶科Theaceae 乔木 621.38 黄山松 Pinus taiwanensis 松科Pinaceae 乔木 515.13 杜茎山 Maesa japonica 紫金牛科Myrsinaceae 灌木 435.24 黑足鳞毛蕨 Dryopteris fuscipes 鳞毛蕨科Dryopteridaceae 草本 431.89 中华薹草 Carex chinensis 莎草科Cyperaceae 草本 407.44 毛竹 Phyllostachys edulis 禾本科Gramineae 乔木状禾本科植物 343.66 青冈 Cyclobalanopsis glauca 壳斗科Fagaceae 乔木 334.72 甜槠 Castanopsis eyrei 壳斗科 乔木 314.95 白茅 Imperata cylindrica 禾本科 草本 269.49 红楠 Machilus thunbergii 樟科Lauraceae 乔木 247.42 草珊瑚 Sarcandra glabra 金粟兰科Chloranthaceae 灌木 240.64 少叶黄杞 Engelhardtia fenzlii 胡桃科Juglandaceae 乔木 230.03 隔药柃 Eurya muricata 山茶科 灌木 221.27 江南卷柏 Selaginella moellendorffii 卷柏科Selaginellaceae 草本 213.64 拟赤杨 Alniphyllum fortunei 安息香科Styracaceae 乔木 200.82 映山红 Rhododendron simsii 杜鹃花科Ericaceae 灌木 195.90 荩草 Arthraxon hispidus 禾本科 草本 183.05 树参 Dendropanax dentiger 五加科Araliaceae 灌木 167.01 石栎 Lithocarpus glaber 壳斗科 乔木 158.91 马尾松 Pinus massoniana 松科 乔木 158.34 山莓 Rubus corchorifolius 蔷薇科Rosaceae 灌木 142.24 矩形叶鼠刺 Itea chinensis var. oblonga 虎耳草科Saxifragaceae 灌木 141.90 虎皮楠 Daphniphyllum oldhami 虎皮楠科Daphniphyllaceae 乔木 140.93 薯豆 Elacocarpus japonicus 杜英科Elaeocarpaceae 乔木 138.80 朱砂根 Ardisia crenata 紫金牛科 灌木 135.49 马银花 Rhododendron ovatum 杜鹃花科 灌木 134.90 毛花连蕊茶 Camellia fraterna 山茶科 灌木 134.69 少花万寿竹 Disporum uniflorum 秋水仙科Colchicaceae 草本 132.29 冬青 Ilex chinensis 冬青科Aquifoliaceae 乔木 125.19 宜昌荚蒾 Viburnum erosum 忍冬科Caprifoliaceae 灌木 123.84 乌药 Lindera aggregata 樟科 灌木 116.19 多花黄精 Polygonatum cyrtonema 百合科Liliaceae 草本 109.51 中华野海棠 Bredia sinensis 野牡丹科Melastomataceae 灌木 108.62 里白 Hicriopteris glauca 里白科Gleicheniaceae 草本 107.89 藜芦 Veratrum nigrum 百合科 草本 106.03 刨花楠 Machilus pauhoi 樟科 乔木 103.29 细枝柃 Eurya loquaiana 山茶科 灌木 99.45 栲 Castanopsis fargesii 壳斗科 乔木 98.24 细叶青冈 Cyclobalanopsis gracilis 壳斗科 乔木 94.52 苦茶槭 Acer ginnala subsp. theiferum 槭树科Aceraceae 灌木 93.11 锥栗 Castanea henryi 壳斗科 乔木 86.52 黄绒润楠 Machilus grijsii 山矾科Symplocaceae 乔木 84.86 米槠 Castanopsis carlesii 樟科 乔木 77.54 老鼠矢 Symplocos stellaris 壳斗科 乔木 74.62 刺毛杜鹃 Rhododendron championiae 杜鹃花科 灌木 74.18 蓝果树 Nyssa sinensis 蓝果树科Nyssaceae 乔木 68.94 江南越橘 Vaccinium mandarinorum 杜鹃花科 灌木 68.88 赤楠 Syzygium buxifolium 桃金娘科Ericaceae 灌木 63.31 五针松 Pinus parviflora 松科 乔木 62.94 檵木 Loropetalum chinensis 金缕梅科Hamamelidaceae 灌木 61.97 光皮桦 Betula luminifera 桦木科Betulaceae 乔木 59.91 地菍 Melastoma dodecandrum 野牡丹科 草本 58.65 贯众 Dryopteris setosa 鳞毛蕨科 草本 57.01 日本蛇根草 Ophiorrhiza japonica 茜草科Rubiaceae 草本 56.71 豹皮樟 Litsea coreana var. sinensis 樟科 灌木 56.41 宽叶金粟兰 Chloranthus henryi 金粟兰科 草本 53.94 大叶青冈 Cyclobalanopsis jenseniana 壳斗科 乔木 53.88 说明:物种重要值为48个样方的累加,包括乔木层、灌木层和草本层重要值排序为前60的物种[5]
下载: 导出CSV
表 3 浙江庆元巾子峰国家森林公园植被群落各群丛环境因子描述
Table 3. Description of environmental factors for each association in Jinzifeng Forest Park of Qingyuan, Zhejiang
群丛 群丛名称 海拔/m 坡向 坡度/(°) 土壤类型 土壤温度/℃ 土壤湿度/% Ⅰ 黄山松-映山红-东方古柯-少花万寿竹群丛 1 200~1 500 北 10~30 黄壤 15~20 16~19 Ⅱ 黄山松-木荷-映山红-多花黄精群丛 1 200~1 400 西北、东北 25~35 黄壤 15~22 10~32 Ⅲ 甜槠-木荷-黄丹木姜子-少花万寿竹群丛 1 000~1 200 东 30~35 黄壤 24~25 12~16 Ⅳ 杉木-毛花连蕊茶-江南卷柏群丛 600~1 300 东、西 10~30 红壤、黄壤 17~25 25~52 Ⅴ 杉木-木荷-杜茎山-芒萁群丛 500~800 东北、东南 20~30 红壤 17~22 18~61 Ⅵ 青冈-木荷-细枝柃-黑足鳞毛蕨群丛 500~800 东、东南 10~30 红壤 18~26 18~28 Ⅶ 拟赤杨-虎皮楠-山矾-日本蛇根草群丛 500~800 东 25~30 红壤 19~23 19~40 Ⅷ 刨花楠-隔药柃-蕨群丛 500~900 东南、南 25~45 红壤 20~23 15~52 Ⅸ 马尾松-木荷-草珊瑚-中华薹草群丛 500~900 西 10~30 红壤 20~24 16~39 Ⅹ 毛竹-山鸡椒-地菍群丛 500~800 西南、西 5~25 红壤 21~25 23~42
下载: 导出CSV
表 4 浙江庆元巾子峰国家森林公园植被群落的典范对应分析排序
Table 4. The CCA analysis of characteristics of the plant communities in Jinzifeng Forest Park of Qingyuan, Zhejiang
排序轴 土壤湿度 土壤温度 坡度 坡向 海拔 土壤类型 特征值 物种-环境
相关性物种数据累计
贡献率/%物种-环境因子关系
数据累计贡献率/%第1排序轴 −0.171 −0.432* −0.038 −0.154 0.962** 0.906** 0.497 0.975 7.3 38.0 第2排序轴 −0.152 −0.699** 0.352* 0.003 0.059 0.044 0.224 0.885 10.6 55.2 第3排序轴 −0.068 0.226 −0.506 0.347* −0.015 0.152 0.184 0.818 13.3 69.2 第4排序轴 0.661** 0.140 −0.304* 0.240 −0.022 −0.163 0.160 0.839 15.7 81.5 说明:**P<0.01, *P<0.05。所有典范轴的显著性监测F=1.629,P=0.002
下载: 导出CSV
-
[1] JONES C C, DEL M R. Patterns of primary succession on the foreland of Coleman Glacier, Washington, USA [J]. Plant Ecol, 2005, 180(1): 105 − 11. [2] 苏日古嘎, 张金屯, 张斌, 等. 松山自然保护区森林群落的数量分类和排序[J]. 生态学报, 2010, 30(10): 2621 − 2629. Suriguga, ZHANG Jintun, ZHANG Bin, et al. Numerical classification and ordination of forest communities in the Songshan National Nature Reserve [J]. Acta Ecol Sin, 2010, 30(10): 2621 − 2629. [3] 郑超超, 伊力塔, 张超, 等. 浙江江山公益林物种种间关系及CCA排序[J]. 生态学报, 2015, 35(22): 7511 − 7521. ZHENG Chaochao, YI Lita, ZHANG Chao, et al. Interspecific relationship and canonical correspondence analysis of the dominant species in ecological service forest of Jiangshan City in Zhejiang Province [J]. Acta Ecol Sin, 2015, 35(22): 7511 − 7521. [4] 张容, 董廷发, 邓晓保, 等. 西双版纳20公顷样地热带森林植被数量分类与排序[J]. 生态学杂志, 2018, 37(2): 347 − 352. ZHANG Rong, DONG Tingfa, DENG Xiaobao, et al. Numerical classification and ordination of tropical vegetaion of a 20-hectare plot in Xishuangbanna, Southwest Yunnan [J]. Chin J Ecol, 2018, 37(2): 347 − 352. [5] 唐志红, 尉秋实, 刘虎俊, 等. 祁连山东段高寒植被群落特征及其与地形气候因子关系研究[J]. 生态学报, 2020, 40(1): 223 − 232. TANG Zhihong, YU Qiushi, LIU Hujun, et al. Characteristics of alpine vegetation community and its relationship to topographic climate factors in the eastern Qilian Mountains [J]. Acta Ecol Sin, 2020, 40(1): 223 − 232. [6] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2018. [7] 许彬, 张金屯, 杨洪晓, 等. 京西百花山植物群落数量分析[J]. 北京师范大学学报(自然科学版), 2006, 42(1): 90 − 94. XU Bin, ZHANG Jintun, YANG Hongxiao, et al. Quantitative analysis of plant communities in Baihua Mountains, Western Beijing [J]. J Beijing Norm Univ Nat Sci, 2006, 42(1): 90 − 94. [8] 齐婷婷, 王晓丽, 冯炘, 等. 庙岛群岛南五岛灌木群落结构及其对环境因子的响应[J]. 西北植物学报, 2015, 35(5): 1044 − 1051. QI Tingting, WANG Xiaoli, FENG Xin, et al. Population structure of shrub in the southern five islands of Miaodao Archipelago and its response to the environmental factors [J]. Acta Bot Boreali-Occident Sin, 2015, 35(5): 1044 − 1051. [9] 范良敏, 叶珍林, 姚丰平, 等. 巾子峰森林公园木本植物区系研究[J]. 浙江林业科技, 2012, 32(6): 27 − 32. FAN Liangmin, YE Zhenlin, YAO Fengping, et al. Study on woody plant flora of Qingyuan Jinzifeng Forest Park in Zhejiang [J]. J Zhejiang For Sci Technol, 2012, 32(6): 27 − 32. [10] 范良敏, 吴启福, 姚丰平, 等. 巾子峰森林公园生态旅游开发与保护探讨[J]. 浙江林业科技, 2002, 22(4): 96 − 98. FAN Liangmin, WU Qifu, YAO Fengping, et al. Ecological tourism development and protection of Jinzifeng Forest Park [J]. J Zhejiang For Sci Technol, 2002, 22(4): 96 − 98. [11] JAN L, SMILAUER P S. Multivariate Analysis of Ecological Data Using CANOCO[M]. Cambridge: Cambridge University Press, 2003. [12] 中国植被编辑委员会. 中国植被[M]. 北京: 科学出版社, 1980. [13] 胡理乐, 毛志宏, 朱教君, 等. 辽东山区天然次生林的数量分类[J]. 生态学报, 2005, 25(11): 2848 − 2854. HU Lile, MAO Zhihong, ZHU Jiaojun, et al. Classification and ordination of secondary forests in montane zone of eastern Liaoning Provine [J]. Acta Ecol Sin, 2005, 25(11): 2848 − 2854. [14] 何惠琴, 李绍才, 孙海龙, 等. 锦屏水电站植被数量分类与排序[J]. 生态学报, 2008, 28(8): 3706 − 3712. HE Huiqin, LI Shaocai, SUN Haihong, et al. Quantitative classification and ordination of Jinping hydropower station, Sichuan Province, China [J]. Acta Ecol Sin, 2008, 28(8): 3706 − 3712. [15] 陶楚, 陈玉凯, 杨小波, 等. 海南铜鼓岭国家级自然保护区植被数量分类与排序[J]. 中国农学通报, 2014, 30(22): 84 − 91. TAO Chu, CHEN Yukai, YANG Xiaobo, et al. Quantitative classification and ordination of vegetations in Tongguling National Nature Reserve, Hainan [J]. Chin Agric Sci Bull, 2014, 30(22): 84 − 91. [16] 王得祥, 蔺雨阳, 雷瑞德, 等. 秦岭山地天然油松林群落结构特征和数量分类研究[J]. 西北植物学报, 2009, 29(5): 1016 − 1025. WANG Dexiang, LIN Yuyang, LEI Ruide, et al. Community composition and classification of natural forest of Chinese pine (Pinus tabulaeformis Carr.) in Qinling Mountains [J]. Acta Bot Boreali-Occident Sin, 2009, 29(5): 1016 − 1025. [17] 王宇超, 周亚福, 王得祥. 秦岭南坡中段主要森林群落类型划分及环境梯度解释[J]. 生态环境学报, 2016, 25(6): 965 − 972. WANG Yuchao, ZHOU Yafu, WANG Dexiang. The quantitative classification and environmental interpretation of forest communities in the middle area of south slope of Qinling Mountains [J]. Ecol Environ Sci, 2016, 25(6): 965 − 972. [18] 朱彦鹏, 梁军, 孙志强, 等. 昆嵛山森林群落数量分类、排序及多样性垂直格局[J]. 林业科学, 2013, 49(4): 55 − 61. ZHU Yanpeng, LIANG Jun, SUN Zhiqiang, et al. Numerical classification, ordination and species diversity along elevation gradients of the forest community in Kunyu Mountain [J]. Sci Silv Sin, 2013, 49(4): 55 − 61. [19] 刘开明, 郑智, 龚大洁. 物种丰富度的垂直分布格局及其形成机制[J]. 生态学杂志, 2017, 36(2): 541 − 544. LIU Kaiming, ZHENG Zhi, GONG Dajie. Elevational patterns of species richness and their underlying mechanism [J]. Chin J Ecol, 2017, 36(2): 541 − 544. [20] SWANSON F J, KRATZ T K, CAINE N. Landform effects on ecosystem patterns and processes [J]. BioScience, 1988, 38(2): 92 − 98. [21] 叶森土, 金超, 吴初平, 等. 浙江松阳县生态公益林群落分类排序及优势种种间关联分析[J]. 浙江农林大学学报, 2020, 37(4): 693 − 701. YE Sentu, JIN Chao, WU Chuping, et al. Classification, ordination and correlation analysis of dominant species of ecological non-commercial forests in Songyang, Zhejiang [J]. J Zhejiang A&F Univ, 2020, 37(4): 693 − 701. [22] 朱国亮, 商天其, 管杰然, 等. 缙云县公益林群落数量分类与排序[J]. 浙江农林大学学报, 2017, 34(1): 68 − 77. ZHU Guoliang, SHANG Tianqi, GUAN Jieran, et al. Numerical classification and ordination of public welfare forest communities in Jinyun [J]. J Zhejiang A&F Univ, 2017, 34(1): 68 − 77. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200393
 点击查看大图
点击查看大图
计量
- 文章访问数: 2375
- HTML全文浏览量: 466
- PDF下载量: 71
- 被引次数: 0
