







-
三峡水库位于生态环境脆弱区,水环境保护一直是三峡库区重点问题。水库建立与运行改变了长江天然河道属性,天然河流转变为人工湖泊型水库,库区水文水情发生重大改变:水体流速变化显著,水体自净能力相较蓄水前也发生变化,库区支流常出现水体富营养化和水华现象[1-5]。三峡水库季节性水位涨落和水库周期性蓄水泄洪,在一定程度上导致水库水动力条件空间分布不均,进而影响水体中污染物扩散与沉积[6]。水质情况优劣对三峡库区社会经济发展与生态环境有着直接影响,水质恶化会破坏水生生态系统,导致生物多样性改变,威胁生态环境与人类健康,因此亟需进行水监测与管理[6]。分析湖泊流域水质的变化特征及其影响因素,有助于了解水质动态变化,加强流域水生态环境管理[7]。近年来,有关三峡水库水质变化研究主要集中在库区长江支流污染物浓度、泥沙淤积和水环境时空评价与库区长江干流污染物浓度时空变化等方面[8-10]。李玲等[11]以三峡水库干流水质为研究对象,结合污染浓度变化,探讨了干流水质变化与水库蓄水调度的关系。秦迪岚等[12]通过对污染物浓度进行聚类分析,研究了三峡库区的汉丰湖水质时空变化与差异。三峡水库水位随季节调度,水动力条件复杂,然而目前针对三峡库区长江干流污染物负荷变化及其与水情关系的定量研究较少。本研究分析三峡库区长江干流入出库断面水体中污染物负荷,探讨水质的时空变化特征及其对水位调度的响应,旨在揭示污染物动态变化特征与影响因素,以期为三峡水库长江干流水质监测与优化管理提供建议。
-
三峡库区是指因三峡工程淹没并有移民任务的地区(28°31′~31°50′N,105°49′~110°30′E),总面积约5.8 万 km2,包含受三峡水库建设直接影响的湖北和重庆的20余个区县[13-14]。库区内地形复杂,以山地丘陵为主,属亚热带湿润季风气候,年均气温18.0 ℃,年均降水量约1 200.0 mm,冬夏季风更替明显,降雨和气温随季风变化有明显季节特征[15]。三峡工程于1994年正式施工。三峡水库在2003年6月首次蓄水至135 m,2006年10月蓄水至156 m,2010年10月蓄水位正式达175 m。三峡水库范围从三斗坪坝址至长江上游江津。三峡水库蓄水后,河道最大长度达660 km,干流河道宽700~1 700 m,库容可达393.0 亿 m3,可拦截洪水221.5 亿 m3。三峡水库建成后,夏季汛期水位低(145 m),汛末至枯水期水位逐渐抬升,冬季枯期水位最高(175 m)。按照三峡水库周期性蓄水水位变化,将水库运行划分丰水期(145 m)、平水期(145~175 m,175~145 m)和枯水期(175 m)。
-
本研究选取三峡库区长江干流入库断面(重庆朱沱)和出库断面(湖北宜昌南津关)为对象进行研究。2013−2018年三峡库区长江干流入出库断面的水质污染物浓度数据来源于中华人民共和国生态环境保护部提供的《全国主要流域重点断面水质监测周报》(
http://www.cnemc.cn )。该周报以周为单位发布了水体污染物氨氮 (NH3-N)和高锰酸钾指数 (CODMn)数据。NH3-N是导致水生植物消失的关键因素之一,CODMn能直接反映水体有机污染程度。三峡水库入出库断面每日水位与流量数据来源于中华人民共和国长江海事局(https://cj.msa.gov.cn/ )。断面污染物负荷计算方法:Wi=10−6CiQ。其中:Wi为入库或出库断面污染物i的输移负荷(t·d−1);Ci为通过断面的水体中污染物i的质量浓度(mg·L−1);Q为通过断面的径流量(m3·d−1)。
-
数据经过检查,剔除异常值处理后,采用SPSS 15.0进行差异检验与皮尔逊相关分析,通过R语言软件绘制概率密度分布曲线与Mann-Kendall趋势检验分析。
Mann-Kendall趋势检验是非参数统计检验法。此方法不必事先设定数据分布特征,适于气象、水文数据的时间序列趋势与突变分析[16-17]。Mann-Kendall趋势检验法通过计算水文序列的趋势变化统计量UF与其反序列UB,并对比UF在显著水平下的表现(显著水平α=0.05时,
$ {U}_{\alpha \left(0.05\right)} $ =1.96,−$ {U}_{\alpha \left(0.05\right)} $ =−1.96;极显著水平α=0.001时,$ {U}_{\alpha \left(0.001\right)} $ =2.58,−$ {U}_{\alpha \left(0.001\right)} $ =−2.58)。当$ {U}_{{\rm{F}}} $ >0,水文序列呈上升趋势;当$ {U}_{{\rm{F}}} $ <0,序列呈下降趋势。$ \left|{U}_{{\rm{F}}}\right| $ <1.96,变化趋势不显著;1.96<$ \left|{U}_{{\rm{F}}}\right| $ <2.56,变化趋势显著;$ \left|{U}_{{\rm{F}}}\right| $ >2.56,变化趋势极显著。 -
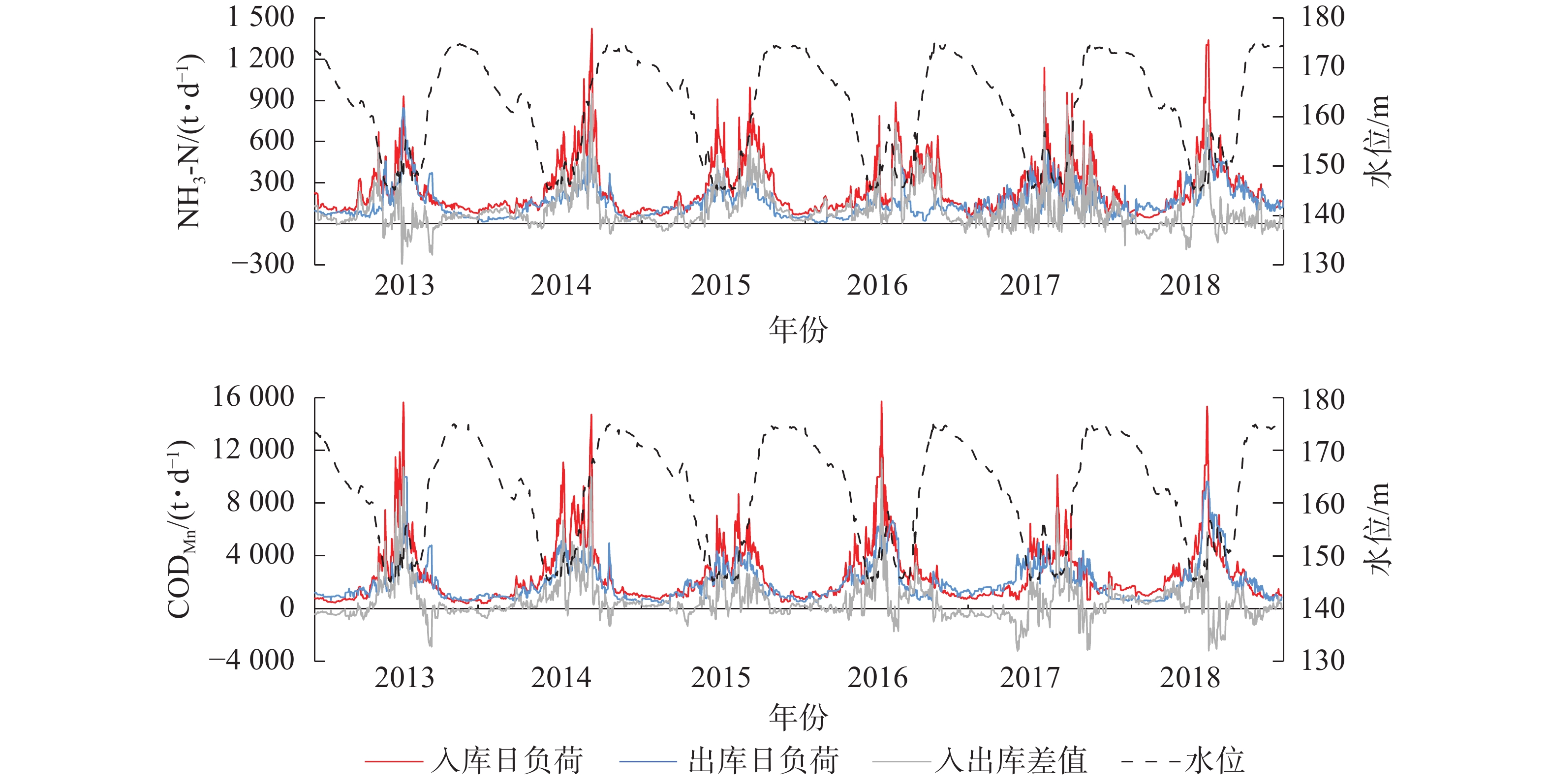
三峡库区长江干流入库与出库断面污染物负荷间的差异,反映了上游来水以及库区区间对入出库水质的影响。三峡水库蓄水后,大坝前的干流水位抬升,出现多处回水河段;干支流水体流速下降,库区上游至下游的水体流速呈下降趋势,出现明显河道型水库特征[18]。2013−2018年入出库断面NH3-N、CODMn日负荷变化见图1。可见,污染物日负荷存在周期性变化:在夏季低水位时期负荷最高,冬季高水位时期最低。对入出库断面污染物负荷做相关分析与配对样本t检验分析显示:①入库断面NH3-N日均负荷256.60 t·d−1,出库断面NH3-N日均负荷为156.45 t·d−1,两者相关系数为0.61(P<0.01),为极显著相关,且存在极显著差异(P<0.01)。②入库断面CODMn日均负荷2 530.84 t·d−1,出库断面为2 012.97 t·d−1,两者相关系数为0.76(P<0.01)),呈极显著相关,且存在极显著差异(P<0.01)。差异检验结果显示:入库断面污染物负荷显著高于出库断面。库区长江干流从天然河道转变为人工水库后,水文形态发生明显变化,如:河道变宽、流速下降等均导致水体在库区滞留时长增加,泥沙沉降作用也随之增强,污染物随泥沙沉入底泥,水体中污染物减少。此外,水库澄清效果沿程累计,伴随沿程污染物不断沉入底泥,水体污染物下降,最终使出库断面水质优于入库断面[18]。
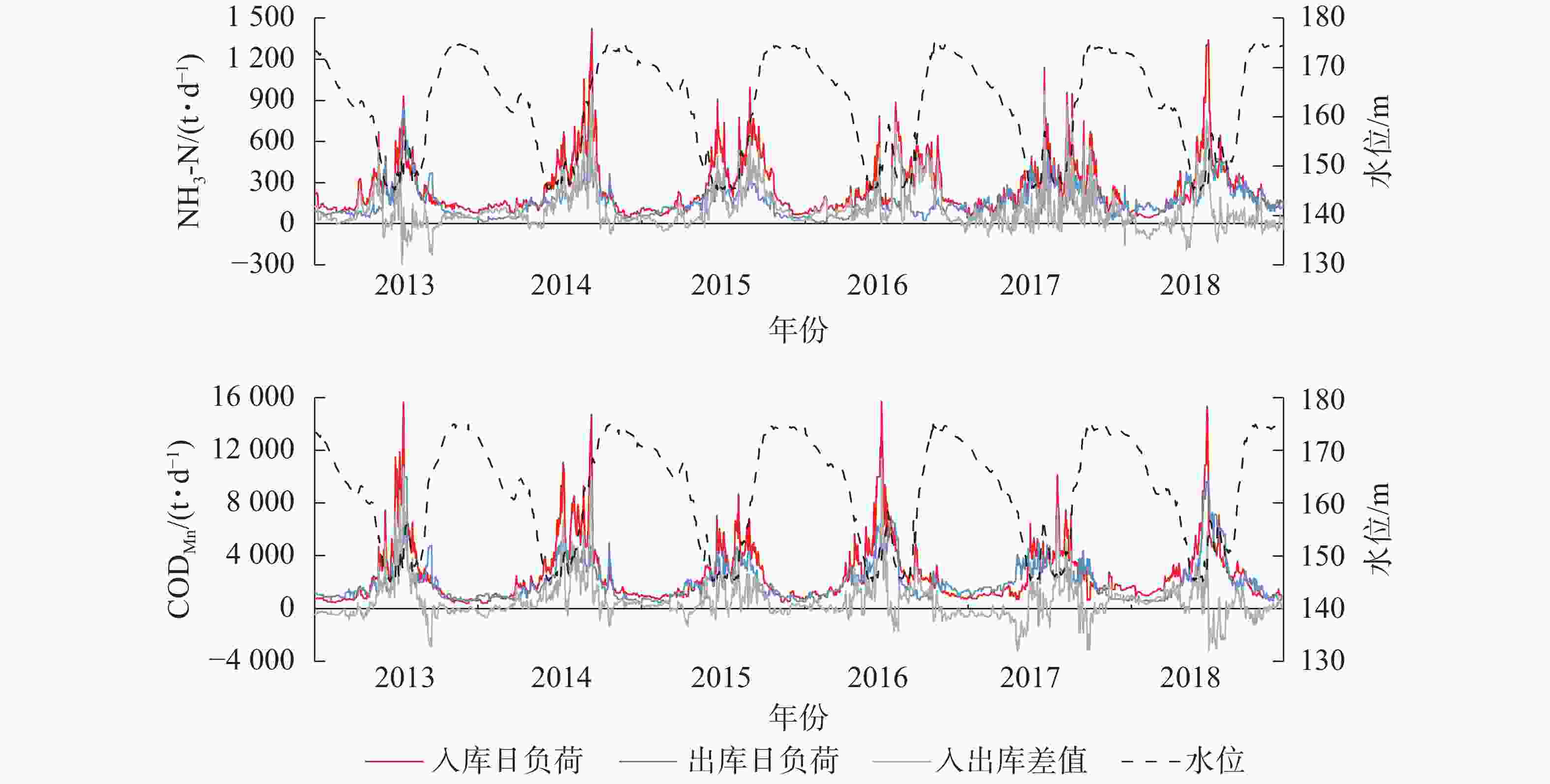
图 1 2013−2018年三峡库区入出库断面污染物日负荷与水位变化
Figure 1. Daily pollutant loads of inlet and outlet and water level changes in the Three Gorges Reservoir from 2013 to 2018
-
2013−2018年三峡水库入库断面NH3-N负荷及CODMn负荷与水位的相关系数分别为−0.44和−0.63,出库断面NH3-N负荷及CODMn负荷水位与水位的相关系数分别为−0.47和−0.58,均呈极显著负相关(P<0.01),说明三峡水库季节性水位调控对污染物负荷有较大影响。
-
因三峡水库水位周期性调度,将水库运行时间划分为:低水位运行的丰水期(6−9月)、人为蓄水调整的平水期(4−5月和10−11月),高水位运行的枯水期(12月至次年3月),不同水位运行期污染物日均负荷统计见表1。相关性分析结果表明:在丰水期和平水期,出库断面污染物日负荷与入库断面污染物日负荷极显著相关(P<0.01,表2);差异分析结果表明:在丰水期、平水期和枯水期,出库断面污染物日负荷与入库断面污染物日负荷差异极显著(P<0.01,表2)。
表 1 不同水位时期入出库断面污染物日均负荷
Table 1. Statistics of average daily pollutant loads of inlet and outlet in the Three Gorges Reservoir at different water level periods
水期 NH3-N/(t·d−1) CODMn/(t·d−1) 入库断面 出库断面 入库断面 出库断面 丰水期 393.75 241.22 4 261.76 3 111.89 平水期 231.20 131.09 1 651.10 1 548.12 枯水期 114.01 78.77 1 060.09 917.83 表 2 不同水位时期入出库断面污染物日负荷相关性分析与差异分析
Table 2. Correlation and differential analysis of daily pollutant loads of inlet and outlet in the Three Gorges Reservoir at different water level periods
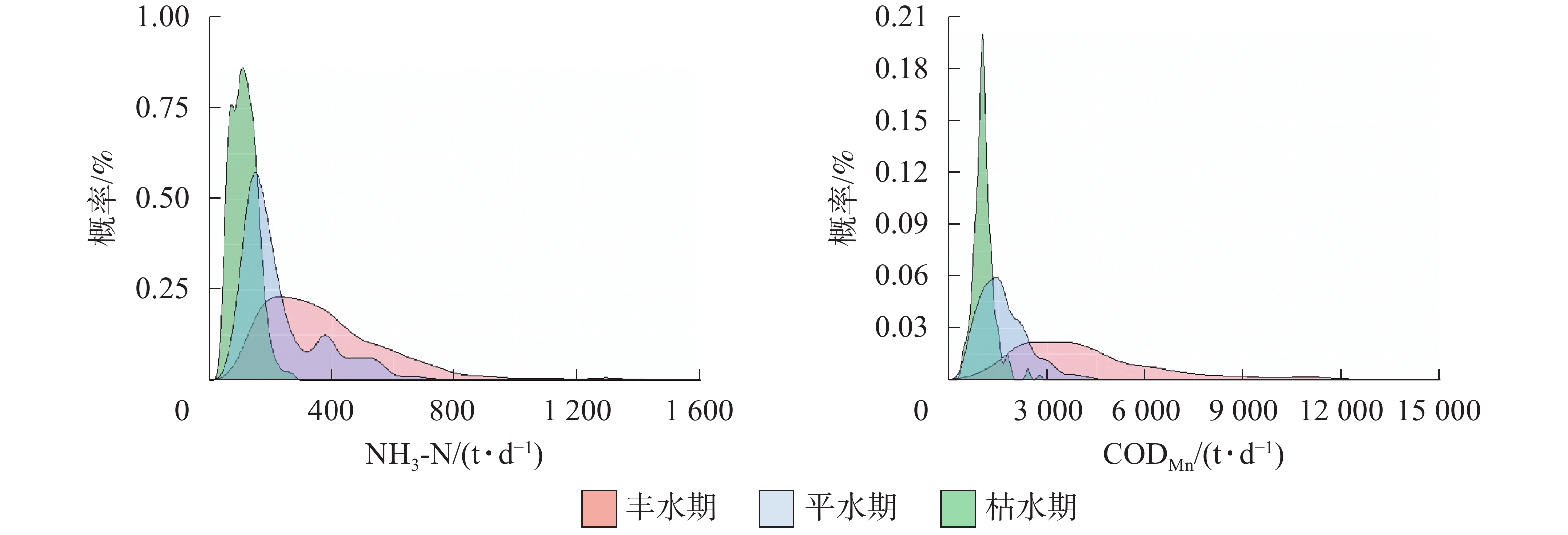
水期 相关系数 t NH3-N CODMn NH3-N CODMn 丰水期 0.41** 0.54** 21.19** 16.22** 平水期 0.29** 0.37** 17.88** 2.95* 枯水期 0.06 0.09 12.52** 6.71** 说明:相关性分析中,**表示极显著相关(P<0.01)。差异 分析t检验中,*表示差异显著(P<0.05);**表示差 异极显著(P<0.01) 如图2所示:①在丰水期,入库断面NH3-N日负荷大于250 t·d−1的概率累计有70.40%,而出库断面仅有35.80%;入库断面CODMn日负荷大于3 000 t·d−1的概率有64.85%,而出库断面仅有38.74%。在丰水期,入库断面NH3-N与CODMn日负荷明显多于出库断面。②在平水期,入库断面NH3-N日负荷大于180 t·d−1的概率有51.37%,而出库断面仅有23.40%;入库断面CODMn日负荷大于1 500 t·d−1的概率有52.65%,出库断面有45.16%。在平水期,入库断面NH3-N日负荷明显多于出库断面。③在枯水期,入库断面NH3-N日负荷大于110 t·d−1的概率有52.49%,而出库断面仅有17.22%;入库断面CODMn日负荷大于1 000 t·d−1的概率有30.29%,出库断面有25.31%。在枯水期,入库断面NH3-N日负荷明显多于出库断面。以上结果表明:在不同水位时期,入库断面污染日负荷均高于出库断面,其中NH3-N日负荷相差很大。

图 2 不同水位时期入出库断面污染物日负荷概率密度曲线图
Figure 2. Probability density line of daily pollutant loads of inlet and outlet in the Three Gorges Reservoir at different water level periods
-
在时间尺度,对入库或出库断面在不同水位运行期的污染物日负荷进行差异分析检验(表3和表4)表明:①在不同水位条件下,入库断面各项污染物负荷均差异显著(P<0.05);②在不同水位条件下,出库断面各项污染物负荷均差异显著(P<0.05);③入出库断面污染物负荷在丰水期最高,枯水期最低的特征,说明水位季节性剧烈波动对三峡库区长江干流入出库断面污染物负荷有较大的影响。
表 3 不同水位时期入库断面污染物日负荷单因素方差分析
Table 3. Differential analysis of daily pollutant loads of inlet in the Three Gorges Reservoir at different water level periods
水期 入库断面NH3-N 水期 入库断面CODMn 丰水期 平水期 枯水期 丰水期 平水期 枯水期 丰水期 162.55* 279.74* 丰水期 2 549.65* 3 140.67* 平水期 −162.55* 117.19* 平水期 −2 549.65* 591.02* 枯水期 −279.74* −117.19* 枯水期 −3 140.67* −591.02* 说明:*表示差异显著(P<0.05) 表 4 不同水位时期出库断面污染物日负荷单因素方差分析
Table 4. Differential analysis of daily pollutant loads of outlet in the Three Gorges Reservoir at different water level periods
水期 出库断面NH3-N 水期 出库断面CODMn 丰水期 平水期 枯水期 丰水期 平水期 枯水期 丰水期 110.13* 162.45* 丰水期 1 563.77* 2 194.07* 平水期 −110.13* 52.32* 平水期 −1 563.77* 630.30* 枯水期 −162.45* −52.32* 枯水期 −2 194.07* −630.30* 说明:*表示差异显著(P<0.05) 分别对入出库污染物负荷在不同水位条件下的状态进行概率密度曲线分析(图3和图4)表明:①入库断面NH3-N日负荷在丰水期时有70.37%的概率大于250 t·d−1,在平水期时有28.75%,在枯水期时很少;入库断面CODMn日负荷在丰水期时有47.39%的概率大于3 800 t·d−1,在平水期和枯水期均不超过3 800 t·d−1。②出库断面NH3-N日负荷在丰水期时有69.61%的概率大于160 t·d−1,在平水期时有42.05%,在枯水期时有不到15.00%;CODMn日负荷在丰水期时有66.23%的概率大于3 800 t·d−1,在平水期时有14.47%,在枯水期时负荷不达3 800 t·d-1。以上结果表明:丰水期时,入库断面NH3-N日负荷与CODMn日负荷均显著高于平水期和枯水期;在丰水期时,出库断面NH3-N日负荷与CODMn日负荷量偏高,而在枯水期时最低。
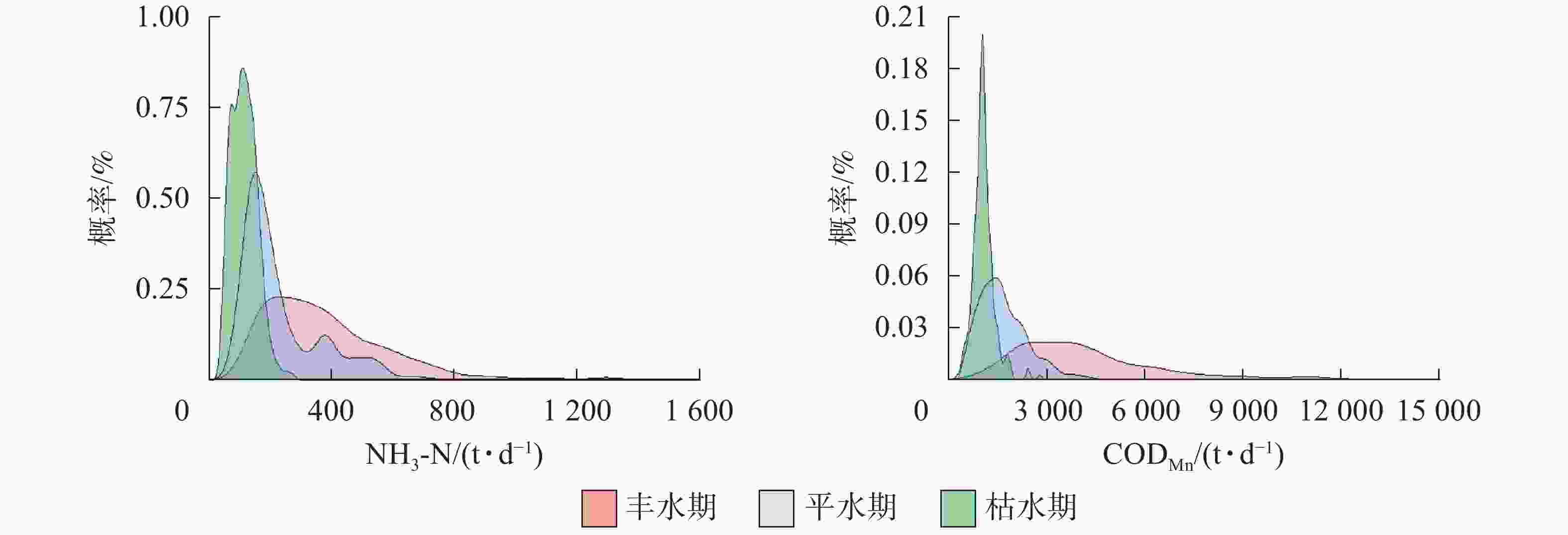
图 3 不同水位时期入库断面污染物日负荷概率密度曲线图
Figure 3. Probability density line of daily pollutant loads of inlet in the Three Gorges Reservoir at different water level periods
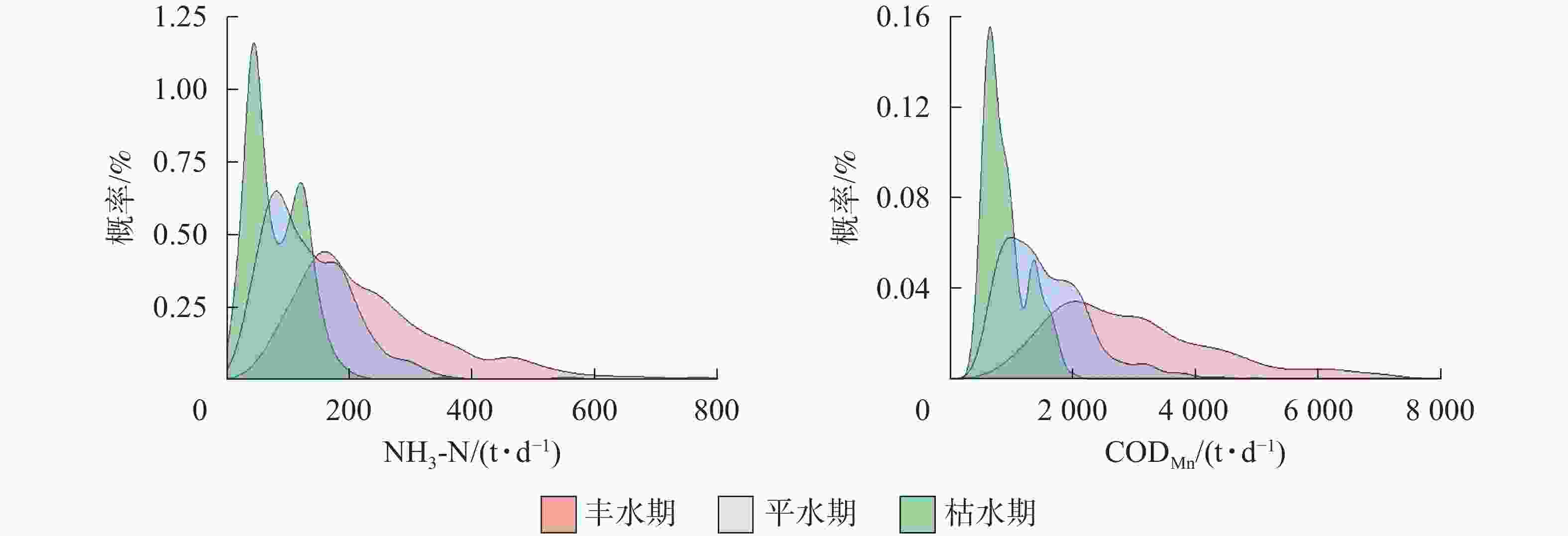
图 4 不同水位时期出库断面污染物日负荷概率密度曲线图
Figure 4. Probability density line of daily pollutant loads of outlet in the Three Gorges Reservoir at different water level periods
-
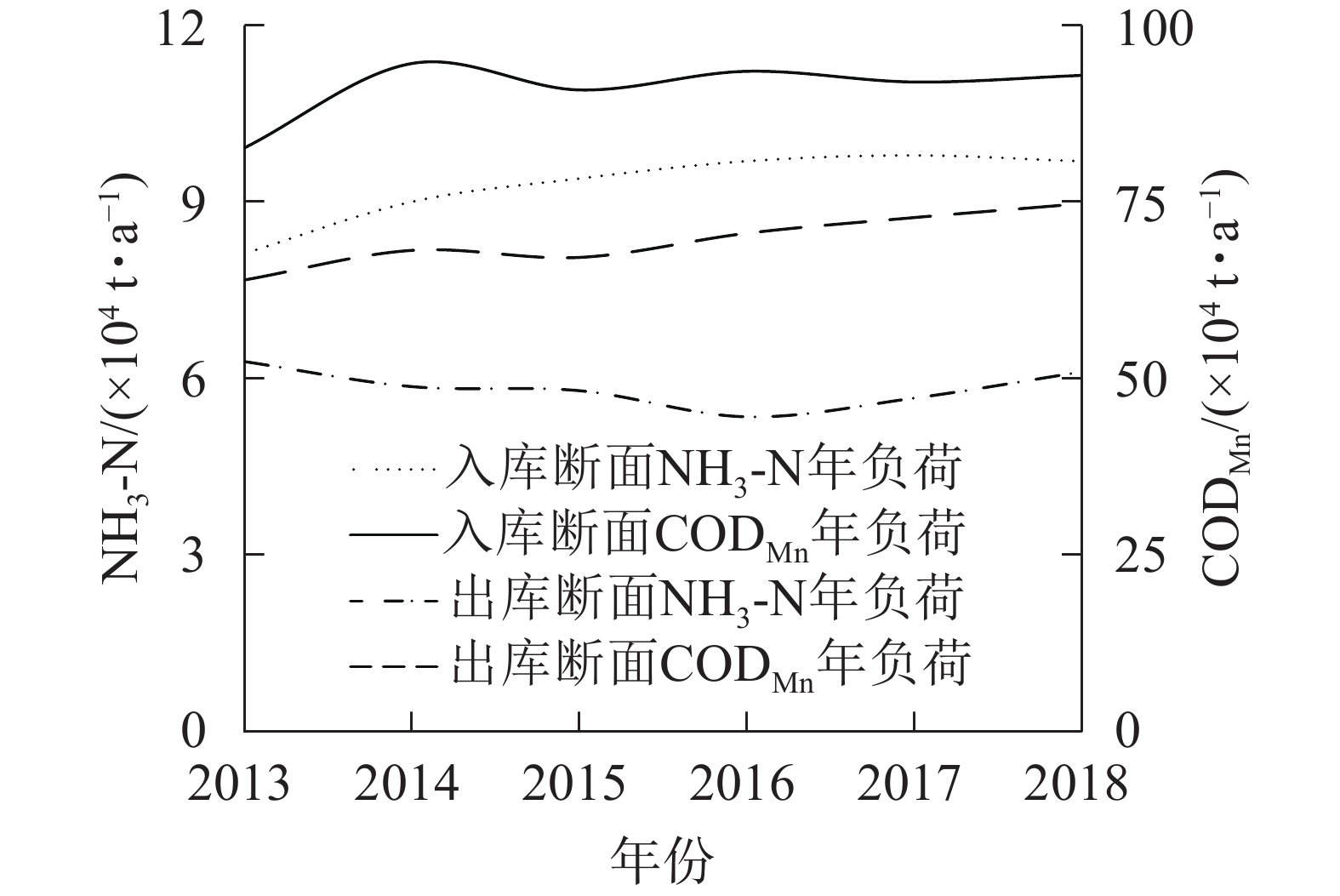
入出库断面2013−2018年污染物年负荷如图5所示,利用R语言对污染物负荷年变化进行Mann-Kendall趋势检验(图6)。结果表明:入库断面NH3-N年负荷与出库断面CODMn年负荷UF曲线与临界值线(Uα=1.96,α=0.05)相交,表明两者呈显著上升趋势(P<0.05),其他断面污染物年负荷无显著变化。
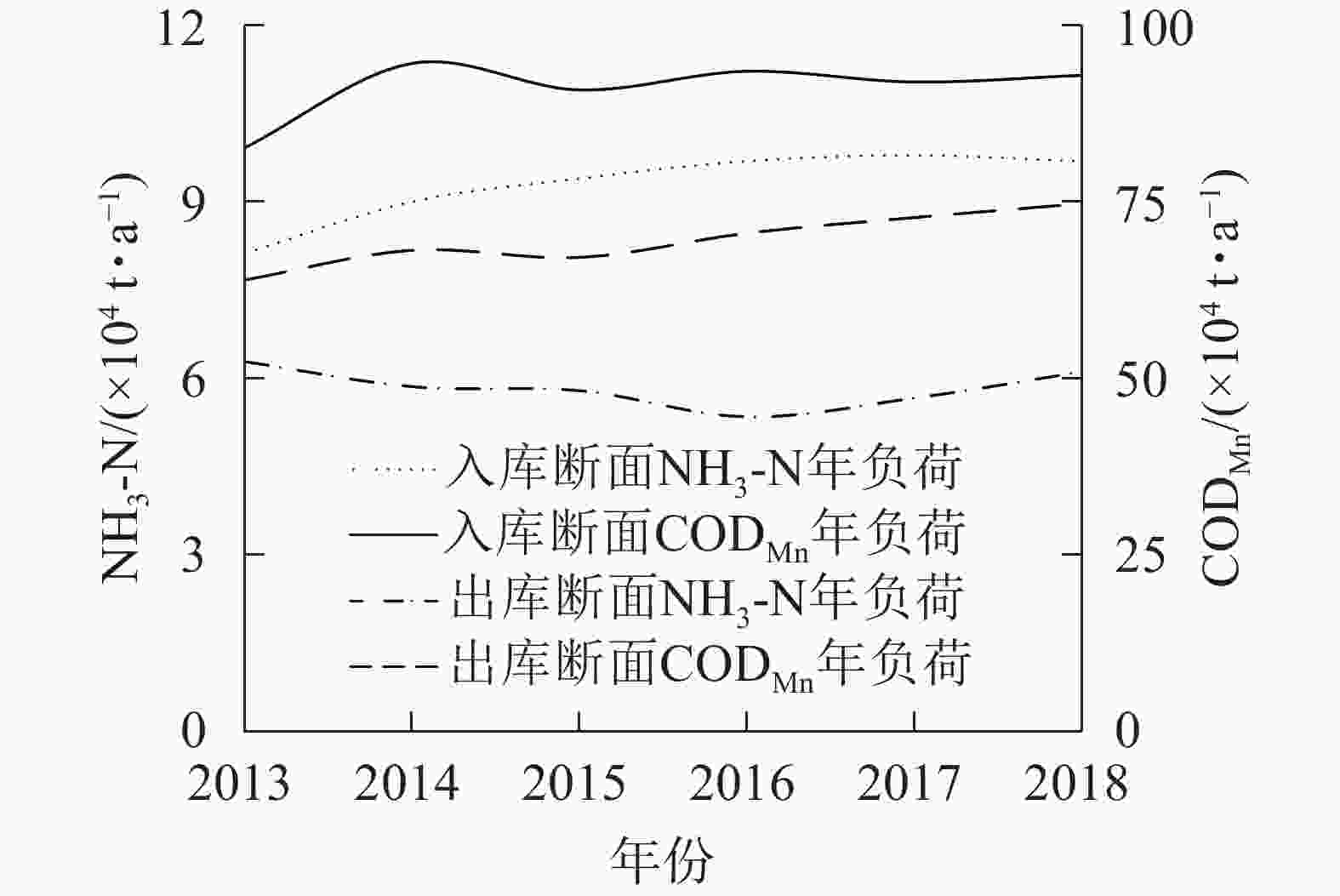
图 5 2013−2018年三峡库区长江干流入出库断面污染物年负荷
Figure 5. Annual pollutant loads of inlet and outlet in the Three Gorges Reservoir from 2013 to 2018
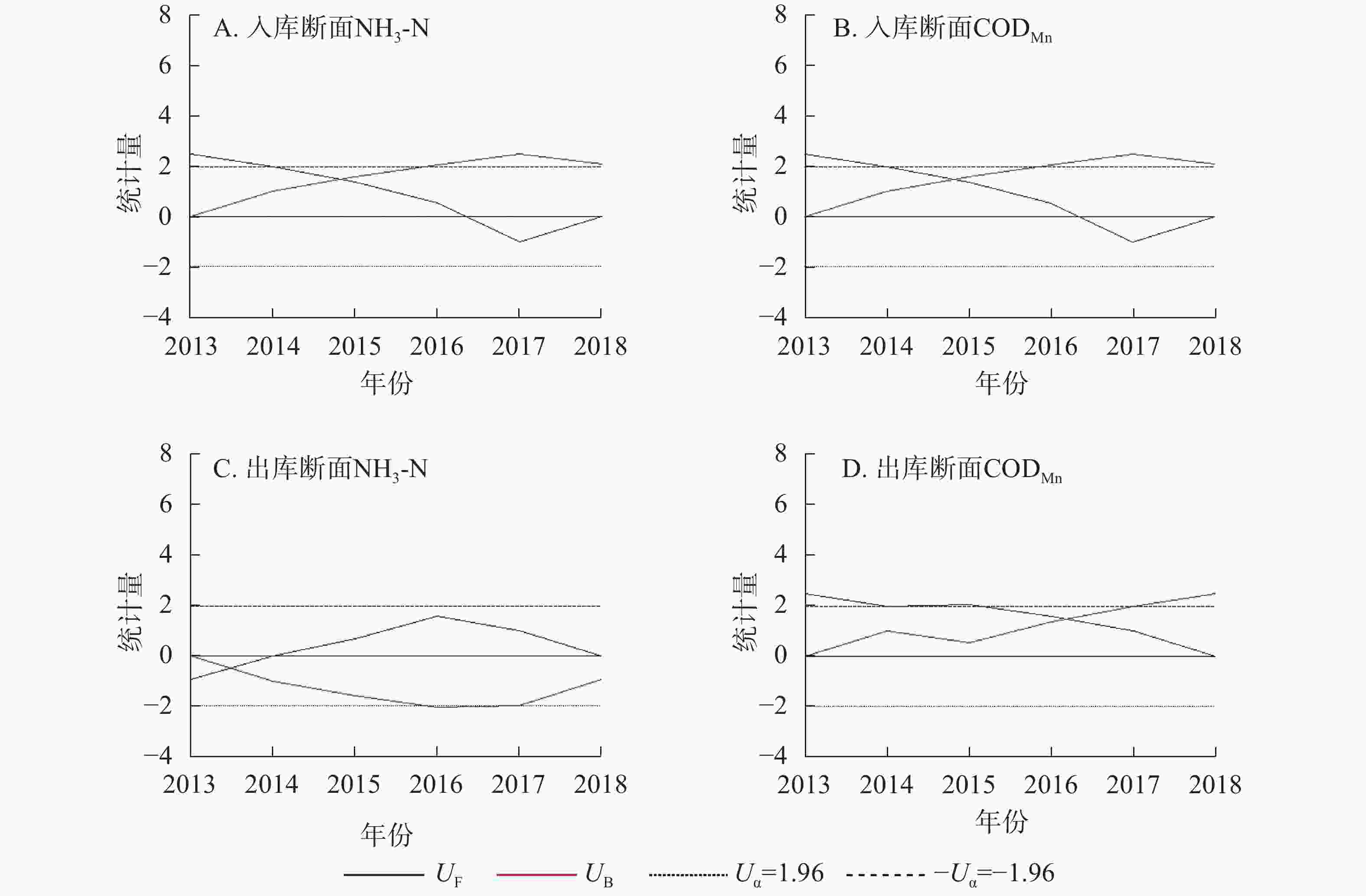
图 6 2013−2018年三峡库区长江干流入出库断面污染物年负荷Mann-Kendall趋势检验曲线
Figure 6. Trend test curve of annual pollutant loads of inlet and outlet in the Three Gorges Reservoir from 2013 to 2018, by Mann-Kendall
-
水质受水位波动变化影响的研究很多,WHITE等[19]利用模型演示出湖泊持续低水位状态会导致水体中悬浮物增加,造成水质恶化。三峡水库运行后,库区水位受人工调度,汛期因泄洪而低水位运行,入库沙量显著增加,泄洪也使得大量泥沙排至下游;冬季高水位运行,水位抬升至175 m,水库大量蓄水。非汛期泄水改变了原有的河道条件,导致三峡库区与天然河道条件差异大[20]。
三峡水库不同水位运行期因蓄水量不同而导致的稀释作用变化,是引发污染物负荷波动的重要因素。2010年三峡工程进入常态化水位调控运行阶段后,水库水质受到季节性因素影响,水温、来水量、泥沙量、水文条件与人类生产生活等因素都会影响水质[21]。在时间尺度上,入库与出库断面污染物负荷变化与三峡水库水位波动高度相关,在夏季汛期的低水位阶段污染物负荷最高,而在冬季枯水期的高水位阶段污染物负荷较低的概率最大。这是因为在丰水期,降雨丰沛,三峡水库来水流量增大,携带污染物进入水体的能力强;汛期降雨冲击力强,在降雨径流的淋溶与冲刷下,污染物随地表径流和泥沙进入水体,引发水质恶化[22]。但受到汛期水库排洪影响,水位下降,此期间水库储水量大幅减少,污染物稀释与降解能力下降,在一定程度上加重了水质恶化[23]。有研究表明[24]:水体中CODMn含量与泥沙显著相关,泥沙与有机质含量决定了CODMn含量,夏季入江水体携带大量泥沙进入水库,是导致CODMn高于其他时期的主要因素。汛期来水来沙量大,携带大量含氮污染物进入水体;进入蓄水期后,水体含沙量降低,清水下泄,进入枯期高水位后水体含氮量会下降[25]。
在枯水期高水位运行时,水库含水量增加,水体流动性下降,悬浮物沉降作用增强,CODMn、总磷、重金属等与悬浮物密切相关的污染物沉降入底泥,并且沉降作用沿程不断累积,接近大坝的出库断面受水位调度影响显著,水体流速变化明显,污染物沉降作用也更加显著[1]。此外,高水位运行期水库蓄水量大,有助于污染物稀释与沉降[18]。同时由于枯期降雨减少,陆地污染物迁移力下降,入库污染物随之减少[19]。
-
三峡水库水质与水文情况、水体污染情况、污染物输入方式,以及三峡库区点源、面源污染输入变化密切相关。有研究认为:三峡库区干流污染物来源主要是农业面源污染、工业生产和生活污水的排放与输入,库区城镇化建设与农业生产活动必然引入新的污染[26]。三峡库区农业污染来源有肥料施用、农作物秸秆燃烧、畜禽养殖等。化肥中的氮素会通过淋溶、吸附、反硝化等途径融入水土,不合理的使用方式也会造成农药化肥大量流失,进入水体造成污染[27]。三峡库区极具区域特色,库区内生态环境受农业面源污染影响严重,尤其是水库蓄水后,天然水文条件发生剧变,会降低水体中污染物的扩散、稀释与转化过程[28]。三峡水库蓄水前后,NH3-N和CODMn单位负荷造成的岸边污染面积比为1∶9和1∶14,伴随水库蓄水,在水流顶托作用下,污染带有明显回溯趋势[29]。
污染物负荷在空间尺度存在的差异,除三峡水库水体发挥蓄清的作用,域外污染物输入也是导致水体污染的重要原因。具体污染物输入来源包括城镇生活污水、工业废水、农业活动、油污废水、垃圾排放等,伴随城镇化加速和农业生产规模扩大,均会带来新的污染输入[18]。位于库首的重庆市大力推行三峡库区畜牧发展工作,畜牧养殖增加也会造成环境污染。同时因三峡大坝建立,人口迁移与设施配套建设而带来的耕地占用与生态环境破坏问题,也引发了水土流失[30]。三峡库区上游发展水平较高,且农村人口占比大,农产畜牧活动多,农药化肥使用量大,外源性污染物输入提高了水体CODMn含量;未充分自净的河水经过上游,在接收当地污染物输入后,向下游流去[17]。上游来水污染、支流污水汇入与沿程的面、点源污染均会引入污染物进入水体。有研究表明:库区污染物指标最低值均出现在库位区域,区域污染物排放冷点区域也位于三峡库位[30]。下游社会经济发展水平较低,污染排放小且距离上游较远,水体在到达下游前得到充分自净,经水库澄清作用后,污染物含量下降[18]。
-
随着经济发展、人口密度增加和城市化程度加快,生活污染排放增大,对水环境也会产生一定影响[25]。三峡水库在2010年进入常规化水位调控后,一方面,水库周期性蓄水后,库区水文形式显著改变,库区水环境变得脆弱,支流常出现水体富营养化与水华现象;另一方面,污染物、污废水处理效率不足,上游又是污染物主要来源,库区污染防治力度有待提高[31]。Mann-Kendall趋势检验结果显示:仍有污染物负荷呈上升趋势,说明库区内污染排放与污染治理仍存在疏忽漏洞。三峡水库蓄水后,氮磷污染是库区水质主要的污染因子,其中大部分来源于上游,库区富营养化问题日益严重[32]。针对三峡库区存在的污染问题,应减少库区人口压力,以达到减轻库区生态环境压力的目的;继续加强开展三峡库区防护林建设工作,积极推进高效农业,鼓励有机肥料使用;积极开展清洁小流域建设,建立生态修复区、生态治理区与生态保护区[33]。要加强库区生态环境管理与审查机制,协调库区生态环境保护与社会发展,实现可持续发展的最终目标。
-
研究结果显示:①研究期间,长江干流入库断面NH3-N、CODMn污染物负荷显著高于出库断面,出库断面污染物负荷与入库断面高度相关。入库断面污染物与支流农业面源污染是水库污染物主要来源。伴随水库澄清作用,污染物沿程下降。②入出库断面污染物负荷受三峡水库水位影响显著,在丰水期水库低水位运行时,污染物负荷最大;在枯水期水库高水位运行时,污染物负荷最小。可见,季节性水库调度对库区水质有着不可忽视的作用。丰水期水库蓄水量少,水体澄清作用下降,但汛期降雨扰乱水环境稳定性,导致污染物含量增加;枯水期水库蓄水量最大,水环境稳定,有助于污染物稀释。③Mann-Kendall趋势检验结果显示:入库断面NH3-N年负荷和出库断面CODMn年负荷呈显著上升趋势,这与社会经济发展、当地生产生活因素相关。污染物治理需加强水质环保工作,提高污水、废水处理效率,促进水环境与社会经济共同发展。
NH3-N and CODMn load in the inlet and outlet sections of the Yangtze River in Three Gorge Reservoir Area
-
摘要:
目的 探究三峡库区长江干流入出库断面污染物负荷变化及其与三峡水库水位的关系。 方法 基于三峡库区长江干流入库断面(重庆朱沱)与出库断面(湖北宜昌南津关)2013−2018年氨氮(NH3-N)和高锰酸钾指数(CODMn)与断面水体流量数据,采用密度分布曲线、Mann-Kendall趋势分析法对污染物负荷情况进行分析。 结果 ①入库断面NH3-N、CODMn负荷极显著高于出库断面,入库断面污染物负荷与出库断面污染物负荷极显著相关(P<0.01)。三峡库区污染主要来自入库断面与沿程支流输入,伴随水库澄清作用,污染物沿程下降。②入出库断面污染物负荷与三峡水库水位波动极显著负相关(P<0.01),低水位运行时污染物负荷最大,高水位运行时污染物负荷最小。③Mann-Kendall趋势分析结果显示:入库断面NH3-N年负荷与出库断面CODMn年负荷呈显著上升趋势(P<0.05)。 结论 三峡低水位运行时水库蓄水少,水体澄清作用下降;高水位运行时水库蓄水量大,水环境稳定,有助于污染物稀释。三峡库区和上游水污染治理不容忽视。图6表4参33 Abstract:Objective This study aims to analyze the variation of pollution load in the inlet and outlet sections of the Yangtze River in Three Gorges Reservoir (TGR) area and its relationship with the water level of TGR. Method Based on the data of ammonia nitrogen (NH3-N) and potassium permanganate index (CODMn) and water flow in TGR area from 2013 to 2018 at the inlet section (Zhutuo, Chongqing) and the outlet section (Nanjinguan, Yichang, Hubei) of the main stream of the Yangtze River, the pollution load and its relationship with the water level of TGR were analyzed through density distribution curve and Mann-kendall trend method. Result (1)The pollution load of NH3-N and CODMn in the inlet section was significantly higher than that of the outlet section, and the pollution load of the inlet and outlet sections was highly correlated (P<0.01). The pollution in TGR area mainly came from the inlet section and tributaries along the way. With the clarification of the reservoir, pollutants decreased along the way. (2) The pollution load at the inlet and outlet sections was negatively correlated with the fluctuation of water level of TGR(P<0.01). (3) Mann-Kendall trend analysis showed that the annual load of NH3-N at the inlet section and that of CODMn at the outlet section increased significantly (P<0.01). Conclusion The pollution load is the largest when TGR is at low water level, and is the smallest when at high water level. When the water level is low, the reservoir has less water storage and the water clarifying effect decreases. When the water level is high, the reservoir has a large water storage capacity and a stable water environment, which helps to dilute pollutants. Water pollution control in TGR area and upstream should not be ignored. [Ch, 6 fig. 4 tab. 33 ref.] -
Key words:
- Three Gorges Reservoir /
- pollution load /
- NH3-N /
- CODMn /
- variation trend
-
图 1 2013−2018年三峡库区入出库断面污染物日负荷与水位变化
Figure 1 Daily pollutant loads of inlet and outlet and water level changes in the Three Gorges Reservoir from 2013 to 2018
图 2 不同水位时期入出库断面污染物日负荷概率密度曲线图
Figure 2 Probability density line of daily pollutant loads of inlet and outlet in the Three Gorges Reservoir at different water level periods
图 3 不同水位时期入库断面污染物日负荷概率密度曲线图
Figure 3 Probability density line of daily pollutant loads of inlet in the Three Gorges Reservoir at different water level periods
图 4 不同水位时期出库断面污染物日负荷概率密度曲线图
Figure 4 Probability density line of daily pollutant loads of outlet in the Three Gorges Reservoir at different water level periods
图 5 2013−2018年三峡库区长江干流入出库断面污染物年负荷
Figure 5 Annual pollutant loads of inlet and outlet in the Three Gorges Reservoir from 2013 to 2018
图 6 2013−2018年三峡库区长江干流入出库断面污染物年负荷Mann-Kendall趋势检验曲线
Figure 6 Trend test curve of annual pollutant loads of inlet and outlet in the Three Gorges Reservoir from 2013 to 2018, by Mann-Kendall
表 1 不同水位时期入出库断面污染物日均负荷
Table 1. Statistics of average daily pollutant loads of inlet and outlet in the Three Gorges Reservoir at different water level periods
水期 NH3-N/(t·d−1) CODMn/(t·d−1) 入库断面 出库断面 入库断面 出库断面 丰水期 393.75 241.22 4 261.76 3 111.89 平水期 231.20 131.09 1 651.10 1 548.12 枯水期 114.01 78.77 1 060.09 917.83  下载: 导出CSV
下载: 导出CSV
表 2 不同水位时期入出库断面污染物日负荷相关性分析与差异分析
Table 2. Correlation and differential analysis of daily pollutant loads of inlet and outlet in the Three Gorges Reservoir at different water level periods
水期 相关系数 t NH3-N CODMn NH3-N CODMn 丰水期 0.41** 0.54** 21.19** 16.22** 平水期 0.29** 0.37** 17.88** 2.95* 枯水期 0.06 0.09 12.52** 6.71** 说明:相关性分析中,**表示极显著相关(P<0.01)。差异 分析t检验中,*表示差异显著(P<0.05);**表示差 异极显著(P<0.01)
下载: 导出CSV
表 3 不同水位时期入库断面污染物日负荷单因素方差分析
Table 3. Differential analysis of daily pollutant loads of inlet in the Three Gorges Reservoir at different water level periods
水期 入库断面NH3-N 水期 入库断面CODMn 丰水期 平水期 枯水期 丰水期 平水期 枯水期 丰水期 162.55* 279.74* 丰水期 2 549.65* 3 140.67* 平水期 −162.55* 117.19* 平水期 −2 549.65* 591.02* 枯水期 −279.74* −117.19* 枯水期 −3 140.67* −591.02* 说明:*表示差异显著(P<0.05)
下载: 导出CSV
表 4 不同水位时期出库断面污染物日负荷单因素方差分析
Table 4. Differential analysis of daily pollutant loads of outlet in the Three Gorges Reservoir at different water level periods
水期 出库断面NH3-N 水期 出库断面CODMn 丰水期 平水期 枯水期 丰水期 平水期 枯水期 丰水期 110.13* 162.45* 丰水期 1 563.77* 2 194.07* 平水期 −110.13* 52.32* 平水期 −1 563.77* 630.30* 枯水期 −162.45* −52.32* 枯水期 −2 194.07* −630.30* 说明:*表示差异显著(P<0.05)
下载: 导出CSV
-
[1] 江婷, 朱慧君, 郭海川, 等. 水质监测质量管理问题及应对措施分析[J]. 中国资源综合利用, 2019, 37(3): 130 − 132. JIANG Ting, ZHU Huijun, GUO Haichuan, et al. Analysis on quality management of water quality monitoring and countermeasures [J]. China Resour Compr Util, 2019, 37(3): 130 − 132. [2] 苏青青, 宋林旭, 刘德富, 等. 三峡水库香溪河沉积物氮含量和氨氮释放特征[J]. 水生态学杂志, 2019, 40(3): 1 − 7. SU Qingqing, SONG Linxu, LIU Defu, et al. Total nitrogen content and ammonia nitrogen release from surface sediments of Xiangxi River, Three Gorges Reservoir Region [J]. J Hydroecol, 2019, 40(3): 1 − 7. [3] 尹真真, 李琎. 三峡水库蓄水前后长江干流主要污染物浓度变化趋势分析研究[J]. 环境科学与管理, 2014, 39(3): 42 − 45. YIN Zhenzhen, LI Jin. Variation tendencies of major pollutants concentration in mainstream of Changjiang River before and after in three Gorges Dam [J]. Environ Sci Manage, 2014, 39(3): 42 − 45. [4] 尹海龙, 徐祖信. 河流综合水质评价方法比较研究[J]. 长江流域资源与环境, 2008, 17(5): 729 − 733. YIN Hailong, XU Zuxin. Comparative study on typical river comprehensive water quality assessment methods [J]. Resour Environ Yangtze Basin, 2008, 17(5): 729 − 733. [5] 蒋增辉, 曾次元, 韩敏奇, 等. 黄浦江上游水源地水质污染分析与突发性水污染事件分级[J]. 净水技术, 2013, 32(4): 15 − 20. JIANG Zenghui, ZENG Ciyuan, HAN Minqi, et al. Analysis of water quality pollution and classification of accidental water pollution events in upstream water surce of Huangpu River [J]. Water Purif Technol, 2013, 32(4): 15 − 20. [6] 李忠武, 赵新娜, 谢更新, 等. 三峡工程蓄水对洞庭湖水环境质量特征的影响[J]. 地理研究, 2013, 32(11): 2021 − 2030. LI Zhongwu, ZHAO Xinna, XIE Gengxin, et al. Water environmental quality properties of Dongting Lake affected by construction of the Three Gorges Project [J]. Geogr Res, 2013, 32(11): 2021 − 2030. [7] 李冰, 杨桂山, 万荣荣, 等. 鄱阳湖出流水质2004−2014年变化及其对水位变化的响应: 对水质监测频率的启示[J]. 长江流域资源与环境, 2017, 26(2): 289 − 296. LI Bing, YANG Guishan, WAN Rongrong, et al. Temporal variability of water quality in Poyang Lake outlet and the associated water level fluctuations: a water quality sampling revelation [J]. Resour Environ Yangtze Basin, 2017, 26(2): 289 − 296. [8] 卓海华, 吴云丽, 刘旻璇, 等. 三峡水库水质变化趋势研究[J]. 长江流域资源与环境, 2017, 26(6): 925 − 936. ZHUO Haihua, WU Yunli, LIU Minxuan, et al. Trend study of water quality in the Three Gorges Reservoir [J]. Resour Environ Yangtze Basin, 2017, 26(6): 925 − 936. [9] 陈永灿, 付健, 刘昭伟, 等. 三峡大坝下游溶解氧变化特性及影响因素分析[J]. 水科学进展, 2009, 20(4): 526 − 530. CHEN Yongcan, FU Jian, LIU Zhaowei, et al. Analysis of the variety and impact factors of dissolved oxygen downstream of Three Gorges Dam after the impoundment [J]. Adv Water Sci, 2009, 20(4): 526 − 530. [10] 崔彦萍, 王保栋, 陈求稳. 三峡正常蓄水后长江口叶绿素a和溶解氧变化及其成因[J]. 生态学报, 2014, 34(21): 6309 − 6316. CUI Yanping, WANG Baodong, CHEN Qiuwen. The spatial-temporal dynamics of chlorophyll a and DO in the Yangtze estuary after normal impoundment of the Three Gorges Reservoir [J]. Acta Ecol Sin, 2014, 34(21): 6309 − 6316. [11] 李玲, 覃春丽. 三峡梯级电站水体溶解氧的累积影响分析[J]. 水力发电学报, 2012, 31(1): 189 − 194. LI Ling, QIN Chunli. Cumulative impact of Three Gorges cascade hydropower stations on dissolved oxygen [J]. J Hydroelectric Eng, 2012, 31(1): 189 − 194. [12] 秦迪岚, 罗岳平, 黄哲, 等. 洞庭湖水环境污染状况与来源分析[J]. 环境科学与技术, 2012, 35(8): 193 − 198. QIN Dilan, LUO Yueping, HUANG Zhe, et al. Pollution status and source analysis of water environment in Dongting Lake [J]. Environ Sci Technol, 2012, 35(8): 193 − 198. [13] 黄祺, 何丙辉, 赵秀兰, 等. 三峡库区汉丰湖水质的时空变化特征分析[J]. 西南大学学报(自然科学版), 2016, 38(3): 136 − 142. HUANG Qi, HE Binghui, ZHAO Xiulan, et al. Analysis on spatiotemporal variation characteristics of water quality of Hanfeng Lake in Three Gorges Reservoir Region of China [J]. J Southwest Univ Nat Sci Ed, 2016, 38(3): 136 − 142. [14] 国洪磊, 周启刚. 三峡库区蓄水前后土地利用变化对生态系统服务价值的影响[J]. 水土保持研究, 2016, 23(5): 222 − 228. GUO Honglei, ZHOU Qigang. Effect of land use change on ecosystem service value pre and post the water storage in the Three Gorges Reservoir Area [J]. Res Soil Water Conserv, 2016, 23(5): 222 − 228. [15] 黄春波, 滕明君, 曾立雄, 等. 长江三峡库区土地利用/覆盖的长期变化[J]. 应用生态学报, 2018, 29(5): 1585 − 1596. HUANG Chunbo, TENG Mingjun, ZENG Lixiong, et al. Long-term changes of land use/cover in the Three Gorges Reservoir Area of the Yangtze River, China [J]. Chin J Appl Ecol, 2018, 29(5): 1585 − 1596. [16] NEETI N, EASTMAN J R. Contextual Mann-kendall approach for the assessment of trend significance in image time series [J]. Trans GIS, 2011, 15(5): 599 − 611. [17] ZHANG Yonggang, ZHU Suyun, TAN Junkun, et al. The influence of water level fluctuation on the stability of landslide in the Three Gorges Reservoir [J]. Arabian J Geosci, 2020, 13(17): 1 − 10. [18] 王小焕, 邵景安, 王金亮, 等. 三峡库区长江干流入出库水质评价及其变化趋势[J]. 环境科学学报, 2017, 37(1): 554 − 565. WANG Xiaohuan, SHAO Jing’an, WANG Jinliang, et al. Water quality assessment and its changing trends in the reservoir inflow and outflow along the Yangtze River mainstream in the Three Gorge Reservoir Area [J]. Acta Sci Circumstant, 2017, 37(1): 554 − 565. [19] WHITE M S, XENOPOULOS M A, HOGSDEN K, et al. Natural lake level fluctuation and associated concordance with water quality and aquatic communities within small lakes of the Laurentian Great Lakes region [J]. Hydrobiologia, 2008, 613(1): 21 − 31. [20] 王丽婧, 翟羽佳, 郑丙辉, 等. 三峡库区及其上游流域水污染防治规划[J]. 环境科学研究, 2012, 25(12): 1370 − 1377. WANG Lijing, ZHAI Yujia, ZHENG Binghui, et al. Study on water pollution prevention planning for the Three Gorges Reservoir Area and its upper reaches [J]. Res Environ Sci, 2012, 25(12): 1370 − 1377. [21] SONDERGAARD M, JENSEN P J, JEPPESEN E. Retention and internal loading of phosphorus in shallow, eutrophic lakes [J]. Sci World J, 2001, 1: 427 − 442. [22] 张馨月, 马沛明, 高千红, 等. 三峡大坝上下游水质时空变化特征[J]. 湖泊科学, 2019, 31(3): 633 − 645. ZHANG Xinyue, MA Peiming, GAO Qianhong, et al. Spatial-temporal variations of water quality in upstream and downstream of Three Gorges Dam [J]. J Lake Sci, 2019, 31(3): 633 − 645. [23] ZHANG Yonggang, ZHANG Zhen, XUE Shuai, et al. Stability analysis of a typical landslide mass in the Three Gorges Reservoir under varying reservoir water levels[J]. Environ Earth Sci, 2020, 79(1): 42. doi: 10.1007/s12665-019-8779-x. [24] ZHAO Jian, FU Guo, LEI Kun, et al. Multivariate analysis of surface water quality in the Three Gorges area of China and implications for water management [J]. J Environ Sci, 2011, 23(9): 1460 − 1471. [25] XU Xibao, YANG Guishan, YAN Yan, et al. Unravelling the effects of large-scale ecological programs on ecological rehabilitation of China’s Three Gorges Dam[J]. J Cleaner Prod, 2020, 256: 120446. doi: 10.1016/j.jclepro.2020.120446. [26] ZHANG Quanfa, LOU Zhiping. The environmental changes and mitigation actions in the Three Gorges Reservoir region, China [J]. Environ Sci Policy, 2011, 14(8): 1132 − 1138. [27] JACKSON S, SLEIGH A C. Resettlement for China’s Three Gorges Dam: socio-economic impact and institutional tensions[J]. Communist Post-Communist Stud, 2000, 33(2): 223 − 241. [28] 娄保锋, 臧小平, 吴炳方. 三峡水库蓄水运用期化学需氧量和氨氮污染负荷研究[J]. 长江流域资源与环境, 2011, 20(10): 1268 − 1273. LOU Baofeng, ZANG Xiaoping, WU Bingfang. Study on pollution loads of CODCr, and ammonia-N into the Three Gorges Reservoir during operation period from first impoundment to completion [J]. Resour Environ Yangtze Basin, 2011, 20(10): 1268 − 1273. [29] 王丽婧, 李虹, 杨正健, 等. 三峡水库蓄水运行初期(2003−2012年)水环境演变特征的“四大效应”[J]. 环境科学研究, 2020, 33(5): 1109 − 1118. WANG Lijing, LI Hong, YANG Zhengjian, et al. Four effects of water environment evolution in early period (2003−2012) after impoundment of the Three Gorges Reservoir [J]. Res Environ Sci, 2020, 33(5): 1109 − 1118. [30] 李崇明, 黄真理. 三峡水库入库污染负荷研究(Ⅰ)蓄水前污染负荷现状[J]. 长江流域资源与环境, 2005, 14(5): 611 − 622. LI Chongming, HUANG Zhenli. Study on the pollutant loads into Three Gorges Reservoir (Ⅰ) pollutant load status before impoundment [J]. Resour Environ Yangtze Basin, 2005, 14(5): 611 − 622. [31] 黄真理. 三峡水库水环境保护研究及其进展[J]. 四川大学学报(工程科学版), 2006, 38(5): 7 − 15. HUANG Zhenli. Water quality protection and advances on Three Gorges Reservoir [J]. J Sichuan Univ Eng Sci Ed, 2006, 38(5): 7 − 15. [32] 江科, 王业春, 张晟, 等. 三峡库区小流域农村生活污水排放格局及污染物特征[J]. 环境科学与技术, 2015, 38(6): 39 − 43. JIANG Ke, WANG Yechun, ZHANG Sheng, et al. Emission pattern and characteristics of rural domestic sewage in the small watershed of the Three Gorges Reservoir Area [J]. Environ Sci Technol, 2015, 38(6): 39 − 43. [33] 秦延文, 赵艳民, 马迎群, 等. 三峡水库氮磷污染防治政策建议: 生态补偿·污染控制·质量考核[J]. 环境科学研究, 2018, 31(1): 1 − 8. QIN Yanwen, ZHAO Yanmin, MA Yingqun, et al. Prevention and control of nitrogen, phosphorus pollution in the Three Gorges Reservoir: ecological Compensation, pollution control, quality assessment [J]. Res Environ Sci, 2018, 31(1): 1 − 8. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200778
 点击查看大图
点击查看大图
计量
- 文章访问数: 1211
- HTML全文浏览量: 413
- PDF下载量: 31
- 被引次数: 0









