





-
全球变暖的持续加剧显著影响了陆地生态系统的生产力状况[1]。植被是陆地生态系统的核心,对物质循环、能量流动以及全球碳平衡具有决定性作用[2−3]。植被覆盖不仅可反映植被生长状况,也是评估生态系统健康的关键指标[4]。因此,深入研究植被覆盖的时空变化及其驱动因素,对于区域生态环境保护和植被恢复具有重要意义,是推动区域可持续健康发展的科学基础。
归一化植被指数(NDVI)是通过遥感技术反映地表植被覆盖度和生长状况的重要指标,已广泛应用于植被动态变化特征[4−6]等研究中。亦有不少学者基于NDVI数据采用像元二分模型(FVC)来探讨植被覆盖度的动态变化特征[7]。在不同区域尺度上植被覆盖对气温和降水变化的响应有很大的差异。例如,在北半球中高纬度地区,气温显著影响植被春季物候的变化[5, 8];在中国,气温升高对植被覆盖的促进作用逐渐增大,而降水则主要影响干旱、半干旱地区的植被状况[9];青藏高原地区[10]、黄土高原北部[11]以及西北干旱和半干旱地区[12]的植被生长主要受降水的影响,而东部沿海平原、黄土高原南部、四川盆地和云贵高原西南部对气温和太阳辐射的响应则更为明显[12−13]。
事实上,植被时空动态变化是气候和人为等因素共同作用的结果。在气候变化和生态修复工程实施的背景下,定量分离这2种因素对植被动态变化的影响是当前研究的热点问题。气候变化和人类活动对植被生长也会产生双重效应。祁鹏卫等[14]研究发现:在重庆市,过去20 a气温是影响NDVI变化的最重要因素,而陈晨等[15]认为:在黄河流域,人为活动在大部分气候区是植被改善的主导因素,而在干旱和半干旱的部分区域,气候变化对NDVI的影响更为明显。可见,气候变化和人类活动对植被的影响尚无定论。
延安市位于黄土高原腹部和黄河中游,水土流失严重,生态环境极其脆弱,是陕西省实施退耕还林还草和黄土高原水土流失综合治理工程的核心区,生态区位十分重要。已有研究表明:延安市自1999年持续开展退耕还林还草工程以来,植被覆盖状况得到了显著改善[16]。现有研究大多围绕黄土高原及黄河流域植被与气候的关系进行了系统分析[17−19],但针对延安市不同时间尺度上的研究还较少。
鉴于此,本研究基于延安市2001—2020年的MODIS-NDVI和同期气象数据,采用Mann-Kendall趋势分析和偏相关分析,并结合残差分析,旨在探究以下2个问题:①延安市2000—2020年NDVI在退耕还林还草工程背景下的变化趋势有何特征?②延安市植被对气候变化和人类活动的响应有何特征?研究结果有助于进一步深化植被对气候变化和人类活动响应机制的理解,以期为当地未来生态环境建设和可持续发展提供科学指导。
-
延安市(35°21′~37°31′N,107°41′~110°31′E)位于陕西省北部、黄土高原的中南部地区,总面积为37 037 km2。该市属黄土高原丘陵沟壑区,地形以高原与丘陵为主,平均海拔为1 200 m,地势“西北高、东南低”。地貌以黄土梁峁、沟壑为主,主要分布于延安北部,占全市总面积的70%。延安市属于暖温带半干旱大陆性季风气候,其中北部为半干旱地区,南部属半湿润地区。受季风环流及地理位置的影响,春秋季干燥少雨,气温波动大;夏季则高温多雨,雨热同期,降水集中;冬季气温较低,且降水稀少。作为全国退耕还林还草工程的关键实施区域,延安市在生态环境保护方面取得了显著成效。据延安市林业局数据显示: 2016—2020年全市累计治理水土流失面积为156.00万 hm2,累计完成营造林为20.90万 hm2。目前,全市共有林业用地面积为298.24万 hm2,森林覆盖率达48.07%。
-
延安市2001—2020年的MODIS-NDVI数据集来自于美国国家航空航天局(
http://ladsweb.modaps.eosdis.nasa.gov ),空间分辨率为250 m,时间分辨率为16 d,统一选取时间跨度为2001—2020年,共460景影像。本研究利用Savitzky-Golay滤波对原始数据进行去噪处理,迭代次数为3,滑动窗口大小为5,重构NDVI序列集。延安市及其周边30个气象站点的同期月降水与月平均气温资料来源于中国气象数据网。为了研究植被与气温、降水的响应关系,选用薄盘光滑样条函数法,基于气象数据曲面拟合的专用软件ANUSPLIN 4.3对延安市气温和降水量的站点数据进行插值。ANUSPLIN软件运行过程中提供了判断插值模型质量的统计参数,可用于确定拟合过程中的最优光滑参数,在插值过程中,考虑到气温受海拔的影响较大,因此在气温插值过程中以高程为协变量进行插值,降水在插值过程中则进行平方根转换以保证其插值非负。具体模型选择如表1所示。
表 1 气象要素空间插值的模型选择
Table 1. Model selection for spatial interpolation of meteorological elements
气象要素 函数选择 变量 协变量 数据转换 样条次数 气温 三变量局部薄盘光滑样条函数 经度、纬度 高程 无 3 降水 双变量薄盘光滑样条函数 经度、纬度 无 平方根转换 2 数字高程模型(DEM)为ASTER GDEM数字高程数据产品,来源于地理空间数据云(
http://www.gscloud.cn/ ),分辨率为30 m。土地利用类型数据来源于中国科学院资源环境科学与数据中心(https://www.resdc.cn/ ),包含2000、2005、2010、2015和2020年5期数据。为研究方便,所有数据统一为WGS1984坐标系,250 m×250 m空间分辨率。 -
目前,Theil-Sen Median方法与Mann-Kendall检验结合已被广泛应用于对植被长时间序列数据的变化趋势分析方面[20]。Theil-Sen Median趋势分析是一种稳健的趋势统计方法,可反映NDVI、气温或降水的变化趋势。Mann-Kendall检验是一种非参数统计检验,本研究用该检验来衡量NDVI、气温和降水序列趋势变化的显著性。它的优点是样本不需要服从某些分布,并且不受离群值的干扰,此方法的具体原理请参考文献[20−21]。
-
采用偏相关分析方法分析延安市年、季节和月NDVI对气候因子的响应。偏相关分析可以将其他要素的影响视为常数,计算某2个变量的相关性,具体原理请参考文献[22]。本研究将置信度水平为0.05的相关性分为不显著正(负)相关和显著正(负)相关4类。
-
残差分析是定量化区分气候变化和人类活动对植被覆盖变化影响最常用的方法[23−24]。该方法首先基于年NDVI、年平均气温和年累计降水量时间序列数据,NDVI作为因变量,以气温和降水量作为自变量,建立二元线性回归模型,计算模型中的各项参数;并基于气温和降水量数据以及回归模型的参数,计算NDVI的预测值(INDVpre),用来表示气候因素对NDVI的影响;最后计算NDVI的观测值(INDVobs)与INDVpre的差值,即NDVI残差(INDVres),借此表征人类活动等其他因素对NDVI变化的影响(以下统称为人类活动)[12]。具体计算公式如下:
$$ {I_{{\text{NDVpre}}}}(i,j) = a \times P(i,j) + b \times T(i,j) + c \text{;} $$ (1) $$ I_{\text{NDVres}} = I_{\text{NDVobs}}-I_{\text{NDVpre}} 。 $$ (2) 式(1)~(2)中:i为像元,j为年份,INDVpre(i, j)为i像元j年份NDVI预测值;P(i, j)为i像元j年份降水量(mm);T(i, j)为i像元j年份平均气温(℃);a和b为回归系数,c为回归常数项。
基于Theil-Sen Median趋势分析法对INDVpre、INDVobs、INDVres的变化趋势进行分析,根据三者的变化斜率(θ)划分植被变化驱动因素,并量化气候因素和人类活动对植被变化的相对贡献率[25],具体判断标准及相对贡献率计算方法见表2。
表 2 气候变化和人类活动对NDVI变化贡献率计算方法
Table 2. Methods for assessing the relative roles of climate variations and human activities in the process of NDVI change
θ(INDVobs) θ(INDVpre) θ(INDVres) 气候变化贡献率/% 人类活动贡献率/% 含义 >0 >0 >0 $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ 气候变化和人类活动驱动植被覆盖增加 >0 <0 100 0 气候变化驱动植被覆盖增加 <0 >0 0 100 人类活动驱动覆盖植被增加 <0 <0 <0 $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ 气候变化和人类活动驱动植被覆盖减少 <0 >0 100 0 气候变化驱动植被覆盖减少 >0 <0 0 100 人类活动驱动植被覆盖减少 说明:θ(INDVpre)、θ(INDVobs)、θ(INDVres)分别为NDVI预测值、观测值、残差的变化斜率。θ>0表示植被受到驱动因素的正向促进作用,θ<0表示植被受到驱动因素的反向抑制作用。 -
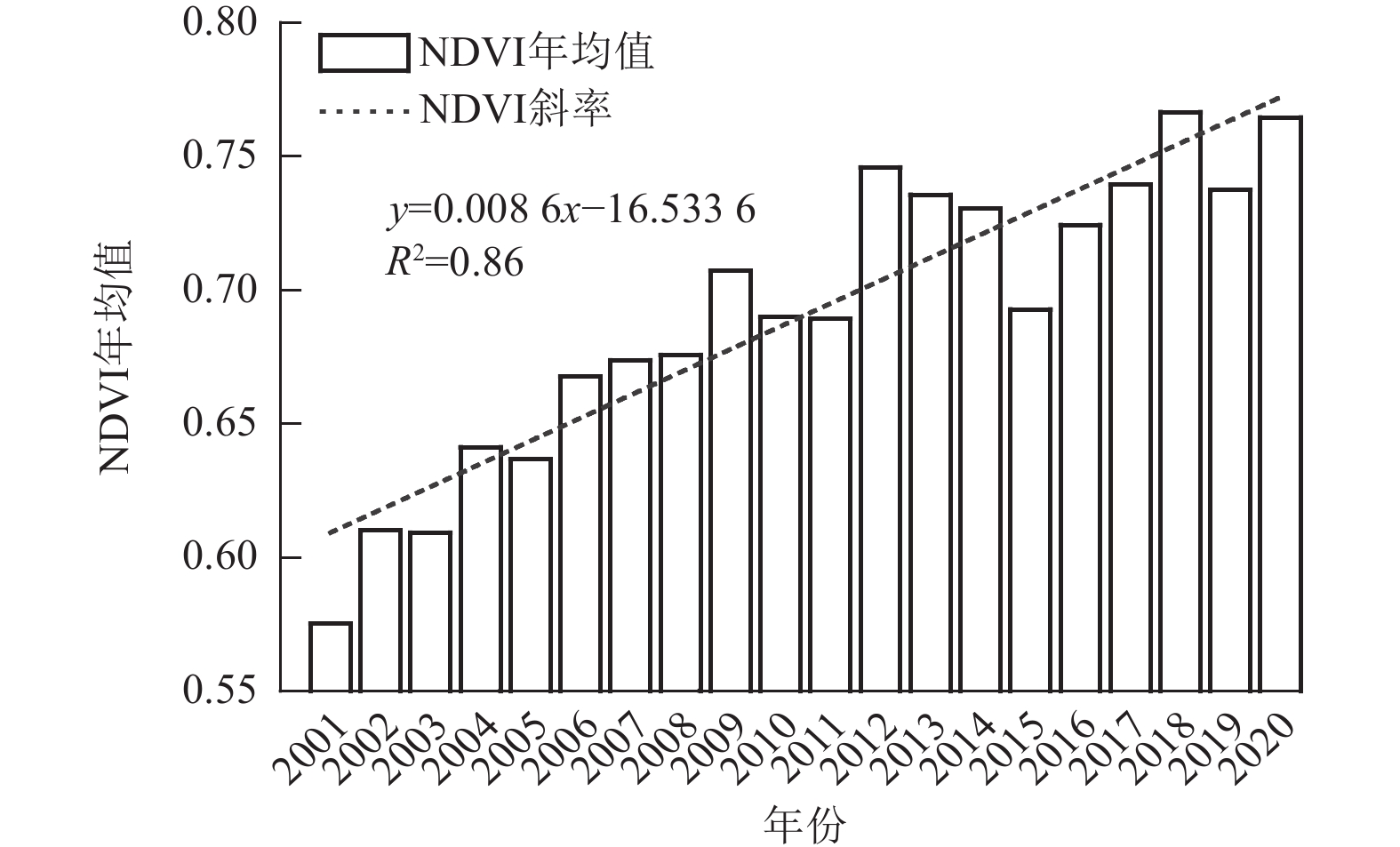
如图1所示:延安市近20 a的NDVI年均值为0.69,呈明显上升趋势,其变化速率为0.86×10−2 a−1(R2=0.86)。具体而言,2001—2009年NDVI上升趋势明显且增速较快;2010—2014年则表现为波动上升,变化相对平缓;而2015—2020年NDVI增长趋势再度提升,其中2015年出现阶段性谷值。在季节尺度上,延安市NDVI的变化趋势与全年趋势相吻合。
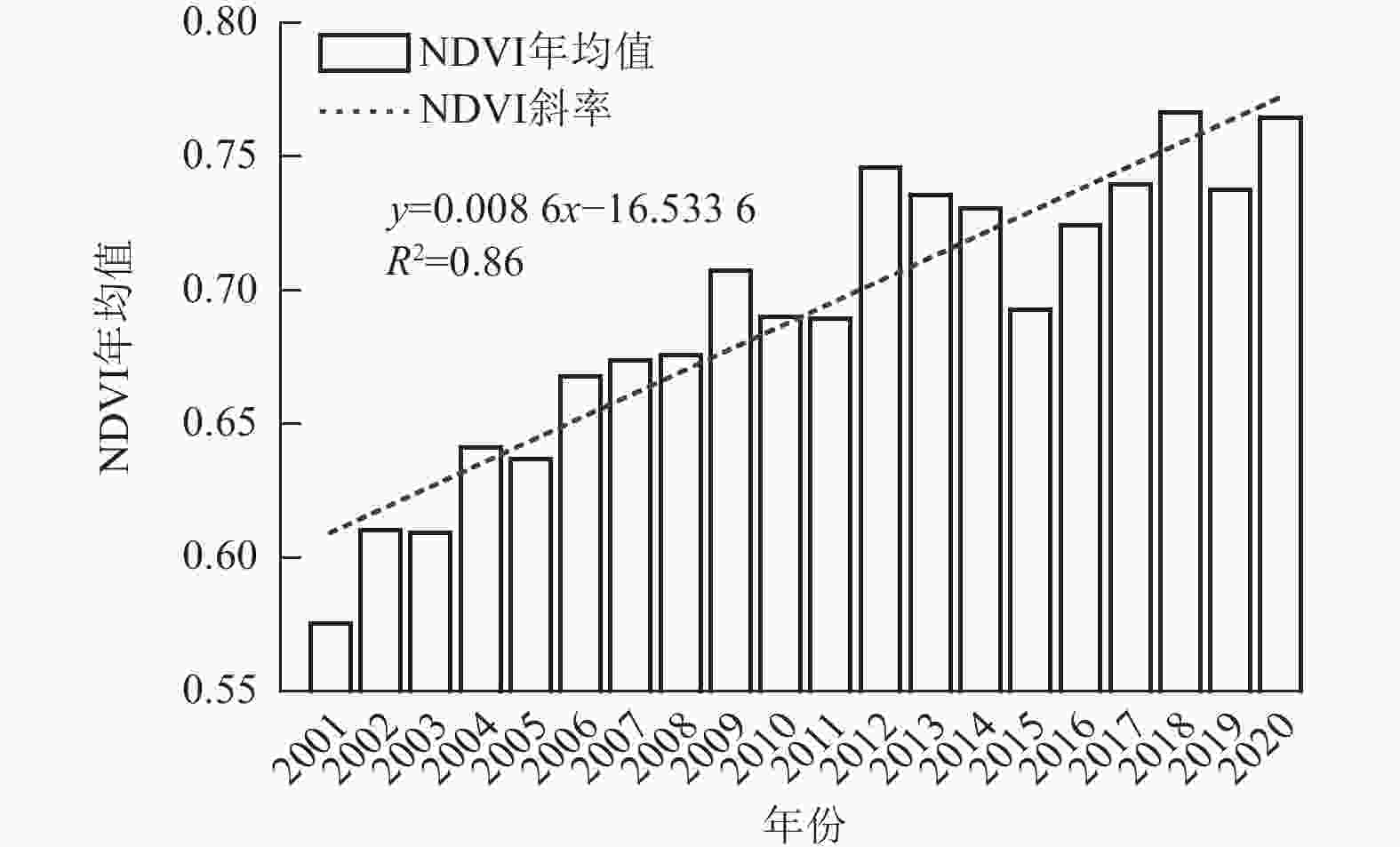
图 1 2001—2020年延安市NDVI年均值变化特征
Figure 1. Characteristics of annual NDVI mean values in Yan’an City from 2001 to 2020
-
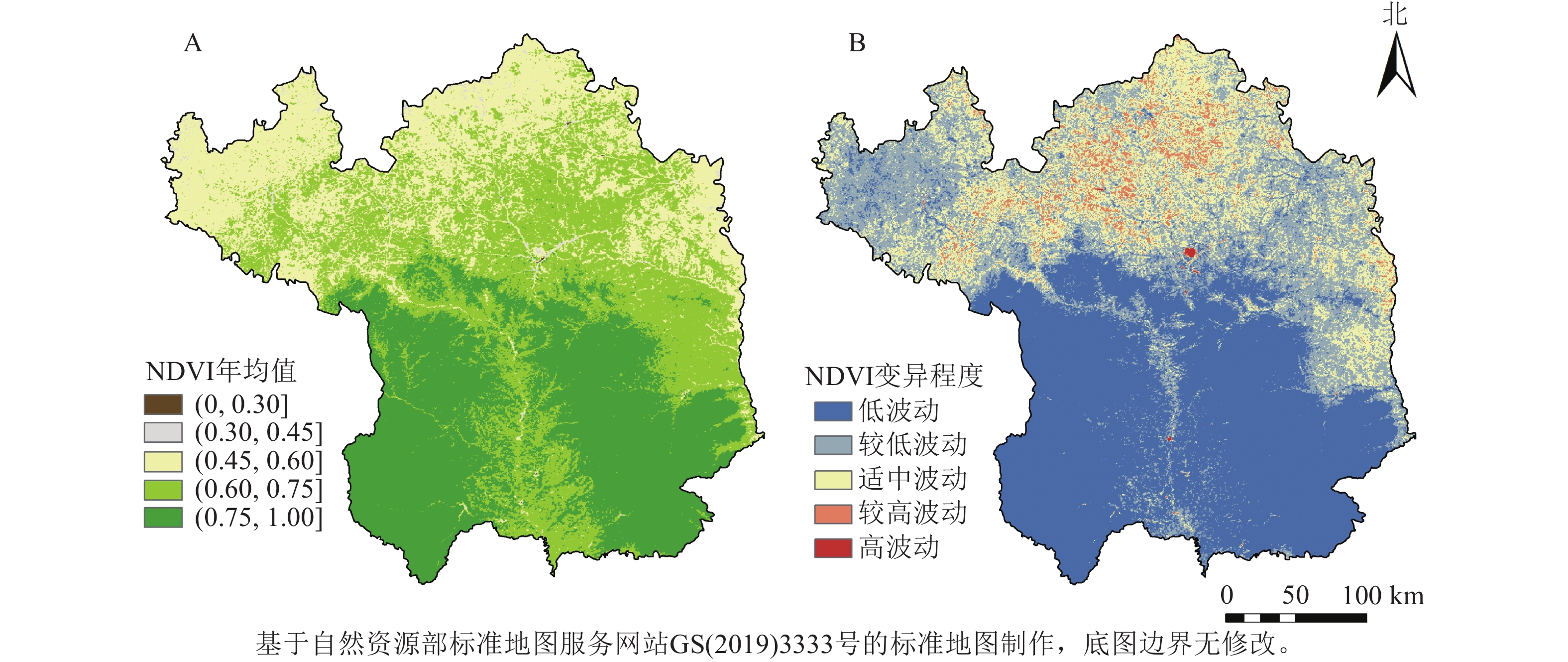
延安市NDVI整体呈“南部高、北部低”的空间分布特征(图2),NDVI均值为(0.75,1]的面积占比最多,为35.82%,主要分布在南部大部分区域;NDVI年均值低值区主要分布在延安北部的黄土丘陵沟壑区。延安市NDVI处于低波动水平等级[NDVI变异系数(CV)≤0.10]的面积占比最大,为45.91%,主要分布在研究区南部,该区域植被类型多为连续分布的草地和林地,受人类活动的干扰较小;处于较高和高波动等级(CV≥0.30)的区域所占研究区面积最小,为0.07%,主要分布在延安市北部,即退耕还林草工程的重点实施区。
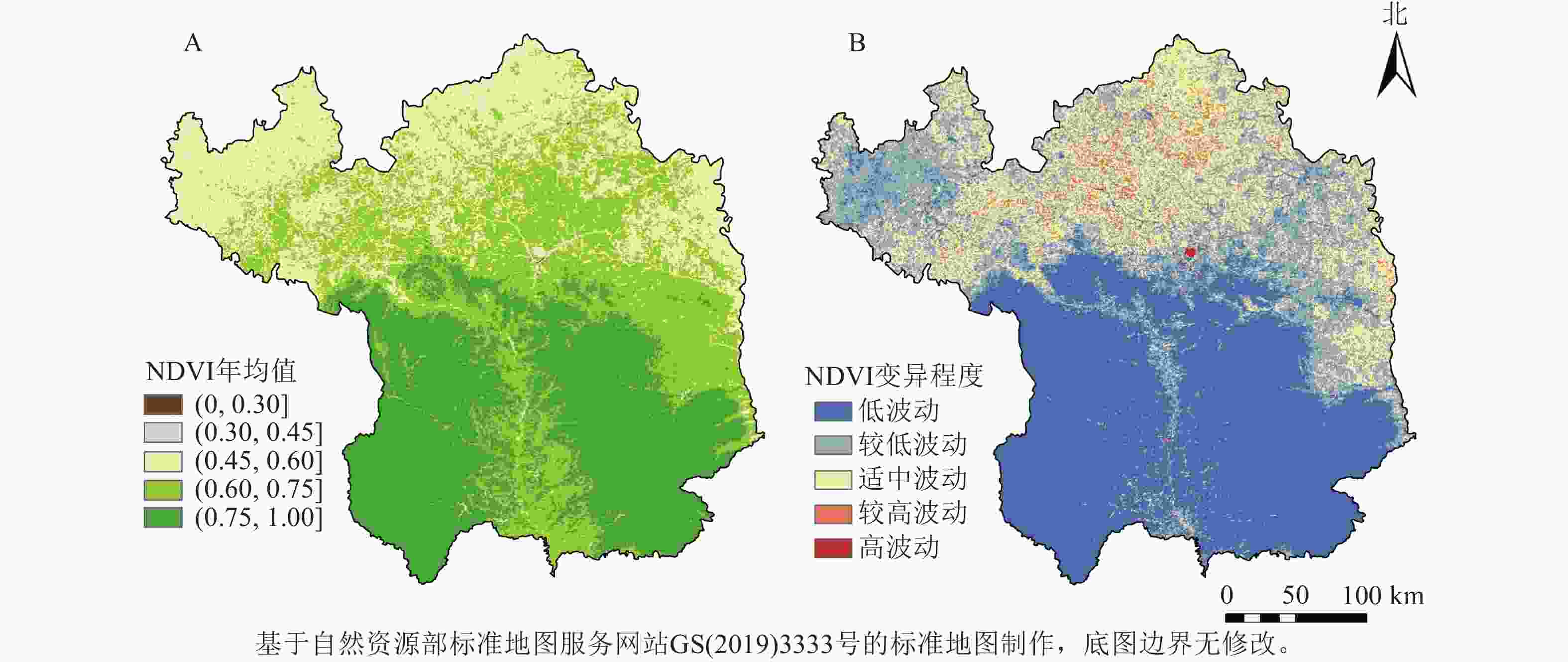
图 2 2001—2020年延安市植被NDVI年均值及变异程度空间分布示意图
Figure 2. Spatial distribution of mean annual NDVI values and degree of variability of vegetation in Yan’an City from 2001 to 2020
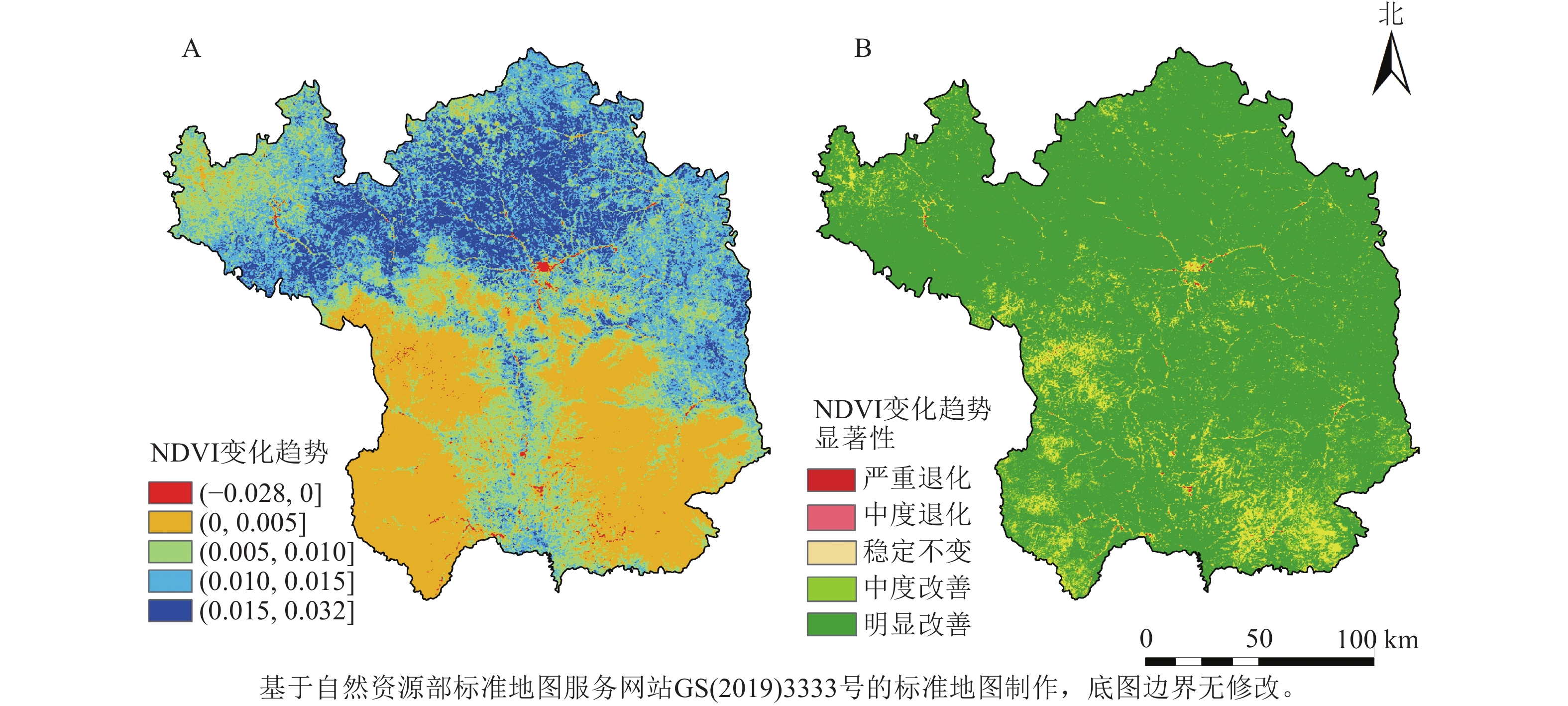
将2001—2020年NDVI变化斜率(β)介于−0.000 5~0.000 5定义为稳定不变区域;低于−0.000 5定义为退化区域;高于0.000 5定义为改善区域。对延安地区NDVI变化进行Mann-Kendall检验,其中置信度在0.05和0.01水平下相应的Z值分别为1.96和2.58,将Mann-Kendall检验结果划分为极显著变化(Z≥2.58或Z≤−2.58)、显著变化(Z>1.96或Z<−1.96)和稳定不变(−1.96≤Z≤1.96),与Theil-Sen Median趋势分析结果叠加分析,得到延安地区年NDVI变化趋势(图3),对季节NDVI变化趋势进行分级统计得到表3。结果显示:延安市NDVI整体呈现显著好转趋势,其中明显改善的区域占比最高,达83.27%。尤为突出的是,植被覆盖度原本较低的北部区域,在研究期间NDVI得到显著改善。严重退化和中度退化的区域零星分布于研究区周边,共占研究区总面积的0.17%。在季节尺度上,春季、夏季、秋季和冬季NDVI明显改善的区域分别占总面积的87.29%、84.99%、76.42%和88.23%。
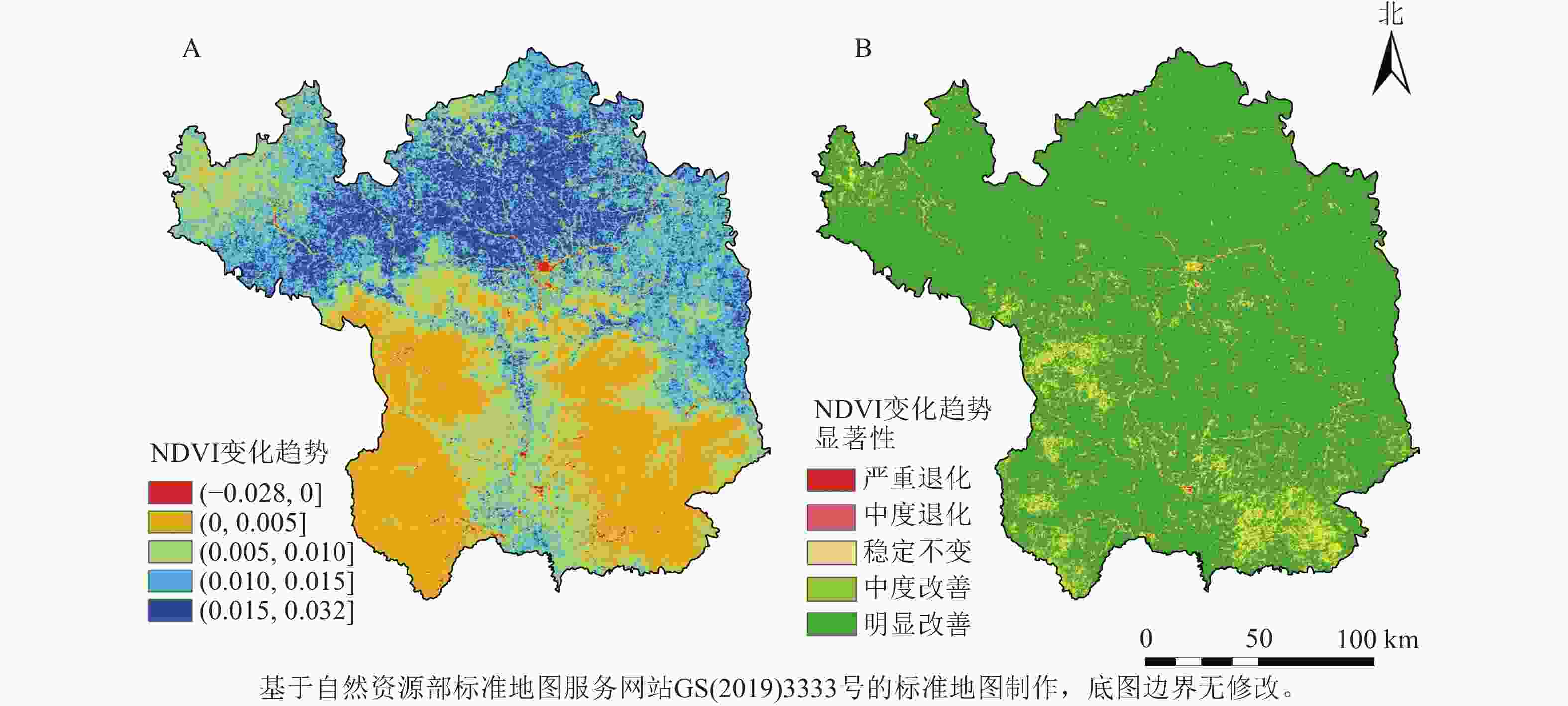
图 3 2001—2020年延安市年NDVI变化趋势及显著性示意图
Figure 3. Trends and significance of annually-averaged NDVI in Yan’an City from 2001 to 2020
表 3 2001—2020年延安市年与季节NDVI变化趋势统计
Table 3. Annual and seasonal NDVI trend statistics in Yan’an City from 2001 to 2020
NDVI变化趋势 β Z 所占面积比例/% 春季/% 夏季/% 秋季/% 冬季/% 显著上升 ≥0.000 5 ≥2.58 83.27 87.29 84.99 76.42 88.23 不显著上升 ≥0.000 5 1.96~2.58 8.59 7.70 8.01 9.13 6.74 稳定不变 −0.000 5~0.000 5 −1.96~1.96 7.97 4.89 6.86 14.33 5.00 不显著下降 <−0.000 5 −2.58~−1.96 0.09 0.08 0.08 0.07 0.02 显著下降 <−0.000 5 ≤−2.58 0.08 0.04 0.06 0.05 0.01 说明:β为NDVI变化斜率。 -
由图4可见:在年尺度上,延市NDVI与气温、降水的整体平均偏相关系数(r)分别为−0.17和0.21,NDVI与气温呈负相关(r<0)的面积占76.52%,与降水呈正相关(r>0)的面积占87.74%。对NDVI与气温、降水的偏相关系数进行t检验发现:NDVI与气温呈显著负相关(P<0.05)的区域集中在延安中部,即洛河和延河两岸;NDVI与降水呈显著正相关(P<0.05)的区域比例为6.96%,主要分布在西北地区。
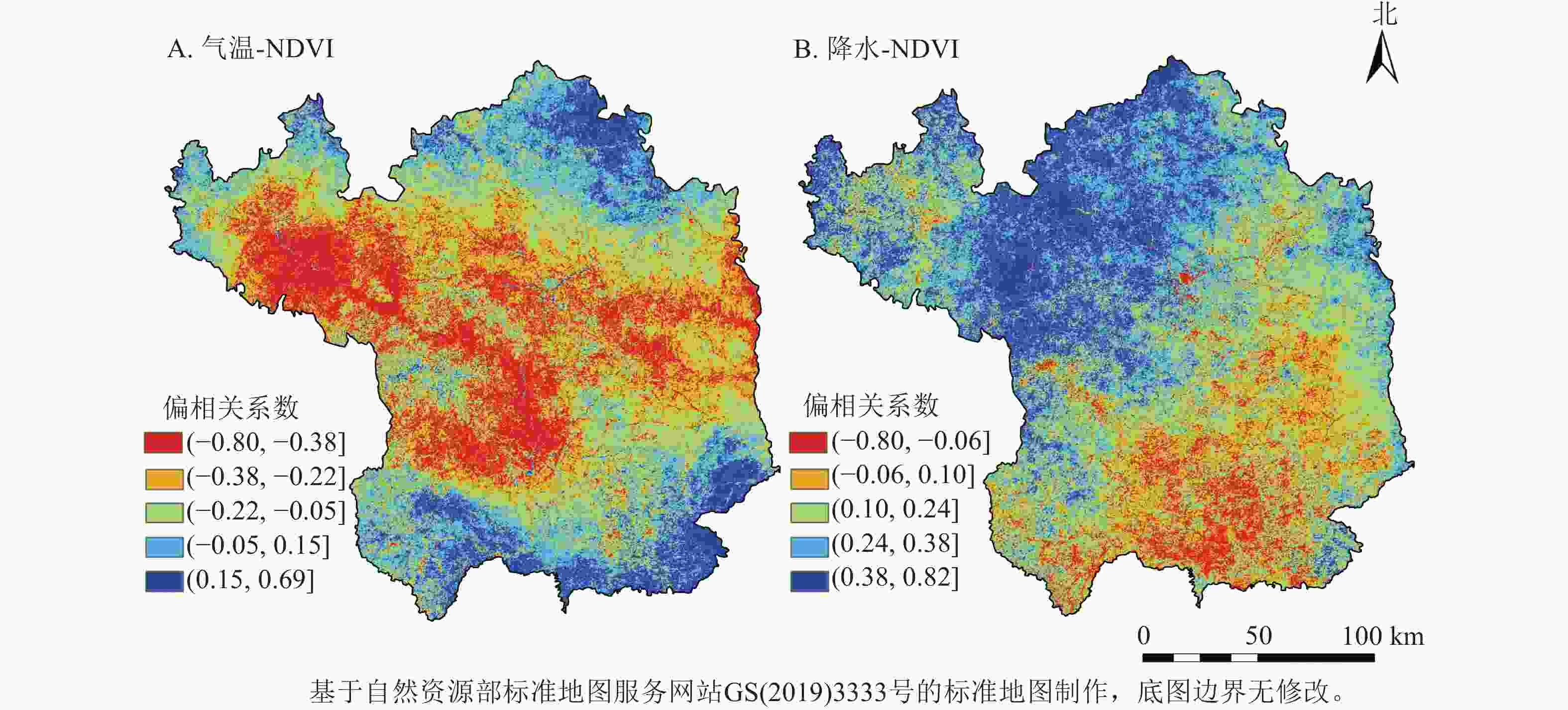
图 4 2001—2020年延安市年NDVI与年气温和降水的相关性示意图
Figure 4. Correlation of annual NDVI with annual temperature and precipitation in Yan’an City from 2001 to 2020
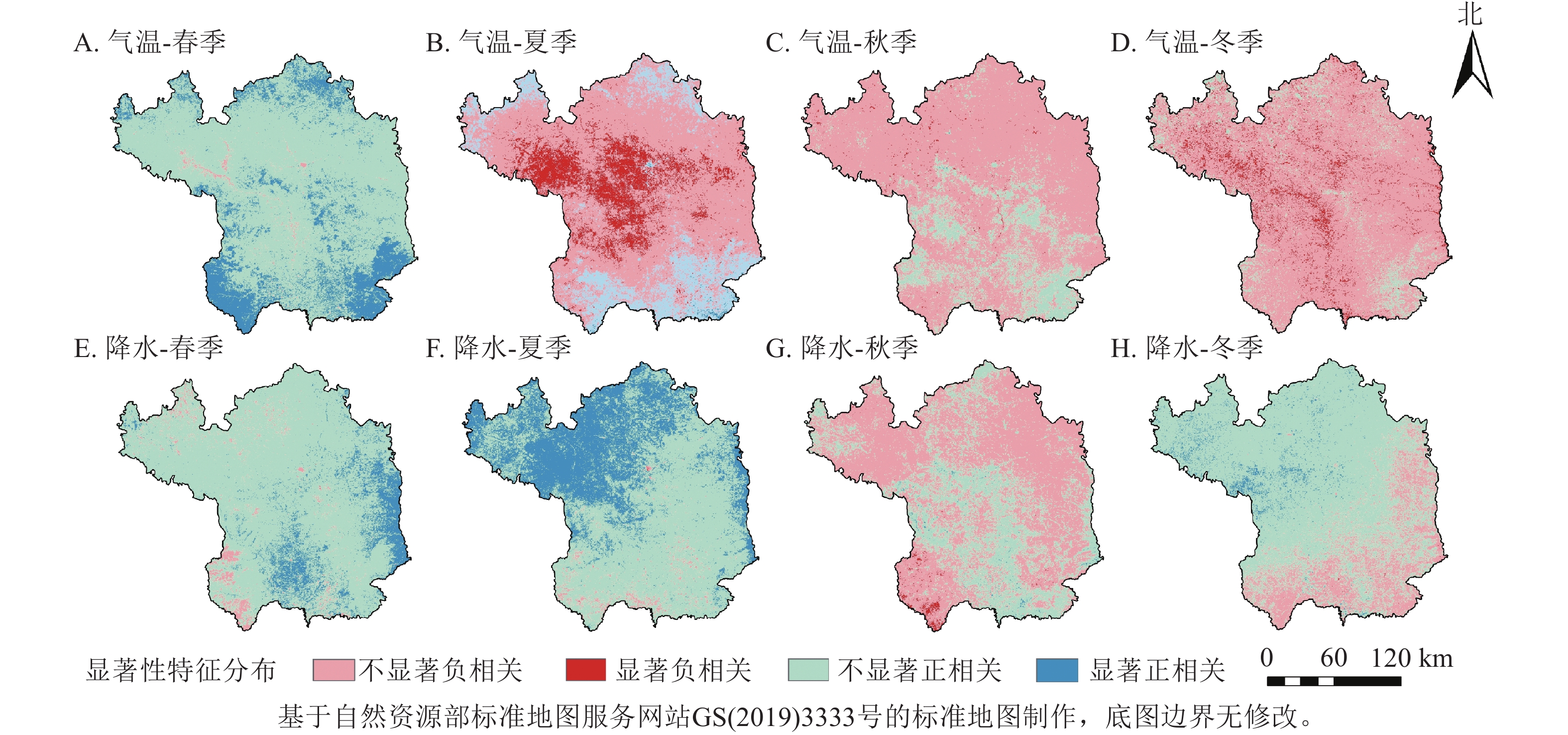
从季节尺度上看(图5),NDVI对气温和降水的响应存在明显的季节差异。春季气温和降水对NDVI存在正向作用,且春季大部分区域气温对NDVI的影响大于降水,说明春季气温对植被生长具有明显的促进作用;夏季大部分区域气温和降水与NDVI的相关性相反,降水对延安市北部和东部毗邻黄河区域的NDVI有明显的促进作用,但气温对北洛河和延河两岸的植被生长有明显的抑制作用;秋季大部分地区与冬季相似,气温和降水与NDVI的相关性都不显著。结合年与季节尺度来说,与气温相比,降水对延安市植被生长的促进作用更大。
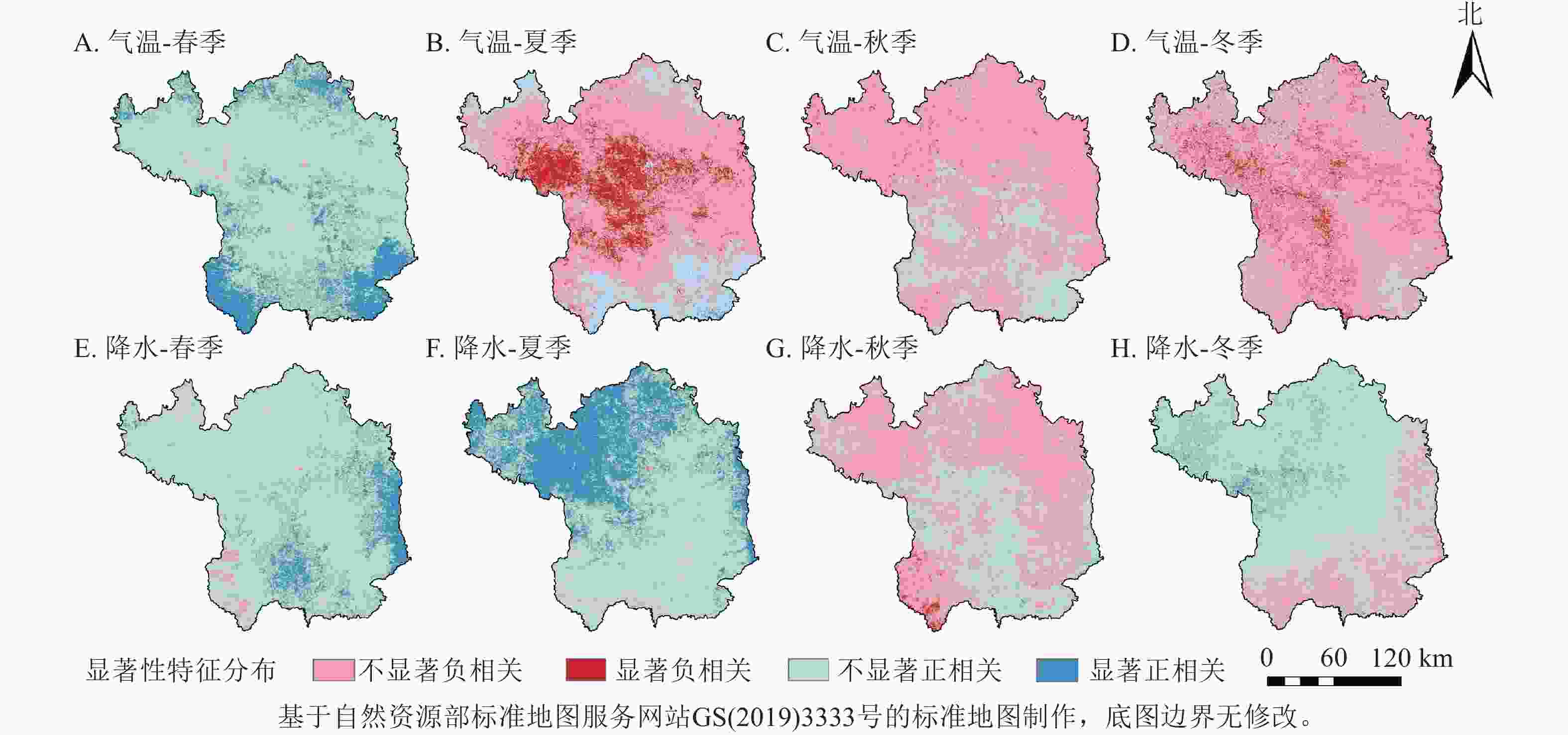
图 5 2001—2020年延安市季节NDVI与气温和降水的相关性示意图
Figure 5. Correlation of seasonal NDVI with temperature and precipitation in Yan’an City from 2001 to 2020
-
通过对比实际年份与预测年份的NDVI数据(R2=0.76),得到NDVI残差序列,并应用Theil-Sen中值趋势分析方法分析残差序列,以揭示人类活动对NDVI变化的影响(图6)。结果表明:近20 a间,延安市NDVI残差的变化范围为−0.28×10−2~0.29×102 a−1,其中98.88%的区域呈现正向残差,表明人类活动对NDVI增长具有正向促进作用。特别是NDVI残差年变化率为0.001~0.015 a−1,超过0.015 a−1的区域占总面积的75.97%。
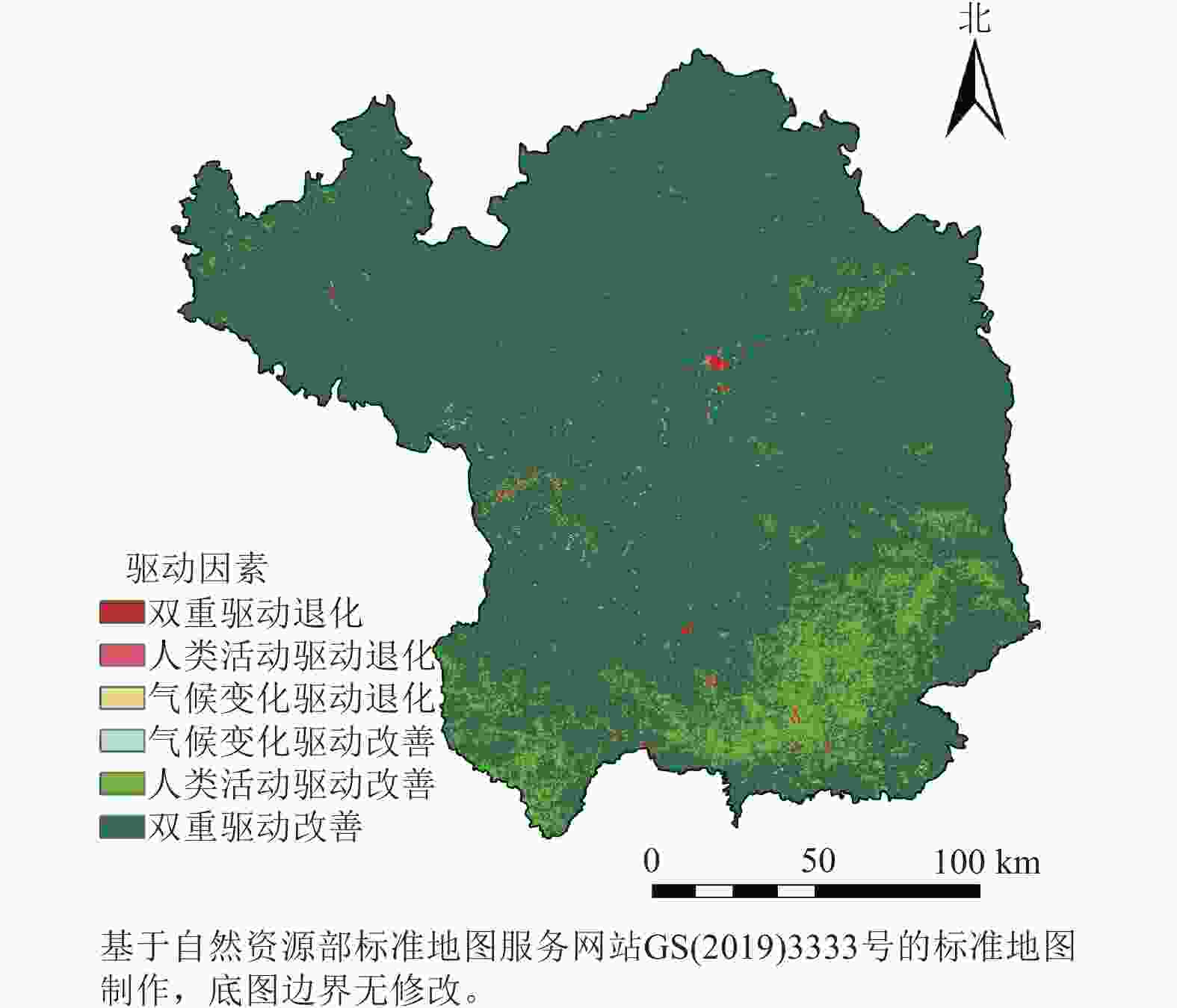
图 6 2001—2020年延安市植被变化驱动因素空间分布示意图
Figure 6. Spatial distribution of driving factor of vegatation change in Yan’an City from 2001 to 2020
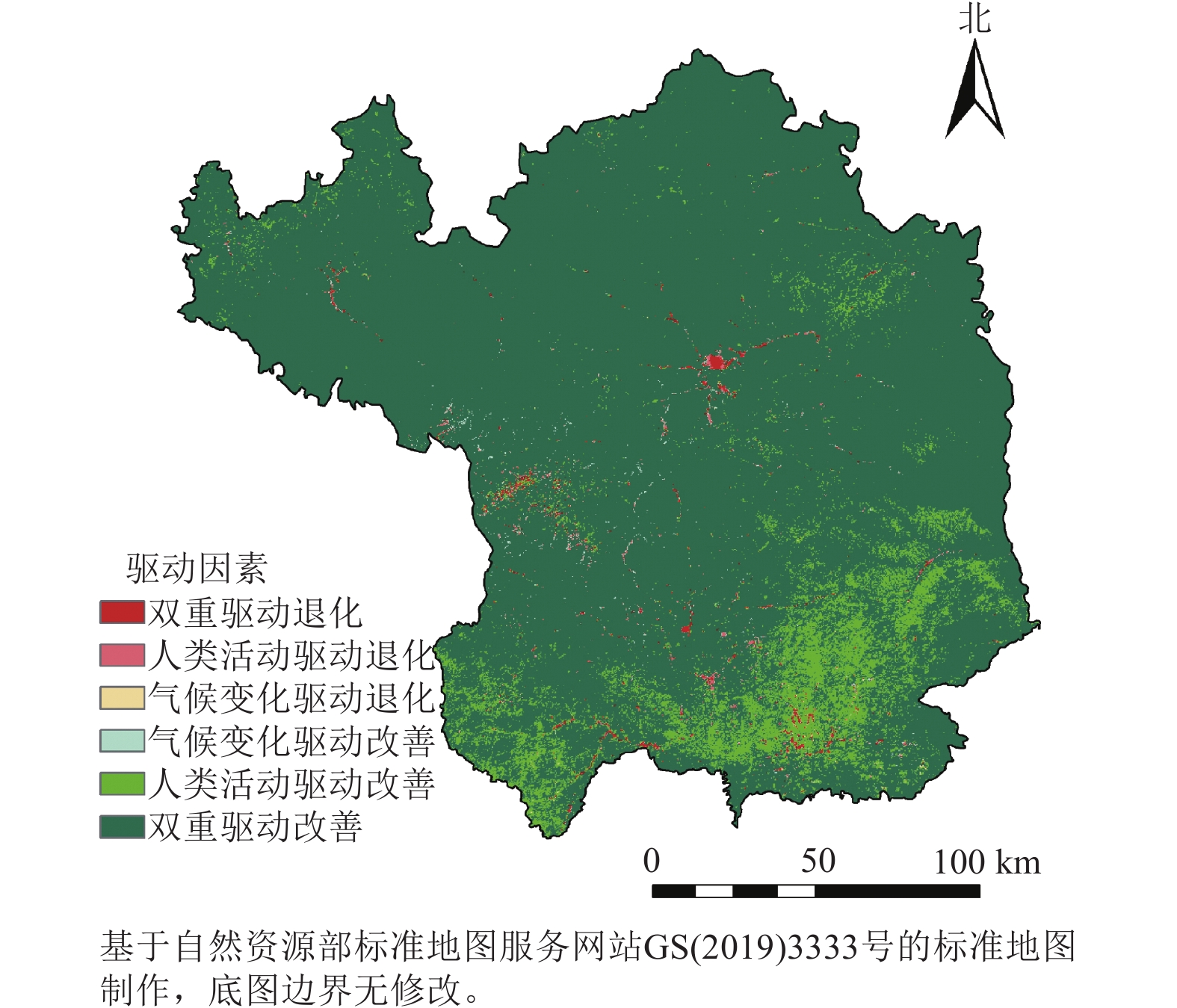
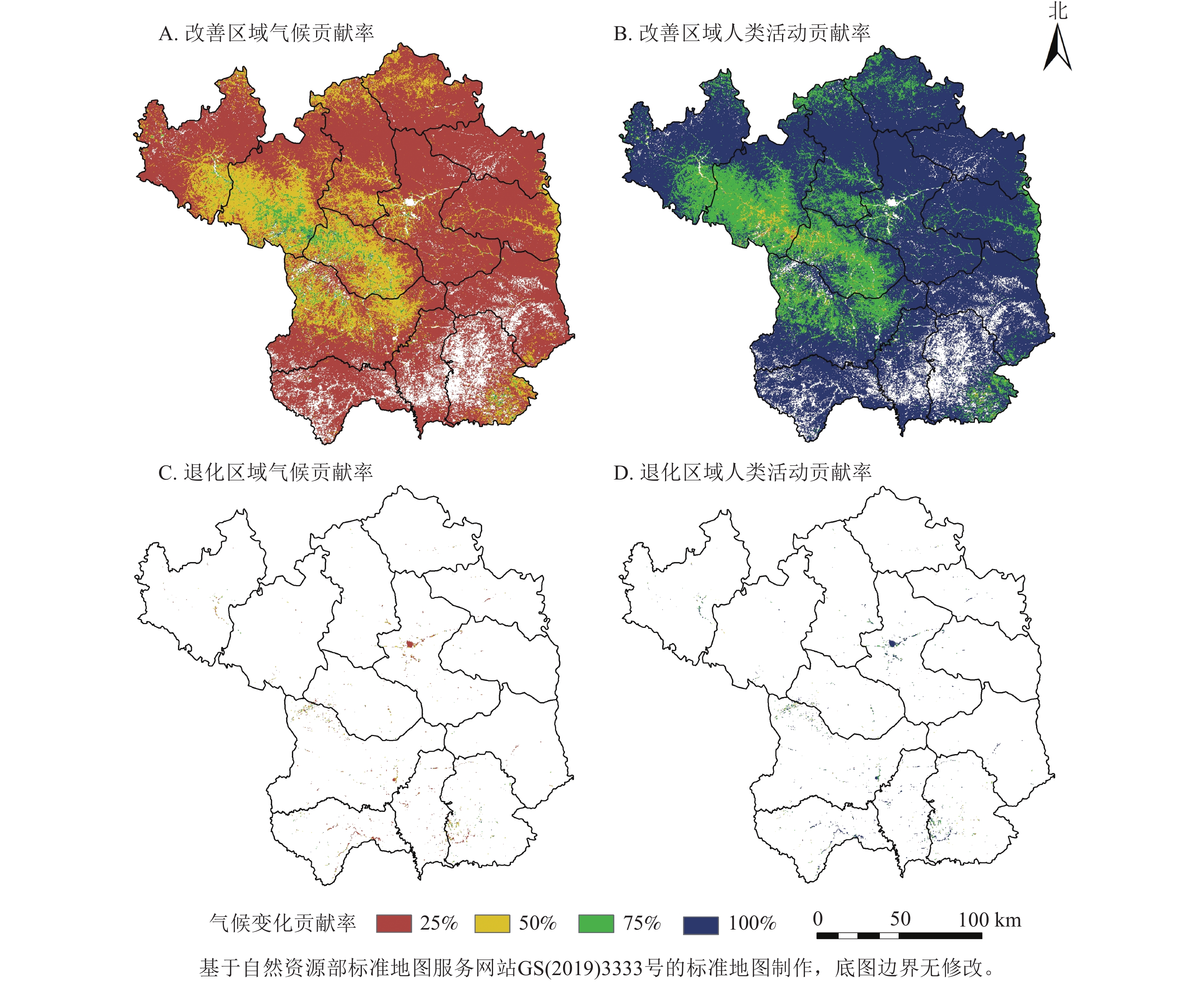
为深入解析气候变化与人类活动对植被生长的具体影响,本研究将NDVI上升趋势定义为植被改善,下降则为退化。延安市植被改善区域占比高达98.88%,其中90.72%的区域由两者共同驱动,广泛分布于延安大部分地区;7.81%的区域主要受人类活动单独影响,集中于延安市南部;而仅0.34%的区域单独受气候变化影响呈现改善。植被退化区域仅占1.22%,其中0.54%由两者共同导致,单独由气候变化和人类活动引起的退化分别占0.13%和0.46%,均呈零散分布。综上所述,植被变化主要受气候变化与人类活动的共同调控。
如图7所示:气候变化对植被的相对贡献率为17.81%,人类活动则为82.19%。在植被改善区,两者的贡献率分别为17.77%和82.23%。具体而言,人类活动对NDVI增长贡献率超过80%和60%~80%的区域,分别占延安市总面积的58.15%和26.86%,主要位于中部和北部;而气候变化的贡献率大多低于20%。这表明人类活动是驱动延安市NDVI增长的主导因素。对于植被退化区,气候变化和人类活动的贡献率分别为24.63%和75.37%,尽管退化区域占比极小,但人类活动对植被NDVI退化贡献率超过80%的区域仍占总面积的0.28%。
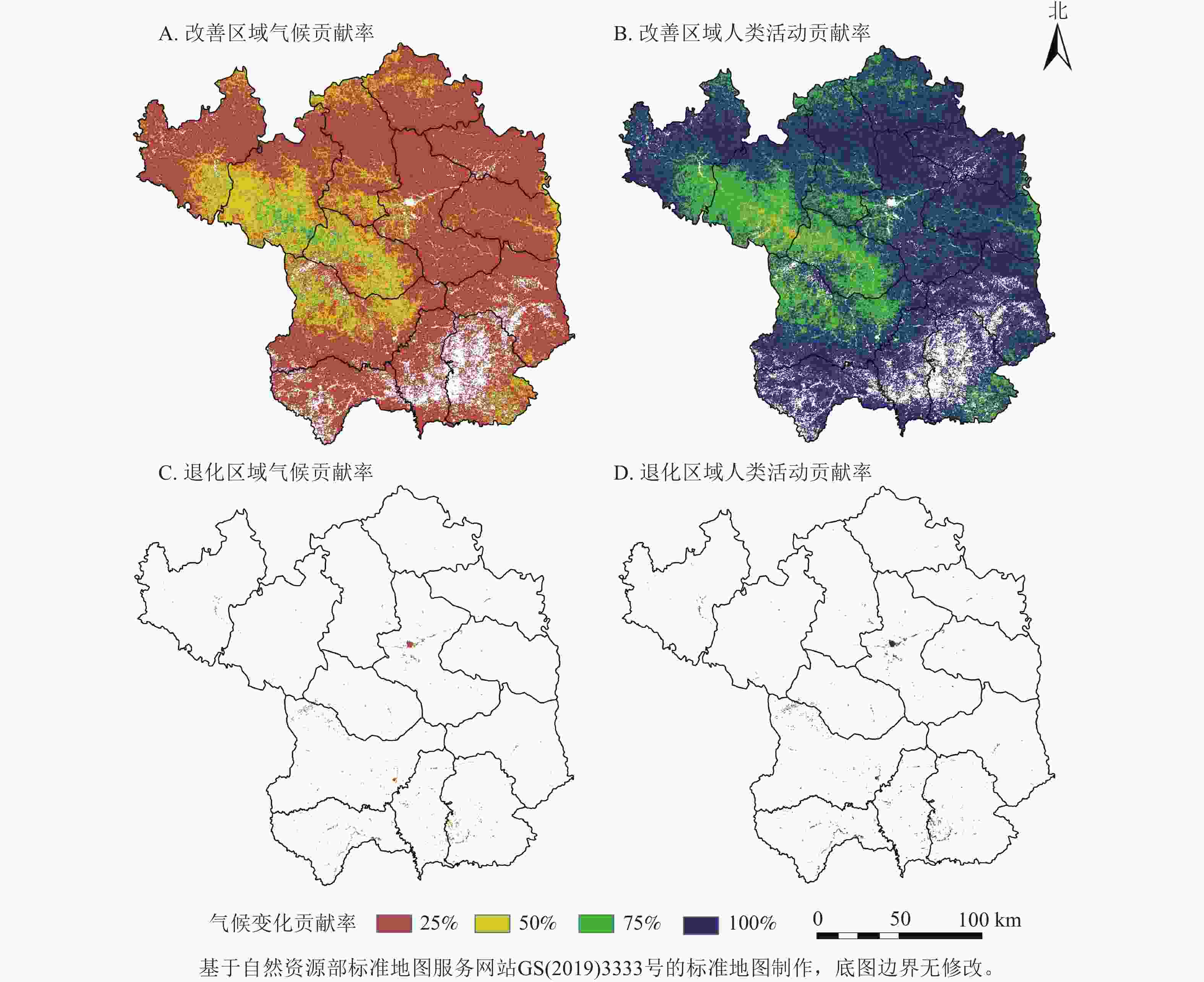
图 7 气候变化和人类活动对植被变化的贡献率
Figure 7. Contribution rate of climate change and human activities to vegatation change
-
本研究发现:近20 a来,延安市NDVI虽存在年际波动,但总体呈改善趋势,这与张小勇等[16]和张家政等[19]的研究结果相符。空间上,延安市北部地区NDVI显著提升,归因于该区域是退耕还林还草工程的重点区域。本研究基于多期土地利用图(2001、2005、2010、2015、2020年),分析了不同土地利用类型下NDVI的变化,结果显示各土地利用类型的NDVI均呈增加态势,表明植被状况整体向好。此外,尽管耕地面积减少,但NDVI均值由0.50增至0.73,反映了退耕还林还草工程促进耕地向林地、草地转变的同时,也提升了耕地生态环境质量。
本研究NDVI对气候因子的响应呈现明显的空间异质性。延安市植被对降水响应更为明显,多数区域与降水呈正相关,这与李依璇等[25]和刘旻霞等[26]的研究结果一致。而气温与NDVI在洛河和延河附近区域呈显著负相关,尤以夏季为甚,这与梁冰洁[23]关于洛河流域水分利用效率与气温呈负相关的研究结果相呼应。水分利用效率可以有效反映植被的光合作用与耗水特征之间的关系,高温加速土壤水分流失,在水资源匮乏地区抑制植被生长。
如何定量区分自然因素与人为因素对NDVI变化的影响是当前研究的热点问题[27−28]。本研究表明:延安市植被NDVI增加是自然与人为因素共同作用的结果,且人类活动为主要驱动力,这与张乐艺等[28]的结论一致。人类活动对植被的影响具有双面性:一方面,林业生态工程促进了植被面积的大幅增加;另一方面,城市化进程中土地扩张对植被恢复构成挑战。总体而言,人类活动的正面影响远超负面影响。
NDVI对气候因子的响应是一个复杂的过程,研究结果往往与研究尺度、数据分辨率和研究方法密切相关[26]。且气候因子对植被的影响不仅限于气温和降水,饱和水汽压、日照时数等因子同样会对NDVI产生影响。未来研究可构建退耕还林还草工程前后的NDVI与气候因子回归模型,以更精确地分离自然与人为因素对NDVI变化的影响。
-
延安市NDVI年均值为0.69,NDVI大于0.60的区域占延安市总面积的70%;空间上,NDVI呈“南部高、北部低”的特征。2001—2020年,NDVI年均值呈明显上升趋势,年增长率达0.86×10−2 a−1。延安市植被整体呈现向好态势,NDVI得到显著改善的区域占比高达83.27%,且春、夏、秋、冬季NDVI明显改善的区域面积占比均超过75.00%。时间尺度上,气温和降水对NDVI的影响存在明显差异;年尺度上,降水对NDVI的影响大于气温;季节尺度上,春季NDVI受气温和降水的正向影响,夏季则主要受降水的正向驱动和气温的负向驱动。气候变化和人类活动对NDVI具有双重效应,受两者共同影响的区域占比高达91.26%。相较于气候因素,人类活动对植被变化的驱动作用更为显著,其相对贡献率达82.19%,远高于气候因素的17.81%。
Spatiotemporal variation and driving factors of normalized difference vegetation index (NDVI) in Yan’an City
-
摘要:
目的 植被是陆地生态系统中的主体,研究区域植被覆盖时空变化并量化气候变化和人类活动的相对贡献,可为区域未来的生态环境建设和可持续发展提供科学指导。 方法 基于延安市2001—2020年的MODIS归一化植被指数(NDVI)数据集和同期气象数据,采用Mann-Kendall趋势分析和偏相关分析、残差分析探讨了NDVI对气候变化和人类活动的响应。 结果 ①2001—2020年,延安市NDVI年均值为0.69,整体呈明显上升趋势,空间上呈“南部高、北部低”的分布格局。②不同尺度上气温和降水对NDVI的影响存在明显差异。年尺度上,NDVI受降水的影响大于气温;季节尺度上,春季NDVI受到气温和降水的双重影响,夏季降水对NDVI的影响较大,秋、冬季大部分区域NDVI与气温、降水相关性不显著。③延安市NDVI整体呈现改善趋势,明显改善区域面积占比为83.27%。④气候变化和人类活动对植被变化的影响具有双重效应,受气候变化和人类活动共同影响面积占比为91.26%,两者对植被变化的相对贡献率分别为17.81%和82.19%。 结论 延安市植被整体呈向好态势,相较于气候因素,人类活动对NDVI变化的驱动作用更强。图7表3参28 -
关键词:
- 归一化植被指数(NDVI) /
- 驱动因素 /
- 气候变化 /
- 人类活动 /
- 延安市
Abstract:Objective Vegetation is a principal component of terrestrial ecosystems. This study intends to investigate the spatiotemporal variation in regional vegetation coverage, and quantify the relative contributions of climate change and human activities, which can provide scientific support for future ecological environment construction and sustainable development of the region. Method Based on the MODIS-normalized difference vegetation index (NDVI) dataset and meteorological data of Yan’an City from 2001 to 2020, Mann-Kendall trend analysis, partial correlation analysis and residual analysis were employed to investigate the response of NDVI to climate change and human activities. Result (1) From 2001 to 2020, the annual mean NDVI value in Yan’an City was 0.69, indicating a significant upward trend, with a spatial distribution pattern of high in the south and low in the north. (2) The impact of temperature and precipitation on NDVI varied significantly at different temporal scales. At the annual scale, NDVI was influenced more by precipitation than temperature. At the seasonal scale, NDVI in spring was influenced by both temperature and precipitation, while precipitation in summer had a greater impact on NDVI. In autumn and winter, the correlation between NDVI and temperature and precipitation was not significant in most regions. (3) The overall NDVI exhibited a trend of improvement, with 83.27% of the area showing significant improvement. (4) The impact of climate change and human activities on vegetation had a dual effect, with 91.26% of the area affected by both factors. The relative contributions of the two to vegetation change were 17.81% and 82.19%, respectively. Conclusion The overall vegetation in Yan’an City demonstrates a positive trend, and compared to climatic factors, human activities have a stronger driving effect on NDVI changes. [Ch, 7 fig. 3 tab. 28 ref.] -
图 1 2001—2020年延安市NDVI年均值变化特征
Figure 1 Characteristics of annual NDVI mean values in Yan’an City from 2001 to 2020
图 2 2001—2020年延安市植被NDVI年均值及变异程度空间分布示意图
Figure 2 Spatial distribution of mean annual NDVI values and degree of variability of vegetation in Yan’an City from 2001 to 2020
图 3 2001—2020年延安市年NDVI变化趋势及显著性示意图
Figure 3 Trends and significance of annually-averaged NDVI in Yan’an City from 2001 to 2020
图 4 2001—2020年延安市年NDVI与年气温和降水的相关性示意图
Figure 4 Correlation of annual NDVI with annual temperature and precipitation in Yan’an City from 2001 to 2020
图 5 2001—2020年延安市季节NDVI与气温和降水的相关性示意图
Figure 5 Correlation of seasonal NDVI with temperature and precipitation in Yan’an City from 2001 to 2020
图 6 2001—2020年延安市植被变化驱动因素空间分布示意图
Figure 6 Spatial distribution of driving factor of vegatation change in Yan’an City from 2001 to 2020
图 7 气候变化和人类活动对植被变化的贡献率
Figure 7 Contribution rate of climate change and human activities to vegatation change
表 1 气象要素空间插值的模型选择
Table 1. Model selection for spatial interpolation of meteorological elements
气象要素 函数选择 变量 协变量 数据转换 样条次数 气温 三变量局部薄盘光滑样条函数 经度、纬度 高程 无 3 降水 双变量薄盘光滑样条函数 经度、纬度 无 平方根转换 2  下载: 导出CSV
下载: 导出CSV
表 2 气候变化和人类活动对NDVI变化贡献率计算方法
Table 2. Methods for assessing the relative roles of climate variations and human activities in the process of NDVI change
θ(INDVobs) θ(INDVpre) θ(INDVres) 气候变化贡献率/% 人类活动贡献率/% 含义 >0 >0 >0 $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ 气候变化和人类活动驱动植被覆盖增加 >0 <0 100 0 气候变化驱动植被覆盖增加 <0 >0 0 100 人类活动驱动覆盖植被增加 <0 <0 <0 $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ $ \dfrac{{{\text{|}}\Delta I_{\text{NDVpre}}|}}{{{\text{|}}\Delta I_{\text{NDVpre}}| + {\text{|}}\Delta I_{\text{NDVres}}|}} $ 气候变化和人类活动驱动植被覆盖减少 <0 >0 100 0 气候变化驱动植被覆盖减少 >0 <0 0 100 人类活动驱动植被覆盖减少 说明:θ(INDVpre)、θ(INDVobs)、θ(INDVres)分别为NDVI预测值、观测值、残差的变化斜率。θ>0表示植被受到驱动因素的正向促进作用,θ<0表示植被受到驱动因素的反向抑制作用。
下载: 导出CSV
表 3 2001—2020年延安市年与季节NDVI变化趋势统计
Table 3. Annual and seasonal NDVI trend statistics in Yan’an City from 2001 to 2020
NDVI变化趋势 β Z 所占面积比例/% 春季/% 夏季/% 秋季/% 冬季/% 显著上升 ≥0.000 5 ≥2.58 83.27 87.29 84.99 76.42 88.23 不显著上升 ≥0.000 5 1.96~2.58 8.59 7.70 8.01 9.13 6.74 稳定不变 −0.000 5~0.000 5 −1.96~1.96 7.97 4.89 6.86 14.33 5.00 不显著下降 <−0.000 5 −2.58~−1.96 0.09 0.08 0.08 0.07 0.02 显著下降 <−0.000 5 ≤−2.58 0.08 0.04 0.06 0.05 0.01 说明:β为NDVI变化斜率。
下载: 导出CSV
-
[1] 樊星, 秦圆圆, 高翔. IPCC第六次评估报告第一工作组报告主要结论解读及建议[J]. 环境保护, 2021, 49 (增刊2): 44 − 48. FAN Xing, QIN Yuanyuan, GAO Xiang. Interpretation of the main conclusions and suggestions of IPCC AR6 working group Ⅰ report [J]. Environmental Protection, 2021, 49 (suppl 2): 44 − 48. [2] 杜勇, 李建柱, 牛凯杰, 等. 1982—2015年永定河山区植被变化及对天然径流的影响[J]. 水利学报, 2021, 52(11): 1309 − 1323. DU Yong, LI Jianzhu, NIU Kaijie, et al. Analysis of vegetation change and its impact on natural runoff in the mountain area of the Yongding River Basin from 1982 to 2015 [J]. Journal of Hydraulic Engineering, 2021, 52(11): 1309 − 1323. [3] 田潇然, 王锦, 余哲修. 丽江坝区植被覆盖度时空变化驱动力研究[J]. 西北林学院学报, 2022, 37(3): 180 − 190. TIAN Xiaoran, WANG Jin, YU Zhexiu. Assessment of forces contributing to fractional vegetation cover spatio-temporal variation in Lijiang Dam area [J]. Journal of Northwest Forestry University, 2022, 37(3): 180 − 190. [4] 国志兴, 王宗明, 宋开山, 等. 1982—2003年东北林区森林植被NDVI与水热条件的相关分析[J]. 生态学杂志, 2007, 26(12): 1930 − 1936. GUO Zhixing, WANG Zongming, SONG Kaishan, et al. Correlations between forest vegetation NDVI and water/thermal condition in northeast China forest regions in 1982−2003 [J]. Chinese Journal of Ecology, 2007, 26(12): 1930 − 1936. [5] 丛楠, 张扬建, 朱军涛. 北半球中高纬度地区近30年植被春季物候温度敏感性[J]. 植物生态学报, 2022, 46(2): 125 − 135. CONG Nan, ZHANG Yangjian, ZHU Juntao. Temperature sensitivity of vegetation phenology in spring in mid- to high-latitude regions of northern Hemisphere during the recent three decades [J]. Chinese Journal of Plant Ecology, 2022, 46(2): 125 − 135. [6] 张云霞, 张云飞, 李晓兵. 地面测量与ASTER影像综合计算植被盖度[J]. 生态学报, 2007, 27(3): 964 − 976. ZHANG Yunxia, ZHANG Yunfei, LI Xiaobing. The synthetically estimating vegetation fractional coverage of grassland using field data and ASTER remote sensing imagine [J]. Acta Ecologica Sinica, 2007, 27(3): 964 − 976. [7] 田地, 刘政, 胡亚林. 福州市植被覆盖度时空特征及与地形因子的关系[J]. 浙江农林大学学报, 2019, 36(6): 1158 − 1165. TIAN Di, LIU Zheng, HU Yalin. Spatial-temporal characteristics of vegetation coverage and the relationship to topographic factors in Fuzhou City [J]. Journal of Zhejiang A&F University, 2019, 36(6): 1158 − 1165. [8] DING Yongxia, LI Zhi, PENG Shouzhang. Global analysis of time-lag and -accumulation effects of climate on vegetation growth [J/OL]. International Journal of Applied Earth Observation and Geoinformation, 2020, 92 : 102179[2023-12-01]. doi: 10.1016/j.jag.2020.102179. [9] 金凯. 中国植被覆盖时空变化及其与气候和人类活动的关系[D]. 杨凌: 西北农林科技大学, 2019. JIN Kai. Spatio-temporal Variations of Vegetation Cover and its Relationships between Climate Change and Human Activities over China [D]. Yangling: Northwest A&F University, 2019. [10] 李红英, 张存桂, 汪生珍, 等. 近40年青藏高原植被动态变化对水热条件的响应[J]. 生态学报, 2022, 42(12): 4770 − 4783. LI Hongying, ZHANG Cungui, WANG Shengzhen, et al. Response of vegetation dynamics to hydrothermal conditions on the Qinghai-Tibet Plateau in the last 40 years [J]. Acta Ecologica Sinica, 2022, 42(12): 4770 − 4783. [11] 逯金鑫, 周荣磊, 刘洋洋, 等. 黄土高原植被覆被时空动态及其影响因素[J]. 水土保持研究, 2023, 30(2): 211 − 219. LU Jinxin, ZHOU Ronglei, LIU Yangyang, et al. Temporal and spatial dynamics of vegetation cover on the Loess Plateau and its influencing factors [J]. Research of Soil and Water Conservation, 2023, 30(2): 211 − 219. [12] WANG Huaijun, LI Zhi, CAO Lei, et al. Response of NDVI of natural vegetation to climate changes and drought in China [J/OL]. Land, 2021, 10 (9): 966[2023-12-01]. doi: 10.3390/land1009096. [13] 涂又, 姜亮亮, 刘睿, 等. 1982—2015年中国植被NDVI时空变化特征及其驱动分析[J]. 农业工程学报, 2021, 37(22): 75 − 84. TU You, JIANG Liangliang, LIU Rui, et al. Spatiotemporal changes of vegetation NDVI and its driving forces in China during 1982−2015 [J]. Transactions of the Chinese Society of Agricultural Engineering, 2021, 37(22): 75 − 84. [14] 祁鹏卫, 张贤. 2000—2019年重庆市植被覆盖时空变化特征及其驱动因素[J]. 生态学报, 2022, 42(13): 5427 − 5436. QI Pengwei, ZHANG Xian. Spatiotemporal variation characteristics and driving factors of vegetation cover in Chongqing from 2000 to 2019 [J]. Acta Ecologica Sinica, 2022, 42(13): 5427 − 5436. [15] 陈晨, 王义民, 黎云云, 等. 黄河流域1982—2015年不同气候区植被时空变化特征及其影响因素[J]. 长江科学院院报, 2022, 39(2): 56 − 62, 81. CHEN Chen, WANG Yimin, LI Yunyun, et al. Vegetation changes and influencing factors in different climatic regions of Yellow River basin from 1982 to 2015 [J]. Journal of Yangtze River Scientific Research Institute, 2022, 39(2): 56 − 62, 81. [16] 张小勇, 卫海燕. 延安市1999-2008年植被恢复的成效分析[J]. 水土保持通报, 2013, 33(2): 218 − 220, 225. ZHANG Xiaoyong, WEI Haiyan. Vegetation restoration effects of implementation of return grain to green program in Yan’an City during 1999−2008 [J]. Bulletin of Soil and Water Conservation, 2013, 33(2): 218 − 220, 225. [17] 赵文武, 吕一河, 郭雯雯, 等. 陕北黄土丘陵沟壑区NDVI与气象因子的相关性分析[J]. 水土保持研究, 2006, 13(2): 112 − 114. ZHAO Wenwu, LÜ Yihe, GUO Wenwen, et al. The correlations between climatic factors and NDVI in hilly and gully area of Loess Plateau in northern Shaanxi [J]. Research of Soil and Water Conservation, 2006, 13(2): 112 − 114. [18] 聂桐, 董国涛, 蒋晓辉, 等. 延安地区植被覆盖度时空变化及驱动力[J]. 水土保持研究, 2021, 28(5): 340 − 346. NIE Tong, DONG Guotao, JIANG Xiaohui, et al. Spatial and temporal variation of vegetation cover in Yan’an City and its driving forces [J]. Research of Soil and Water Conservation, 2021, 28(5): 340 − 346. [19] 张家政, 闵志强, 王得军, 等. 基于延安市不同植被类型NDVI对气象因子影响及空间异质性[J]. 西北林学院学报, 2021, 36(6): 55 − 64. ZHANG Jiazheng, MIN Zhiqiang, WANG Dejun, et al. Effects of NDVI on meteorological factors and spatial heterogeneity based on different vegetation types in Yan’an City [J]. Journal of Northwest Forestry University, 2021, 36(6): 55 − 64. [20] JIANG Weiguo, YUAN Lihua, WANG Wenjie, et al. Spatio-temporal analysis of vegetation variation in the Yellow River Basin [J]. Ecological Indicators, 2015, 51: 117 − 126. [21] SEN P K. Estimates of the regression coefficient based on Kendall’s tau [J]. Journal of the American Statistical Association, 1968, 63(324): 1379 − 1389. [22] FENSHOLT R, LANGANKE T, RASMUSSEN K, et al. Greenness in semi-arid areas across the globe 1981−2007 an earth observing satellite based analysis of trends and drivers [J]. Remote Sensing of Environment, 2012, 121: 144 − 158. [23] 梁冰洁. 气候变化背景下的北洛河流域植被生态系统时空格局演变分析[D]. 郑州: 郑州大学, 2017. LIANG Bingjie. Analysis of Spatio-temporal Variation of Vegetation Ecosystem in Beiluo River Basin in the Context of Climate Change [D]. Zhengzhou: Zhengzhou University, 2017. [24] QI Xiangzhen, JIA Junhe, LIU Huiyu, et al. Relative importance of climate change and human activities for vegetation changes on China’s silk road economic belt over multiple timescales [J]. Catena, 2019, 180: 224 − 237. [25] 李依璇, 朱清科, 石若莹, 等. 2000—2018年黄土高原植被覆盖时空变化及影响因素[J]. 中国水土保持科学, 2021, 19(4): 60 − 68. LI Yixuan, ZHU Qingke, SHI Ruoying, et al. Spatial and temporal changes of vegetation cover and its influencing factors in the Loess Plateau from 2000 to 2018 [J]. Science of Soil and Water Conservation, 2021, 19(4): 60 − 68. [26] 刘旻霞, 赵瑞东, 邵鹏, 等. 近15 a黄土高原植被覆盖时空变化及驱动力分析[J]. 干旱区地理, 2018, 41(1): 99 − 108. LIU Minxia, ZHAO Ruidong, SHAO Peng, et al. Temporal and spatial variation of vegetation coverage and its driving forces in the Loess Plateau from 2001 to 2015 [J]. Arid Land Geography, 2018, 41(1): 99 − 108. [27] 卢乔倩, 江涛, 柳丹丽, 等. 中国不同植被覆盖类型NDVI对气温和降水的响应特征[J]. 生态环境学报, 2020, 29(1): 23 − 34. LU Qiaoqian, JIANG Tao, LIU Danli, et al. The response characteristics of NDVI with different vegetation cover types to temperature and precipitation in China [J]. Ecology and Environmental Sciences, 2020, 29(1): 23 − 34. [28] 张乐艺, 李霞, 冯京辉, 等. 2000-2018年黄河流域NDVI时空变化及其对气候和人类活动的双重响应[J]. 水土保持通报, 2021, 41(5): 276 − 286. ZHANG Leyi, LI Xia, FENG Jinghui, et al. Spatial-temporal changes of NDVI in Yellow River basin and its dual response to climate change and human activities during 2000−2018 [J]. Bulletin of Soil and Water Conservation, 2021, 41(5): 276 − 286. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230610
 点击查看大图
点击查看大图
计量
- 文章访问数: 1726
- HTML全文浏览量: 478
- PDF下载量: 26
- 被引次数: 0
