





-
铁路、公路等基础设施建设会破坏和占压地表植被,形成大量的裸露坡面,遇到降雨极易发生水土流失,甚至出现滑坡、泥石流等次生地质灾害。裸露坡面常常具有坡度陡、坡体稳定性低、水分条件差和土壤瘠薄等特征,是不利于植被生长的困难立地。客土喷播绿化是裸露坡面恢复植被最快速最有效的方式之一,喷播后灌溉养护对植被生长至关重要[1]。大量调查发现:客土喷播后普遍存在过度灌溉,产生坡面径流,造成水土流失和水资源浪费;同时喷播基质通气不畅也会影响植被生长。可见,确定适合植被生长且能保证灌溉时坡面不产流的客土喷播基质含水量已成为当前亟需解决的问题。目前,关于适宜含水量研究大多集中在林地土壤与林木之间,如夏江宝等[2]对贝壳堤岛旱柳Salix matsudana光合效率的土壤水分临界效应及其阈值进行了分级研究,景雄等[3]对毛竹Phyllostachys edulis实生苗土壤水分有效性及生产力进行了分级研究,张淑勇等[4]对黄刺玫Rosa xanthina叶片光合生理参数的土壤水分阈值响应及其生产力进行了分级研究等,客土喷播基质适宜含水量与植被生长的关系研究则较少。以往的研究大都只关注了植物某一个生长阶段的土壤水分适宜含水量阈值[2, 5-6],缺乏对不同季节植被生长与基质水分关系的研究。鉴于此,本研究以北方地区常用的喷播修复植物黑麦草Lolium perenne作为研究对象,利用种植盆模拟客土喷播绿化,通过控制不同客土喷播基质水分梯度,分析夏、秋季黑麦草光合特性日变化对不同喷播基质水分的响应规律,以叶片净光合速率(Pn)和水分利用效率(EWU)作为“产”“效”来评价黑麦草生产力和水分利用能力的依据,并进行季节间比较,建立夏、秋季黑麦草喷播基质适宜含水量阈值分级,以期为北京至张家口的公路、铁路等冬季奥林匹克运动会交通廊道以及自然条件相近地区的工程创面客土喷播恢复植被灌溉养护提供参考。
-
研究区河北省张家口市涿鹿县为北京冬季奥林匹克运动会延庆赛区和张家口崇礼赛区廊道沿线,高速公路G6和G7之间,地理坐标为40°26′20″N,115°17′03″E。涿鹿县属温带半干旱大陆性季风气候,年均气温为9.1 ℃,极端最高气温为39.2 ℃,极端最低气温为−23.9 ℃,年均降水量为367 mm,年均蒸发量为1 600 mm,无霜期为169 d,年平均积温为2 100~3 400 ℃,风向以西北为主,平均风速2~3 m·s−1,土壤为沙壤质褐土。
-
喷播基质材料为客土(取自河北省涿鹿县苗圃)、木纤维[长1~3 cm,中矿复地生态环境技术研究院(北京)有限公司]、保水剂(3005KCE,美国艾森公司)、黏合剂(A30,美国艾森公司)、稻壳和黑麦草种子(北京布莱特草业有限公司)。喷播基质层和种子层的材料配比见表1。黑麦草播种量为4 g·m−2。
表 1 基质层和种子层的材料配比
Table 1. Material ratio of matrix layer and seed layer
喷播层次 客土/
%木纤
维/%稻壳/
%复合肥/
(g·m−3)保水剂/
(g·m−3)黏合剂/
(g·m−3)基质层(10 cm) 70 10 20 300 200 150 种子层(3 cm) 67 33 说明:客土、木纤维和稻壳为体积比 -
利用种植盆试验模拟客土喷播绿化,种植盆上口直径50 cm、盆底直径40 cm、高15 cm,底部打孔便于排水。使用恒睿牌HKP125型客土喷播机。2021年4月26日,根据表1的材料配比将基质层和种子层分上、下2层先后喷播到种植盆内,采用微喷灌雾化喷头对喷播基质灌溉养护,保持喷播基质充分湿润(每次灌溉以喷播基质表面不积水为准),保证种子出苗有充足的水分。
2021年5月26日开始控制喷播基质含水量(为质量含水量,下同),用环刀法测得喷播基质的田间持水量为30.36%,容重为1.12 g·cm−3。喷播基质含水量设置5个水分梯度,分别为30.36%、25.81%、21.25%、16.70%和12.14%,即喷播基质相对含水量(CRW)为100%、85%、70%、55%和40%,每个水分梯度设置3个重复。每天16:00用TDR350土壤水分速测仪(美国Spectrum公司)测定CRW(通过容重换算为质量含水量),每盆测定重复3次取平均值,并根据公式计算耗水量:w设−w测=m耗/m干。其中:w设为设计质量含水量(%);w测为实测质量含水量(%),根据TDR350实测值和容重换算;m耗为每盆黑麦草耗水量(g);m干为每盆喷播基质干质量(g),可由基质体积和容重计算得出。使用微喷灌雾化喷头对喷播基质补充水分,为避免降水影响,试验在透明通风遮雨大棚内进行。
-
于夏季(2021年8月5日,即控水2个月后)、秋季(2021年10月11日,即控水4个月后)选择连续3 d晴朗无云的天气,使用Li-6400XT便携式光合作用测定仪(标准叶室,Li-COR)测定黑麦草叶片Pn (μmol·m−2·s−1)、蒸腾速率Tr (mmol·m−2·s−1)、气孔导度Gs (mol·m−2·s−1)、胞间二氧化碳(CO2)摩尔浓度Ci (μmol·mol−1)等生理参数以及大气CO2摩尔浓度Ca (μmol·mol−1)、光合有效辐射PAR (μmol·m−2·s−1)、气温Ta (℃)和相对湿度Rh (%)等环境因子,并根据公式EWU=Pn/Tr计算水分利用效率、Ls=1−Ci/Ca计算气孔限制值。测定时间为8:00—16:00,隔2 h测1次,每个种植盆选取3株生长健康、长势一致的黑麦草,每株选取3片叶,每片叶记录3次读数,取平均值。
-
运用Excel 2016整理光合参数与基质相对含水量数据;SPSS 22.0进行差异显著性检验LSD;Origin 2018进行作图和多项式拟合建立回归模型,使用F检验对回归模型进行显著性检验。
-
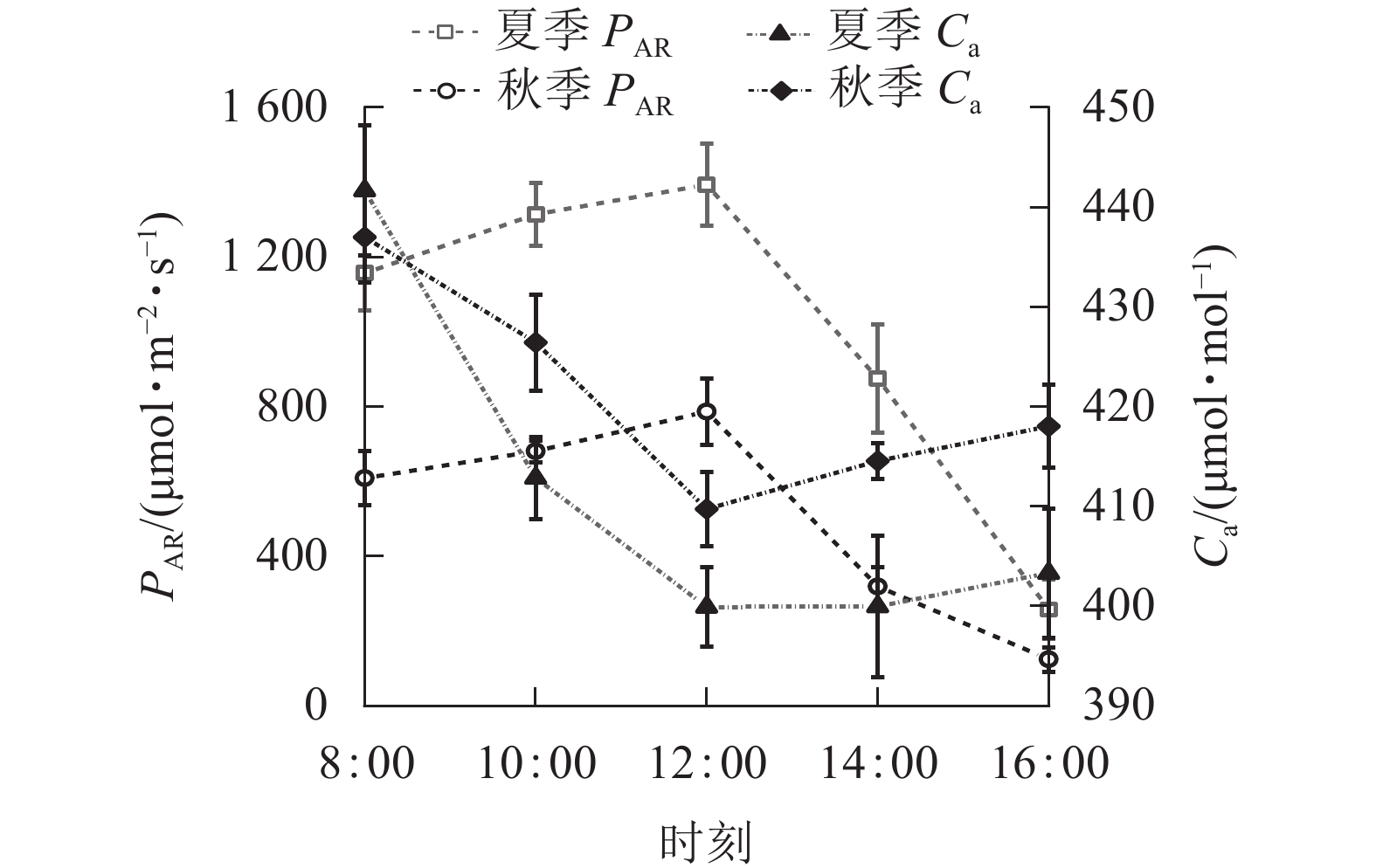
由图1可知:夏、秋季PAR的日变化为单峰曲线,均为先升高后下降,峰值均出现在12:00,夏季峰值为(1 393.71±110.04) μmol·m−2·s−1,秋季为(786.73±88.74) μmol·m−2·s−1。夏季PAR日均值(999.75±459.61) μmol·m−2·s−1大于秋季(504.07±274.09) μmol·m−2·s−1。夏、秋季Ca日变化为“V”型曲线,8:00—12:00下降,之后上升。秋季Ca日均值(421.15±17.65) μmol·mol−1大于夏季(411.54 ±10.76) μmol·mol−1,两者相差较小,仅为2.30%。
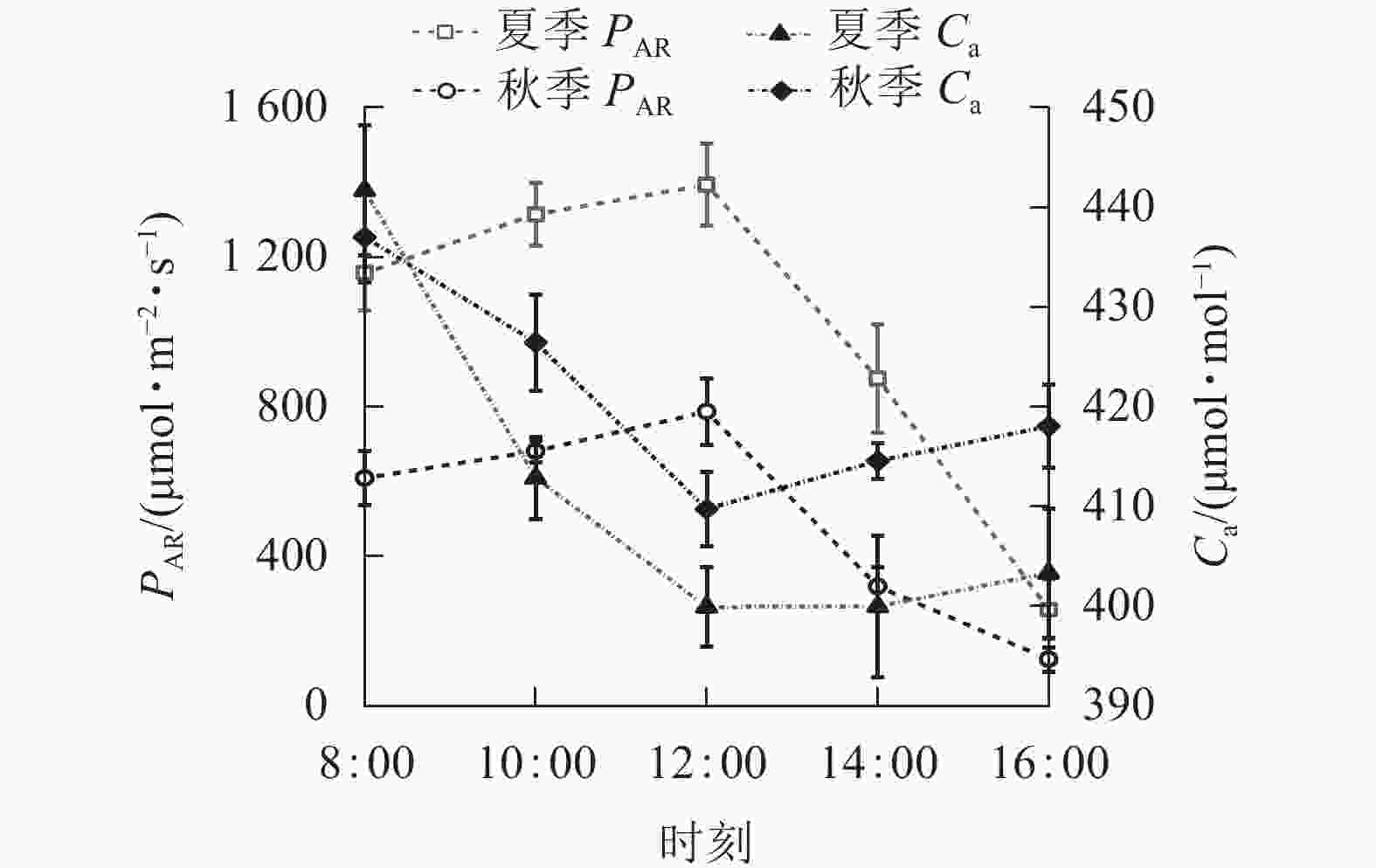
图 1 夏、秋季光合有效辐射(PAR)和大气CO2摩尔浓度(Ca)的日变化
Figure 1. Diurnal variation of photosynthetically active radiation (PAR) and atmospheric CO2 concentration (Ca) in summer and autumn
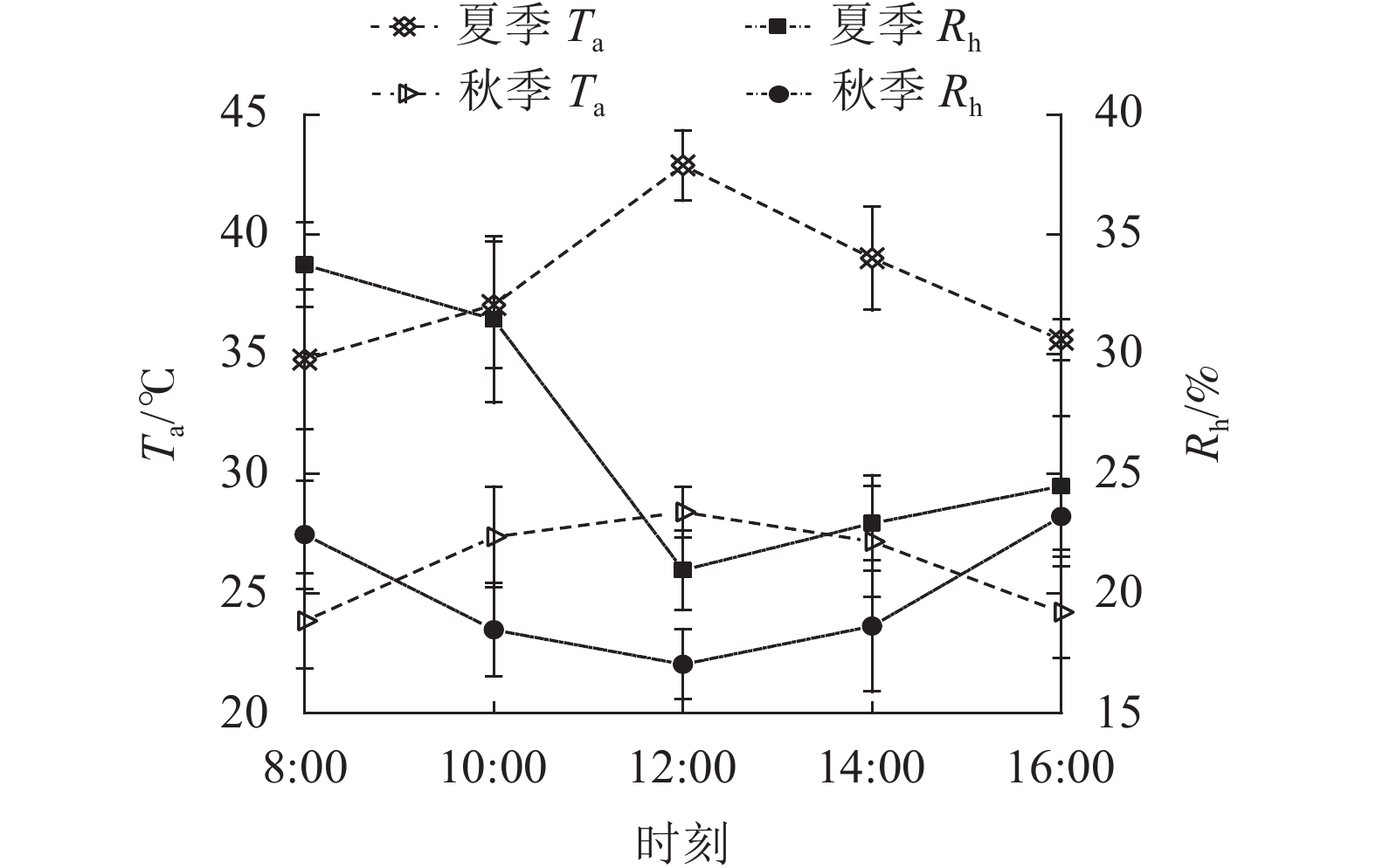
由图2可知:夏、秋季Ta的日变化与PAR相似,也为单峰曲线,在12:00达最大值。夏季Ta最大为(42.88±1.46) ℃,秋季为(28.41±1.06) ℃。夏季日均值(37.87±3.23) ℃大于秋季(26.21±2.03) ℃。夏、秋季Rh的日变化与Ta相反,12:00前下降,之后上升,夏、秋季Rh最低值分别为20.98%±1.65%和17.05%±1.47%。夏季Rh日均值(26.72%±5.56%)大于秋季(19.98%±2.70%)。
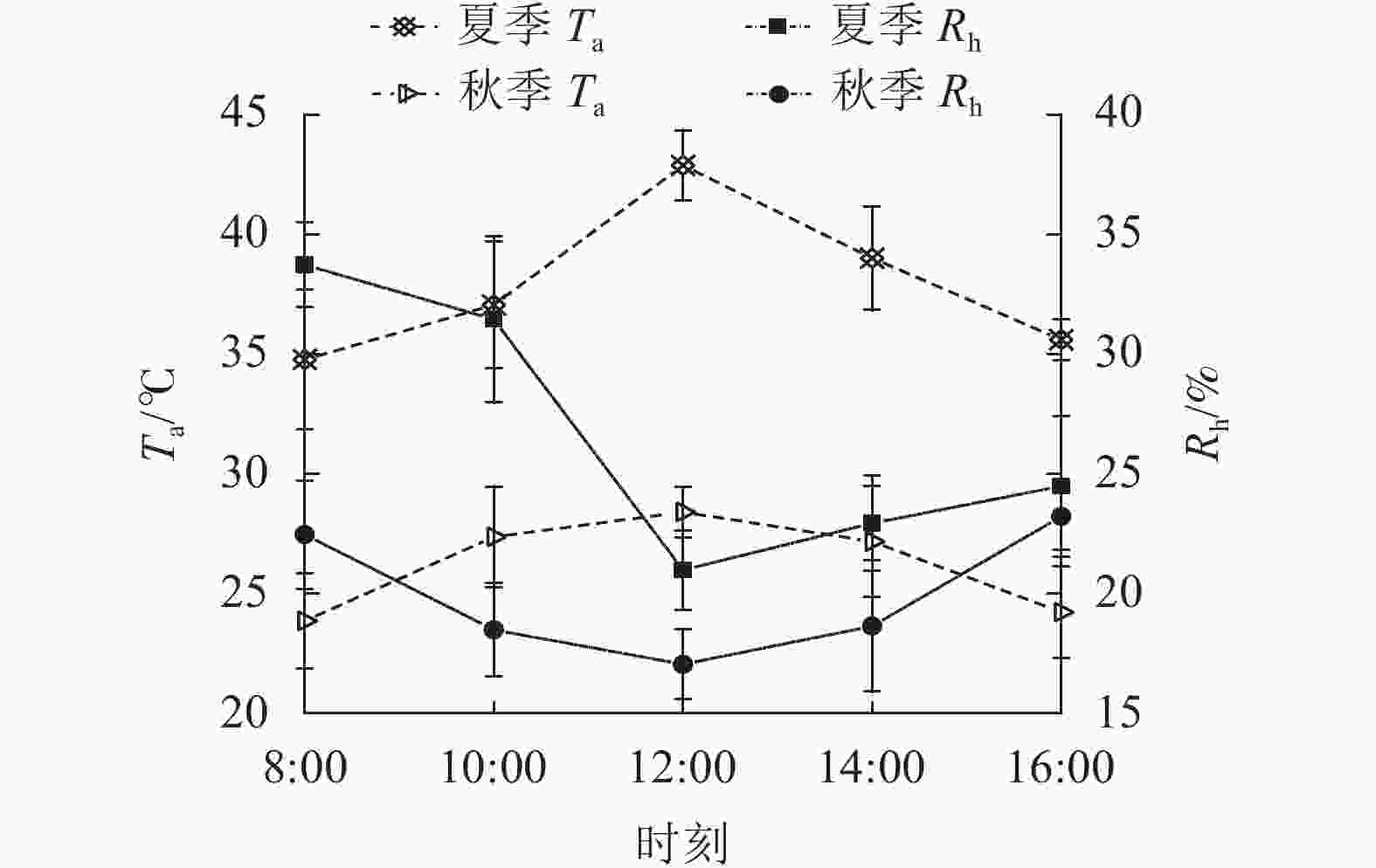
图 2 夏、秋季气温(Ta)和相对湿度(Rh)的日变化的日变化
Figure 2. Diurnal variation of temperature (Ta) and relative humidity (Rh) in summer and autumn
-
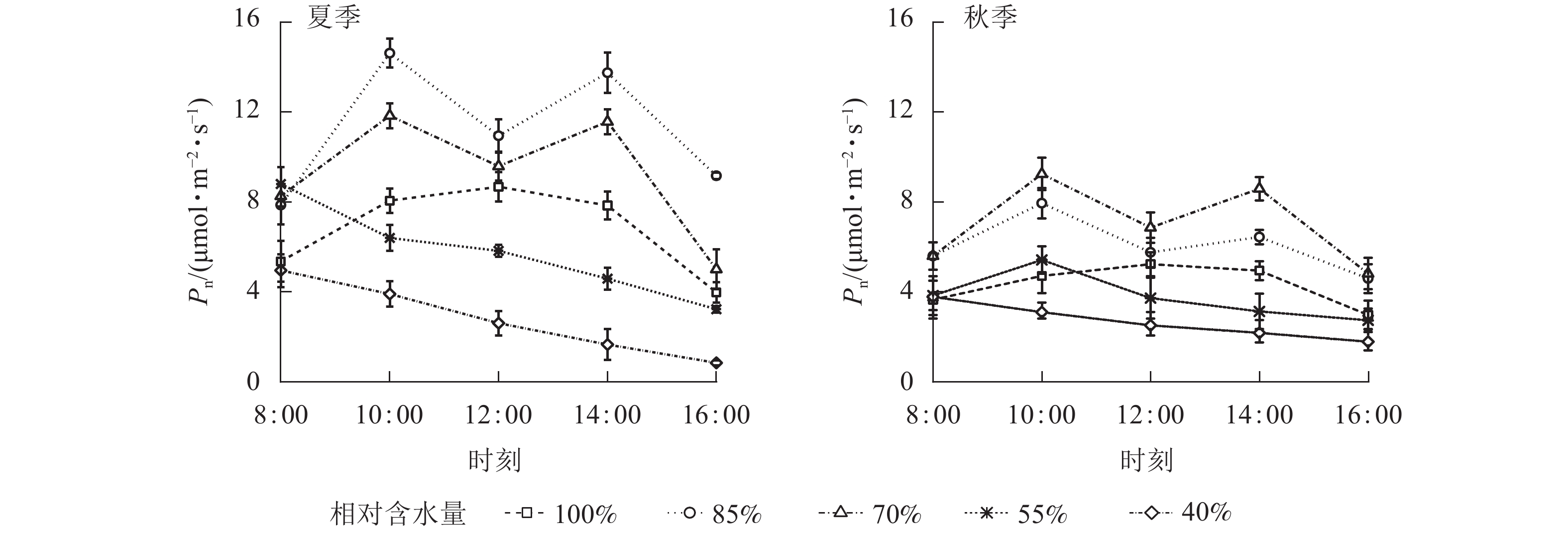
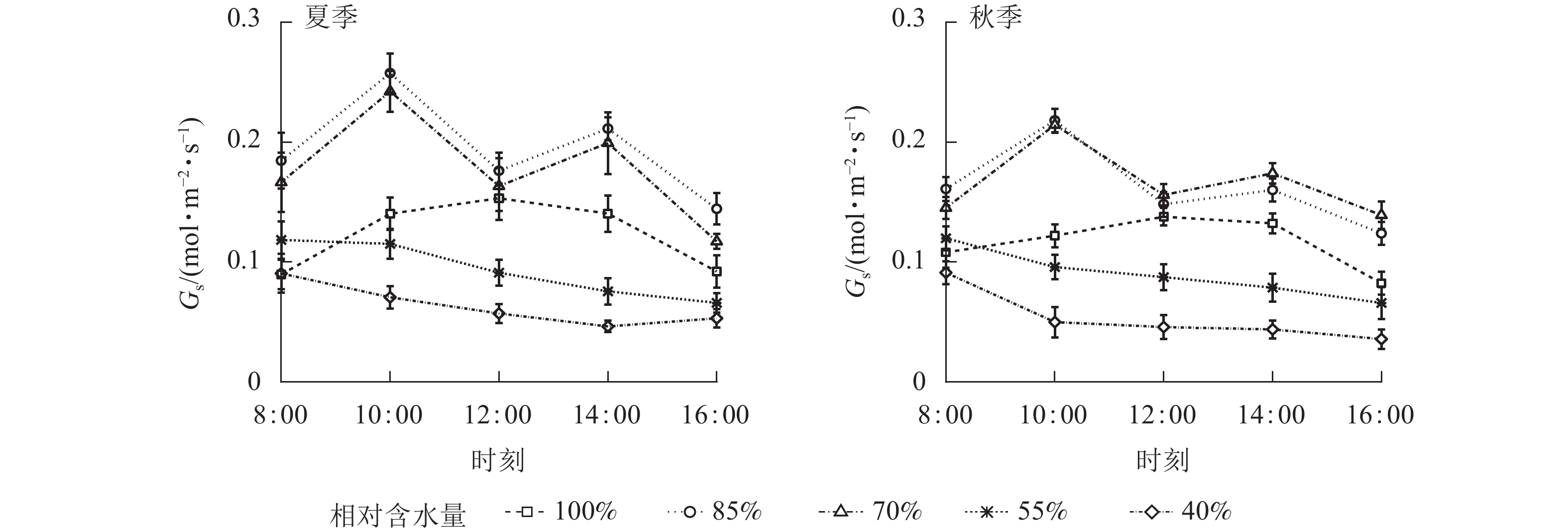
夏、秋季黑麦草叶片Pn日变化对CRW有明显的阈值响应(图3)。当CRW为70%~85%时,Pn的变化呈双峰曲线,均出现光合“午休”现象,上午和下午各出现1个峰值,此水分范围内,Pn在全天各时段均最高。当CRW增加至100%时,Pn呈单峰曲线,峰值出现在12:00。当CRW降低到55%和40%时,Pn为单峰曲线,峰值均出现在8:00(但秋季CRW为55%时Pn峰值出现在10:00),Pn在全天各时段均处于较低水平,表明CRW低于55%会严重抑制植物的光合作用。由表2可知:Pn日均值对CRW也有明显的阈值响应。当CRW为85%时,夏季Pn日均值最大,达(11.17±3.08) μmol·m−2·s−1,与其他水分梯度有显著差异(P<0.05)。秋季的Pn日均值在CRW为70%时达最大,为(7.02±1.97) μmol·m−2·s−1,与其他水分梯度也有显著差异(P<0.05)。夏季Pn日均值均大于秋季,CRW为55%~100%时两季差异达到显著(P<0.05)。CRW为40%时,两季Pn日均值均较低,可见当CRW较低时植物光合作用将受到严重影响。综上所述,夏、秋两季维持黑麦草较高Pn的CRW为70%~85%,高于或低于此范围,Pn明显受到抑制。

图 3 夏、秋季不同喷播基质含水量下黑麦草净光合速率(Pn)的日变化
Figure 3. Diurnal variation of net photosynthetic rate (Pn) of L. perenne under different spraying substrate water content in summer and autumn
表 2 夏、秋季不同喷播基质含水量下黑麦草光合生理参数的日均值变化
Table 2. Change of daily mean of photosynthetic physiological parameters of L. perenne under different spraying substrate water content in summer and autumn
CRW/% Pn/(μmol·m−2·s−1) Tr/(mmol·m−2·s−1) EWU/(mol·mol−1) 夏季 秋季 夏季 秋季 夏季 秋季 100 6.79±2.01 Abc 4.30±0.95 Bb 5.59±1.17 Aab 2.75±0.16 Bab 1.32±0.20 Bbc 1.56±0.26 Acd 85 11.17±3.08 Aa 6.07±1.24 Ba 6.83±1.12 Aa 3.13±0.40 Ba 1.61±0.22 Ba 1.92±0.22 Ab 70 9.26±2.79 Aab 7.02±1.97 Ba 6.76±0.63 Aa 2.92±0.59 Ba 1.43±0.15 Bab 2.37±0.25 Aa 55 5.77±2.09 Ac 3.77±1.03 Bb 4.91±0.93 Ab 2.35±0.30 Bbc 1.20±0.12 Bbc 1.63±0.20 Ac 40 2.80±1.66 Ae 2.74±0.78 Ab 3.03±0.87 Ac 2.16±0.28 Ac 1.01±0.28 Ac 1.28±0.16 Ad 说明:同列不同小写字母、同行不同大写字母均表示差异显著(P<0.05) -
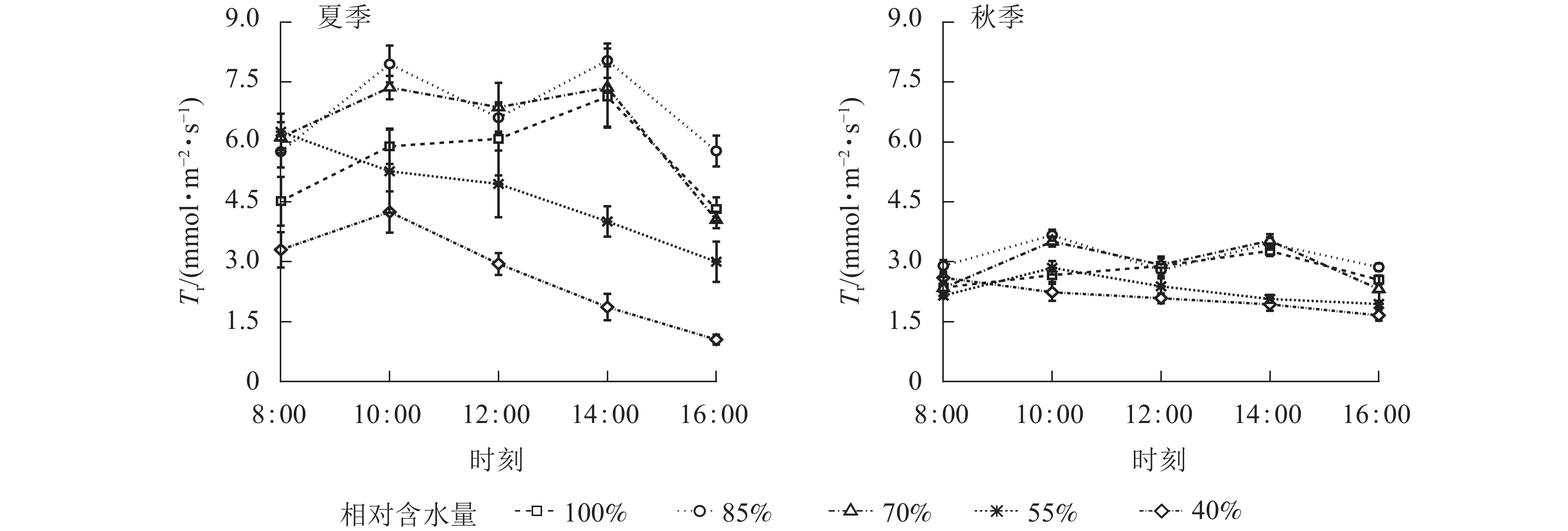
夏、秋季黑麦草叶片Tr日变化规律与Pn基本相似(图4),当CRW为70%~85%时,黑麦草Tr的日变化呈双峰曲线。当CRW增加至100%时,Tr呈现单峰曲线,夏、秋季峰值均出现在14:00,但日均值却低于CRW为70%~85%时。表明基质水分充足可有效延缓Tr“午休”,但会降低Tr。当CRW≤55%时,Tr呈单峰曲线,峰值出现在8:00或10:00,全天各时段均处于较低的水平。结合表2可知:当CRW≥55%时,夏季Tr日均值显著高于秋季(P<0.05)),可见不同季节气候环境对植物Tr影响较大。当CRW为40%时,Tr日均值显著低于其他水分梯度(P<0.05),表明水分胁迫严重限制Tr。综上所述,CRW过高或过低均会降低黑麦草Tr,当CRW为70%~85%时,黑麦草会保持较高的Tr,保障植物正常生理活动。
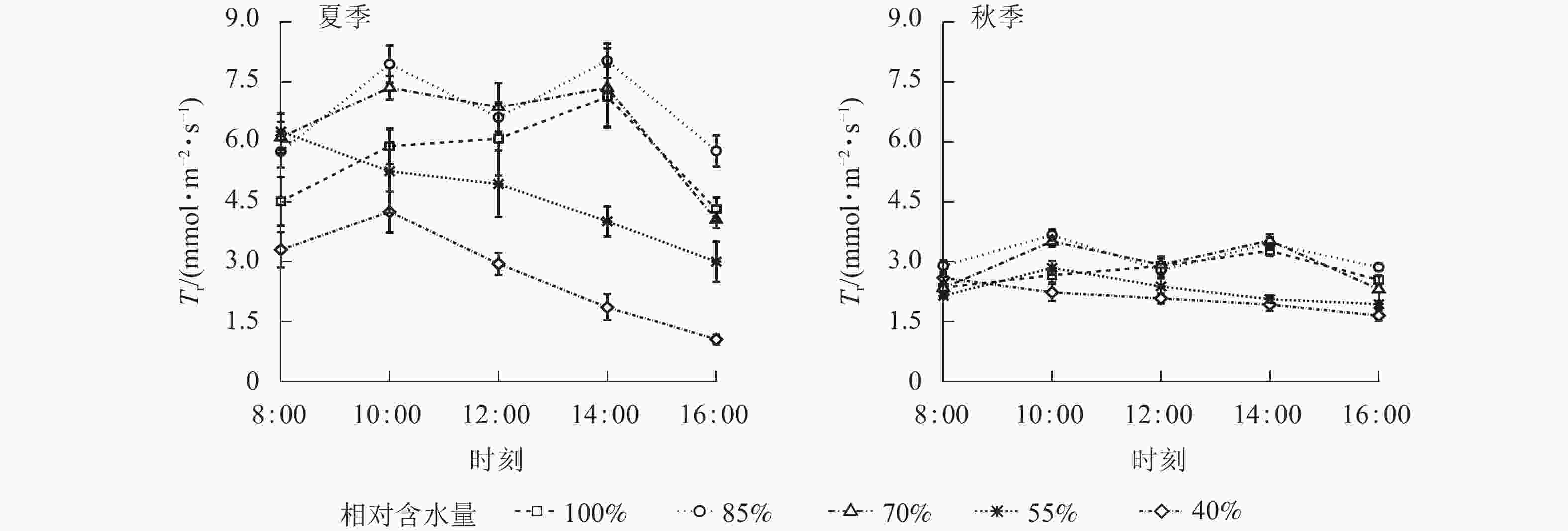
图 4 夏、秋季不同喷播基质含水量下黑麦草蒸腾速率(Tr)的日变化
Figure 4. Diurnal variation of transpiration rate (Tr) of L. perenne under different spraying substrate water content in summer and autumn
-
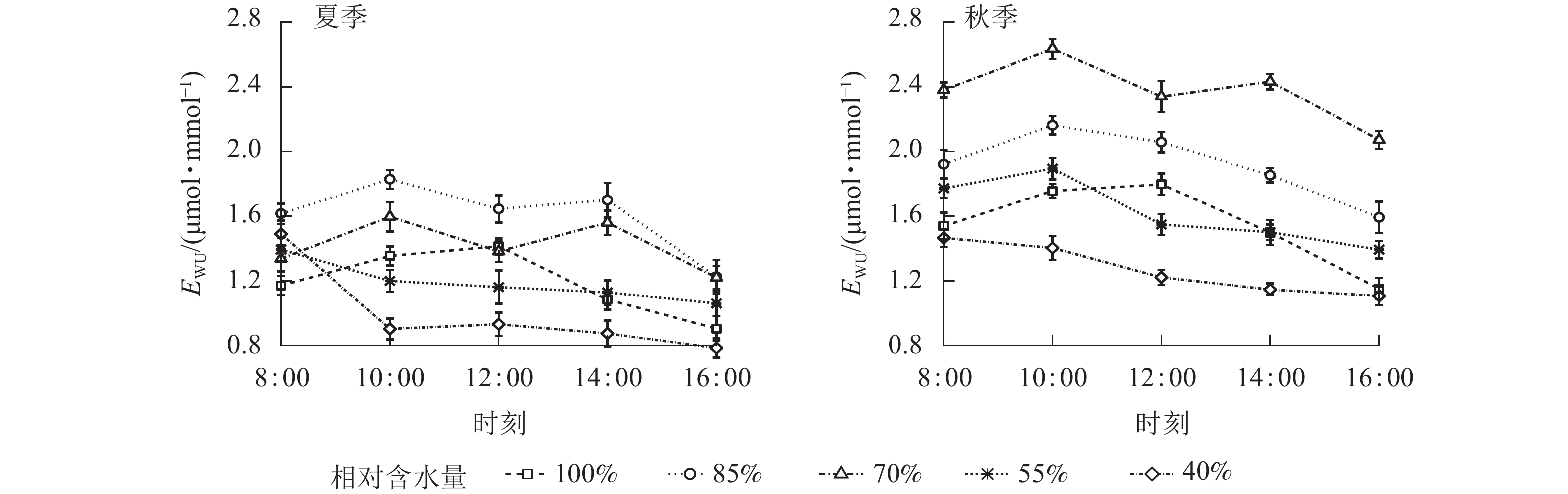
EWU日变化对基质含水量有明显的阈值响应(图5)。CRW为70%~85%时,EWU为双峰曲线(但秋季CRW=85%时为单峰曲线),全天各时段EWU均高于其他水分梯度。当CRW增加至100%时,EWU表现为单峰曲线,峰值出现在12:00。CRW为40%~55%时,EWU峰值出现在8:00或10:00,之后不断降低。结合表2可知:当CRW≥55%时,秋季EWU日均值显著高于夏季(P<0.05),CRW为40%时秋季EWU日均值高于夏季,但不差异显著。CRW为40%和100%时,EWU日均值均显著低于其他水分梯度(P<0.05),表明CRW过高或过低都会降低EWU。综上所述,夏、秋季维持黑麦草同时具有较高Pn和EWU的CRW为70%~85%,在这个水分范围内,Tr也保持较高水平,有利于植物的光合作用。

图 5 夏、秋季不同喷播基质含水量下黑麦草水分利用效率(EWU)的日变化
Figure 5. Diurnal variation of water use efficiency (EWU) of L. perenne under different spraying substrate water content in summer and autumn
-
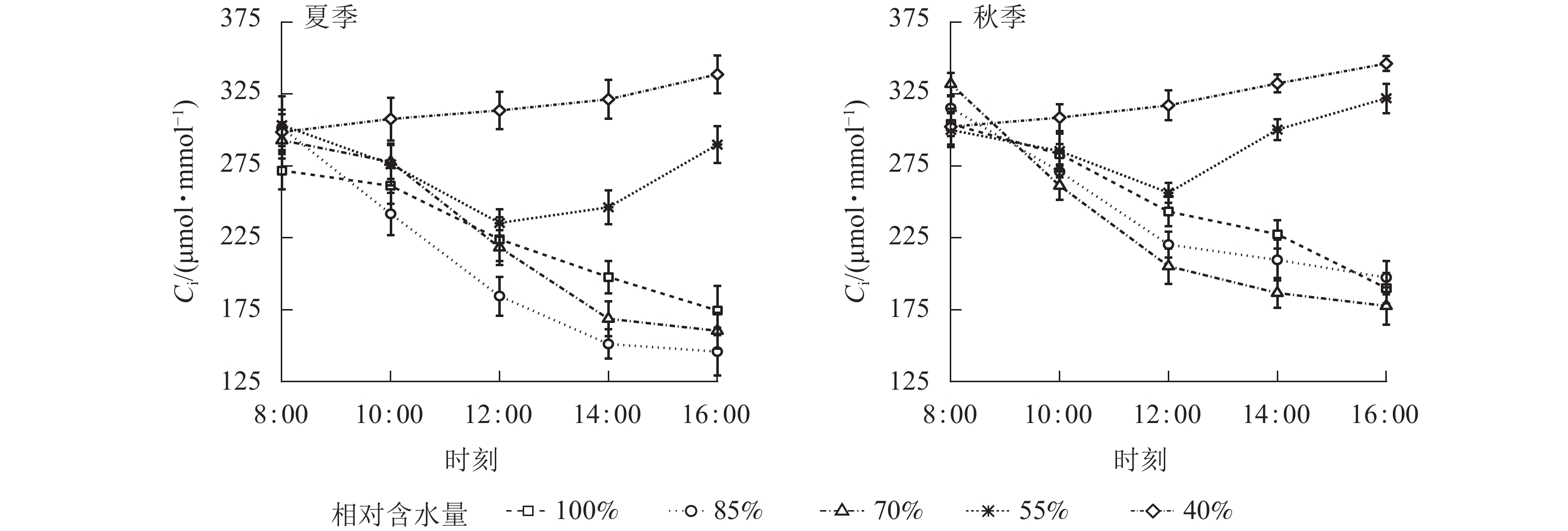
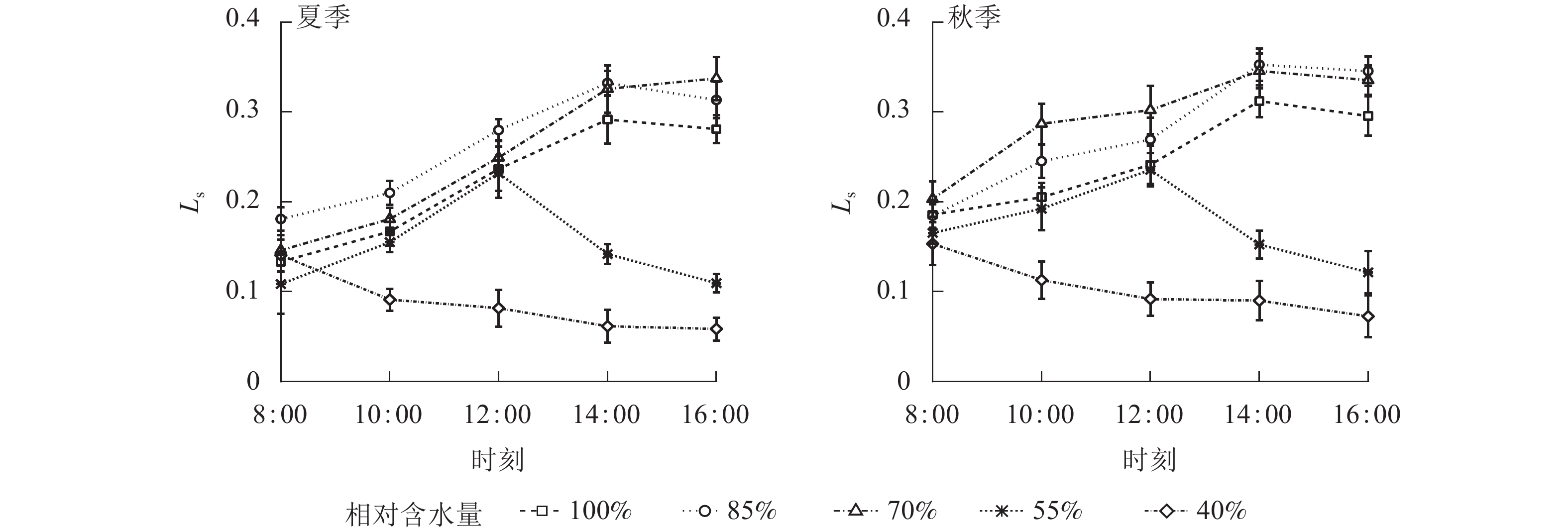
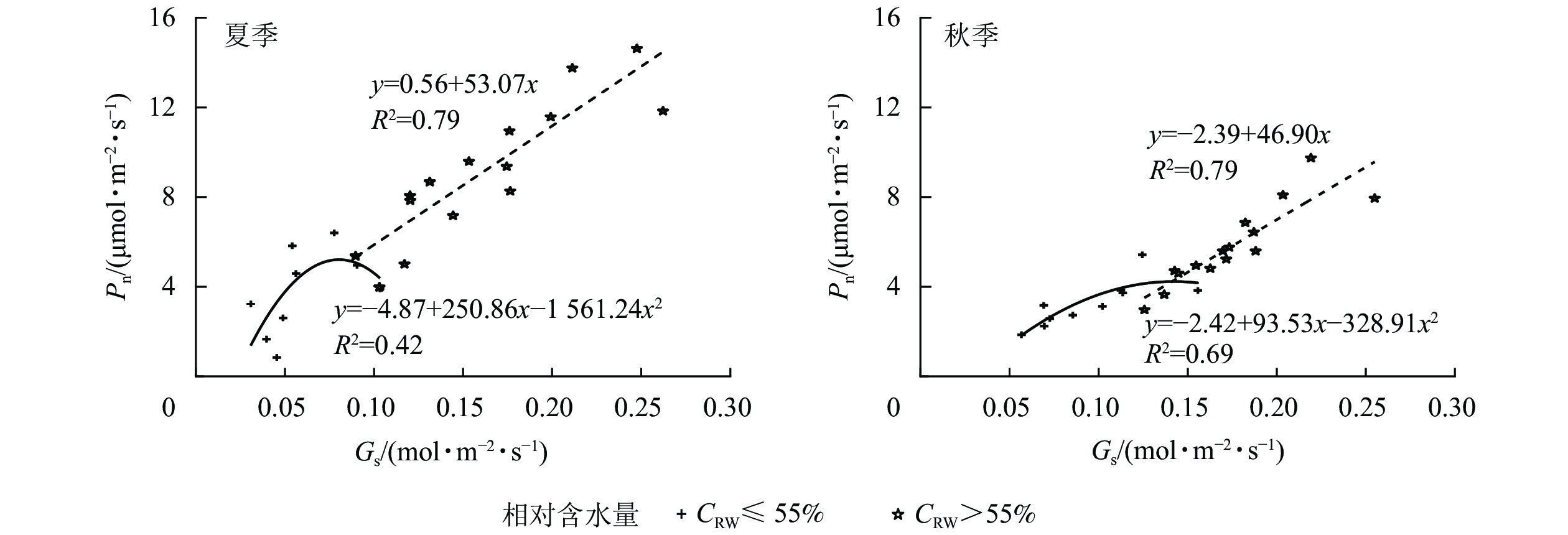
夏、秋季黑麦草Gs对CRW具有明显的阈值响应(图6),当CRW为70%~85%时,Gs呈现双峰曲线。当CRW=100%时,Gs为单峰曲线,峰值出现在12:00。当CRW为40%~55%时,全天Gs峰值出现在8:00,之后一直降低,维持在较低水平。Ci和Ls对CRW的阈值响应表现不同的变化规律(图7和图8),上午和下午表现也不同。CRW为70%~100%时,Pn下降,Gs和Ci明显下降,Ls明显升高,表明Pn下降原因是气孔限制。CRW=55%时,上午Pn下降,Gs和Ci明显下降,Ls升高,但下午Pn下降,Gs和Ls下降,Ci反而升高,可见限制黑麦草Pn的原因上午和下午不同,上午以气孔限制为主,气孔关闭导致CO2供应不足,下午以非气孔限制为主,水分胁迫导致植物叶片光合结构受损,Pn下降。当CRW=40%时,Ci从8:00开始上升且一直处于较高水平,而Ls全天都较低,表明水分胁迫严重损坏了植物叶片光合结构,降低了光合作用有关酶的活性,从而降低了Pn。由图9可知:夏、秋季不同CRW范围内Pn和Gs的正比关系不同,当CRW>55%时,随着Gs增大,Pn线性增大,Pn和Gs为线性正比关系;当CRW≤55%时,Pn和Gs为非线性关系。因此,当CRW=55%时,黑麦草不仅发生了Pn限制机制的转变,其Pn和Gs之间的关系也发生转变。综上所述,在CRW=55%时出现上午、下午Ci和Ls变化相反的情况,表明此基质含水量是黑麦草叶片Pn下降由气孔限制为主转变为非气孔限制为主的临界点。
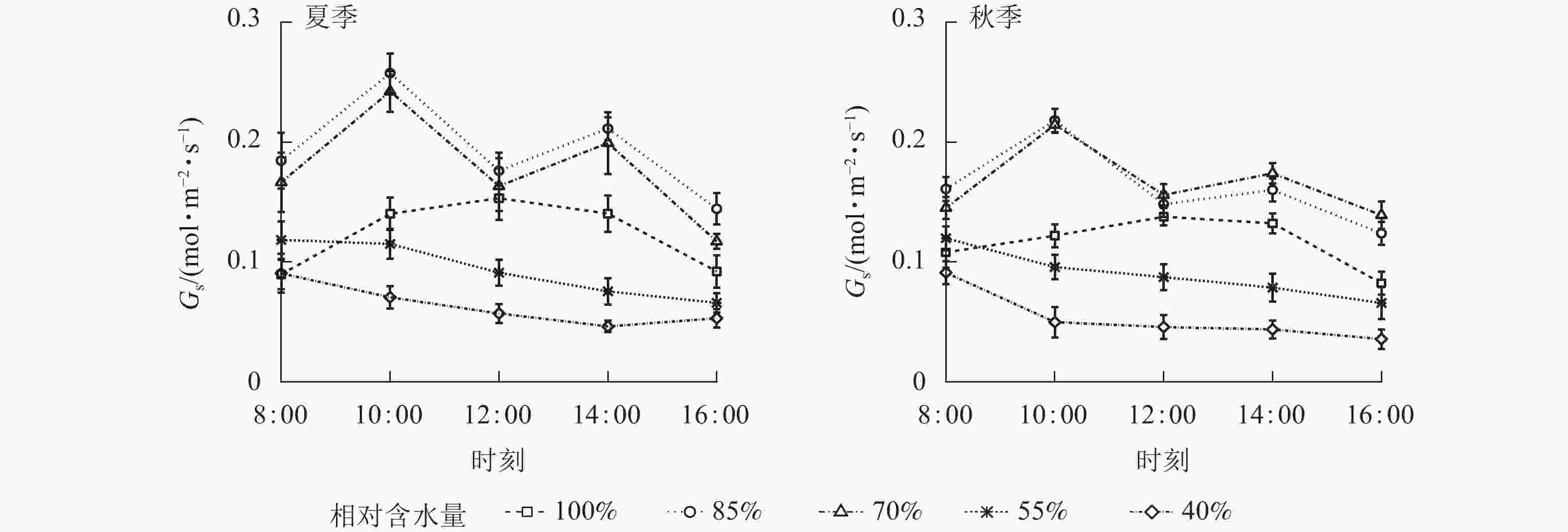
图 6 夏、秋季不同喷播基质含水量下黑麦草气孔导度(Gs)的日变化
Figure 6. Diurnal variation of stomatal conductance (Gs) of L. perenne under different water content of spraying substrate in summer and autumn
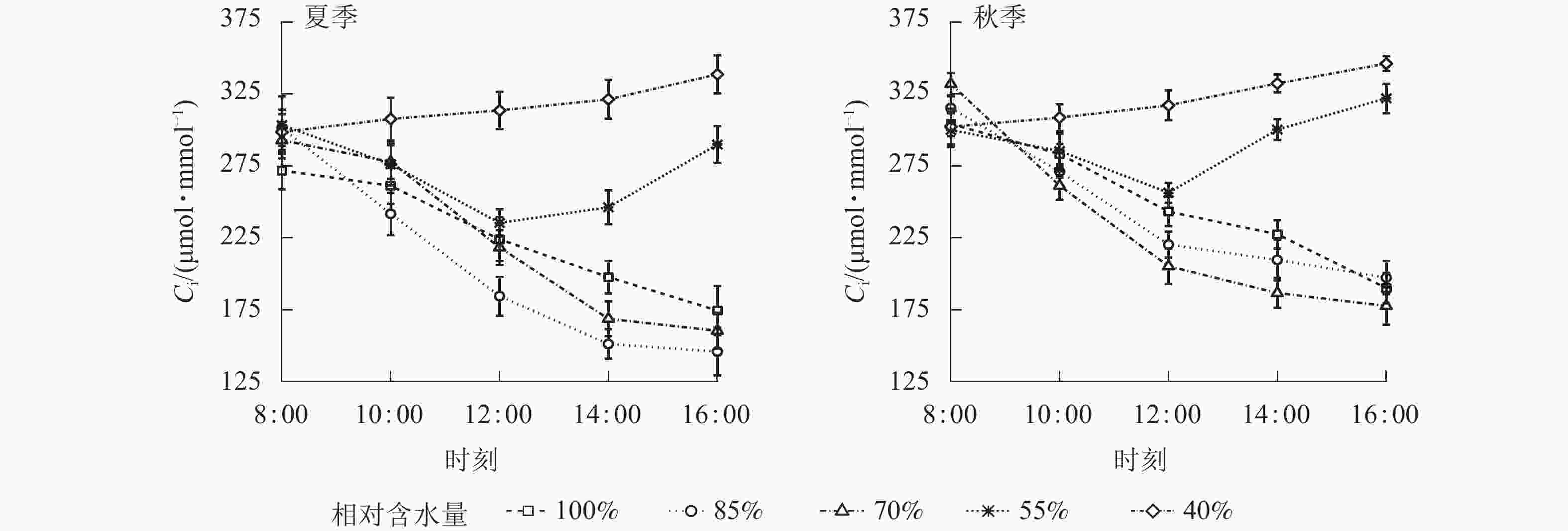
图 7 夏、秋季不同喷播基质含水量下黑麦草胞间CO2摩尔浓度(Ci)的日变化
Figure 7. Diurnal variation of intercellular CO2 concentration (Ci) of L. perenne under different water content of spraying substrate in summer and autumn
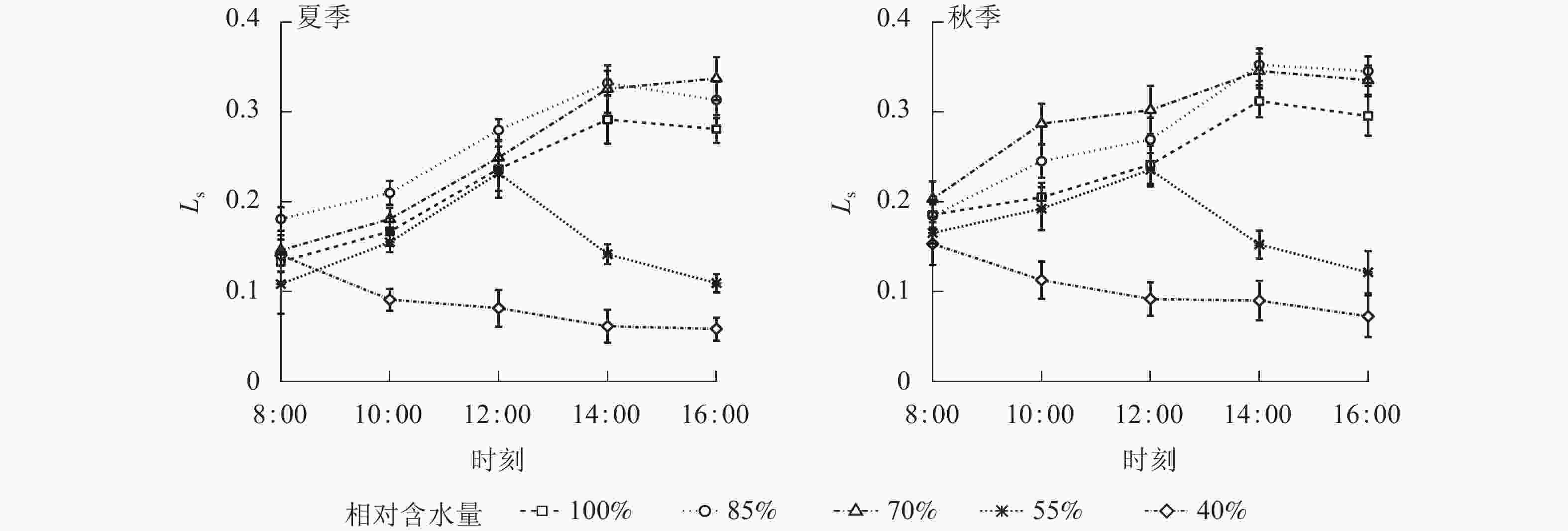
图 8 夏、秋季不同喷播基质含水量下黑麦草气孔限制值(Ls)的日变化
Figure 8. Diurnal variation of stomatal limit value (Ls) of L. perenne under different spraying substrate water content in summer and autumn
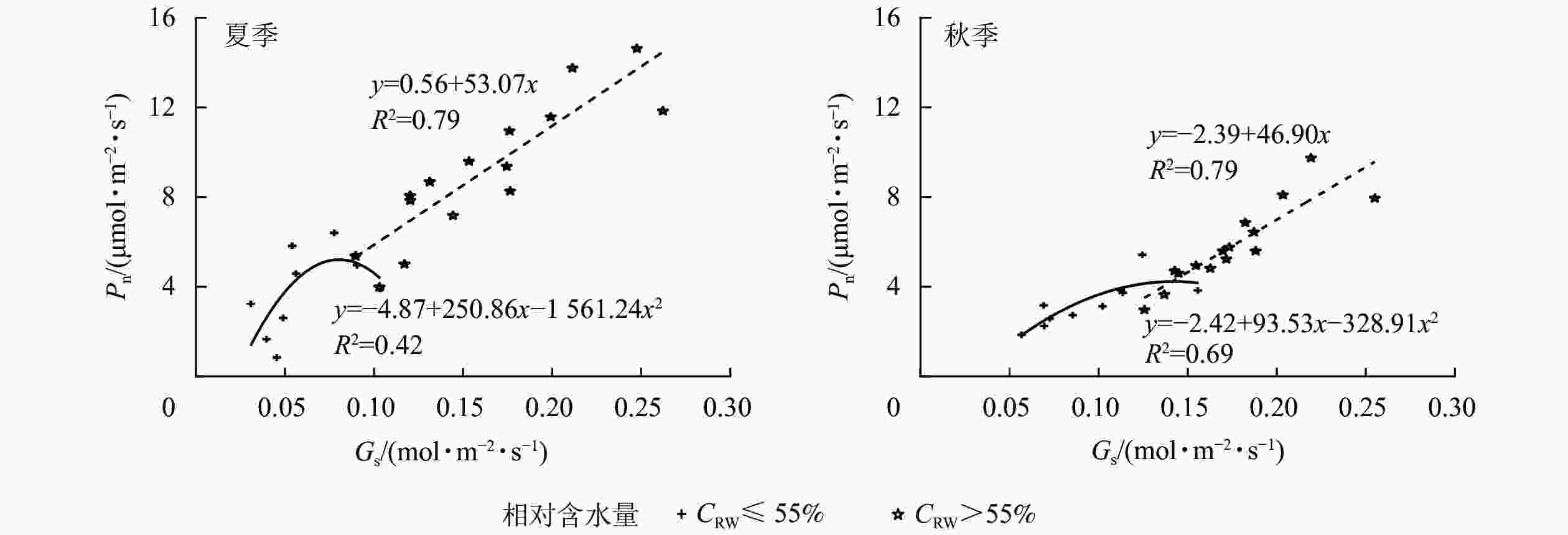
图 9 夏、秋季黑麦草净光合速率(Pn)和气孔导度(Gs)的关系
Figure 9. Relationship between net photosynthetic rate (Pn) and stomatal conductance (Gs) of L. perenne in summer and autumn
-
为进一步确定黑麦草喷播基质相对含水量(CRW)分级临界值,对黑麦草Pn、Tr、EWU和Gs的日均值与CRW构建回归模型(表3)。由Pn与CRW的回归模型知:夏、秋季Pn达最大值的CRW分别为78.17%、76.02%,其对应的最大Pn分别为9.68和 6.33 μmol·m−2·s−1。令Pn=0,求出夏、秋季水合补偿点的CRW分别为35.02%、30.83%(CRW大于100%的点均已舍去)。根据回归模型的积分式[2]求出CRW为40%~100%时黑麦草夏季Pn平均值为7.77 μmol·m−2·s−1,对应的CRW分别为58.98%和97.36%。同理可求出黑麦草秋季Pn平均值为5.29 μmol·m−2·s−1,对应的CRW分别为57.71%和94.33%。由此可以确定黑麦草夏、秋季Pn达到中等以上水平的CRW分别为58.98%~97.36%、57.71%~94.33%。
表 3 夏、秋季黑麦草光合参数与喷播基质相对含水量的回归模型
Table 3. Regression model between photosynthetic parameters of L. perenne and relative water content of spraying substrate in summer and autumn
参数 季节 回归模型 决定系数 F P Pn 夏季 y=−22.092 7+0.813 0x−0.005 2x2 0.88 78.98 9.12×10−11 秋季 y=−11.584 0+0.471 3x−0.003 1x2 0.81 45.60 1.49×10−8 Tr 夏季 y=−9.497 1+0.398 7x−0.002 5x2 0.94 595.83 0.000 秋季 y=−0.574 0+0.083 8x−0.000 5x2 0.83 39.08 5.74×10−8 EWU 夏季 y=−0.844 9+0.061 0x−0.000 4x2 0.80 31.05 3.93×10−7 秋季 y=−2.344 8+0.122 2x−0.000 83x2 0.76 35.29 1.37×10−7 Gs 夏季 y=−0.354 2+0.013 5x−0.000 086x2 0.78 39.97 4.73×10−8 秋季 y=−0.319 7+0.012 2x−0.000 077x2 0.83 53.94 3.30×10−9 说明:y表示各参数,x表示喷播基质相对含水量(CRW) 根据EWU与CRW的回归模型,求出夏、秋季EWU达最大值的CRW分别为76.25%、73.61%,对应的最大值分别为1.48和 2.15 μmol·mmol−1。令EWU=0,求出夏、秋季的对应的CRW分别为15.41%、22.68%(CRW大于100%的点均已舍去)。根据回归模型的积分式求出CRW为40%~100%时黑麦草夏季EWU的平均值为1.35 μmol·mmol−1,对应的CRW分别为58.17%和94.33%。同理可求出黑麦草秋季EWU的平均值为1.89 μmol·mmol−1,对应的CRW分别为55.81%和91.42%。由此确定黑麦草夏、秋季EWU达到中等以上水平的CRW分别为58.17%~94.33%、55.81%~91.42%。
-
将Pn和EWU取最大值点、平均值点、最低值点和Pn下降气孔限制转折点的喷播基质CRW临界值,作为黑麦草喷播基质适宜含水量阈值分级临界点,建立喷播基质适宜含水量的阈值分级(表4)。此分级标准将Pn和EWU作为“产”“效”来评价黑麦草生产力和水分利用能力的依据,建立了黑麦草喷播基质适宜含水量阈值分级。以Pn=0时的水合补偿点作为临界点,低于此临界点划为“无产无效水”范围。Pn下降原因由气孔限制为主转为非气孔限制为主对应的CRW称为“Pn气孔限制转折点”。Pn和EWU取最大值时的CRW确定为“高产高效水”临界值点。依据Pn、EWU与CRW的回归模型积分式求解二者的平均值来确定Pn和EWU达到中等以上水平的临界点,在此范围内称为“中产”“中效”,此范围外称为“低产”“低效”。为更清晰地展示5种阈值分级类型,借助坐标轴对其划分参数和数值进行展示(图10)。
表 4 基于光合特性的黑麦草喷播基质适宜含水量阈值分级
Table 4. Threshold gradient of suitable water content of L. perenne spraying substrate based on photosynthetic characteristics
季节 临界值指标 临界点对应的CRW/% 基质适宜含水量阈值分级类型 基质适宜含水量阈值/% 夏季 Pn=0 35.02 无产无效水 <35.02 Pn(sl→nsl) 55.00 低产低效水 35.02~55.00,97.36~100.00 Pn取平均值(Pn-ave) 58.98~97.36 中产中效水 78.17~97.36 Pn取最大值(Pn-max) 78.17 中产高效水 55.00~76.25 EWU取最大值(EWU-max) 76.25 高产高效水 76.25~78.17 EWU取平均值(EWU-ave) 58.17~94.33 秋季 Pn=0 30.83 无产无效水 <30.83 Pn(sl→nsl) 55.00 低产低效水 30.83~55.00,94.33~100.00 Pn取平均值(Pn-ave) 57.71~94.33 中产中效水 76.02~94.33 Pn取最大值(Pn-max) 76.02 中产高效水 55.00~73.61 EWU取最大值(EWU-max) 73.61 高产高效水 73.61~76.02 EWU取平均值(EWU-ave) 55.81~91.42 说明:Pn=0为水合补偿点,Pn(sl→nsl)为Pn气孔限制转折点 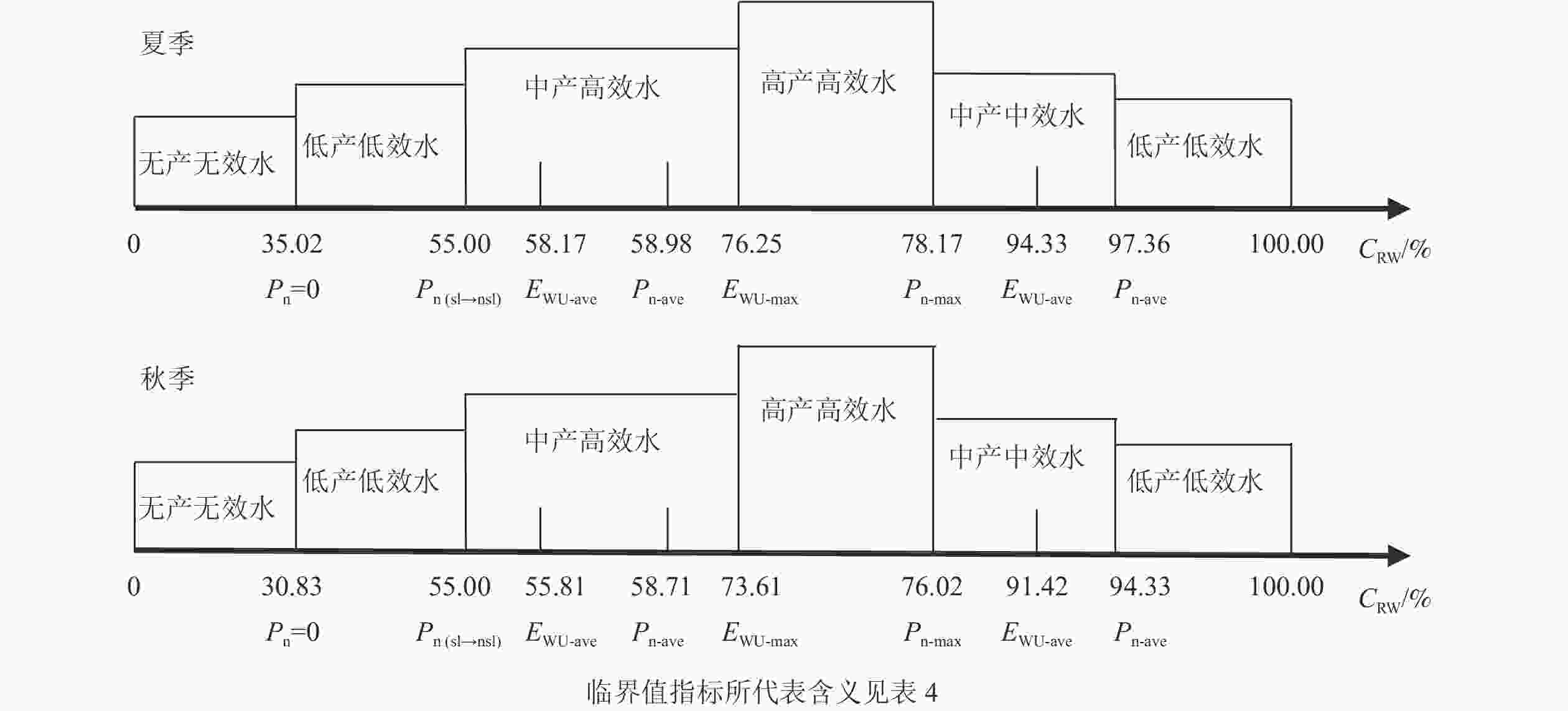
图 10 夏、秋季黑麦草喷播基质适宜含水量阈值分级坐标轴图示
Figure 10. Coordinate graphic figures of spraying substrate suitable water content threshold gradient of L. perenne in summer and autumn
-
夏、秋季黑麦草光合生理参数(Pn、Tr、EWU、Gs、Ci和Ls)日变化对喷播基质含水量的阈值响应规律与黄刺玫[5]、文冠果Xanthoceras sorbifolia[6]、连翘Forsythia suspensa[7]、山杏Prunus sibirica[8]、羊草Leymus chinensis和紫花苜蓿Medicago sativa[9]等对土壤水分阈值响应的规律一致,即CRW过高或过低均会抑制植物光合作用。CRW为70%~85%时,夏、秋季Pn、Tr日变化均呈现双峰曲线,在12:00表现出“光合午休”现象。主要原因是中午气温最高,高温影响植物光合酶的活性,降低Pn;空气相对湿度低,叶片表面饱和水汽压差增大,叶片气孔保卫细胞失水过多,导致部分气孔关闭,降低Tr和Pn[10]。CRW为100%时,夏、秋季Pn日变化均呈现单峰曲线,峰值出现在12:00,但Pn日均值并不高。表明水分充足可以延缓植物光合午休,但CRW过高,喷播基质孔隙较小,不利于根系呼吸,影响根系吸收营养元素,造成光合叶绿素含量降低,从而降低Pn[11]。CRW为100%时,夏、秋季Tr日变化的峰值延迟到14:00。已有研究表明:当水分充足时光照强度是影响Tr的主要因子,光合辐射可以促进叶片气孔开放,从而增强Tr[12-13]。CRW为40%~55%时,夏、秋季Pn和Tr均处于较低水平,原因是严重水分胁迫下植物为减少体内水分散失增加了气孔阻力[4],导致Pn和Tr降低。研究表明:适度的干旱胁迫能有效提高植物的水分利用效率[14-15],与本研究观点一致,即CRW为70%~85%时黑麦草EWU达最大值,并非在CRW最高的时候。秋季EWU显著高于夏季,主要原因是秋季Tr的降低幅度比Pn的降低幅度要更大,这与许多学者[16-18]的研究结果一致。
夏、秋季黑麦草Gs日变化与Pn的变化规律基本相似,但通过对Pn和Gs的关系拟合可知:Pn和Gs在CRW≤55%时两者为非线性关系,CRW>55%时为线性正比关系,这与郎莹等[19]的研究结果一致。轻度水分胁迫下,叶片气孔部分关闭,Gs下降,进入叶片CO2减少,因此Ci降低,Ls升高,但是当CRW为55%时,下午时段Gs下降,Ci升高,表明水分胁迫可能破坏了叶片的光合结构,导致叶片吸收CO2、光合作用能力下降。这也进一步说明,在CRW为55%时,黑麦草Pn下降原因已经由气孔限制为主转变为非气孔限制为主。已有研究表明:当植物光合作用受到非气孔限制时,水分胁迫可能开始损坏光合结构[20-21],叶绿体受损并且不可逆[22],当CRW进一步降低,植物叶子变黄甚至脱落[21]。因此,CRW=55%被认为是黑麦草喷播基质适宜含水量阈值分级的临界点。
采用Pn和EWU作为土壤水分的“产”“效”指标可评价土壤水分有效性和适宜含水量范围[2-3, 5, 7, 23],主要方法有3类:第1类为聚类分析法[4, 24],即通过试验获取多个水分梯度下的Pn和EWU进行聚类分析,得到不同的水分分级临界点。由于获取的水分梯度随机性较大,该方法缺乏足够代表性。第2类为极限值法,即通过获取Pn和EWU与CRW的定量关系,找出Pn和EWU的最低值、最大值点和气孔限制转折点,以此来划分水分分级临界点。但此法并未对中等水平的“产”“效”进行划分[2,6]。第3类为回归方程拟合法,即通过建立植物Pn和EWU与CRW的回归模型,计算Pn的水合补偿点、Pn和EWU最低值点、最大值点和平均值点对应的土壤水分,并以此作为土壤水分有效性阈值分级临界点。该方法对土壤水分分级比较完整[2-3, 21]。本研究结合第2类和第3类方法,即采用回归方程拟合法计算临界值点再结合Pn气孔限制转折点来确定喷播基质适宜含水量阈值分级标准。在拟合时采用了Pn和EWU的日平均值与CRW,相比只测上午光合数据[2, 4, 6, 21]的研究更具有代表性。本研究确定的“无产无效水”“低产低效水”“中产中效水”“中产高效水”和“高产高效水”5种喷播基质适宜含水量阈值分级类型,可以根据不同的工程绿化养护要求和黑麦草不同生长阶段对水分的需求来选择利用。例如,在裸露边坡等困难立地最突出的特征是干旱和缺水,坡面工程绿化以防治水土流失和提高水分利用效率为目标,而不是充分供水达到最高产量 [24-25]。因此既满足边坡植被修复要求,又不因灌溉量过大而造成坡面水土流失、影响植物生长和浪费水资源等问题,可以保持喷播基质含水量在“中产高效水”(55.00%≤CRW≤76.25%和55.00%≤CRW≤73.61%)的范围,以此为标准进行灌溉。
-
夏、秋季黑麦草净光合速率水合补偿点的喷播基质相对含水量分别为35.02%和30.83%,即实际质量含水量分别为10.63%和9.36%,喷播基质含水量低于此值光合作用无效。夏、秋季黑麦草净光合速率下降由气孔限制转变为非气孔限制的喷播基质相对含水量均为55%,即实际质量含水量为16.70%,喷播基质含水量低于此值将对黑麦草叶片光合结构造成不可逆性损坏,建议灌溉养护时保持基质含水量不能低于此水分范围。客土喷播绿化以快速恢复植被为目标时可以保持喷播基质含水量在“高产高效水”范围,以此为标准进行灌溉,夏、秋季分别为76.25%≤CRW≤78.17%和73.61%≤CRW≤76.02%,即实际质量含水量分别为23.15%~23.73%和22.35%~23.08%。客土喷播绿化以提高水分利用效率并恢复基本植被(即恢复到当地自然植被盖度为准)为目标时,可以保持喷播基质含水量在“中产高效水”范围,以此为标准进行灌溉,夏、秋季分别为55.00%≤CRW≤76.25%和55.00%≤CRW≤73.61%,即实际质量含水量分别为16.70%~23.15%和16.70%~22.35%。
Threshold gradient of water content of spraying substrate based on photosynthetic characteristics of Lolium perenne in summer and autumn
-
摘要:
目的 针对客土喷播基质过度灌溉造成坡面水土流失、影响植物生长等问题,提出植被生长的喷播基质适宜含水量阈值。 方法 采用种植盆模拟裸露创面客土喷播试验,设计5种喷播基质水分梯度;使用Li-6400XT便携式光合仪测定夏、秋季黑麦草Lolium perenne的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间二氧化碳(CO2)摩尔浓度(Ci)等光合参数,分析夏、秋季黑麦草光合参数与喷播基质含水量的关系。 结果 ①夏、秋季黑麦草净光合速率水合补偿点的喷播基质相对含水量(CRW)分别为35.02%和30.83%(即实际质量含水量分别为10.63%和9.36%)。②夏、秋季黑麦草Pn下降,由气孔限制转变为非气孔限制的喷播基质CRW均为55.00%(即实际质量含水量为16.70%)。③黑麦草喷播基质含水量阈值可分为5种类型,即“无产无效水”“低产低效水”“中产中效水”“中产高效水”“高产高效水”。 结论 ①客土喷播绿化以快速恢复植被为目标时,可保持喷播基质含水量在“高产高效水”范围,以此为标准进行灌溉,夏季为76.25%≤CRW≤78.17%,其实际质量含水量为23.15%~23.73%;秋季为73.61%≤CRW≤76.02%,其实际质量含水量为22.35%~23.08%。②客土喷播绿化以提高水分利用效率并恢复基本植被(即恢复到当地自然植被盖度)为目标时,可保持喷播基质含水量在“中产高效水”范围,以此为标准进行灌溉,夏季为55.00%≤CRW≤76.25%,其实际质量含水量为16.70%~23.15%;秋季为55.00%≤CRW≤73.61%,其实际质量含水量为16.70%~22.35%。图10表4参25 Abstract:Objective This study, with an analysis of how the over-irrigation of soil spraying substrate has caused soil erosion and affected plant growth, is aimed to put forward an optimal threshold of suitable water content of spraying substrate for vegetation growth. Method First, planting pots were used to simulate the experiment of soil spraying on bare slopes, and 5 kinds of water gradients of spraying substrates were designed. Then, efforts were made to measure the net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (Gs) and intercellular CO2 concentration (Ci) of Lolium perenne by Li-6400XT portable photosynthetic analyzer in summer and autumn before an analysis was conducted of the relationship between photosynthetic parameter of L. perenne and water content of spraying substrates. Result (1) The relative water content (CRW) of the spraying substrate at the net photosynthetic rate hydration compensation point of L. perenne in summer and autumn were 35.02% and 30.83% respectively (with the actual substrate mass water content being 10.63% and 9.36% respectively). (2) In summer and autumn, CRW of sprayed substrate, where the Pn of L. perenne decreased from stomatal limitation to non-stomatal limitation, was 55.00% (with the actual substrate mass water content being 16.70%). (3) The spraying substrate water content threshold of L. perenne comes in five types, namely non-productivity and non-efficiency water, low productivity and low efficiency water, middle productivity and middle efficiency water, middle productivity and high efficiency water and high productivity and high efficiency water. Conclusion When the goal of soil spraying and greening is to quickly restore vegetation, the water content of the spraying substrate can be kept in the range of high productivity and high efficiency water, based on which irrigation is carried out: in summer, it is 76.25%≤CRW≤78.17%, with its actual mass water content being 23.15%−23.73% whereas in autumn, it is 73.61%≤CRW≤76.02%, with its actual mass water content being 22.35%−23.08%. On the other hand, when the goal of soil spraying greening is to improve the water use efficiency and restore the basic vegetation (that is, to restore the local natural vegetation coverage), the water content of the spraying substrate can be kept in the range of “middle productivity and high efficiency water ”, based on which irrigation is carried out: in summer, it is 55.00%≤CRW≤76.25%, with its actual mass water content being 16.70%−23.15% whereas in autumn, it is 55.00%≤CRW≤73.61%, with its actual mass water content being 16.70%−22.35%. [Ch, 10 fig. 4 tab. 25 ref.] -
图 1 夏、秋季光合有效辐射(PAR)和大气CO2摩尔浓度(Ca)的日变化
Figure 1 Diurnal variation of photosynthetically active radiation (PAR) and atmospheric CO2 concentration (Ca) in summer and autumn
图 2 夏、秋季气温(Ta)和相对湿度(Rh)的日变化的日变化
Figure 2 Diurnal variation of temperature (Ta) and relative humidity (Rh) in summer and autumn
图 3 夏、秋季不同喷播基质含水量下黑麦草净光合速率(Pn)的日变化
Figure 3 Diurnal variation of net photosynthetic rate (Pn) of L. perenne under different spraying substrate water content in summer and autumn
图 4 夏、秋季不同喷播基质含水量下黑麦草蒸腾速率(Tr)的日变化
Figure 4 Diurnal variation of transpiration rate (Tr) of L. perenne under different spraying substrate water content in summer and autumn
图 5 夏、秋季不同喷播基质含水量下黑麦草水分利用效率(EWU)的日变化
Figure 5 Diurnal variation of water use efficiency (EWU) of L. perenne under different spraying substrate water content in summer and autumn
图 6 夏、秋季不同喷播基质含水量下黑麦草气孔导度(Gs)的日变化
Figure 6 Diurnal variation of stomatal conductance (Gs) of L. perenne under different water content of spraying substrate in summer and autumn
图 7 夏、秋季不同喷播基质含水量下黑麦草胞间CO2摩尔浓度(Ci)的日变化
Figure 7 Diurnal variation of intercellular CO2 concentration (Ci) of L. perenne under different water content of spraying substrate in summer and autumn
图 8 夏、秋季不同喷播基质含水量下黑麦草气孔限制值(Ls)的日变化
Figure 8 Diurnal variation of stomatal limit value (Ls) of L. perenne under different spraying substrate water content in summer and autumn
图 9 夏、秋季黑麦草净光合速率(Pn)和气孔导度(Gs)的关系
Figure 9 Relationship between net photosynthetic rate (Pn) and stomatal conductance (Gs) of L. perenne in summer and autumn
图 10 夏、秋季黑麦草喷播基质适宜含水量阈值分级坐标轴图示
Figure 10 Coordinate graphic figures of spraying substrate suitable water content threshold gradient of L. perenne in summer and autumn
表 1 基质层和种子层的材料配比
Table 1. Material ratio of matrix layer and seed layer
喷播层次 客土/
%木纤
维/%稻壳/
%复合肥/
(g·m−3)保水剂/
(g·m−3)黏合剂/
(g·m−3)基质层(10 cm) 70 10 20 300 200 150 种子层(3 cm) 67 33 说明:客土、木纤维和稻壳为体积比  下载: 导出CSV
下载: 导出CSV
表 2 夏、秋季不同喷播基质含水量下黑麦草光合生理参数的日均值变化
Table 2. Change of daily mean of photosynthetic physiological parameters of L. perenne under different spraying substrate water content in summer and autumn
CRW/% Pn/(μmol·m−2·s−1) Tr/(mmol·m−2·s−1) EWU/(mol·mol−1) 夏季 秋季 夏季 秋季 夏季 秋季 100 6.79±2.01 Abc 4.30±0.95 Bb 5.59±1.17 Aab 2.75±0.16 Bab 1.32±0.20 Bbc 1.56±0.26 Acd 85 11.17±3.08 Aa 6.07±1.24 Ba 6.83±1.12 Aa 3.13±0.40 Ba 1.61±0.22 Ba 1.92±0.22 Ab 70 9.26±2.79 Aab 7.02±1.97 Ba 6.76±0.63 Aa 2.92±0.59 Ba 1.43±0.15 Bab 2.37±0.25 Aa 55 5.77±2.09 Ac 3.77±1.03 Bb 4.91±0.93 Ab 2.35±0.30 Bbc 1.20±0.12 Bbc 1.63±0.20 Ac 40 2.80±1.66 Ae 2.74±0.78 Ab 3.03±0.87 Ac 2.16±0.28 Ac 1.01±0.28 Ac 1.28±0.16 Ad 说明:同列不同小写字母、同行不同大写字母均表示差异显著(P<0.05)
下载: 导出CSV
表 3 夏、秋季黑麦草光合参数与喷播基质相对含水量的回归模型
Table 3. Regression model between photosynthetic parameters of L. perenne and relative water content of spraying substrate in summer and autumn
参数 季节 回归模型 决定系数 F P Pn 夏季 y=−22.092 7+0.813 0x−0.005 2x2 0.88 78.98 9.12×10−11 秋季 y=−11.584 0+0.471 3x−0.003 1x2 0.81 45.60 1.49×10−8 Tr 夏季 y=−9.497 1+0.398 7x−0.002 5x2 0.94 595.83 0.000 秋季 y=−0.574 0+0.083 8x−0.000 5x2 0.83 39.08 5.74×10−8 EWU 夏季 y=−0.844 9+0.061 0x−0.000 4x2 0.80 31.05 3.93×10−7 秋季 y=−2.344 8+0.122 2x−0.000 83x2 0.76 35.29 1.37×10−7 Gs 夏季 y=−0.354 2+0.013 5x−0.000 086x2 0.78 39.97 4.73×10−8 秋季 y=−0.319 7+0.012 2x−0.000 077x2 0.83 53.94 3.30×10−9 说明:y表示各参数,x表示喷播基质相对含水量(CRW)
下载: 导出CSV
表 4 基于光合特性的黑麦草喷播基质适宜含水量阈值分级
Table 4. Threshold gradient of suitable water content of L. perenne spraying substrate based on photosynthetic characteristics
季节 临界值指标 临界点对应的CRW/% 基质适宜含水量阈值分级类型 基质适宜含水量阈值/% 夏季 Pn=0 35.02 无产无效水 <35.02 Pn(sl→nsl) 55.00 低产低效水 35.02~55.00,97.36~100.00 Pn取平均值(Pn-ave) 58.98~97.36 中产中效水 78.17~97.36 Pn取最大值(Pn-max) 78.17 中产高效水 55.00~76.25 EWU取最大值(EWU-max) 76.25 高产高效水 76.25~78.17 EWU取平均值(EWU-ave) 58.17~94.33 秋季 Pn=0 30.83 无产无效水 <30.83 Pn(sl→nsl) 55.00 低产低效水 30.83~55.00,94.33~100.00 Pn取平均值(Pn-ave) 57.71~94.33 中产中效水 76.02~94.33 Pn取最大值(Pn-max) 76.02 中产高效水 55.00~73.61 EWU取最大值(EWU-max) 73.61 高产高效水 73.61~76.02 EWU取平均值(EWU-ave) 55.81~91.42 说明:Pn=0为水合补偿点,Pn(sl→nsl)为Pn气孔限制转折点
下载: 导出CSV
-
[1] 王胤飞. 不同肥料及浓度对边坡修复植物生长的影响研究[D]. 北京: 中国地质大学, 2020. WANG Yinfei. Effects of Different Fertilizers and Concentrations on the Growth of Slope Restoration Plants [D]. Beijing: China University of Geosciences, 2020. [2] 夏江宝, 张淑勇, 赵自国, 等. 贝壳堤岛旱柳光合效率的土壤水分临界效应及其阈值分级[J]. 植物生态学报, 2013, 37(9): 851 − 860. XIA Jiangbao, ZHANG Shuyong, ZHAO Ziguo, et al. Critical effect of photosynthetic efficiency in Salix matsudana to soil moisture and its threshold grade in shell ridge island [J]. Chinese Journal of Plant Ecology, 2013, 37(9): 851 − 860. [3] 景雄, 范少辉, 蔡春菊, 等. 基于光合特性的毛竹实生苗土壤水分有效性及生产力分级[J]. 生态学杂志, 2021, 40(10): 3088 − 3097. JING Xiong, FAN Shaohui, CAI Chunju, et al. Classification of soil water availability and productivity of Moso bamboo (Phyllostachys edulis) seedlings based on photosynthetic characteristics [J]. Chinese Journal of Ecology, 2021, 40(10): 3088 − 3097. [4] 张淑勇, 夏江宝, 张光灿, 等. 黄刺玫叶片光合生理参数的土壤水分阈值响应及其生产力分级[J]. 生态学报, 2014, 34(10): 2519 − 2528. ZHANG Shuyong, XIA Jiangbao, ZHANG Guangcan, et al. Threshold effect of soil moisture on photosynthetic and physiological parameters in Rosa xanthina L. and its photosynthetic productivity classification [J]. Acta Ecologica Sinica, 2014, 34(10): 2519 − 2528. [5] 夏宣宣, 张淑勇, 张光灿, 等. 黄土丘陵区土壤水分对黄刺玫叶片光响应特征参数的影响[J]. 生态学报, 2016, 36(16): 5142 − 5149. XIA Xuanxuan, ZHANG Shuyong, ZHANG Guangcan, et al. Effects of soil moisture on the photosynthetic light reaction of Rosa xanthina L. in a loess hilly region [J]. Acta Ecologica Sinica, 2016, 36(16): 5142 − 5149. [6] 程甜甜, 张光灿, 张淑勇, 等. 不同土壤水分条件下文冠果的光合作用日变化[J]. 西北植物学报, 2016, 36(9): 1828 − 1835. CHENG Tiantian, ZHANG Guangcan, ZHANG Shuyong, et al. Photosynthesis diurnal variation of Xanthoceras sorbifolia Bunge under different soil water conditions [J]. Acta Botanica Boreali-Occidentalia Sinica, 2016, 36(9): 1828 − 1835. [7] 郎莹, 汪明. 春、夏季土壤水分对连翘光合作用的影响[J]. 生态学报, 2015, 35(9): 3043 − 3051. LANG Ying, WANG Ming. Effects of soil water on photosynthesis of Forsythia suspensa (Thunb. ) Vahl. in spring and summer [J]. Acta Ecologica Sinica, 2015, 35(9): 3043 − 3051. [8] 吴芹, 张光灿, 裴斌, 等. 不同土壤水分下山杏光合作用CO2响应过程及其模拟[J]. 应用生态学报, 2013, 24(6): 1517 − 1524. WU Qin, ZHANG Guangcan, PEI Bin, et al. CO2 response process and its simulation of Prunus sibirica photosynthesis under different soil moisture conditions [J]. Chinese Journal of Applied Ecology, 2013, 24(6): 1517 − 1524. [9] 于晓娜, 赵杰, 祁瑜, 等. 羊草和紫花苜蓿生长特征及光合特性对不同土壤水分的响应[J]. 生态学报, 2014, 34(21): 6067 − 6075. YU Xiaona, ZHAO Jie, QI Yu, et al. Effects of soil moisture content on morphological and photosynthetic characteristics of Leymus chinensis and Medicago sativa [J]. Acta Ecologica Sinica, 2014, 34(21): 6067 − 6075. [10] 张硕, 王姝怡, 李晴, 等. 近200年来新疆安迪尔地区红柳沙包稳定氧同位素特征与气候变化[J]. 干旱区资源与环境, 2019, 33(6): 119 − 125. ZHANG Shuo, WANG Shuyi, LI Qing, et al. The characteristics of stable oxygen isotope in Tamarix cone and climate change in recent 200 years in Andier, Xinjiang [J]. Journal of Arid Land Resources and Environment, 2019, 33(6): 119 − 125. [11] 岳文俊. 水氮供应和施肥频率对温室甜瓜生长和养分吸收的影响[D]. 杨凌: 西北农林科技大学, 2015. YUE Wenjun. Effects of Water, Nitrogen and Fertigation Frequency on Greenhouse Muskmelon Growth and Nutrient Absorption [D]. Yangling: Northwest A&F University, 2015. [12] 王鹏, 李海梅. 三种灌木的蒸腾耗水特性研究[J]. 北方园艺, 2011(2): 92 − 94. WANG Peng, LI Haimei. Study on characteristics transpiration water consumption of three types of shrubs [J]. Northern Horticulture, 2011(2): 92 − 94. [13] 王兴鹏, 文卿琳, 张维江. 毛乌素沙地南缘赖草水分生理特性及其影响因子的初步研究[J]. 塔里木大学学报, 2006, 18(1): 38 − 42. WANG Xingpeng, WEN Qinglin, ZHANG Weijiang. Study on the physiological character of A. dasystachys and influential factor in the south of MaoWuSu Sandlot [J]. Journal of Tarim University, 2006, 18(1): 38 − 42. [14] 王晓, 夏江宝, 周东兴, 等. 黄河三角洲贝壳砂干旱生境杠柳(Periploca sepium)叶片的光合作用特征[J]. 中国沙漠, 2019, 39(4): 139 − 148. WANG Xiao, XIA Jiangbao, ZHOU Dongxing, et al. Photosynthesis characteristics of Periploca sepium under drought stress in shell-sand habitat in the Yellow River Delta [J]. Journal of Desert Research, 2019, 39(4): 139 − 148. [15] BOUTRAA T, AKHKHA A, AL-SHOAIBI A A, et al. Effect of water stress on growth and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in Saudi Arabia [J]. Journal of Taibah University for Science, 2010, 3: 39 − 48. [16] LI Ping, LIU Wenxin, ZHANG Yinan, et al. Fungal canker pathogens trigger carbon starvation by inhibiting carbon metabolism in poplar stems [J/OL]. Scientific Report, 2019, 9: 10111[2022-01-11]. doi: 10.1038/s41598-019-46635-5. [17] 陈平, 孟平, 张劲松, 等. 两种药用植物生长和水分利用效率对干旱胁迫的响应[J]. 应用生态学报, 2014, 25(5): 1300 − 1306. CHEN Ping, MENG Ping, ZHANG Jinsong, et al. Effects of drought stress on growth and water use efficiency of two edicinal plants [J]. Chinese Journal of Applied Ecology, 2014, 25(5): 1300 − 1306. [18] 张子荷, 龚吉蕊, 晏欣, 等. 放牧干扰对内蒙古草甸草原羊草光合特性的影响[J]. 草业学报, 2018, 27(11): 36 − 48. ZHANG Zihe, GONG Jirui, YAN Xin, et al. Effects of grazing on photosynthetic characteristics of Leymus chinensis in meadow steppe of Inner Mongolia, China [J]. Acta Prataculturae Sinica, 2018, 27(11): 36 − 48. [19] 郎莹, 汪明. 春夏两季连翘光合作用的土壤水分阈值效应及生产力分级[J]. 林业科学, 2016, 52(2): 38 − 46. LANG Ying, WANG Ming. Threshold effect of photosynthesis in Forsythia suspense to soil water and its photosynthetic productivity grading in spring and summer [J]. Scientia Silvae Sinica, 2016, 52(2): 38 − 46. [20] 许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯, 1997, 33(4): 241 − 244. XU Daquan. Some problems in stomatal limitation analysis of photosynthesis [J]. Plant Physiology Communications, 1997, 33(4): 241 − 244. [21] 张光灿, 刘霞, 贺康宁. 黄土半干旱区刺槐和侧柏林地土壤水分有效性及生产力分级研究[J]. 应用生态学报, 2003, 14(6): 858 − 862. ZHANG Guangcan, LIU Xia, HE Kangning. Grading of Robinia pseudoacacia and Platycladus orientalis woodland soil’s water availability and productivity in semi-arid region of Loess Plateau [J]. Chinese Journal of Applied Ecology, 2003, 14(6): 858 − 862. [22] 王晓. 水分胁迫对贝壳堤植被杠柳光合效率及其水分生理的影响[D]. 哈尔滨: 东北农业大学, 2018. WANG Xiao. Effect of Photosynthetic Efficiency and Water Physiological Characteristics on Periploca sepium Bunge of Shell Ridge under Water Stress [D]. Harbin: Northeast Agricultural University, 2018. [23] 贺康宁, 田阳, 史常青, 等. 黄土半干旱区集水造林条件下林木生长适宜的土壤水分环境[J]. 林业科学, 2003, 39(1): 10 − 16. HE Kangning, TIAN Yang, SHI Changqing, et al. The suitable soil moisture condition for forest growth in catchment afforestation in semi-arid region on Loess Plateaus [J]. Scientia Silvae Sinica, 2003, 39(1): 10 − 16. [24] 夏江宝, 张光灿, 孙景宽, 等. 山杏叶片光合生理参数对土壤水分和光照强度的阈值效应[J]. 植物生态学报, 2011, 35(3): 322 − 329. XIA Jiangbao, ZHANG Guangcan, SUN Jingkuan, et al. Threshold effects of photosynthetic and physiological parameters in Prunus sibirica to soil moisture and light intensity [J]. Chinese Journal of Plant Ecology, 2011, 35(3): 322 − 329. [25] 秦景, 贺康宁, 朱艳艳. 库布齐沙漠几种常见灌木光合生理特征与土壤含水量的关系[J]. 北京林业大学学报, 2009, 31(1): 37 − 43. QIN Jing, HE Kangning, ZHU Yanyan. Relationship between photosynthetic physiological characteristics of several common shrub species and soil water content in Hobg Desert of Inner Mongolia, northern China [J]. Journal of Beijing Forestry University, 2009, 31(1): 37 − 43. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220116
 点击查看大图
点击查看大图
计量
- 文章访问数: 1486
- HTML全文浏览量: 266
- PDF下载量: 52
- 被引次数: 0
