





-
水分是影响植物生长和发育的重要环境因子,对植物的分布起着决定性作用[1]。全球气候变化背景下,大气降水格局不断发生改变[2−4],导致中国热带亚热带地区因降水时空分布不均引发的季节性干旱形势越发严峻[5−6],且气候模型预测这一趋势还将进一步加剧[7],这会对该地区植被的生长、固碳及生态系统的稳定性产生巨大威胁[8]。植物吸收和利用水分来源的模式在一定程度上决定了其对环境水分状况发生改变时的响应强度和结果[9−10],也影响着在干旱胁迫期水分管理对策的选择和判断。因此,对植物水分利用来源特征的了解,不但有助于评估和预测降水格局变化对未来植被结构的影响,还对在水分胁迫期精准指导林分水分管理和调控具有重大意义。
氢氧稳定同位素示踪技术是目前国内外确定植物水分来源及其吸收土壤水分层位最有效、最准确的方法[11−12]。基于除盐生植物和超旱生植物外,植物根系从吸水到传输至叶片和未栓化枝条前氢氧稳定同位素不发生分馏的现象,通过测定植物木质部和潜在水分来源的氢氧同位素值,结合溯源混合模型,可以有效确定植物根系吸收水分的来源[12−13]。目前,该技术已在国内外广泛应用,如王平元等[14]利用该技术发现:浅层土壤水是斜叶榕Ficus tinctoria全年最主要的水分来源;STRATTON等[15]利用该技术对夏威夷干旱森林中8种木本植物水分利用来源研究发现:除常绿物种多型铁心Metrosideros polymorpha和干旱落叶物种夏威夷雷诺木Reynoldsia sandwicensis主要利用深层土壤水外,其他6种植物均主要利用浅层土壤水。
桉树是华南地区最重要的速生丰产林树种,其巨大的林分生产力和碳汇功能关系着中国的木材供给安全和碳收支平衡。然而,研究却发现:桉树的生长受水分胁迫的限制比较严重[16−17]。全球气候变化背景下桉树种植区季节性干旱程度的不断加剧[6],使得桉树人工林的生产力及其碳汇功能面临严重下降的风险。在季节性干旱期制定科学合理的调控对策,对桉树水分利用来源特征的了解势在必行。尾巨桉Eucalyptus urophylla×E. grandis是中国最具代表性的桉树,约占桉树种植总面积的1/3,然而,利用氢氧稳定同位素对典型立地条件下尾巨桉人工林水分利用来源特征的研究鲜有报道。为此,本研究选取雷州半岛尾巨桉人工林为研究对象,在旱雨季典型月份对尾巨桉人工林木质部水、土壤水、雨水和地下水氢稳定同位素值(δD)和氧稳定同位素值(δ18O)进行测定,分析尾巨桉人工林水分利用来源的旱雨季变化,揭示其适应季节性干旱的水分来源利用策略,为桉树产业的可持续经营管理和水资源高效利用提供理论依据和数据支持。
-
研究地设置在广东湛江桉树林生态系统国家定位观测研究站内(21°16′N,110°05′E),该区海拔为80~220 m,属典型的海洋性季风气候。研究区多年平均气温和降水量分别为23.1 ℃和1319. 5 mm,其中极端最低气温为1.4 ℃,常发生在1月,极端最高气温为38.1 ℃,常发生在7月。研究区降水时空分布极为不均,高度集中在5—10月(雨季),占全年降水量的77%~85%[18],是典型的季节性干旱胁迫区。根据世界土壤资源参考数据库[19],研究地点的土壤被归类为砖红壤(暗红湿润铁铝土,Rhodi-Udic Ferralosols),由玄武岩的风化沉积物发展而成,呈酸性,0~80 cm土层深度的平均pH为4.9。
本研究的尾巨桉人工林营造于2012年7月,造林无性系为DH32-29,现存密度为1345 株·hm−2,叶面积指数为1.82,平均胸径为16.54 cm,平均树高为19.74 m,平均冠幅为3.12 m。在试验林内,设置3个具有代表性的样地,每个样地面积为400 m2。试验林林下植被丰富,主要灌木有野牡丹Melastoma candidum、马樱丹Lantana camara等;主要草本植物有飞扬草Euphorbia hirta、飞机草Eupatorium odoratum、蟛蜞菊Wedelia chinensis以及白花鬼针草HerbaBidentis等。
-
在尾巨桉人工林试验区附近开阔地带安置自动气象观测系统,装有Campbell公司CR3000型数据采集器、HMP155A空气温湿度传感器、TE525MM雨量筒及LI190SB型光合有效辐射传感器,可连续观测试验期间的大气温度(T, ℃)、空气相对湿度(HR, %)、降水量(R, mm)及光合有效辐射(PAR, μmol·s−1·m−2)等气象指标特征,隔10 min观测1次。
-
木质部样品:在3个固定样地内标记用于固定取样的植物个体3株,分别在雨季典型月份(6—8月)和旱季典型月份(12—2月)采集植株非绿色栓化枝条样本(直径为0.3~0.5 cm,长为3.0~5.0 cm),每株3个重复。迅速去除韧皮部,装入12 mL螺口玻璃采样瓶中,用parafilm膜密封,放入装好干冰的干冰桶带回。在进行水分抽提前放超低温冰箱−20 ℃冷冻保存。
土壤样品:采集植物样本的同时,在采样株附近挖取土壤剖面,分别在10、20、30、40、60、80、100、150和200 cm土层深处采集土壤样品,装入12 mL的螺口玻璃瓶中,并用parafilm膜密封,迅速放入装好干冰的干冰桶带回。在进行水分抽提前放超低温冰箱−20 ℃冷冻保存。稳定同位素取样的同时,对各土层进行环刀取土,用于准确测定各土层土壤含水量(烘干法)。
地下水样品:取距研究点最近的井水代表地下水(水位观测井),每次3个重复。将水装入12 mL螺口玻璃瓶中,parafilm膜密封,迅速装入4 ℃保温箱中带回实验室。在样品测定前放冰箱4 ℃冷藏保存。取样日期及频率与植物木质部取样保持一致。
大气降水样品:该地区的降水均以雨水的形式出现。采用国际原子能机构全球大气降水网络计划(GNIP)推荐的雨水收集装置采集雨样。收集过程中将蒸发作用的影响降到最低,尽量避免收集的雨水与外界空气交换。对观测期间所有单次雨量超过5 mm的降水分别采样记录,于降水事件结束后从收集瓶内取出2 mL(<2 mL时全部取出)样品装入采样瓶,parafilm膜密封,迅速装入4 ℃的保温箱中带回。在样品测定前放冰箱4 ℃冷藏保存。
-
首先利用超低温(−196 ℃)真空蒸馏抽提系统将植物木质部中的水和土壤样品中的水抽提出来,其次利用稳定同位素质谱仪(Delta V Advantage,美国)测定δD和δ18O;液态水δD和δ18O均以V-SMOW (Vienna standard mean ocean water)为标准;样品稳定同位素值计算公式为:δX=(Rsam/Rstd−1) ×1000‰。其中:δX为测定的对应样品的稳定同位素值,Rsam为样品中元素的重轻同位素比值;Rstd为国际通用标准物中元素重轻同位素之比(如D/H,18O/16O)。
-
在R 4.2.0中使用贝叶斯混合模型(MixSIAR)计算尾巨桉对各潜在水分来源的相对利用比例。考虑到提取或根系吸收水分过程可能存在氢同位素分馏效应,本研究利用该模型基于各层土壤水及地下水的δ18O实测值(source data)和茎木质部水的δ18O的实测值(mixture data)运行,其中马尔科夫链蒙特卡洛方法(Markov Chain Monte Carlo,MCMC)运行步长选择“Very long”,模型误差选取“Process+residual”,由此估算得到的每个水源对桉树水分利用的贡献率。
采用Excel 2016处理数据和制作表格,利用SPSS 19.0软件对所有数据进行统计分析并检验差异的显著性。用Origin 2020进行图表绘制。
-
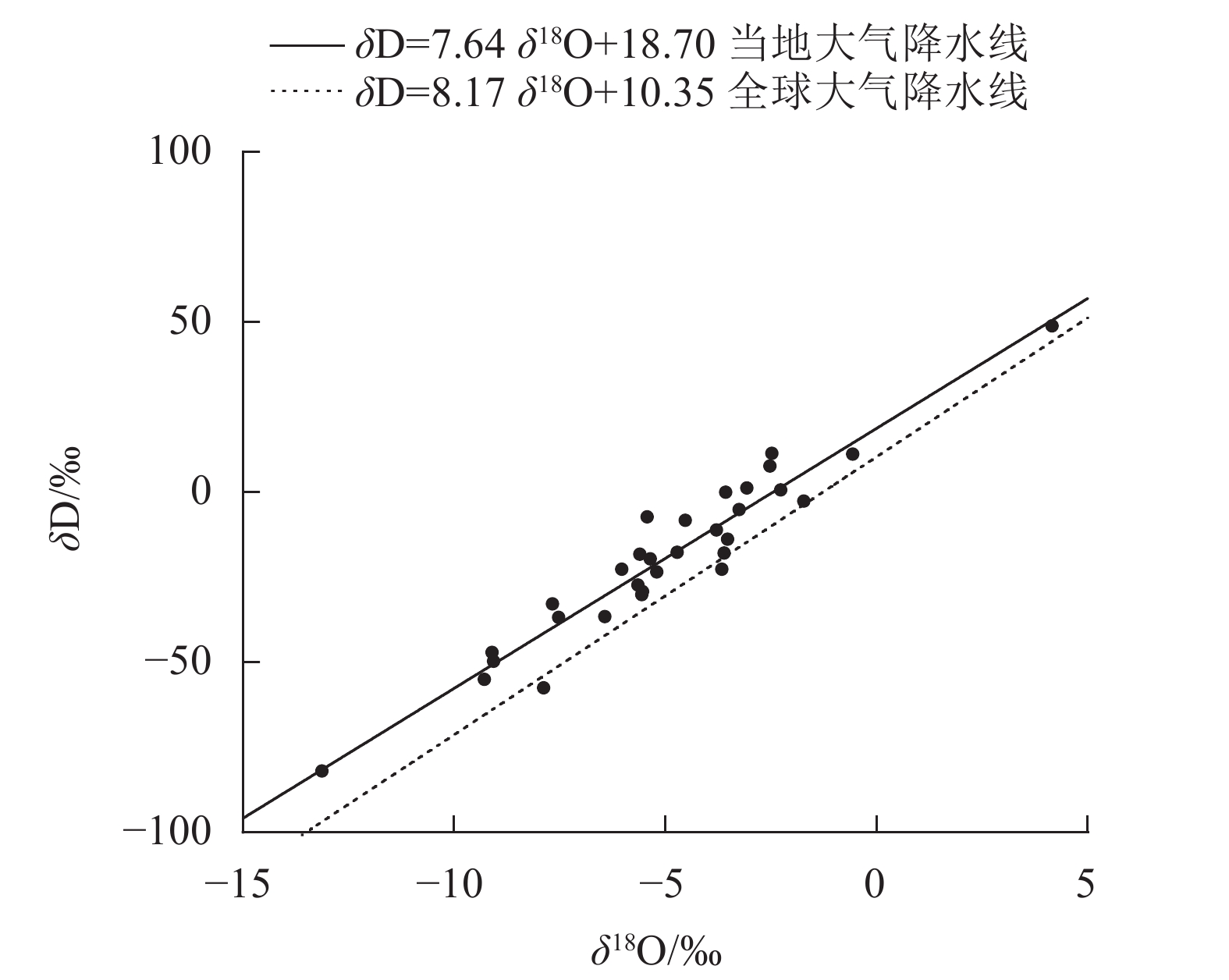
整个研究期间(2021年1月至2022年4月)总降水量为1531.5 mm,降水主要在5—10月,其降水量为976.5 mm(图1),占2021年全年降水量(1208.4 mm)的80.8%,旱雨季明显。其中在旱季典型月份取样期间的2月出现气候反常,降水量达111.6 mm。另外,对研究期间降水的δ18O监测发现:在1—4月δ18O平均为−3.12‰,5—10月δ18O平均为−6.75‰,11月至次年4月δ18O平均为−2.98‰,表现为旱季数据偏正,雨季数据偏负的季节变化规律;同时根据当地降水的δ18O和δD,线性拟合出当地大气降水线方程(图2):δD=7.64δ18O+18.70 (R2=0.919,P<0.001),与全球大气降水线方程相比,斜率偏小,表明当地大气降水在降落过程中发生了蒸发富集现象。
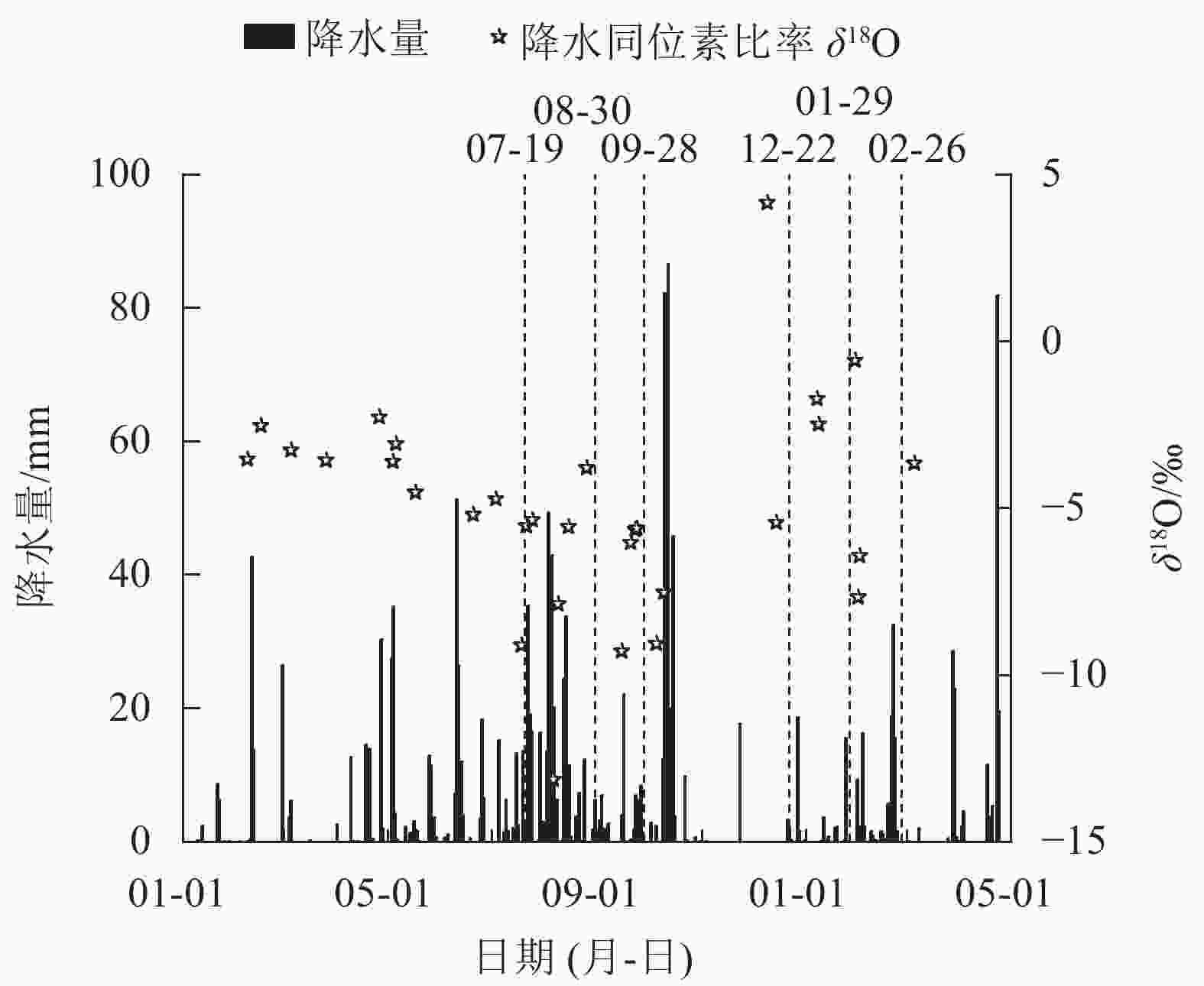
图 1 研究期间研究降水量的分布和降水的δ18O值
Figure 1. Distribution of precipitation and δ18O values of precipitation during the study period
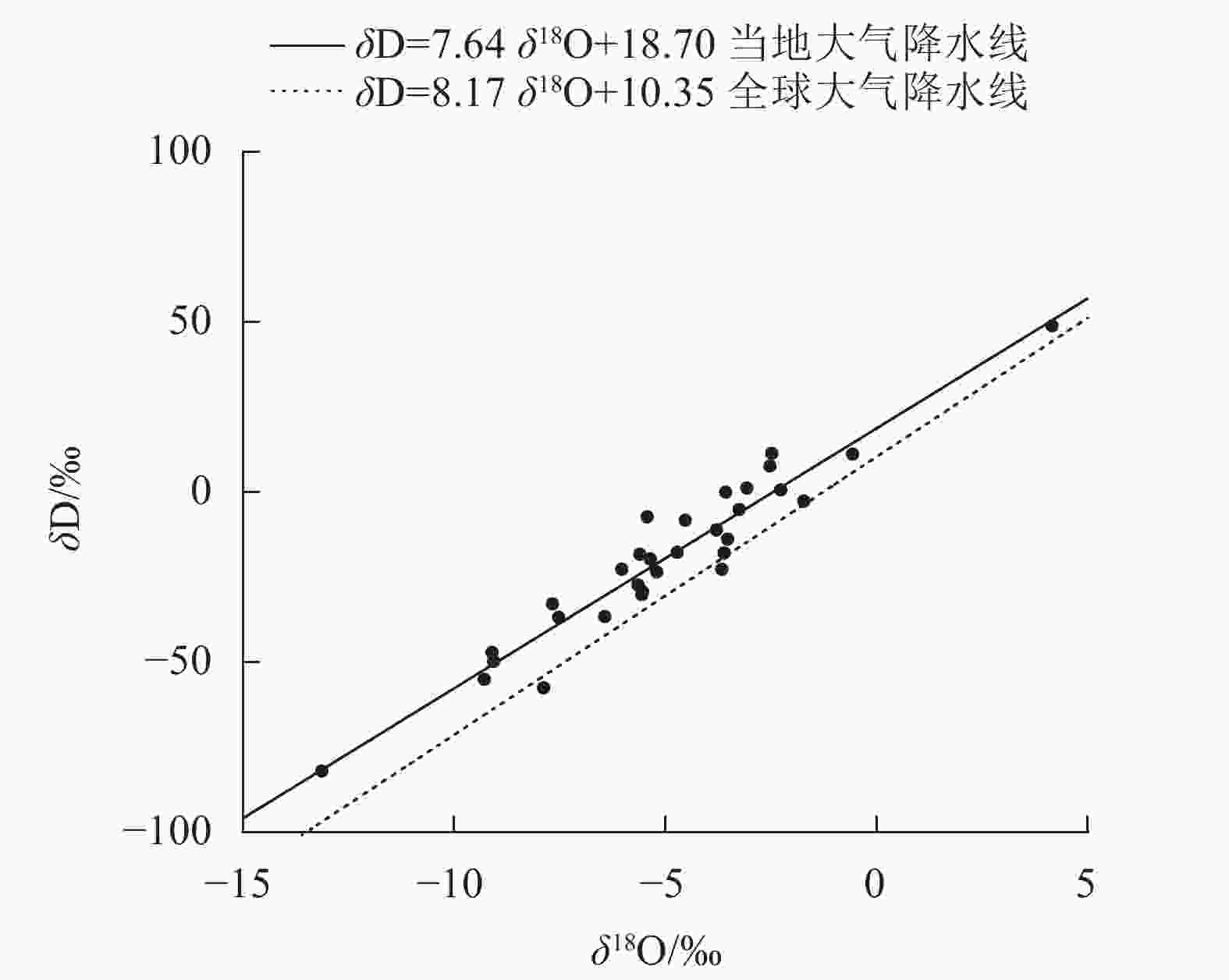
图 2 研究区当地大气降水线和全球大气降水线比较
Figure 2. Comparison of local meteoric water line in the study area and global meteoric water line
-
由图3可见:各月土壤含水量均随土壤深度的增加而增加,且越靠近表层的土壤受蒸发的影响越剧烈,变化幅度越大。雨季典型月份7、8、9月土壤含水量为24.5%~32.1%,0~200 cm土层平均含水量为29.2%,其中表层0~40 cm的平均土壤含水量也高达27.5%,土壤水分较为充沛;而旱季典型月份12和1月的土壤含水量为15.4%~26.8%,0~200 cm土层平均含水量为22.9%,显著小于雨季平均含水量(P<0.05),其表层0~40 cm的平均土壤含水量仅为19.3%,显著小于雨季表层土壤含水量及旱季深层土壤含水量25.8% (P<0.05),表明旱季水分较为亏缺,且浅层土壤亏缺较深层严重。旱季2月大量降水后(111.6 mm),土壤水分得到迅速补充,平均土壤含水量为28.3%,表层0~40 cm的平均土壤含水量提高到27.4%,已与雨季表层土壤含水量无显著差异。
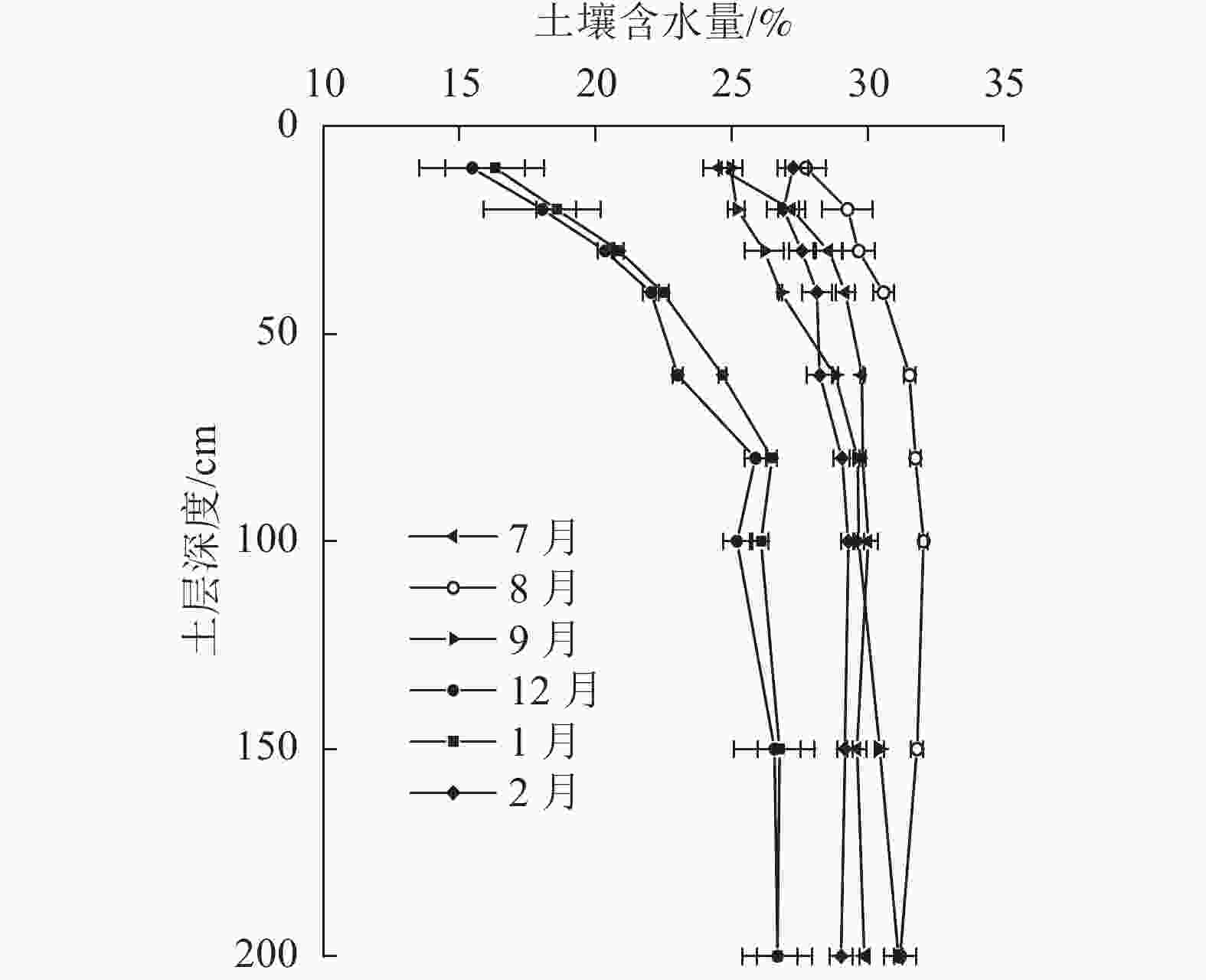
图 3 尾巨桉人工林不同月份土壤含水量变化特征
Figure 3. Change characteristics of soil water content of E. urophylla×E. grandis plantation in different months
-
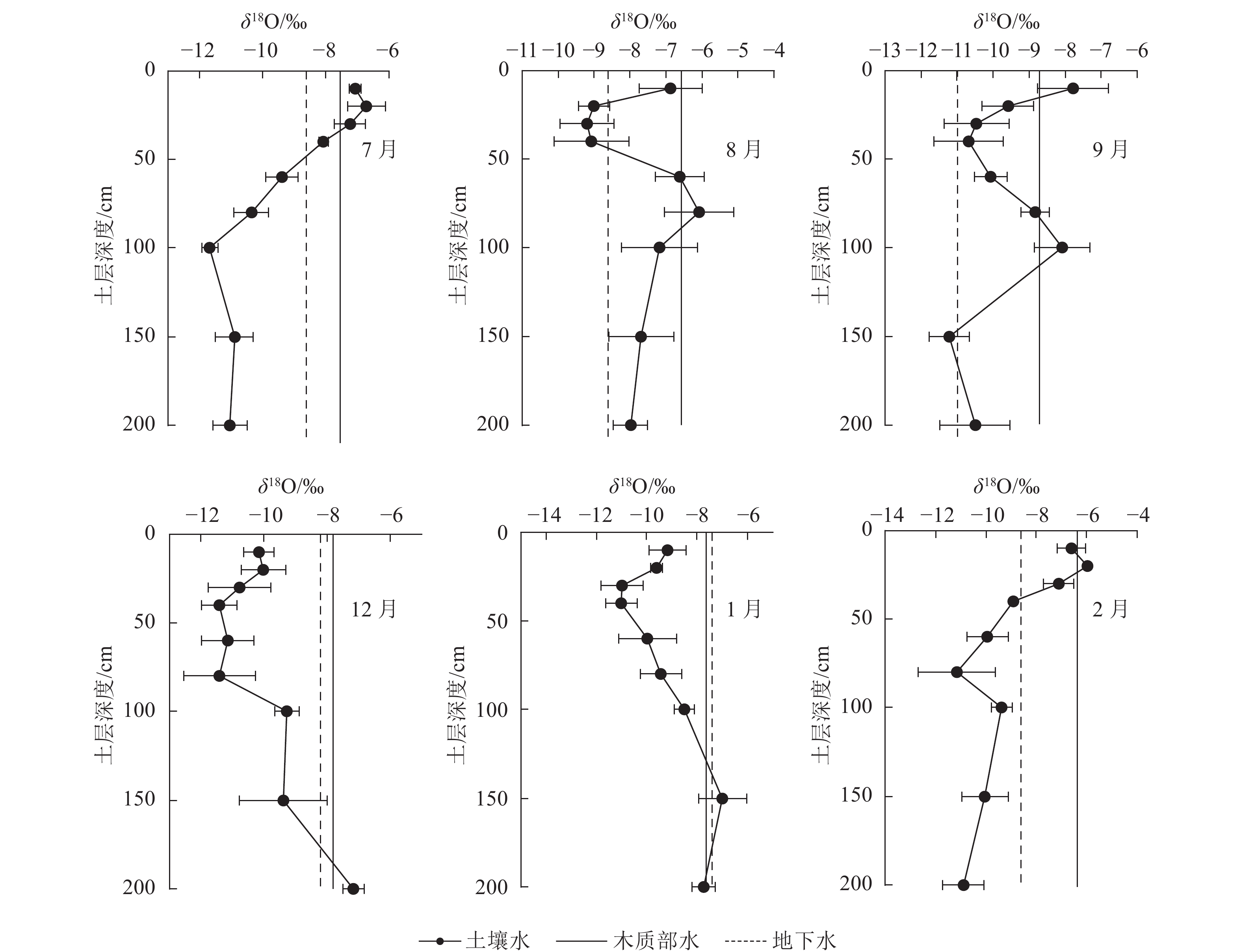
如图4所示:受不同月份近期降水与土壤原有水分同位素差异的联合影响,不同月份各土层中土壤水δ18O的变化特征不同,但表层0~40 cm土壤水的δ18O均随土层深度的增加而减小。这主要是由于雨水降落到地表并向土壤入渗的过程中发生了蒸发现象使得重同位素富集导致的。
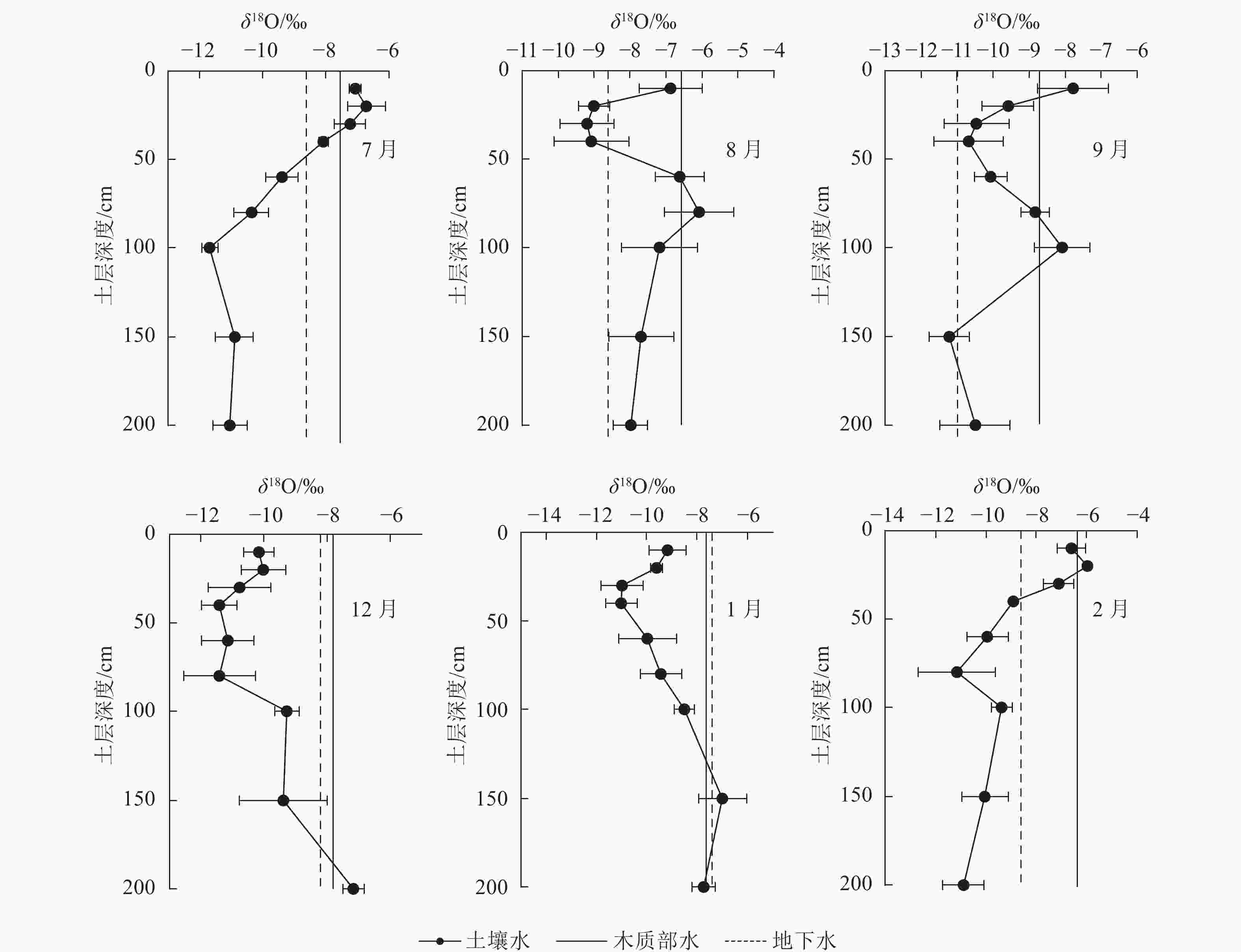
图 4 不同取样时间植物木质部水、各土层土壤水以及地下水的δ18O值
Figure 4. δ18O of stem water, soil water and groundwater at different sampling times
各月木质部水的δ18O也有所不同,说明尾巨桉在不同月份的水分来源可能有差异。在雨季,7月尾巨桉木质部水的δ18O与0~40 cm土层土壤水δ18O接近,且在30~40 cm土层处有1个交点,说明尾巨桉7月主要利用0~40 cm土层的土壤水;8月尾巨桉木质部水的δ18O与10及60~100 cm土层土壤水δ18O接近,且分别在60及80~100 cm土层处各有1个交点,说明尾巨桉8月主要利用10及40~100 cm土层的土壤水;9月尾巨桉木质部水的δ18O与0~20、80~100 cm土层土壤水δ18O接近,并分别相交于10~20 cm、80及100 cm附近,说明尾巨桉9月主要利用0~20及80~100 cm土层的土壤水。综合雨季典型月份水分来源的定性分析可以看出,尾巨桉在雨季主要利用0~100 cm土层的土壤水。
在旱季,12月尾巨桉木质部水的δ18O与地下水、150~200 cm土层土壤水δ18O接近,并相交于150~200 cm土层,表明尾巨桉12月主要利用150~200 cm土层的土壤水和地下水;1月,尾巨桉木质部水的δ18O分别在100~150及150~200 cm土层处相交,并与地下水、100 cm土层土壤水、150 cm土层土壤水和200 cm土层土壤水的δ18O接近,说明尾巨桉1月主要利用100~200 cm土层土壤水和地下水。2月,尾巨桉木质部水δ18O与0~30 cm土层处土壤水δ18O接近,且在10~20、20~30 cm土层处各有交点,说明尾巨桉2月主要利用0~30 cm土层的土壤水。综合旱季典型月份水分来源的定性分析可以看出:尾巨桉在旱季的干旱月份主要利用100~200 cm土层的土壤水和地下水,但旱季水分得到补充后(2月),又转为主要利用0~30 cm土层的土壤水。
-
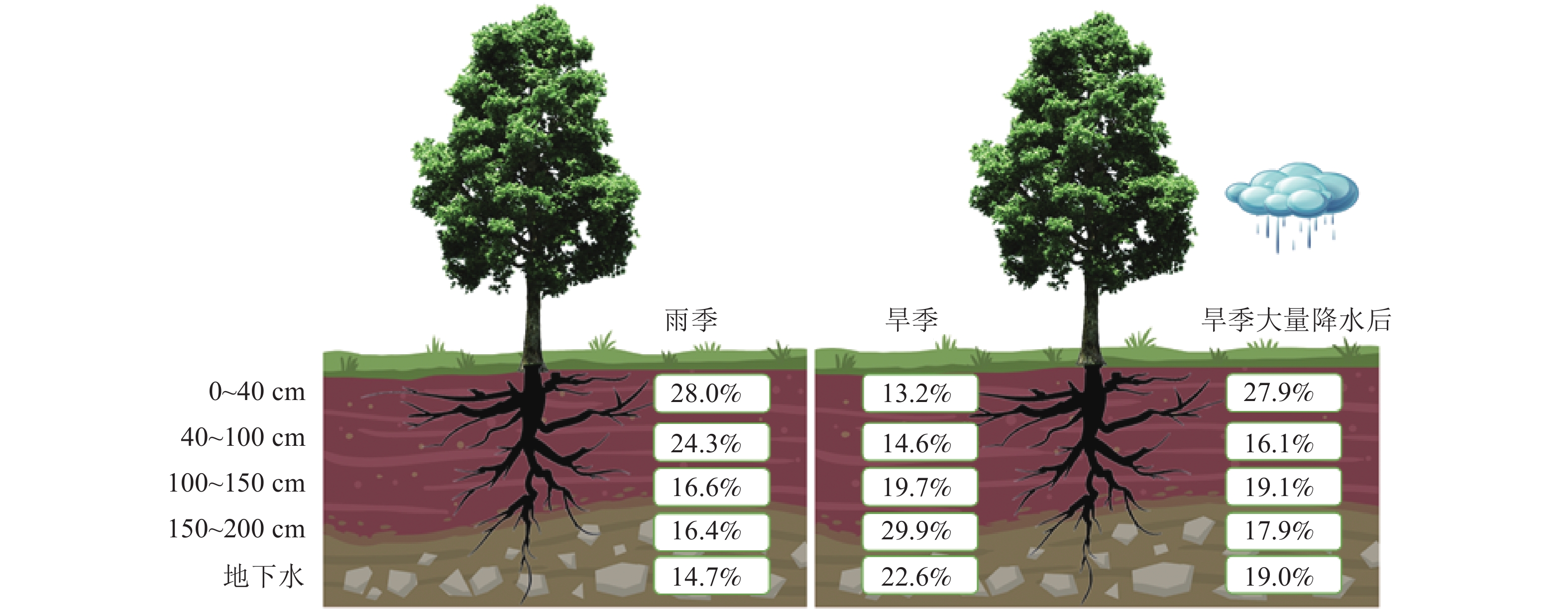
根据同位素质量守恒原理,通过MixSIAR模型对尾巨桉旱雨季典型月份的潜在水源利用比例进行定量估算(表1),结果表明:不同月份尾巨桉对不同深度土壤水和地下水的利用比例存在一定差异。7月尾巨桉对0~40 cm土层土壤水的利用比例最高,为45.5%,对其他土层水源的利用比例分别为12.1% (40~100 cm)、12.2% (100~150 cm)、11.7% (150~200 cm)和18.4% (地下水);8月主要利用40~100和100~150 cm土层土壤水,利用比例分别为30.9%和22.5%,而对其他土层水源利用比例相对较低:0~40 cm为15.8%、150~200 cm为18.0%、地下水为12.8%;9月则主要利用0~40和40~100 cm土层的土壤水,利用比例分别为22.7%和29.8%,对其他土层水源的利用比例分别为15.2% (100~150 cm)、19.4% (150~200 cm)以及13.0% (地下水)。12月降水较少,取样前仅为5.4 mm,此时尾巨桉主要利用150~200 cm土层的土壤水和地下水,利用比例分别为39.6%和23.3%,而对浅层土壤水的利用比例较低,其中0~40 cm土层为11.0%,40~100 cm土层为11.2%,100~150 cm土层为15.0%;1月同样降水极少,仅为24.7 mm,其主要利用100 cm土层以下的土壤水和地下水,利用比例分别为24.4% (100~150 cm)、20.3% (150~200 cm)和21.8% (地下水)。2月气候反常,降水量达111.6 mm,土壤水分得到大量补充,此时尾巨桉对0~40 cm土层土壤水的利用比例达27.9%,对其他土层水源利用比例相当,其中40~100 cm土层为16.1%,100~150 cm土层为19.1%,150~200 cm土层为17.9%,地下水为19.0%。
表 1 不同月份尾巨桉人工林对各潜在水源的利用比例
Table 1. Proportions of potential water sources for E. urophylla × E.grandis plantation in different months
土层深度/cm 各月尾巨按人工林对潜在水源的利用率/% 7月 8月 9月 12月 1月 2月 贡献率 范围 贡献率 范围 贡献率 范围 贡献率 范围 贡献率 范围 贡献率 范围 0~40 45.5 3.5~80.6 15.8 0.4~51.1 22.7 0.7~64.3 11.0 0.7~38.1 15.4 0.5~49.2 27.9 1.2~75.0 40~100 12.1 0.3~41.7 30.9 1.0~73.7 29.8 1.2~72.6 11.2 0.4~35.6 18.1 0.5~54.3 16.1 0.5~51.4 100~150 12.2 0.4~43.0 22.5 0.8~65.5 15.2 0.5~50.6 15.0 0.6~46.4 24.4 0.9~63.4 19.1 0.6~56.1 150~200 11.7 0.3~42.7 18.0 0.6~56.9 19.4 0.8~56.7 39.6 3.3~74.2 20.3 0.6~61.5 17.9 0.6~54.8 地下水 18.4 0.9~55.0 12.8 0.5~44.0 13.0 0.3~44.8 23.3 0.8~62.7 21.8 0.7~63.1 19.0 1.2~56.7 说明:贡献率表示植物对来源水的利用比例,范围表示来源水贡献率所出现的频率。 对尾巨桉旱季和雨季典型月份水分利用来源比例统计分析发现,尾巨桉旱雨季平均的主要水分利用来源存在明显差异(图5)。雨季尾巨桉主要利用0~40和40~100 cm土层土壤水,利用比例分别为28.0%和24.3%,而旱季则主要利用150~200 cm土层的土壤水和地下水,利用比例分别为29.9%和22.6%;此外,旱季大量降水后,尾巨桉会增加对表层土壤水的利用比例,降低深层土壤水和地下水的利用比例,如2月大量降水后,对0~40和40~100 cm土层土壤水的利用比例较12和1月的平均值分别增加了14.7%和1.5%,达27.9%和16.1%,提高率分别为111.4%和10.3%。而对100~150、150~200 cm土层的土壤水和地下水的利用率分别减少了0.6%、12.0%和3.6%。
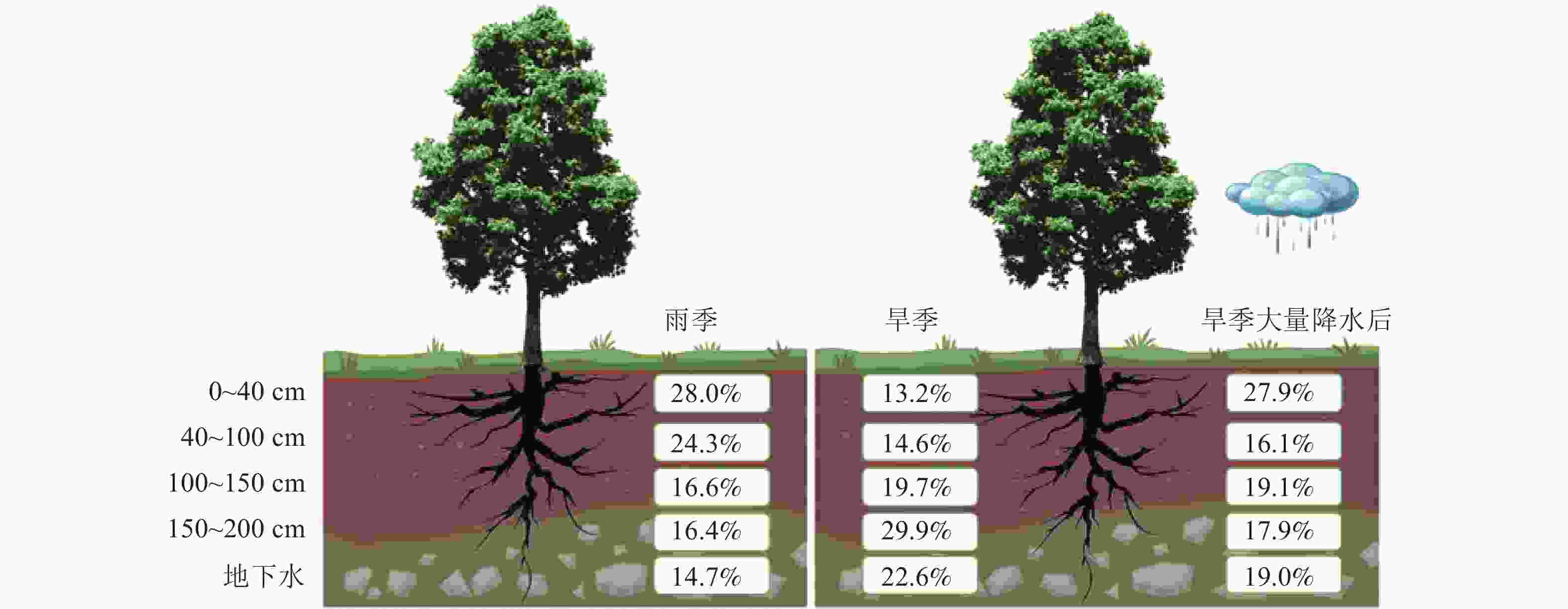
图 5 尾巨桉人工林雨季、旱季及旱季大量补水后的各潜在水源的利用比例变化
Figure 5. Changes in the proportion of each potential water source used in the wet and dry seasons and after substantial water supply in the dry season in E. urophylla × E. grandis plantations
-
中国的雷州半岛地区属于典型的海洋性季风气候,全年降水量主要在5—10月,旱雨季分明[20]。受低纬度高热和高辐射的影响,该地区的雨水在下落过程中往往会发生蒸发作用,导致同位素富集,因此本研究得到的当地大气降水线的斜率较全球大气降水线小。这与曾祥明等[21]对贵州普定县陈旗流域大气降水线的研究结果相似。此外,本研究还发现:当地降水的δ18O还表现出雨季偏负旱季偏正的季节性变化规律。这主要是由于雨季降水的水汽主要来源于低纬度海洋。受海洋水汽的影响,空气湿润且降水量大,蒸发作用较弱,重同位素富集作用轻,而旱季受大陆性气团的影响,空气干燥且降水量小,蒸发作用强,重同位素富集作用强导致的[22]。
林地的土壤水分及其同位素组成受降水、蒸发以及入渗等过程的综合影响[23],因此不同时间、不同地点及不同土层深度的土壤含水量及其δ18O往往存在较大差异[24]。众所周知,越接近地表的土壤受降水和蒸发等环境作用的影响就越大,因此表层土壤的含水量往往最小,且变化幅度最大。随着土层的加深,受外界环境影响不断减弱,土壤含水量逐渐增加并趋于稳定[25]。这与本研究对尾巨桉各土层深度土壤含水量的研究结果相符。受降水时空分配不均的影响,7—9月0~200 cm土层平均土壤含水量显著大于12和1月(P<0.05),且越接近土壤表层这种差异越明显,因此,旱季干旱月份浅层土壤的水分亏缺程度要比深层土壤更为严重,这也与表层土壤受到蒸发作用最为强烈有关。另外,地表土壤水分的蒸发,还会导致表层土壤水δ18O的富集[26],因此各月0~40 cm土层的土壤δ18O均随土层的加深而逐渐偏负,表现为明显的分馏效应。随着土层深度的增加,这种蒸发分馏效应逐渐减弱,再加上深层土壤水δ18O不但受近期降水的影响,还与更早降水入渗补给混合形成的新土壤水的原水分同位素有关[27−28],因此,40~200 cm土层土壤水的δ18O变化规律各月有所不同,但基本遵循先增大后减小或趋于稳定的变化规律。综合降水、入渗和蒸发的影响,尾巨桉人工林0~200 cm土层土壤水分的δ18O基本遵循先减小后增大再减小或趋于稳定的状态,这与杨爱国等[12]对科尔沁沙地杨树Populus土壤水以及靳宇蓉等[29]对黄土高原黄绵土土壤水δ18O的分布特征的研究结果保持一致。
环境中可利用水资源在时间或空间上经常发生变化,植物往往会通过一定的水分利用来源策略来提升其适应水分胁迫的生存能力[30−32],不同植物适应干旱的水分利用策略可能不同。本研究中尾巨桉雨季主要利用0~40和40~100 cm的浅层土壤水分,而旱季干旱月份则主要利用150~200 cm的深层土壤水和地下水,并且在旱季土壤水分得到补充后尾巨桉又会增加浅层土壤水的利用比例,并降低深层土壤水和地下水的利用比例。这与丁亚丽等[33]对喀斯特地区3年生尾巨桉的水分利用深度有所不同,可能是由于林龄的不同尾巨桉根系分布特征也不同以及喀斯特地区土壤特性共同导致的[34−36]。由两者的研究结论可以推测:尾巨桉可以随着环境可利用水资源的变化灵活地改变其主要的水分利用来源的策略。这种策略对尾巨桉适应季节性干旱的意义重大[1, 37],同时也能最大程度减小根系吸收水分时的能量消耗[36, 38]。相似的研究结论也在其他树种中被发现,如北京山区的侧柏Platycladus orientalis雨季主要利用表层0~20 cm的土壤水,而旱季时期则主要利用深层土壤水[39];在雨季,德国西南部的欧洲冷杉Abies alba 70%的水来自于浅层土壤,而旱季则主要利用深层土壤水[40]。与上述水分利用策略不同的是干旱区的物种面对季节性干旱可能采用更加保守的水分利用策略,如亚洲中部的红柳Tamarix ramosissima[30]以及西南亚高山地区的岷江冷杉Abies faxoniana[41]均主要依靠地下水生存,其生理活动和对地下水的利用比例均不受上层土壤水分含量变化的影响。古尔班通古特沙漠南缘的白梭梭Haloxylon persicum无论春季、夏季和秋季均主要利用100~300 cm的深层土壤水(42.6%~68.8%)[42−43],但同一地区的梭梭H. ammodendron在水分充足期(春季),主要利用0~40 cm的浅层土壤水(62%~95%)[42−44],这与尾巨桉雨季水分利用策略相似,但在干旱期(夏秋季),梭梭对地下水的利用比例高达68%~100%,且在外界降水后并不能增加其对土壤水分利用比例[42−44]。这与本研究对尾巨桉旱季大量降水后水分利用来源的研究结果不同,显示了梭梭在极端干旱条件下保守的水分利用策略。不同植物应对干旱时水分利用来源策略的差异,可能是其对环境长期适应的结果。全球气候变化背景下,桉树种植区的降水格局正在不断发生变化。本研究对尾巨桉水分利用来源特征的定量研究,对于评估未来降水格局对桉树产业的影响,精准指导桉树产业的可持续经营管理和水资源高效利用均有重要意义。
Difference in water use sources of Eucalyptus urophylla×E. grandis plantation in the Leizhou Peninsula during dry and rainy seasons
-
摘要:
目的 探讨桉树人工林适应季节性干旱的水分利用来源策略,为桉树产业的可持续经营管理和水资源高效利用提供数据支持。 方法 以雷州半岛分布面积最大的尾巨桉Eucalyptus urophylla×E. grandis为研究对象,通过测定旱雨季典型月份尾巨桉木质部水、各土层土壤水和地下水的氢稳定同位素值(δD)和氧稳定同位素值(δ18O),并利用贝叶斯混合模型(MixSIAR)量化尾巨桉旱雨季对各土层土壤水或地下水的利用比例,深入探讨了尾巨桉旱雨季水分利用来源的差异。 结果 当地大气降水存在明显的蒸发富集现象,且其δ18O呈明显的雨季偏负,旱季偏正的季节变化规律;各月土壤含水量均随深度的加深而增加最后趋于稳定,旱季土壤含水量显著(P<0.05)小于雨季,且越接近地表差异越大。尾巨桉在雨季主要利用0~40和40~100 cm土层的土壤水,利用比例分别为28.0%和24.3%,而旱季主要利用150~200 cm土层的土壤水和地下水,利用比例分别为29.9%和22.6%。旱季大量降水后,土壤水分尤其是浅层土壤水得到大量补充,尾巨桉对0~40和40~100 cm土层土壤水的利用比例分别提高了111.4%和10.3%,对100~150、150~200 cm土层土壤水和地下水的利用比例分别降低了3.1%、40.1%和15.9%。 结论 尾巨桉面对环境可利用水资源的变化,具有灵活多变的水分利用来源策略,这表明尾巨桉对季节性干旱具有较强的适应能力。图5表1参44 Abstract:Objective The objective is to explore the water use strategies of eucalyptus plantations to adapt to seasonal drought, so as to provide data support for assessing the impact of future rainfall patterns on eucalyptus industry and accurately guiding the sustainable management of eucalyptus industry and efficient use of water resources. Method Eucalyptus urophylla × E. grandis, the most widespread eucalyptus species in the Leizhou Peninsula, was taken as the research object. The differences in water use sources during dry and rainy seasons were discussed in depth by measuring the δD and δ18O values of xylem water, soil water in each soil layer and groundwater during typical dry and rainy seasons, and the MixSIAR model was used to quantify the utilization proportion of soil water and groundwater in each soil layer by E. urophylla × E. grandis in dry and rainy seasons. Result There was an obvious evaporative enrichment in local atmospheric precipitation, and the δ18O value showed a clear seasonal pattern of negative in rainy season and positive in dry season. Soil moisture content in each month increased with the depth and eventually became stable. Soil moisture content was significantly lower in dry season than in rainy season (P<0.05), and the difference was even greater close to the surface. E. urophylla × E. grandis mainly used 0−40 and 40−100 cm soil water in rainy season, with the utilization ratio of 28.0% and 24.3% respectively. In dry season it mainly used 50−200 cm soil water and groundwater, with the utilization ratio of 29.9% and 22.6% respectively. After a large amount of precipitation in dry season, soil water, especially shallow soil water, was greatly replenished, and the proportion of surface soil water used by E. urophylla × E. grandis in the 0−40 and 40−100 cm layers increased by 111.4% and 10.3% respectively, while the utilization proportion of 100−150 and 150−200 cm soil water and groundwater decreased by 3.1%, 40.1% and 15.9%, respectively. Conclusion E. urophylla × E. grandis has a flexible water use source strategy in the face of changes in environmental available water resources, indicating that it has strong adaptability to seasonal drought. [Ch, 5 fig. 1 tab. 44 ref.] -
图 1 研究期间研究降水量的分布和降水的δ18O值
垂直短划线指示的是植物茎样及土壤样品的取样时间
Figure 1 Distribution of precipitation and δ18O values of precipitation during the study period
图 2 研究区当地大气降水线和全球大气降水线比较
Figure 2 Comparison of local meteoric water line in the study area and global meteoric water line
图 3 尾巨桉人工林不同月份土壤含水量变化特征
Figure 3 Change characteristics of soil water content of E. urophylla×E. grandis plantation in different months
图 4 不同取样时间植物木质部水、各土层土壤水以及地下水的δ18O值
Figure 4 δ18O of stem water, soil water and groundwater at different sampling times
图 5 尾巨桉人工林雨季、旱季及旱季大量补水后的各潜在水源的利用比例变化
Figure 5 Changes in the proportion of each potential water source used in the wet and dry seasons and after substantial water supply in the dry season in E. urophylla × E. grandis plantations
表 1 不同月份尾巨桉人工林对各潜在水源的利用比例
Table 1. Proportions of potential water sources for E. urophylla × E.grandis plantation in different months
土层深度/cm 各月尾巨按人工林对潜在水源的利用率/% 7月 8月 9月 12月 1月 2月 贡献率 范围 贡献率 范围 贡献率 范围 贡献率 范围 贡献率 范围 贡献率 范围 0~40 45.5 3.5~80.6 15.8 0.4~51.1 22.7 0.7~64.3 11.0 0.7~38.1 15.4 0.5~49.2 27.9 1.2~75.0 40~100 12.1 0.3~41.7 30.9 1.0~73.7 29.8 1.2~72.6 11.2 0.4~35.6 18.1 0.5~54.3 16.1 0.5~51.4 100~150 12.2 0.4~43.0 22.5 0.8~65.5 15.2 0.5~50.6 15.0 0.6~46.4 24.4 0.9~63.4 19.1 0.6~56.1 150~200 11.7 0.3~42.7 18.0 0.6~56.9 19.4 0.8~56.7 39.6 3.3~74.2 20.3 0.6~61.5 17.9 0.6~54.8 地下水 18.4 0.9~55.0 12.8 0.5~44.0 13.0 0.3~44.8 23.3 0.8~62.7 21.8 0.7~63.1 19.0 1.2~56.7 说明:贡献率表示植物对来源水的利用比例,范围表示来源水贡献率所出现的频率。  下载: 导出CSV
下载: 导出CSV
-
[1] 王锐, 章新平, 戴军杰, 等. 亚热带地区不同林分下植物水分利用的季节差异[J]. 生态环境学报, 2020, 29(4): 665 − 675. WANG Rui, ZHANG Xinping, DAI Junjie, et al. Seasonal differences in water-uptake pattern of plants under different forest types in subtropical regions [J]. Ecology Environment Science, 2020, 29(4): 665 − 675. [2] MA Jian, ZHOU Lei, FOLTZ G R, et al. Hydrological cycle changes under global warming and their effects on multiscale climate variability [J]. Annals of the New York Academy of Sciences, 2020, 1472(1): 21 − 48. [3] WANG Xuyang, LI Yuqiang, WANG Mingming, et al. Changes in daily extreme temperature and precipitation events in mainland China from 1960 to 2016 under global warming [J]. International Journal of Climatology, 2021, 41(2): 1465 − 1483. [4] YADUVANSHI A, NKEMELAN T, BENDAPUDI R, et al. Temperature and rainfall extremes change under current and future global warming levels across Indian climate zones[J/OL]. Weather Climate Extremes, 2021, 31: 100291[2022-06-18]. doi: 10.1016/j.wace.2020.100291. [5] HAO Zengchao, SINGH V P, XIA Youlong. Seasonal drought prediction: advances, challenges, and future prospects [J]. Reviews of Geophysics, 2018, 56(1): 108 − 141. [6] ZHOU Guoyi, WEI Xiaohua, WU Yiping, et al. Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China [J]. Global Change Biology, 2011, 17(12): 3736 − 3746. [7] CHEN Sisi, YUAN Xing. CMIP6 projects less frequent seasonal soil moisture droughts over China in response to different warming levels[J/OL]. Environmental Research Letters, 2021, 16(4): 044053[2022-06-23]. doi: 10.1088/1748-9326/abe782. [8] GUPTA A, RICO M A, CANO-DELGADO A I. The physiology of plant responses to drought [J]. Science, 2020, 368(6488): 266 − 269. [9] EWE S M L, STERNBERG L S L, BUSCH D E. Water-use patterns of woody species in pineland and hammock communities of south Florida [J]. Forest Ecology and Management, 1999, 118(1/3): 139 − 148. [10] 徐庆, 冀春雷, 王海英, 等. 氢氧碳稳定同位素在植物水分利用策略研究中的应用[J]. 世界林业研究, 2009, 22(4): 41 − 46. XU Qing, JI Chunlei, WANG Haiying, et al. Use of stable isotopes of hydrogen, oxygen and carbon to identify water use strategy by plants [J]. World Forestry Research, 2009, 22(4): 41 − 46. [11] 徐庆, 刘世荣, 安树青, 等. 四川卧龙亚高山暗针叶林土壤水的氢稳定同位素特征[J]. 林业科学, 2007, 43(1): 8 − 14. XU Qing, LIU Shirong, AN Shuqing, et al. Characteristics of hydrogen stable isotope in soil water of sub-alpine dark coniferous forest in Wolong, Sichuan Province [J]. Scientia Silvae Sinicae, 2007, 43(1): 8 − 14. [12] 杨爱国, 付志祥, 王玲莉, 等. 科尔沁沙地杨树水分利用策略[J]. 北京林业大学学报, 2018, 40(5): 63 − 72. YANG Aiguo, FU Zhixiang, WANG Lingli, et al. Strategies on water utilization of poplar in Horqin Sandy Land of northern China [J]. Journal of Beijing Forestry University, 2018, 40(5): 63 − 72. [13] SPRENGER M, LEISTERT H, GIMBEL K, et al. Illuminating hydrological processes at the soil-vegetation-atmosphere interface with water stable isotopes [J]. Reviews of Geophysics, 2016, 54(3): 674 − 704. [14] 王平元, 刘文杰, 李金涛. 西双版纳绞杀植物斜叶榕的水分利用策略[J]. 应用生态学报, 2010, 21(4): 836 − 842. WANG Pingyuan, LIU Wenjie, LI Jintao. Water use strategy of Ficus tinctoria in tropical rainforest region of Xishuangbanna, Southwestern China [J]. Chinese Journal of Applied Ecology, 2010, 21(4): 836 − 842. [15] STRATTON L C, GOLDSTEIN G, MEINZER F C. Temporal and spatial partitioning of water resources among eight woody species in a Hawaiian dry forest [J]. Oecologia, 2000, 124(3): 309 − 317. [16] CAMPOE O C, ALVARES C A, CARNEIRO R L, et al. Climate and genotype influences on carbon fluxes and partitioning in Eucalyptus plantations [J/OL]. Forest Ecology and Management, 2020, 475: 118445[2022-06-23]. doi: 10.1016/j.foreco.2020.118445. [17] WANG Zhichao, DU Apeng, XU Yuxing, et al. Factors limiting the growth of Eucalyptus and the characteristics of growth and water use under water and fertilizer management in the dry season of Leizhou Peninsula, China [J]. Agronomy, 2019, 9(10): 590 − 607. [18] XU Yuxing, DU Apeng, WANG Zhichao, et al. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity[J/OL]. Forest Ecology and Management, 2020, 456: 117683[2022-06-08]. doi: 10.1016/j.foreco.2019.117683. [19] IUSS Working Group WRB. World Reference Base for Soil Resources[R]. Rome: FAO, 2006. [20] 王春林, 邹菊香, 麦北坚, 等. 近50年华南气象干旱时空特征及其变化趋势[J]. 生态学报, 2015, 35(3): 595 − 602. WANG Chunlin, ZOU Juxiang, MAI Beijian, et al. Temporal-spatial characteristics and its variation trend of meteorological drought in recent 50 years, South China [J]. Acta Ecologica Sinica, 2015, 35(3): 595 − 602. [21] 曾祥明, 徐宪立, 钟飞霞, 等. MixSIAR和IsoSource模型解析植物水分来源的比较研究[J]. 生态学报, 2020, 40(16): 5611 − 5619. ZENG Xiangming, XU Xianli, ZHONG Feixia, et al. Comparative study of MixSIAR and IsoSource models in the analysis of plant water sources [J]. Acta Ecologica Sinica, 2020, 40(16): 5611 − 5619. [22] 杨俊华. 中国大气降水δ18O的时空分布及其影响因素[D]. 北京: 中国科学院大学, 2013. YANG Junhua. Spatial and Temporal Distribution of δ18O in Precipitation Over China and Its Influence Factors[D]. Beijing: University of Chinese Academy of Sciences, 2013. [23] 王海燕, 刘廷玺, 王力, 等. 科尔沁沙地坨甸交错区土壤水分的空间变异规律[J]. 干旱区研究, 2013, 30(3): 438 − 443. WANG Haiyan, LIU Tingxi, WANG Li, et al. Spatial variation of soil moisture content in the dune-meadow ecotone in the Horqin Sandy Land [J]. Arid Zone Reseach, 2013, 30(3): 438 − 443. [24] 刘自强, 余新晓, 娄源海, 等. 北京山区栓皮栎林水分来源及生长季动态规律[J]. 北京林业大学学报, 2016, 38(7): 40 − 47. LIU Ziqiang, YU Xinxiao, LOU Yuanhai, et al. Water use strategy of Quercus variabilis in Beijing mountainous area [J]. Journal of Beijing Forestry University, 2016, 38(7): 40 − 47. [25] 王勇, 赵成义, 王丹丹, 等. 塔里木河流域不同林龄胡杨与柽柳的水分利用策略研究[J]. 水土保持学报, 2017, 31(6): 157 − 163. WANG Yong, ZHAO Chengyi, WANG Dandan, et al. Water use strategies of Populus euphratica and Tamarix ramosissima at different ages in Tarim River Basin [J]. Journal of Soil Water Conservation, 2017, 31(6): 157 − 163. [26] 容丽, 王世杰, 俞国松, 等. 荔波喀斯特森林4种木本植物水分来源的稳定同位素分析[J]. 林业科学, 2012, 48(7): 14 − 22. RONG Li, WANG Shijie, YU Guosong, et al. Stable isotope analysis of water sources of four woody species in the Libo Karst forest [J]. Scientia Silvae Sinicae, 2012, 48(7): 14 − 22. [27] EHLERINGER J R, DAWSON T E. Water uptake by plants: perspectives from stable isotope composition [J]. Plant,Cell and Environment, 1992, 15(9): 1073 − 1082. [28] EVARISTO J, JASECHKO S, MCDONNELL J J. Global separation of plant transpiration from groundwater and streamflow [J]. Nature, 2015, 525(7567): 91 − 94. [29] 靳宇蓉, 鲁克新, 李鹏, 等. 基于稳定同位素的土壤水分运动特征[J]. 土壤学报, 2015, 52(4): 792 − 801. JIN Yurong, LU Kexin, LI Peng, et al. Research on soil water movement based on stable isotopes [J]. Acta Pedologica Sinica, 2015, 52(4): 792 − 801. [30] XU Hao, LI Yan. Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events [J]. Plant and Soil, 2006, 285(1): 5 − 17. [31] HU Haiying, ZHU Lin, LI Huixia, et al. Seasonal changes in the water-use strategies of three herbaceous species in a native desert steppe of Ningxia, China [J]. Journal of Arid Land, 2021, 13(2): 109 − 122. [32] JIA Guodong, LIU Ziqiang, CHEN Lixin, et al. Distinguish water utilization strategies of trees growing on earth‐rocky mountainous area with transpiration and water isotopes [J]. Ecology and Evolution, 2017, 7(24): 10640 − 10651. [33] 丁亚丽, 陈洪松, 聂云鹏, 等. 基于稳定同位素的喀斯特坡地尾巨桉水分利用特征[J]. 应用生态学报, 2016, 27(9): 2729 − 2736. DING Yali, CHEN Hongsong, NIE Yunpeng, et al. Water use strategy of Eucalyptus urophylla × E. grandis on karst hillslope based on isotope analysis [J]. Chinese Journal of Applied Ecology, 2016, 27(9): 2729 − 2736. [34] 高娅, 贾志清, 李清雪, 等. 降雨对高寒沙地不同林龄中间锦鸡儿水分利用特征的影响[J]. 应用生态学报, 2021, 32(6): 1935 − 1942. GAO Ya, JIA Zhiqing, LI Qingxue, et al. Effects of precipitation on water use characteristics of Caragana intermedia plantations with different stand ages in alpine sandy land [J]. Chinese Journal of Applied Ecology, 2021, 32(6): 1935 − 1942. [35] 刘丽颖, 贾志清, 朱雅娟, 等. 青海共和盆地不同林龄乌柳林的水分利用策略[J]. 林业科学研究, 2012, 25(5): 597 − 603. LIU Liying, JIA Zhiqing, ZHU Yajuan, et al. Water use strategy of Salix cheilophila stands with different ages in Gonghe Basin, Qinghai Province [J]. Foretry Reseach, 2012, 25(5): 597 − 603. [36] 陈洪松, 聂云鹏, 王克林. 岩溶山区水分时空异质性及植物适应机理研究进展[J]. 生态学报, 2013, 33(2): 317 − 326. CHEN Hongsong, NIE Yunpeng, WANG Kelin. Spatio-temporal heterogeneity of water and plant adaptation mechanisms in karst regions: a review [J]. Acta Ecologica Sinica, 2013, 33(2): 317 − 326. [37] WANG Pingyuan, LIU Wenjie, ZHANG Jiaolin, et al. Seasonal and spatial variations of water use among riparian vegetation in tropical monsoon region of SW China[J/OL]. Ecohydrology, 2019, 12(4): e2085[2022-07-07]. doi: 10.1002/eco.2085. [38] SCHENK H J, JACKSON R B. The global biogeography of roots [J]. Ecological Monographs, 2002, 72(3): 311 − 328. [39] 刘自强, 余新晓, 贾国栋, 等. 北京山区侧柏和栓皮栎的水分利用特征[J]. 林业科学, 2016, 52(9): 22 − 30. LIU Ziqiang, YU Xinxiao, JIA Guodong, et al. Water use characteristics of Platycladus orientalis and Quercus variabilis in Beijing Mountain Area [J]. Scientia Silvae Sinicae, 2016, 52(9): 22 − 30. [40] MAGH R K, EIFERLE C, BURZLAFF T, et al. Competition for water rather than facilitation in mixed beech-fir forests after drying-wetting cycle[J/OL]. Journal of Hydrology, 2020, 587: 124944[2022-06-23]. doi: 10.1016/j.jhydrol.2020.124944. [41] XU Qing, LI Harbin, CHEN Jiquan, et al. Water use patterns of three species in subalpine forest, Southwest China: the deuterium isotope approach [J]. Ecohydrology, 2011, 4(2): 236 − 244. [42] WU Xue, ZHENG Xinjun, YIN Xinwei, et al. Seasonal variation in the groundwater dependency of two dominant woody species in a desert region of Central Asia [J]. Plant and Soil, 2019, 444(1): 39 − 55. [43] DAI Yue, ZHENG Xinjun, TANG Lisong, et al. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert [J]. Plant and Soil, 2015, 389(1/2): 73 − 87. [44] 戴岳, 郑新军, 唐立松, 等. 古尔班通古特沙漠南缘梭梭水分利用动态[J]. 植物生态学报, 2014, 38(11): 1214 − 1225. DAI Yue, ZHENG Xinjun, TANG Lisong, et al. Dynamics of water usage in Haloxylonammodendron in the southern edge of the Gurbantünggüt Desert [J]. Chinese Journal of Plant Ecology, 2014, 38(11): 1214 − 1225. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220481
 点击查看大图
点击查看大图

计量
- 文章访问数: 1604
- HTML全文浏览量: 487
- PDF下载量: 82
- 被引次数: 0
