




-
文冠果Xanthoceras sorbifolium为无患子科Sapindaceae文冠果属Xanthoceras植物,是中国北方特有的木本油料树种,其种仁含油率高,且富含不饱和脂肪酸,具有很高的食用价值和营养价值[1]。文冠果具有较强的生理适应性,抗寒旱、耐瘠薄,是北方造林绿化和防风固沙的优良生态树种[2]。文冠果不同的种质资源适应干旱的能力有明显的差异,丰产和抗旱的种质资源较为匮乏[3-4]。在全球水资源短缺问题不断加重的情况下,开展文冠果抗旱性评价,筛选文冠果抗旱种质资源对推动文冠果进一步扩大种植和推广及改善生态环境具有重要意义[5]。叶片是植物进行光合和蒸腾的主要部位,当植物长期生长在干旱少雨的地方时,其形态结构会发生变化来响应干旱胁迫,因此叶片的形态指标可以反映植物对干旱环境的适应情况[6-7]。基于叶片形态特征,研究者们发现文冠果叶片可通过提高蜡质层厚度和栅海比以及增加上表皮厚度来响应干旱,叶片厚度、栅海比、气孔密度和气孔开口面积指数等都可作为评价文冠果抗旱性的主要指标[8-10]。目前,中国通过叶片解剖结构指标来综合评价不同文冠果种质资源的抗旱性研究较少,因此,本研究基于文冠果种质资源圃内前期筛选的200份丰产型文冠果种质资源,通过观测叶片解剖结构的特征,采用因子和聚类分析的方法评价文冠果不同种质资源的抗旱性,以期为文冠果丰产抗旱型新品种选育提供理论依据。
-
样品采集自内蒙古自治区通辽市开鲁县东风林场文冠果种质资源圃,43°40′~ 43°42′N,121°34′~121°37′E。该种质资源圃是2005年从全国范围内收集文冠果适生区内多个种源(新疆、宁夏、甘肃、内蒙古、黑龙江、吉林、辽宁、北京、山东、陕西、山西、河南、河北等)播种建立的,位于内蒙古东部,属大陆性温带半干旱季风气候。年平均气温 为5.9 ℃,年平均降水量为 338.3 mm,无霜期约148 d。
-
选择树龄一致(15年生)、无病虫害、长势良好的实生丰产型文冠果植株,于2019年,在经历自然干旱且叶片完全展开时(7月下旬),在每株树正西方向中部选取朝向一致的6片成熟叶片,放入甲醛-乙酸-乙醇固定液(FAA)中固定。
-
将FAA固定液中的叶片取出,在每个叶片主脉中部选取0.5 cm×0.5 cm正方形组织块。采用常规石蜡切片法对叶片进行脱水、透明、浸蜡和包埋等处理,然后使用徕卡切片机切片,厚度为8~10 μm,番红固绿染色后用中性树胶封片,之后使用奥林巴斯BX51显微镜进行观察并拍照。使用Digimizer软件测量叶片厚度(leaf thickness, LT),上表皮厚度(thickness of upper epidermis, TU),下表皮厚度 (thickness of lower epidermis, TL),栅栏组织厚度(thickness of palisade tissue, TP),海绵组织厚度 (thickness of spongy tissue, TS),蜡质层厚度(leaf cuticle thickness, LCT),导管直径(vessel diameter, VD)。计算栅海比(palisade tissue and spongy tissue ratio, P/S),上表皮占比(proportion of the upper epidermis, PU),下表皮占比(proportion of lower epidermis, PL),蜡质层占比(proportion of leaf cuticle thickness, PLCT),组织结构紧密度(tightness of leaf palisade tissue, CTR)和组织结构疏松度(looseness of leaf spongy tissue, SR),每次测量选取3个视野,每个视野测定重复10次,所有数据为30个测定值的平均值。其中:栅海比(P/S)=栅栏组织厚度(TP)/海绵组织厚度(TS)×100%;上表皮占比(PU)=上表皮厚度(PU)/叶片厚度(LT)×100%;下表皮占比(PL)=下表皮厚度(PL)/叶片厚度(LT)×100%;蜡质层占比(PLCT)=蜡质层厚度(PLCT)/叶片厚度(LT)×100%;组织结构紧密度(CTR)=栅栏组织厚度(TP)/叶片厚度(LT)×100%;组织结构疏松度(SR)=海绵组织厚度(TS)/叶片厚度(LT)×100%。
-
数据使用 Excel 2019、SPSS 26.0与Rstudio 软件进行整理,根据不同文冠果种质资源的叶片解剖结构特征进行因子分析和聚类分析及方差分析。因子分析法是根据研究指标之间的相互关系,把多个具有错综复杂关系的变量归结为少数几个不相关的综合因子的一种多元统计分析方法[11]。因子分析前,先进行KMO (Kaiser-Meyer-Olkin)检验和巴特利特(Bartlett)球形检验,判定变量是否适合做因子分析。KMO值,即抽样适度测定值,取值范围为0~1,KMO值越接近于1 (>0.5),变量之间的相关性越强,原有变量越适合作因子分析;巴特利特球形检验(Bartlett)卡方值的显著水平P<0.05,因子分析的效果比较好[12-13]。为了确保结果的可靠性,通过SPSS软件采用标准差标准化(Z标准化)法对13个评价指标进行标准化处理,进行KMO检验和巴特利特(Bartlett)球形检验后运用分析模块进行因子分析,按照特征根≥1的原则提取公因子。并采用最大方差法对因子载荷矩阵进行旋转,再对旋转后的指标变量进行因子分析,从而明确各公因子代表的变量和意义。
同时采用隶属函数分析法对种质资源的抗旱性进行综合评价[14],隶属函数值越高,抗旱性越强,反之,抗旱性越弱[15]。其中,隶属函数值的计算公式如下:正相关:Ypq=(λpq−λqmin)/(λqmax−λqmin);负相关:Ypq=1−(λpq−λqmin)/ (λqmax−λqmin)。其中:Ypq第p组的第q个指标的隶属函数值;λpq为第p组的第q个指标的测定值;λqmax与λqmin分别为第q个指标的最大值和最小值。
-
由表1可见:叶片解剖结构13个指标的变异系数为18.78%~51.17%,说明200份文冠果种质资源的叶片解剖结构存在明显差异。其中,蜡质层厚度(LCT)为1.23~10.44 μm,在各材料中的变异系数最大,为51.17%。相比于其他性状,组织结构紧密度(CTR)的变异系数为18.78%,变异相对较小。各指标相关性分析(表2)发现:大多数性状之间显著(P<0.05)或极显著(P<0.01)相关。其中叶片厚度(TL)与上表皮厚度(TU)、下表皮厚度(TL)、栅栏组织厚度(TP)、海绵组织厚度(TS)和蜡质层厚度(LCT)等性状呈极显著相关(P<0.01)。组织结构疏松度(SR)与栅海比(P/S)、导管直径(VD)、蜡质层厚度(LCT)等性状呈极显著负相关(P<0.01)。性状之间的相关性表明:用单个叶片解剖结构指标来评价文冠果种质资源的抗旱性具有片面性,不能全面准确地评价干旱情况下文冠果叶片特性。
表 1 文冠果叶片解剖结构13个指标统计分析
Table 1. Statistics of 13 characters of leaf anatomical structure in X. sorbifolium
指标 最小
值/μm最大
值/μm均值/
μm标准
偏差变异系
数/%叶片厚度(LT) 60.22 270.04 140.90 50.99 36.19 上表皮厚度(TU) 3.97 24.10 11.95 5.22 43.65 下表皮厚度(TL) 4.26 21.36 10.13 4.04 39.87 栅栏组织厚度(TP) 22.34 182.31 62.66 28.22 45.04 海绵组织厚度(TS) 20.30 104.51 51.90 17.08 32.92 蜡质层厚度(LCT) 1.23 10.44 3.86 1.97 51.17 导管直径(VD) 4.03 25.01 9.74 4.41 45.31 栅海比(P/S) 35.26 263.46 121.69 38.93 32.00 上表皮占比(PU) 4.27 17.96 8.40 1.73 20.59 下表皮占比(PL) 4.19 21.34 7.26 1.84 25.28 蜡质层占比(PLCT) 1.25 5.43 2.68 0.74 27.47 组织结构紧密度(CTR) 23.08 93.76 43.79 8.22 18.78 组织结构疏松度(SR) 22.81 90.89 37.93 7.72 20.35 说明:样本数为200 表 2 200份文冠果叶片解剖结构13个性状的相关性分析
Table 2. Correlation between 13 traits of leaf anatomical structure in 200 X. sorbifolium samples
指标 LT TU TL TP TS LCT VD P/S PU PL PLCT CTR SR LT 1 TU 0.890** 1 TL 0.823** 0.881** 1 TP 0.903** 0.827** 0.754** 1 TS 0.832** 0.726** 0.734** 0.653** 1 LCT 0.832** 0.772** 0.769** 0.783** 0.719** 1 VD 0.851** 0.86** 0.829** 0.785** 0.711** 0.741** 1 P/S 0.393** 0.405** 0.313** 0.678** −0.075 0.366** 0.362** 1 PU 0.139* 0.552** 0.452** 0.187** 0.115 0.195** 0.334** 0.152** 1 PL −0.110 0.146* 0.447** −0.061 0.016 0.056 0.117 −0.064 0.595** 1 PLCT 0.214** 0.263** 0.357** 0.252** 0.233** 0.691** 0.261** 0.157* 0.221** 0.309** 1 CTR 0.230** 0.276** 0.251** 0.602** −0.021 0.266** 0.248** 0.838** 0.227** 0.137 0.213** 1 SR −0.394** −0.354** −0.220** −0.493** 0.154* −0.281** −0.323** −0.792** 0.010 0.293** 0.018 −0.412** 1 说明:缩写指代的指标见表1。*表示显著相关 (P<0.05);**表示极显著相关 (P<0.01) -
本研究中13个指标的KMO值为0.715,巴特利特(Bartlett)球形检验近似卡方值为4 891.701,显著性P=0.00(<0.05),说明本研究适合使用因子分析法来统计分析。在13个因子中,共提取出4个公因子F1、F2、F3、F4。4个公因子经旋转后的方差贡献率分别为42.154%、22.300%、14.885%、10.650%,累计贡献率达89.988%,能够较好地反映13个评价指标的信息。各因子的特征值、贡献率以及累计贡献率见表3。表4为旋转后的公因子载荷矩阵及得分系数矩阵,其中F1公因子在叶片厚度、海绵组织厚度、上表皮厚度、导管直径、下表皮厚度、栅栏组织厚度和蜡质层厚度上载荷较大,且这些变量均与叶片厚度呈显著正相关,因此F1公因子定义为厚度因子;F2公因子在栅海比、组织结构紧密度和组织结构疏松度上载荷较大,可定义为组织结构因子,其中组织结构疏松度对组织结构因子的影响是负向的;F3公因子的主要载荷量在上表皮占比和下表皮占比上,可定义为表皮占比因子;F4公因子中蜡质层占比最大,成为蜡质层占比因子。
表 3 主要评价指标贡献率
Table 3. Total variance contribution
指标 初始特征值 旋转后主因子 特征根 贡献
率/%累积贡
献率/%特征根 贡献
率/%累积贡
献率/%LT 6.554 50.418 50.418 5.480 42.154 42.154 TU 2.303 17.714 68.132 2.899 22.300 64.454 TL 1.799 13.837 81.969 1.935 14.885 79.339 TP 1.043 8.019 89.988 1.384 10.650 89.988 TS 0.608 4.677 94.665 LCT 0.409 3.145 97.811 VD 0.179 1.377 99.188 P/S 0.037 0.288 99.476 PU 0.023 0.176 99.652 PL 0.018 0.137 99.789 PLCT 0.014 0.109 99.898 CTR 0.009 0.066 99.964 SR 0.005 0.036 100 说明:缩写指代的指标见表1 表 4 旋转后的因子载荷及得分系数
Table 4. Factor loadings and factor scores coefficient after rotated
指标 变量 因子载荷 因子得分 F1 F2 F3 F4 F1 F2 F3 F4 LT X1 0.961 0.225 −0.105 0.051 0.211 −0.017 −0.115 −0.081 TU X2 0.900 0.250 0.282 0.003 0.179 0.006 0.116 −0.172 TL X3 0.848 0.137 0.397 0.157 0.151 −0.035 0.160 −0.044 TP X4 0.800 0.537 −0.025 0.122 0.123 0.126 −0.068 −0.012 TS X5 0.913 −0.263 −0.035 0.139 0.236 −0.209 −0.092 0.001 LCT X6 0.788 0.194 −0.021 0.558 0.095 −0.018 −0.133 0.379 VD X7 0.880 0.207 0.155 0.045 0.180 −0.011 0.038 −0.113 P/S X8 0.179 0.970 0.025 0.074 −0.074 0.370 0.006 0.020 PU X9 0.225 0.115 0.868 −0.039 0.003 0.034 0.490 −0.197 PL X10 −0.018 −0.115 0.890 0.229 −0.072 −0.037 0.473 0.075 PLCT X11 0.182 0.063 0.168 0.942 −0.106 −0.008 −0.052 0.779 CTR X12 0.062 0.827 0.208 0.202 −0.115 0.326 0.096 0.125 SR X13 −0.196 −0.821 0.207 0.148 0.009 −0.309 0.096 0.142 说明:缩写指代的指标见表1 由表4可以得到各指标在4个公因子上的得分。将各项指标变量定义为X1,X2
$, \cdots , $ X13,其在4个公因子上的得分表示为F1、F2、F3、F4,综合得分表示为F。根据4个公因子的权重,求得综合得分(F)的表达式为F=(λ1F1+λ2F2+λ3F3+λ4F4)/(λ1+λ2+λ3+λ4),其中λi (i=1, 2
$, \cdots , $ 4)分别为4个公因子的特征根。即:F=0.066X1+0.084X2+0.083X3+0.076X4+0.044X5+0.063X6+0.075X7+0.060X8+0.068X9+0.044X10+0.032X11+0.058X12−0.040X13。
其中:Xi (i=1, 2
$, \;\cdots , $ 13)为各样本经过标准化后各性状指标对应的数值。由上述表达式指标前的系数值(即权重)可以看出:13项指标中上表皮厚度(0.084)、下表皮厚度(0.083)、栅栏组织厚度(0.076)和导管直径(0.075)指标最能反映文冠果种质资源的抗旱能力,而组织结构疏松度(−0.040)与综合得分呈负相关关系,即叶片组织结构越疏松,综合得分越低,抗旱性越差。
-
基于因子分析得到的4个公因子的得分及综合得分,以Euclidean距离为度量标准,利用Ward法对200份种质资源进行系统聚类分析,将200份种质资源分成了A、B共2组,其中A组包括90份种质资源,B组包括110份种质资源。根据公式得到A组文冠果种质资源的综合得分为0.610,远高于B组得分−0.449(表5)。由图1可以看出:A组的叶片栅栏组织排列紧密且占比较大,紧密的栅栏组织可以有效地减少叶片的蒸腾作用,提高叶片的保水能力,从而提高植株适应干旱的能力。而B组的叶片栅栏组织和海绵组织则呈疏松状排列,细胞间隙较大,在植物受到干旱胁迫时,叶片不能维持稳定的含水量,从而影响植株的生长。
表 5 2种类型文冠果叶片解剖结构的隶属函数分析
Table 5. Subordinative function of 2 types of X. sorbifolium leaf anatomical structure
类型 隶属函数值 综合评价值 综合得分F LT TU TL TP TS LCT VD P/S PU PL PLCT CTR SR A组 0.610 0.638 0.563 0.411 0.471 0.476 0.467 0.461 0.346 0.195 0.433 0.338 0.820 0.479 0.610 B组 0.205 0.203 0.168 0.125 0.254 0.131 0.118 0.310 0.265 0.164 0.274 0.256 0.257 0.210 −0.449 说明:缩写指代的指标见表1 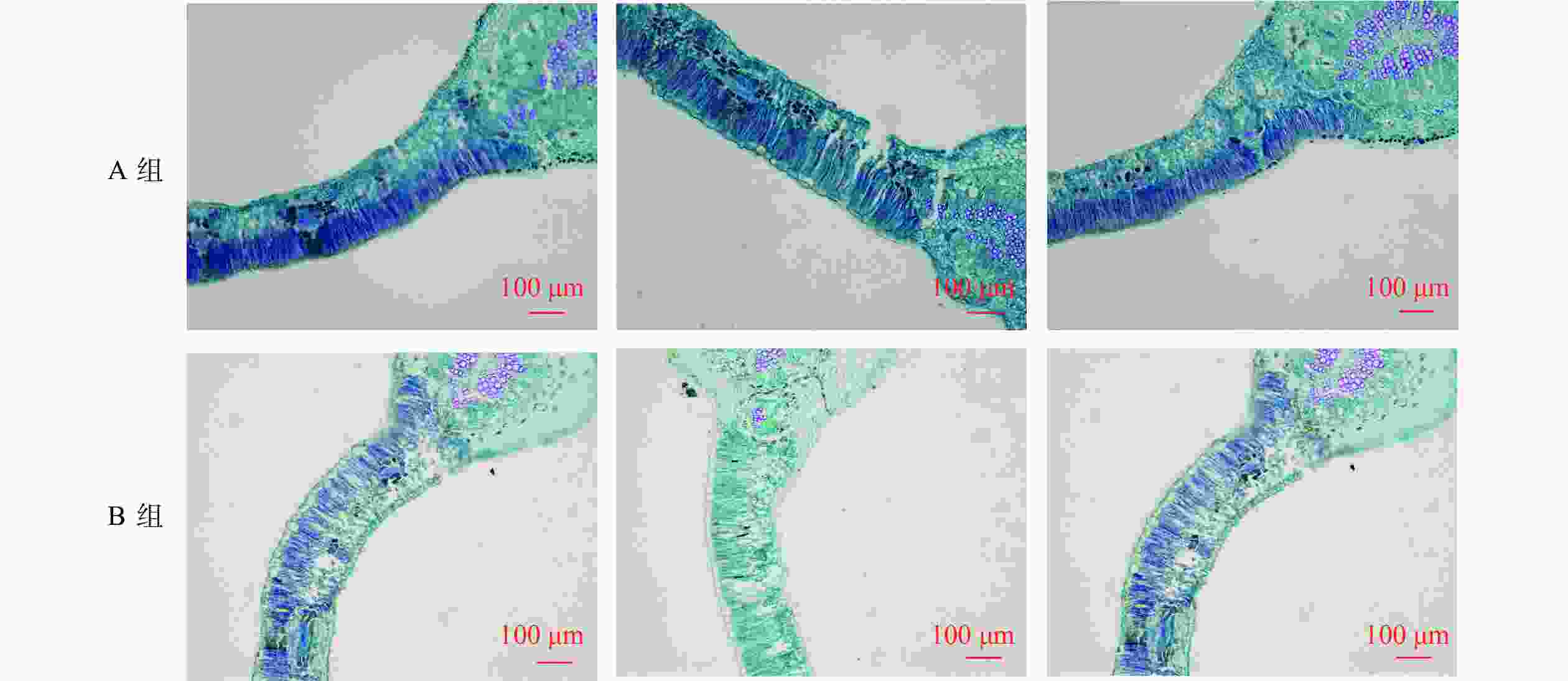
图 1 2种类型文冠果种质资源的叶片解剖结构特征
Figure 1. Leaf anatomical structure characteristics of 2 types of X. sorbifolium
对2组种质资源指标数据进行隶属函数分析(表5):A组的隶属函数值为0.479,高于B组(0.210),这与因子分析的综合得分结果一致,表明因子分析法对于评价文冠果种质资源抗旱性的适用性。方差分析结果(表6)表明:A组和B组种质资源的各项指标中除下表皮占比不存在显著差异外,其余指标均存在极显著差异。A组的叶片厚度、上/下表皮厚度、栅栏组织厚度和蜡质层厚度等都显著高于B组,而B组的组织结构疏松度较高,为40.321%。这表明在同一干旱环境条件下,A组种质资源叶片的保水能力要强于B组,即A组的抗旱性要强于B组,A组的文冠果种质资源更适宜在干旱地区种植。
表 6 2种类型文冠果叶片解剖结构的方差分析
Table 6. Variance of 2 types of X. sorbifolium leaf anatomical structure
类型 LT/μm TU/μm TL/μm TP/μm TS/μm LCT/μm VD/μm P/S/% PU/% PL/% PLCT/% CTR/% SR/% A组 188.117 16.820 13.879 88.037 64.863 5.618 13.818 140.499 9.011 7.540 3.060 46.987 35.053 B组 103.134 8.062 7.123 42.304 41.716 2.439 6.499 106.100 7.889 7.001 2.393 41.157 40.321 P 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.910 0.000 0.000 0.000 说明:缩写指代的指标见表1 -
叶片形态的变化是植物对干旱胁迫的重要响应机制之一。长期生长在干旱环境中的植物会形成多种抗旱、耐旱的叶片结构特征,其中叶片解剖结构的多项指标在植物抗旱性评价中广泛应用[16-18],如叶片厚度[19]、上表皮厚度、下表皮厚度[19]、栅海比[20-21]、组织结构紧密度[22]等都是评价植物抗旱性的主要指标,但使用单一指标评价植物的抗旱性具有片面性,对多项指标进行综合分析是准确评价植物种质资源抗旱性的关键。本研究基于13项叶片解剖结构指标对文冠果种质资源的抗旱性进行了评价,结果表明:文冠果不同种质资源的叶片解剖结构指标的变异范围较大,且指标之间具有显著相关性。为使综合评价结果更加客观合理,采用因子分析法从上述指标中提取出4个公因子:厚度指数因子、组织指数因子、表皮占比因子和蜡质层占比因子,其中载荷量较大的上/下表皮厚度、栅栏组织厚度和导管直径可作为评价文冠果抗旱性的主要指标。
基于因子分析的结果,结合聚类和隶属函数分析法将种质资源划分成为A组和B组。其中A组的90份种质资源具有较高的综合得分和隶属函数值,在表皮厚度、栅栏组织厚度、组织结构紧实度和导管直径等方面均具有较好的表现,即在干旱胁迫下,A组种质资源相比于B组具有较强的适应能力。叶片越厚,储水能力越强[22-24],栅栏组织厚实,细胞排列紧密能有效减少叶片水分的流失[25-26],而较大的导管直径可提高水分运输效率,使植株更好地适应干旱环境[27]。因子分析和隶属函数分析结果的一致性表明:因子分析法能够准确地评价并筛选出抗旱性文冠果种质资源,筛选出的A组种质资源能兼顾丰产和节水抗旱的优点,可为干旱地区提供适宜的种质材料。
叶片解剖结构特征是反映植物抗旱性和筛选抗旱型种质资源的一个重要方面[28-29]。抗旱性则是植物长期适应干旱环境而产生的一种复杂的性状,涉及植物的形态构造、生理生化反应等多方面[30-32]。在今后的研究中,可以综合考虑多种因素并结合生产实践来评价并筛选抗旱种质资源。
Leaf anatomical structure and evaluation of drought resistance of different germplasm resources of Xanthoceras sorbifolium
-
摘要:
目的 评价文冠果Xanthoceras sorbifolium不同种质资源在干旱地区的适应能力,为干旱、半干旱地区筛选抗旱文冠果种质提供依据。 方法 以内蒙古自治区通辽市文冠果种质资源圃内前期筛选的200份丰产型文冠果种质资源的叶片为材料,通过制作石蜡切片对叶片解剖结构进行观测,测定了叶片厚度、栅栏组织厚度、海绵组织厚度、蜡质层厚度等13个与文冠果抗旱性相关的指标,并基于因子分析、聚类分析和隶属函数分析的方法评价不同文冠果种质资源的抗旱性。 结果 文冠果叶片解剖结构指标之间具有显著的相关性,基于因子分析法提取了4个公因子,对变异的累计贡献率达89.99%,涵盖了13个指标的大部分信息,对文冠果抗旱性的影响程度依次为:F1 (厚度指数因子)>F2 (组织指数因子)>F3 (表皮占比因子)>F4 (蜡质层占比因子)。 结论 F1公因子中载荷较大的上/下表皮厚度、栅栏组织厚度和导管直径可作为评价文冠果抗旱性的主要指标。筛选出的90份抗旱型种质资源能兼顾丰产和节水抗旱的优点,可为干旱地区提供适宜的种质材料。图1表6参32 Abstract:Objective This study aims to evaluate the adaptability of different germplasm resources of Xanthoceras sorbifolium in arid areas, so as to provide a basis for screening drought-resistant germplasm resources for arid and semi-arid areas. Method The leaves of 200 high-yield X. sorbifolium germplasm resources were collected in Tongliao Germplasm Resources Nursery in Inner Mongolia Autonomous Region. The anatomical structure of leaves was observed by making paraffin section, and 13 indexes related to drought resistance were measured, including leaf thickness, palisade tissue thickness, sponge tissue thickness and wax layer thickness. The drought resistance of different germplasm resources was evaluated by factor analysis, cluster analysis and membership function analysis. Result The anatomical structure indexes of leaves showed significant correlation. Based on factor analysis, 4 common factors were extracted, and the cumulative contribution rate to variation was 89.99%, basically covering most of the information of the 13 indicators. The degree of influence on drought resistance ranging from large to small was as follows: F1 (thickness index factor), F2 (tissue index factor), F3 (epidermal proportion factor), and F4 (wax layer proportion factor). Conclusion The thickness of upper/lower epidermis, thickness of palisade tissue and vessel diameter, which are the most important factors in F1, can be used as the main indicators for drought resistance evaluation of X. sorbifolium. The selected 90 drought-resistant germplasm resources, with advantges of high-yield, water-saving and drought-risistant, could provide materials for the breeding of drought-resistant X. sorbifolium. [Ch, 1 fig. 6 tab. 32 ref.] -
图 1 2种类型文冠果种质资源的叶片解剖结构特征
Figure 1 Leaf anatomical structure characteristics of 2 types of X. sorbifolium
表 1 文冠果叶片解剖结构13个指标统计分析
Table 1. Statistics of 13 characters of leaf anatomical structure in X. sorbifolium
指标 最小
值/μm最大
值/μm均值/
μm标准
偏差变异系
数/%叶片厚度(LT) 60.22 270.04 140.90 50.99 36.19 上表皮厚度(TU) 3.97 24.10 11.95 5.22 43.65 下表皮厚度(TL) 4.26 21.36 10.13 4.04 39.87 栅栏组织厚度(TP) 22.34 182.31 62.66 28.22 45.04 海绵组织厚度(TS) 20.30 104.51 51.90 17.08 32.92 蜡质层厚度(LCT) 1.23 10.44 3.86 1.97 51.17 导管直径(VD) 4.03 25.01 9.74 4.41 45.31 栅海比(P/S) 35.26 263.46 121.69 38.93 32.00 上表皮占比(PU) 4.27 17.96 8.40 1.73 20.59 下表皮占比(PL) 4.19 21.34 7.26 1.84 25.28 蜡质层占比(PLCT) 1.25 5.43 2.68 0.74 27.47 组织结构紧密度(CTR) 23.08 93.76 43.79 8.22 18.78 组织结构疏松度(SR) 22.81 90.89 37.93 7.72 20.35 说明:样本数为200  下载: 导出CSV
下载: 导出CSV
表 2 200份文冠果叶片解剖结构13个性状的相关性分析
Table 2. Correlation between 13 traits of leaf anatomical structure in 200 X. sorbifolium samples
指标 LT TU TL TP TS LCT VD P/S PU PL PLCT CTR SR LT 1 TU 0.890** 1 TL 0.823** 0.881** 1 TP 0.903** 0.827** 0.754** 1 TS 0.832** 0.726** 0.734** 0.653** 1 LCT 0.832** 0.772** 0.769** 0.783** 0.719** 1 VD 0.851** 0.86** 0.829** 0.785** 0.711** 0.741** 1 P/S 0.393** 0.405** 0.313** 0.678** −0.075 0.366** 0.362** 1 PU 0.139* 0.552** 0.452** 0.187** 0.115 0.195** 0.334** 0.152** 1 PL −0.110 0.146* 0.447** −0.061 0.016 0.056 0.117 −0.064 0.595** 1 PLCT 0.214** 0.263** 0.357** 0.252** 0.233** 0.691** 0.261** 0.157* 0.221** 0.309** 1 CTR 0.230** 0.276** 0.251** 0.602** −0.021 0.266** 0.248** 0.838** 0.227** 0.137 0.213** 1 SR −0.394** −0.354** −0.220** −0.493** 0.154* −0.281** −0.323** −0.792** 0.010 0.293** 0.018 −0.412** 1 说明:缩写指代的指标见表1。*表示显著相关 (P<0.05);**表示极显著相关 (P<0.01)
下载: 导出CSV
表 3 主要评价指标贡献率
Table 3. Total variance contribution
指标 初始特征值 旋转后主因子 特征根 贡献
率/%累积贡
献率/%特征根 贡献
率/%累积贡
献率/%LT 6.554 50.418 50.418 5.480 42.154 42.154 TU 2.303 17.714 68.132 2.899 22.300 64.454 TL 1.799 13.837 81.969 1.935 14.885 79.339 TP 1.043 8.019 89.988 1.384 10.650 89.988 TS 0.608 4.677 94.665 LCT 0.409 3.145 97.811 VD 0.179 1.377 99.188 P/S 0.037 0.288 99.476 PU 0.023 0.176 99.652 PL 0.018 0.137 99.789 PLCT 0.014 0.109 99.898 CTR 0.009 0.066 99.964 SR 0.005 0.036 100 说明:缩写指代的指标见表1
下载: 导出CSV
表 4 旋转后的因子载荷及得分系数
Table 4. Factor loadings and factor scores coefficient after rotated
指标 变量 因子载荷 因子得分 F1 F2 F3 F4 F1 F2 F3 F4 LT X1 0.961 0.225 −0.105 0.051 0.211 −0.017 −0.115 −0.081 TU X2 0.900 0.250 0.282 0.003 0.179 0.006 0.116 −0.172 TL X3 0.848 0.137 0.397 0.157 0.151 −0.035 0.160 −0.044 TP X4 0.800 0.537 −0.025 0.122 0.123 0.126 −0.068 −0.012 TS X5 0.913 −0.263 −0.035 0.139 0.236 −0.209 −0.092 0.001 LCT X6 0.788 0.194 −0.021 0.558 0.095 −0.018 −0.133 0.379 VD X7 0.880 0.207 0.155 0.045 0.180 −0.011 0.038 −0.113 P/S X8 0.179 0.970 0.025 0.074 −0.074 0.370 0.006 0.020 PU X9 0.225 0.115 0.868 −0.039 0.003 0.034 0.490 −0.197 PL X10 −0.018 −0.115 0.890 0.229 −0.072 −0.037 0.473 0.075 PLCT X11 0.182 0.063 0.168 0.942 −0.106 −0.008 −0.052 0.779 CTR X12 0.062 0.827 0.208 0.202 −0.115 0.326 0.096 0.125 SR X13 −0.196 −0.821 0.207 0.148 0.009 −0.309 0.096 0.142 说明:缩写指代的指标见表1
下载: 导出CSV
表 5 2种类型文冠果叶片解剖结构的隶属函数分析
Table 5. Subordinative function of 2 types of X. sorbifolium leaf anatomical structure
类型 隶属函数值 综合评价值 综合得分F LT TU TL TP TS LCT VD P/S PU PL PLCT CTR SR A组 0.610 0.638 0.563 0.411 0.471 0.476 0.467 0.461 0.346 0.195 0.433 0.338 0.820 0.479 0.610 B组 0.205 0.203 0.168 0.125 0.254 0.131 0.118 0.310 0.265 0.164 0.274 0.256 0.257 0.210 −0.449 说明:缩写指代的指标见表1
下载: 导出CSV
表 6 2种类型文冠果叶片解剖结构的方差分析
Table 6. Variance of 2 types of X. sorbifolium leaf anatomical structure
类型 LT/μm TU/μm TL/μm TP/μm TS/μm LCT/μm VD/μm P/S/% PU/% PL/% PLCT/% CTR/% SR/% A组 188.117 16.820 13.879 88.037 64.863 5.618 13.818 140.499 9.011 7.540 3.060 46.987 35.053 B组 103.134 8.062 7.123 42.304 41.716 2.439 6.499 106.100 7.889 7.001 2.393 41.157 40.321 P 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.910 0.000 0.000 0.000 说明:缩写指代的指标见表1
下载: 导出CSV
-
[1] YU Haiyan, FAN Siqi, BI Quanxin, et al. Seed morphology, oil content and fatty acid composition variability assessment in yellow horn (Xanthoceras sorbifolium Bunge) germplasm for optimum biodiesel production [J]. Industrial Crops and Products, 2017, 97: 425 − 430. [2] 麻云霞. 文冠果种子特性变异及优良砧用种源选择[D]. 呼和浩特: 内蒙古农业大学, 2021. MA Yunxia. Variation of Xanthoceras sorbifolium Bunge Seed Characteristics and Selection of Superior Rootstock Provenance[D]. Huhhot: Inner Mongolia Agricultural University, 2021. [3] 于海燕, 胡潇予, 何春霞, 等. 文冠果不同种源叶片结构对水分胁迫的差异性响应[J]. 北京林业大学学报, 2019, 41(1): 57 − 63. YU Haiyan, HU Xiaoyu, HE Chunxia, et al. Differential response of water stress on leaf morphological anatomical structures of varied provenances Xanthocera sorbifolium [J]. Journal of Beijing Forestry University, 2019, 41(1): 57 − 63. [4] 田梦妮, 毕泉鑫, 贺晓辉, 等. 文冠果叶片解剖结构的抗旱性分析与评价[J]. 分子植物育种, 2019, 17(24): 8261 − 8270. TIAN Mengni, BI Quanxin, HE Xiaohui, et al. Analysis and evaluation of drought resistance of Xanthoceras sorbifolium leaf anatomical structure [J]. Molecular Plant Breeding, 2019, 17(24): 8261 − 8270. [5] 赵阳, 毕泉鑫, 句娇, 等. 文冠果种子及苗期生长性状地理种源变异[J]. 林业科学研究, 2019, 32(1): 163 − 171. ZHAO Yang, BI Quanxin, JU Jiao, et al. Geographic variation of seed and seedling growing traits in Xanthoceras sorbifolium [J]. Forest Research, 2019, 32(1): 163 − 171. [6] 李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报, 2005, 22(增刊 1): 118 − 127. LI Fanglan, BAO Weikai. Responses of the morphological and anatomical structure of the plant leaf to environmental change [J]. Chinese Bulletin of Botany, 2005, 22(suppl 1): 118 − 127. [7] 岑湘涛, 沈伟, 牛俊乐, 等. 基于植物叶片解剖结构的抗逆性评价研究进展[J]. 北方园艺, 2021(18): 140 − 147. CEN Xiangtao, SHEN Wei, NIU Junle, et al. Research progress of stress resistance evaluation based on the anatomy of plant leaves [J]. Northern Horticulture, 2021(18): 140 − 147. [8] 赵雪, 张秀珍, 牟洪香, 等. 文冠果幼苗叶片解剖结构和光合作用对干旱胁迫的响应[J]. 北方园艺, 2017(13): 38 − 44. ZHAO Xue, ZHANG Xiuzhen, MU Hongxiang, et al. Effect of drought stress on anatomical structure and photosynthesis of Xanthoceras sorbifolia leaves in seedling [J]. Northern Horticulture, 2017(13): 38 − 44. [9] 王孟珂, 田梦妮, 毕泉鑫, 等. 基于气孔性状的文冠果种质资源抗旱性评价及抗旱资源筛选[J]. 植物研究, 2021, 41(6): 957 − 964. WANG Mengke, TIAN Mengni, BI Quanxin, et al. Evaluation of drought tolerance based on stomatal characters and selection of germplasm resources from Xanthoceras sorbifolia [J]. Bulletin of Botanical Research, 2021, 41(6): 957 − 964. [10] 马小芬, 王兴芳, 李强, 等. 不同种源地文冠果叶片解剖结构比较及抗旱性分析[J]. 干旱区资源与环境, 2013, 27(6): 92 − 96. MA Xiaofen, WANG Xingfang, LI Qiang, et al. The analysis of drought resistance and the comparison of anatomical structuresof the leave of Xanthoceras sorbifolia Bunge introduced from different regions [J]. Journal of Arid Land Resources and Environment, 2013, 27(6): 92 − 96. [11] 韩永亮, 李世云, 路正营, 等. 62份陆地棉种质资源苗期抗旱性综合评价及耐旱种质筛选[J]. 干旱地区农业研究, 2021, 39(6): 28 − 38. HAN Yongliang, LI Shiyun, LU Zhengying, et al. Comprehensive identification and selection of drought resistance of 62 cotton varieties (lines) at cotton seedling stage [J]. Agricultural Research in the Arid Areas, 2021, 39(6): 28 − 38. [12] 宋俊乔. 水稻叶片形态、解剖结构与抗旱性的关系研究[D]. 武汉: 华中农业大学, 2010. SONG Junqiao. Research of the Relationship between Morphologic and Anatomic Structure and Drought Resistance of Rice[D]. Wuhan: Huazhong Agricultural University, 2010. [13] 赵永超. 基于因子分析和聚类分析的马铃薯节水灌溉绩效评价研究[D]. 昆明: 云南师范大学, 2021. ZHAO Yongchao. Study on Performance Evaluation of Potato Water-saving Irrigation Based on Factor Analysis and Cluster Analysis: A Case of Wenshan Prefecture in Yunnan Province[D]. Kunming: Yunnan Normal University, 2021. [14] 高源, 王霞, 高树仁, 等. 基于隶属函数法分析80份玉米自交系萌芽期的抗旱性[J/OL]. 分子植物育种, 2021-09-28[2022-01-28]. http://kns.cnki.net/kcms/detail/46.1068.S.20210927.2137.006.html. GAO Yuan, WANG Xia, GAO Shuren, et al. Analysis of drought resistance of 80 maize inbred lines based on membership function at germination stage[J/OL]. Molecular Plant Breeding, 2021-09-28[2022-01-28]. http://kns.cnki.net/kcms/detail/46.1068.S.20210927.2137.006.html. [15] 马胜, 齐恩芳, 文国宏, 等. 基于叶片显微结构综合评价马铃薯不同品种的抗旱性[J]. 中国马铃薯, 2021, 35(6): 500 − 506. MA Sheng, QI Enfang, WEN Guohong, et al. Comprehensive evaluation of drought resistance of different potato varieties based on leaf microstructure [J]. Chinese Potato Journal, 2021, 35(6): 500 − 506. [16] 郭燕, 张树航, 李颖, 等. 中国板栗238份品种(系)叶片形态、解剖结构及其抗旱性评价[J]. 园艺学报, 2020, 47(6): 1033 − 1046. GUO Yan, ZHANG Shuhang, LI Ying, et al. Studies on the leaf morphology, anatomical structure and drought resistance evaluation of 238 Chinese chestnut varieties (strains) [J]. Acta Horticulturae Sinica, 2020, 47(6): 1033 − 1046. [17] 高晓宁, 赵冰, 刘旭梅, 等. 4个杜鹃花品种对干旱胁迫的生理响应及抗旱性评价[J]. 浙江农林大学学报, 2017, 34(4): 597 − 607. GAO Xiaoning, ZHAO Bing, LIU Xiaomei, et al. Physiological response to drought stress and drought resistance evaluation of four Rhododendron cultivars [J]. Journal of Zhejiang A&F University, 2017, 34(4): 597 − 607. [18] BRITO C, DINIS L T, MOUTINHO-PEREIRA J, et al. Drought stress effects and olive tree acclimation under a changing climate [J]. Plants, 2019, 8(7): 232. [19] 赵君茹, 朱周俊, 肖诗鑫, 等. 高州油茶叶片解剖结构及抗旱性评价[J]. 分子植物育种, 2022, 20(16): 5435 − 5443. ZHAO Junru, ZHU Zhoujun, XIAO Shixin, et al. Study on drought resistance of leaf anatomical structure of Camellia gauchowensis Chang clone [J]. Molecular Plant Breeding, 2022, 20(16): 5435 − 5443. [20] 朱凯琳. 花楸属植物叶解剖特征及其环境适应性研究[D]. 南京: 南京林业大学, 2021. ZHU Kailin. Leaf Anatomical Characteristics and Environmental Adaptability of Sorbus Species[D]. Nanjing: Nanjing Forestry University, 2021. [21] 王烟霞, 樊军锋, 程玮哲, 等. 基于叶片解剖结构的12个杨树无性系抗旱性分析[J]. 西北农林科技大学学报(自然科学版), 2021, 49(11): 147 − 154. WANG Yanxia, FAN Junfeng, CHENG Weizhe, et al. Drought resistances analysis of 12 poplar clones based on leaf anatomical structures [J]. Journal of Northwest A&F University (Natural Science Edition), 2021, 49(11): 147 − 154. [22] 何小三, 王玉娟, 徐林初, 等. 干旱胁迫对不同油茶品种叶片解剖结构的影响[J]. 中南林业科技大学学报, 2020, 40(10): 1 − 17. HE Xiaosan, WANG Yujuan, XU Linchu, et al. Effects of drought stress on leaf anatomical structure of different Camellia oleifera cultivars [J]. Journal of Central South University of Forestry &Technology, 2020, 40(10): 1 − 17. [23] 刘光辉, 陈全家, 吴鹏昊, 等. 棉花花铃期抗旱性综合评价及指标筛选[J]. 植物遗传资源学报, 2016, 17(1): 53 − 62, 69. LIU Guanghui, CHEN Quanjia, WU Penghao, et al. Screening and comprehensive evaluation of drought resistance indices of cotton at blossming and boll-forming Stages [J]. Journal of Plant Genetic Resources, 2016, 17(1): 53 − 62, 69. [24] VELIKOVA V, ARENA C, IZZO L G, et al. Functional and structural leaf plasticity determine photosynthetic performances during drought stress and recovery in two Platanus orientalis populations from contrasting habitats[J/OL]. International Journal of Molecular Sciences, 2020, 21(11): 3912[2022-01-30]. doi: 10.3390/ijms21113912. [25] 吴丽君, 李志辉, 杨模华, 等. 赤皮青冈幼苗叶片解剖结构对干旱胁迫的响应[J]. 应用生态学报, 2015, 26(12): 3619 − 3626. WU Lijun, LI Zhihui, YANG Mohua, et al. Response of leaf anatomical characteristics of Cyclobalanopsis gilva seedlings to drought stress [J]. Chinese Journal of Applied Ecology, 2015, 26(12): 3619 − 3626. [26] MANSOOR U, FATIMA S, HAMEED M, et al. Structural modifications for drought tolerance in stem and leaves of Cenchrus ciliaris L. ecotypes from the Cholistan Desert[J/OL]. Flora, 2019, 261: 151485[2022-01-30]. doi: 10.1016/j.flora.2019.151485. [27] 李昭良. 不同桑树品种茎叶解剖结构的耐旱性特征研究[D]. 杨凌: 西北农林科技大学, 2014. LI Zhaoliang. Studies on the Drought Resistance Characteristics of Anatomical Structure of the Leaves and Stems of Different Mulberry Varieties[D]. Yangling: Northwest A&F University, 2014. [28] HU Yang, YANG Lu, GAO Chao, et al. A comparative study on the leaf anatomical structure of Camellia oleifera in a low-hot valley area in Guizhou Province, China[J/OL]. PLoS One, 2022, 17(1): e0262509[2022-01-30]. doi: 10.1371/journal.pone.0261942. [29] GONG Zhizhong, XIONG Liming, SHI Huazhong, et al. Plant abiotic stress response and nutrient use efficiency [J]. Science China-Life Sciences, 2020, 63(5): 663 − 674. [30] BACELAR E A, CORREIA C M, MOUTINHO-PEREIRA J M, et al. Sclerophylly and leaf anatomical traits of five field-grown olive cultivars growing under drought conditions [J]. Tree Physiology, 2004, 24(2): 233 − 239. [31] 张刚. 文冠果的抗旱性研究[D]. 杨凌: 西北农林科技大学, 2014. ZHANG Gang. Studies on Drought Resistance of Xanthoceras sorbifolia Seedlings[D]. Yangling: Northwest A&F University, 2014. [32] 刘彬, 麻文俊, 王军辉, 等. 基于叶片解剖结构的砂生槐群体抗旱性评价[J]. 植物研究, 2017, 37(3): 325 − 333. LIU Bin, MA Wenjun, WANG Junhui, et al. Drought-resistance of Sophora moorcroftiana from different population from leaf anatomical structure [J]. Bulletin of Botanical Research, 2017, 37(3): 325 − 333. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220202
 点击查看大图
点击查看大图

计量
- 文章访问数: 2438
- HTML全文浏览量: 432
- PDF下载量: 51
- 被引次数: 0



