




-
百山祖冷杉Abies beshanzuensis隶属于松科Pinaceae冷杉属Abies,于1963年被首次发现,并于1976年被正式鉴定及命名为新物种,目前仅自然分布于浙江省庆元县百山祖海拔约1 750 m的区域[1],被列为国家一级重点保护野生植物[2],且被列为全球最濒危的12种植物之一[2–3]。该物种起源古老,是低纬度地区的冷杉属等高寒植物在第四纪冰川末期从低海拔向高海拔山体退缩的佐证,自然生长的成熟个体现仅存3株,被誉为“植物活化石”。由于其环境适应能力弱且自然繁殖能力低,该物种长期处于极度濒危的状态[4]。该物种对于这一地区的植物区系发生和演变,以及植物学、生态学、地理学和气候学等具有很高的科研价值[5]。
生长量是反映植物生长状况最直接的指标之一,被用来直观地显示环境对植物的生存状态造成的影响[6–7]。叶绿素荧光参数是一种非破坏性的测量技术,专门用于评估光系统Ⅱ(PSⅡ)的活性,能够在不破坏叶片细胞结构的前提下,快速获取相关参数信息。此技术目前已在植物生理学研究中得到广泛应用[8–9],尤其在深入探究光合作用机制、植物对环境变化的适应性响应、遗传多样性分析以及生态系统多样性评估等方面发挥了重要作用[10–11]。因此,近年来,叶绿素荧光技术已在多个领域广泛应用,包括高海拔植物光合适应性的研究[12−14]、植物在不同胁迫条件下光合生理响应的探讨[15−17]、不同物种间光合生理特性差异性的分析[18–19]、遮阴对植物光合特性影响的研究[20–21],以及从光合作用角度探讨濒危物种致濒机制的解析等[22−24]。为了改善百山祖冷杉的濒危状况,针对其濒危机制及抢救性保护措施展开了深入研究。近年来,除了传统的嫁接繁育技术,一些利用转录组学、代谢组学等手段对其遗传多样性、濒危机制及保护策略的研究也取得了新的认识[25–26]。选择1年生小枝和叶片作为研究对象的主要原因:其一、叶片在成熟后光合能力强,叶绿素荧光参数相对稳定和可靠;其二、取材量相对较少,对植株损坏较小,尤其是原生母树;其三、2年生及以上年龄枝条上的叶片可能因各种原因引起落叶或损坏,对统计数据会有影响。因此,本研究通过对比自然条件下的百山祖冷杉原生母树与嫁接树1年生枝叶的生长指标,并在主要生长季节持续监测其叶片的叶绿素荧光参数,旨在深入揭示百山祖冷杉原生母树与嫁接树在生长状态及叶绿素荧光参数的差异,进而为百山祖冷杉的精准保护策略提供科学依据。
-
采样点位于浙江省庆元县钱江源-百山祖国家公园,原生母树采样点位于海拔约1 750 m的山地林中,无遮阴、高光强,土壤瘠薄;嫁接树采样点位于海拔约1 550 m的百山祖国家公园庆元保护中心百管站苗圃内(嫁接株不能种植在原生母树周围,否则花粉会对原生母树所结种子产生影响),生境条件相对优越,有大树和西面房屋部分遮阴,土壤较肥厚。选择生长正常的原生母树(树龄约170 a)和以日本冷杉 Abies firma为砧木的嫁接树(树龄约42 a)的健康1年生枝条。于2023年生长季4—9月,每月进行1次采样。
-
2023年8月,选取原生母树与嫁接树长势良好且具有代表性的1年生枝条,使用游标卡尺(精度0.01 cm)测量枝条上叶片的长度和宽度、小枝的长度和直径。使用分析天平(精度0.000 1 g)称量1年生枝条的鲜质量和枝条上叶片鲜质量,并对叶片数量进行计数。均采样3次进行测量,取其平均值。
-
叶绿素荧光测定采用沈宗根等的方法[27],使用德国Mini-PAM便携式调制叶绿素荧光仪测定叶片叶绿素荧光参数。选取百山祖冷杉原生母树与嫁接树1年生枝条上的叶片,将叶片置于黑暗处进行暗处理30 min后,激发光的光强(PAR)梯度设置为0、81、130、195、268、396、540、816、1 190 μmol·m−2·s−1,依次测定叶片的光合量子产量(Y)、电子传递速率(RET)、光化学猝灭系数(CPQ)和非光化学猝灭系数(CNPQ),重复3次。测定时始终使叶片垂直于入射光,为减少误差,不同处理的叶片交叉测定。4—9月每月测定1次,共计6次。
-
使用软件Excel 2019和SPSS 26对数据进行统计分析和方差分析,采用Pearson相关系数对各项指标进行相关性分析,使用软件GraphPad Prism 9.5进行作图和分析。
-
如表1所示:百山祖冷杉原生母树的1年生枝叶生长量指标小枝长、叶片数、叶长、小枝直径、叶鲜质量、小枝鲜质量均显著小于嫁接树(P<0.05)。8月,原生母树的小枝长为56.39 mm±2.66 mm,而嫁接树的小枝长为78.66 mm±5.87 mm;原生母树的小枝直径为3.20 mm±0.15 mm,嫁接树的小枝直径为3.71 mm±0.21 mm;原生母树叶片数为67.33片±2.91片,嫁接树叶片数为79.33片±2.40片;母树叶长为33.76 mm±2.56 mm;嫁接树叶长为45.85 mm±0.38 mm;原生母树叶鲜质量为2.32 g±0.25 g,嫁接树叶鲜质量为4.93 g±0.21 g;原生母树小枝鲜质量为0.36 g±0.03 g,嫁接树小枝鲜质量为0.69 g±0.02 g。而叶宽在原生母树和嫁接树之间无显著差异。
表 1 百山祖冷杉原生母树与嫁接树1年生枝叶的生长量
Table 1. Growth of current-year branches and leaves of the original mother and grafted trees of A. beshanzuensis
样树 小枝长/mm 小枝直径/mm 叶片数/片 叶长/mm 叶宽/mm 叶鲜质量/g 小枝鲜质量/g 原生母树 56.39±2.66 b 3.20±0.15 b 67.33±2.91 b 33.76±2.56 b 3.49±0.22 a 2.32±0.25 b 0.36±0.03 b 嫁接树 78.66±5.87 a 3.71±0.21 a 79.33±2.40 a 45.85±0.38 a 3.93±0.45 a 4.93±0.21 a 0.69±0.02 a 说明:数据为平均值±标准误,同列不同小写字母表示差异显著(P<0.05,独立样本t检验)。 -
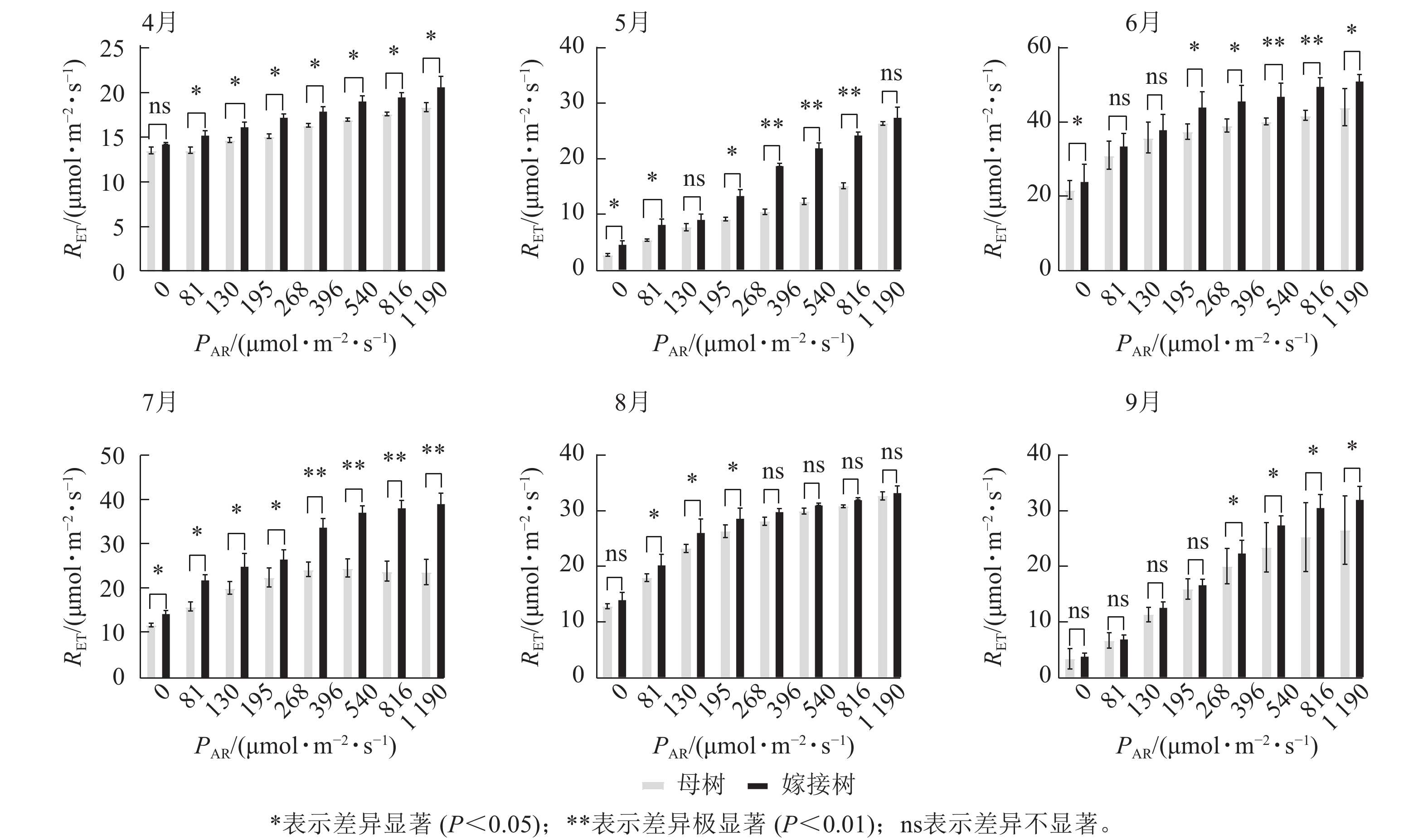
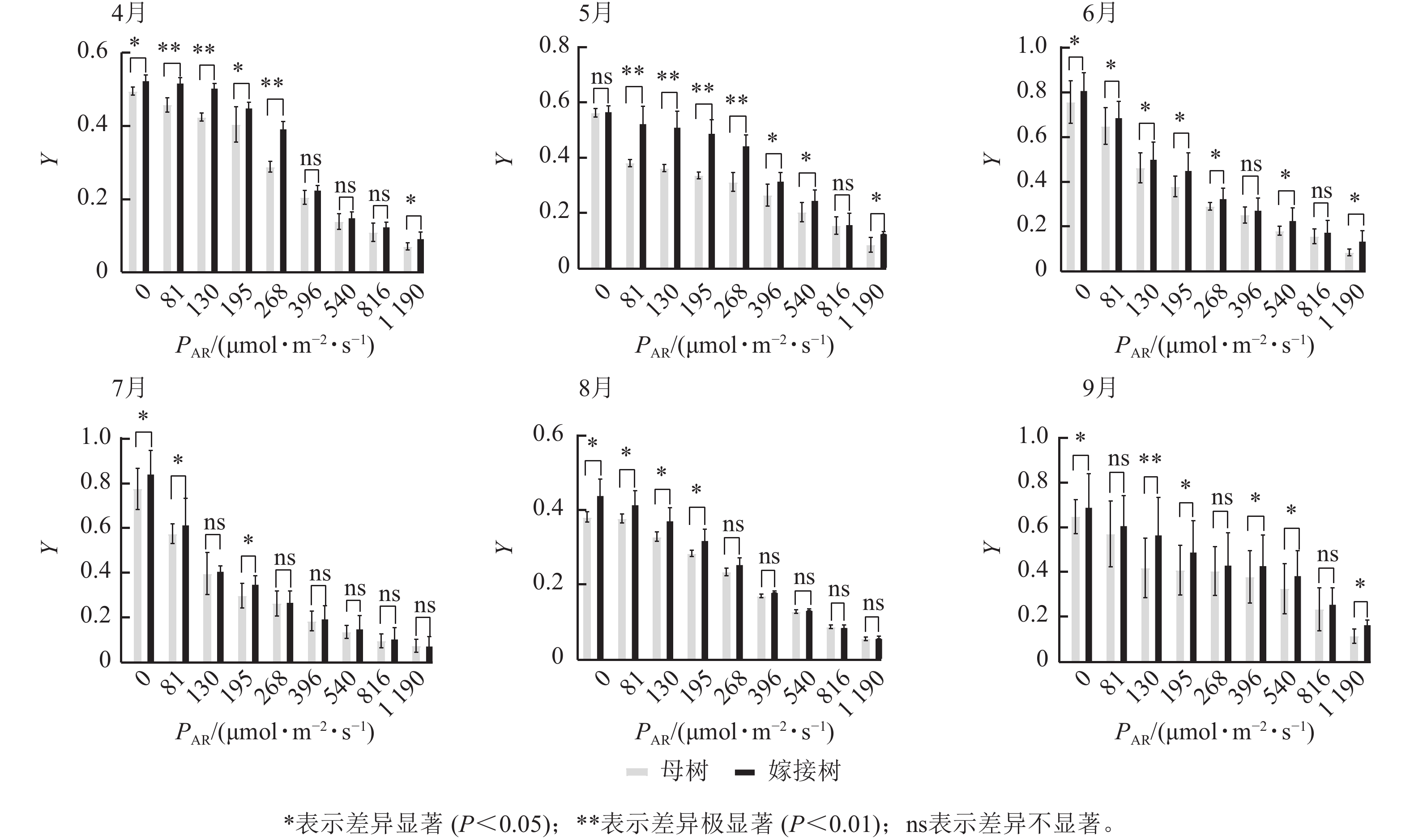
6个月的持续监测(图1)表明:百山祖冷杉原生母树与嫁接树的叶片光合量子产量均随光强的增高而降低。在整个监测期间,同一光强下原生母树的光合量子产量均低于嫁接树:4月,在0、195、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在81、130、268 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);5月,在396、540、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在81~268 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);6月,在0、81、130、195、268、540、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05);7月,在0、81、195 μmol·m−2·s−1等光强条件下差异显著(P<0.05);8月,0~195 μmol·m−2·s−1等光强条件下差异显著;9月,在0、195、396、540、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在130 μmol·m−2·s−1光强条件下差异极显著(P<0.01)。这表明原生母树的光能捕获效率低于嫁接树。图2显示:原生母树与嫁接树的电子传递速率均随着光强的增高而增加,表明百山祖冷杉具有较强的光合能力。然而,在同一光强下,原生母树的电子传递速率均低于嫁接树:4月,除81 μmol·m−2·s−1光强条件外,其余光强条件下均差异显著(P<0.05);5月,在81、130、268 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在396、540、816 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);6月,在81、268、396、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05),在540、816 μmol·m−2·s−1等光强条件下差异极显著(P<0.05);7月,在81~268 μmol·m−2·s−1等光强条件下差异显著(P<0.05),≥396 μmol·m−2·s−1光强条件下差异极显著(P<0.01);8月,仅在130~268 μmol·m−2·s−1光强条件下差异显著(P<0.05);9月,≥396 μmol·m−2·s−1等光强条件下差异显著(P<0.05)。这表明原生母树的电子传递速率较嫁接树弱。综合以上2个指标发现:原生母树的光合能力显著低于嫁接树(P<0.01)。
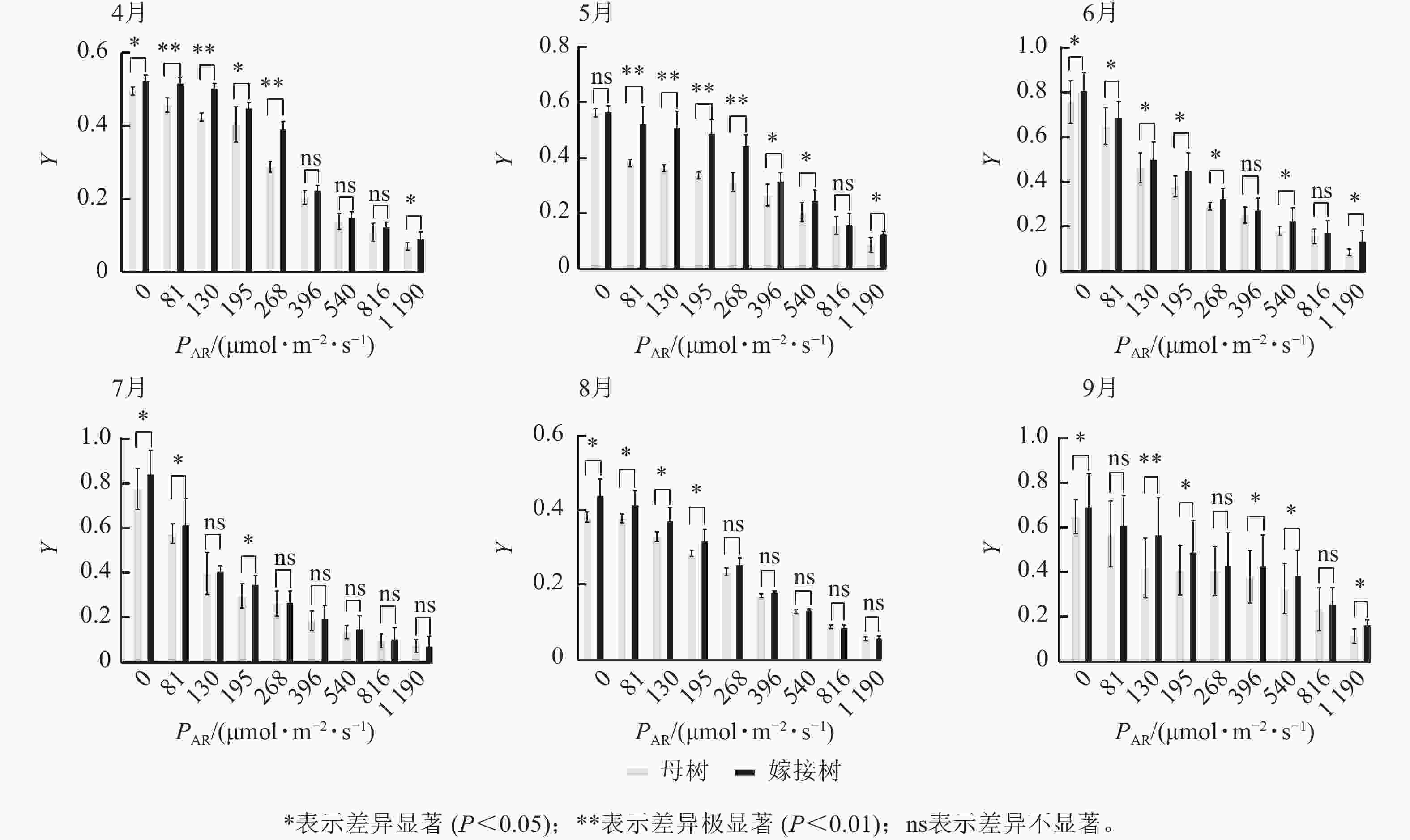
图 1 4—9月百山祖冷杉原生母树与嫁接树的光合量子产量(Y)对光强(PAR)的响应比较
Figure 1. Comparison of the response curves of photosynthetic quantum yield (Y) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
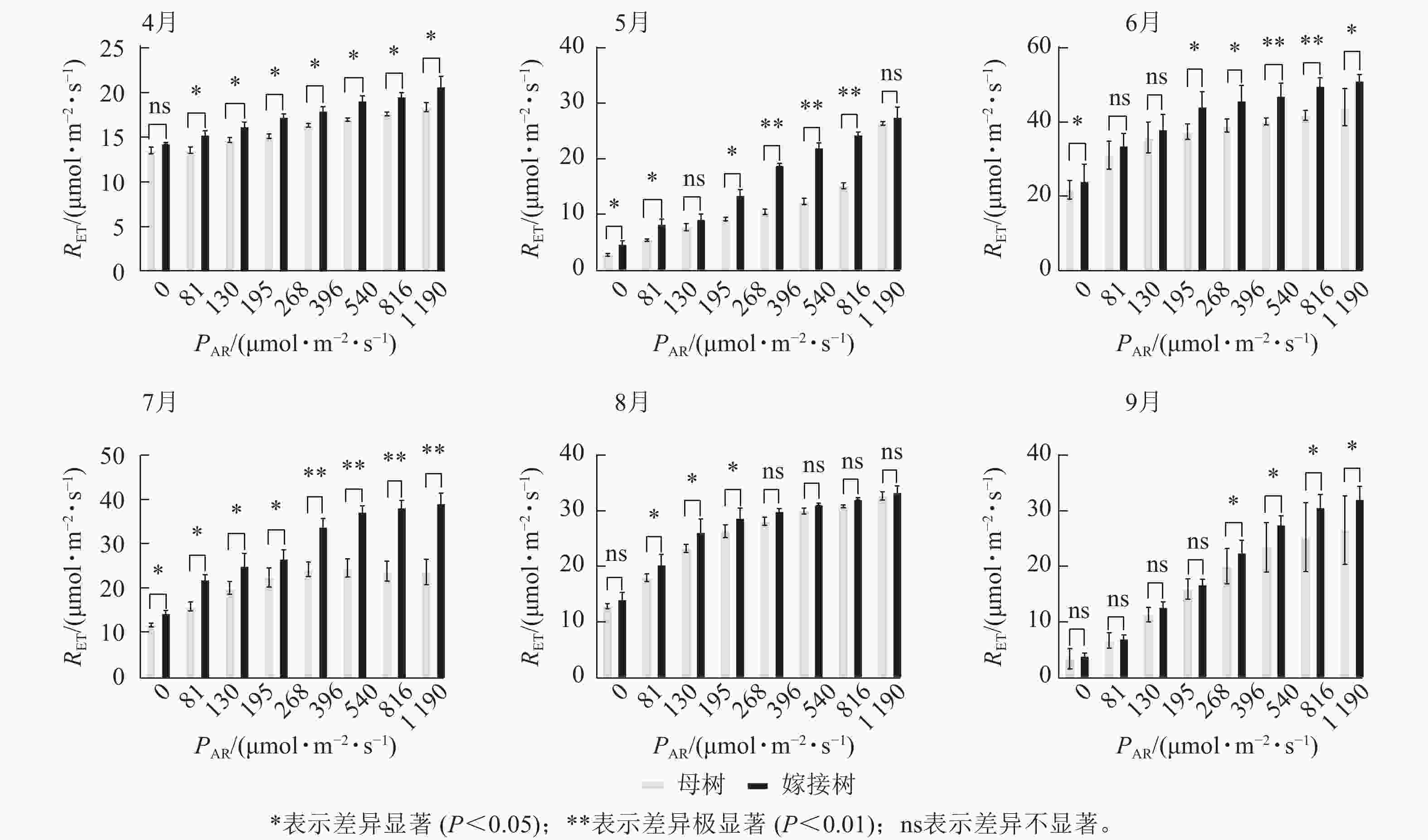
图 2 4—9月百山祖冷杉原生母树与嫁接树的电子传递速率(RET)对光强(PAR)的响应比较
Figure 2. Comparison of the response curves of electron transport rate (RET) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
-
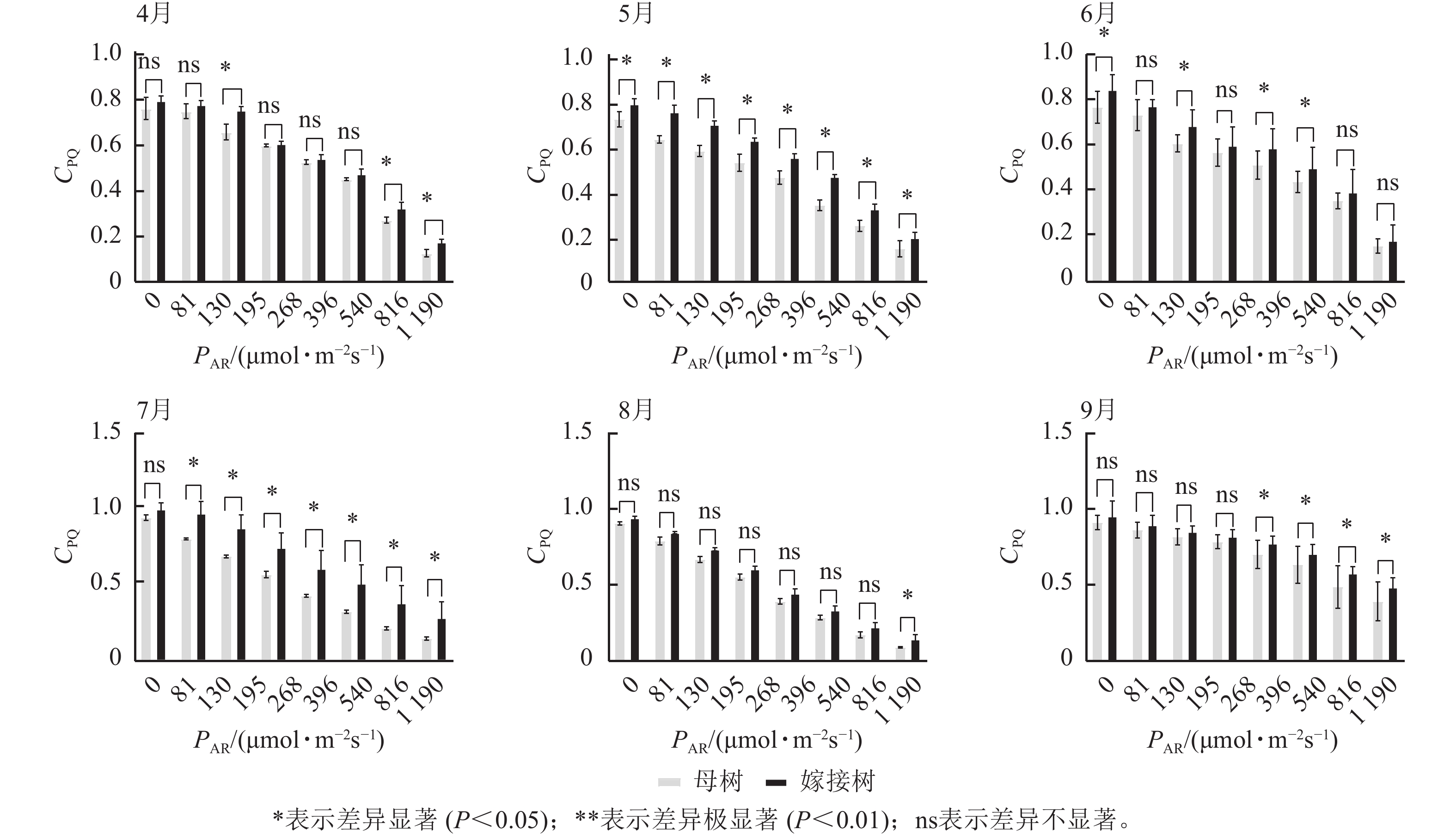
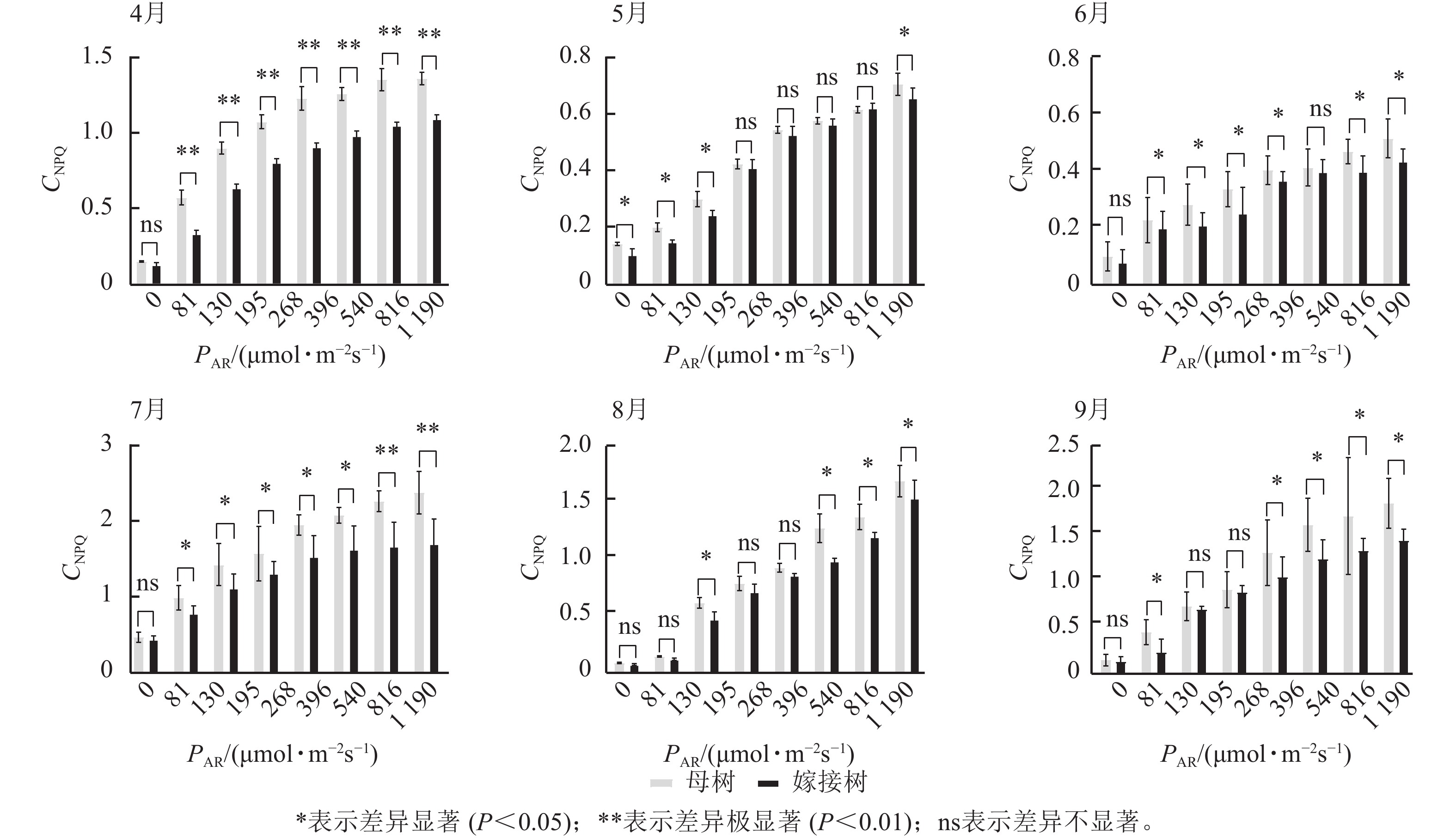
图3显示:原生母树与嫁接树的光化学猝灭系数随着光合有效辐射的升高而降低,且在同一光强下,原生母树的光化学猝灭系数均低于嫁接树:4月,在195、816、
1190 μmol·m−2·s−1等光强条件下差异显著(P<0.05);5月,所有实验的光强条件下均差异显著(P<0.05);6月,在81、195、396、540 μmol·m−2·s−1等光强条件下差异显著(P<0.05);7月,除81 μmol·m−2·s−1外,其他光强下均差异显著(P<0.05);8月,仅1190 μmol·m−2·s−1光强条件下差异显著(P<0.05);9月,≥396 μmol·m−2·s−1光强条件均差异显著(P<0.05)。图4显示:原生母树与嫁接树的非光化学猝灭系数随着光强的升高而升高,光合作用系统完整,并未因受到环境胁迫而破坏。且同一光强下原生母树的非光化学猝灭系数均高于嫁接树:4月,除81 μmol·m−2·s−1外,其他光强下均差异极显著(P<0.01); 5月,在81、130、195、1 190 μmol·m−2·s−1等光强条件下差异显著(P<0.05);6月,除81、816 μmol·m−2·s−1外,其他光强下均差异显著(P<0.05);7月,在130~540 μmol·m−2·s−1光强条件下差异显著(P<0.05),在816、1190 μmol·m−2·s−1等光强条件下差异极显著(P<0.01);8月,在195和≥540 μmol·m−2·s−1等光强条件下差异显著(P<0.05),其余光强条件下无显著差异;9月,除81、195、268 μmol·m−2·s−1等光强条件下无显著差异外,其他光强条件下均具显著差异(P<0.05)。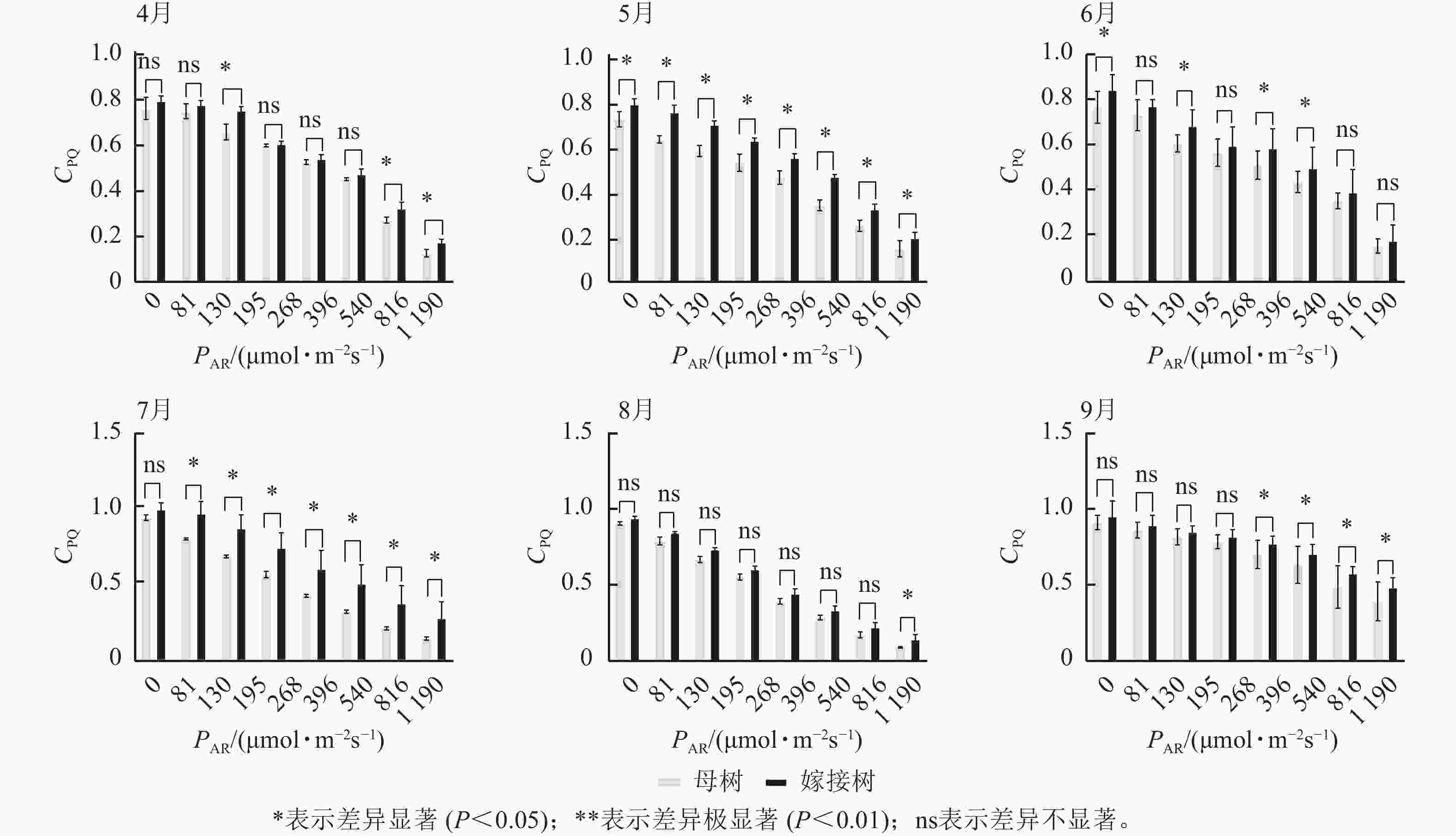
图 3 4—9月百山祖冷杉原生母树与嫁接树的光化学猝灭系数(CPQ)和对光强(PAR)的响应比较
Figure 3. Comparison of the response curves of photochemical quenching coefficent (CPQ) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
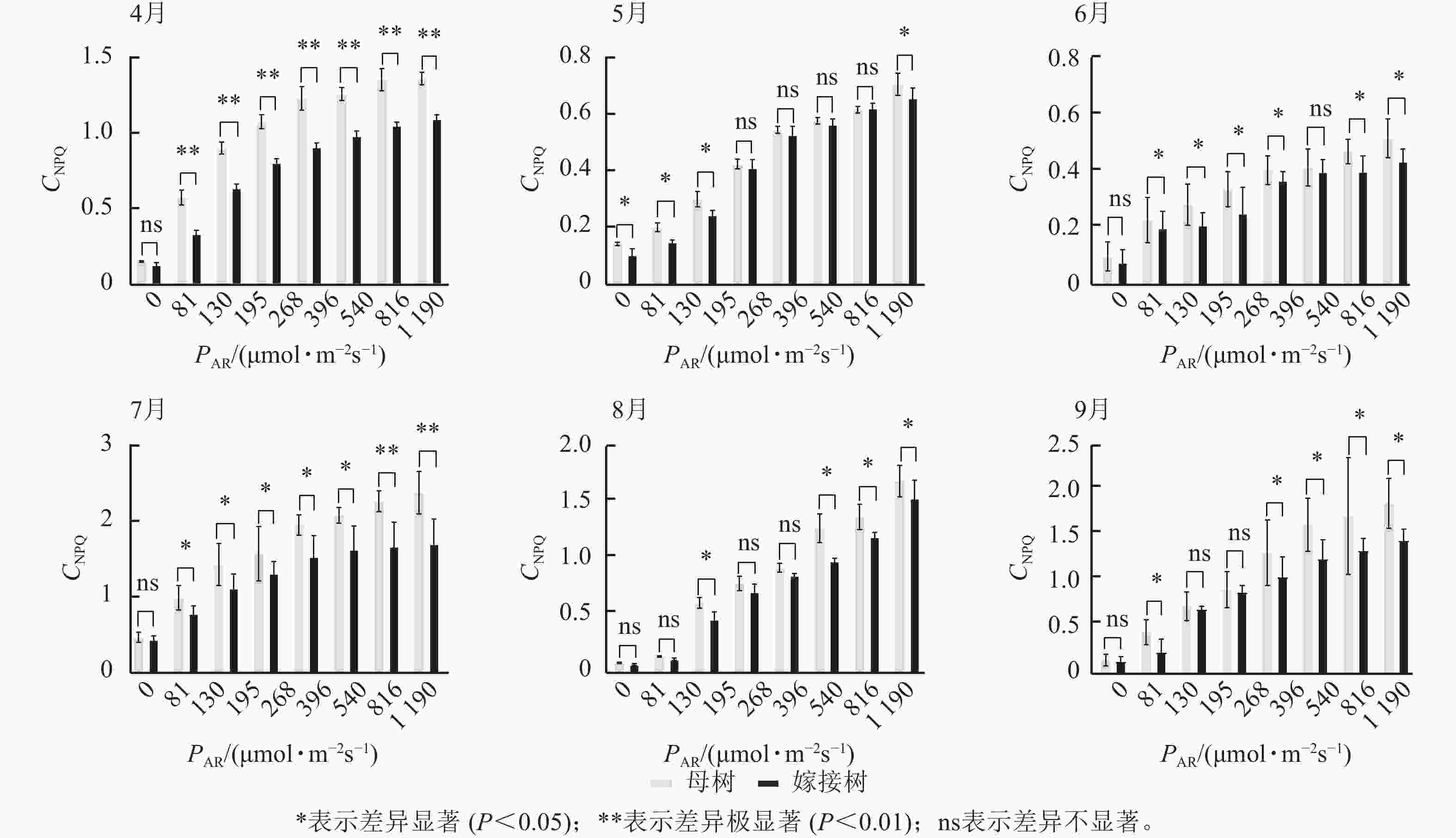
图 4 4—9月百山祖冷杉原生母树与嫁接树的非光化学猝灭系数(CNPQ)和对光强(PAR)的响应比较
Figure 4. Comparison of the response curves of photochemical quenching coefficent (CNPQ) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
-
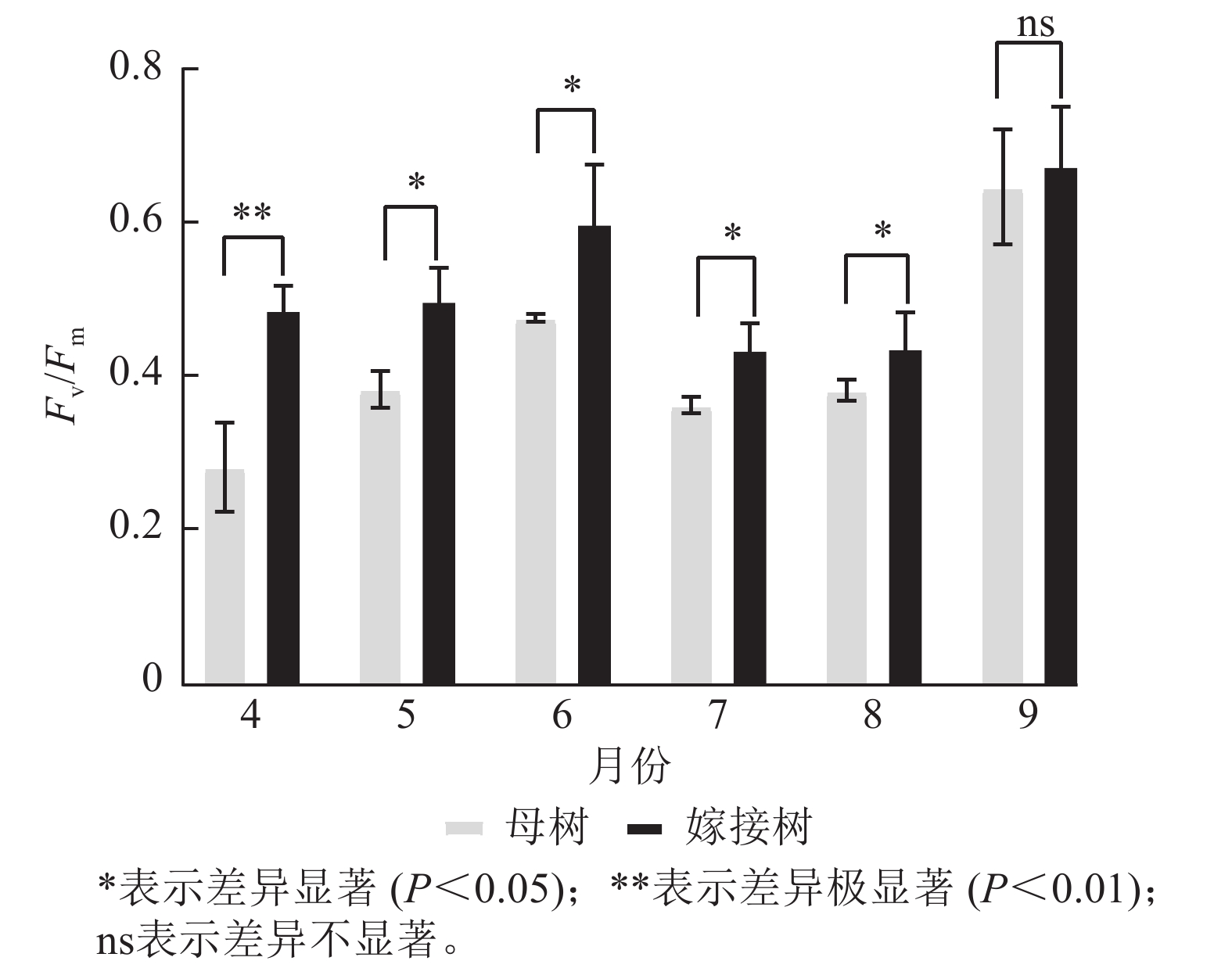
图5显示:原生母树与嫁接树的最大光化学量子产量均在4—6月呈上升趋势:在6—8月呈下降趋势,8—9月开始上升。且嫁接树的最大光化学量子产量均高于原生母树,4月差异极显著(P<0.01),5—8月则差异显著(P<0.05)。
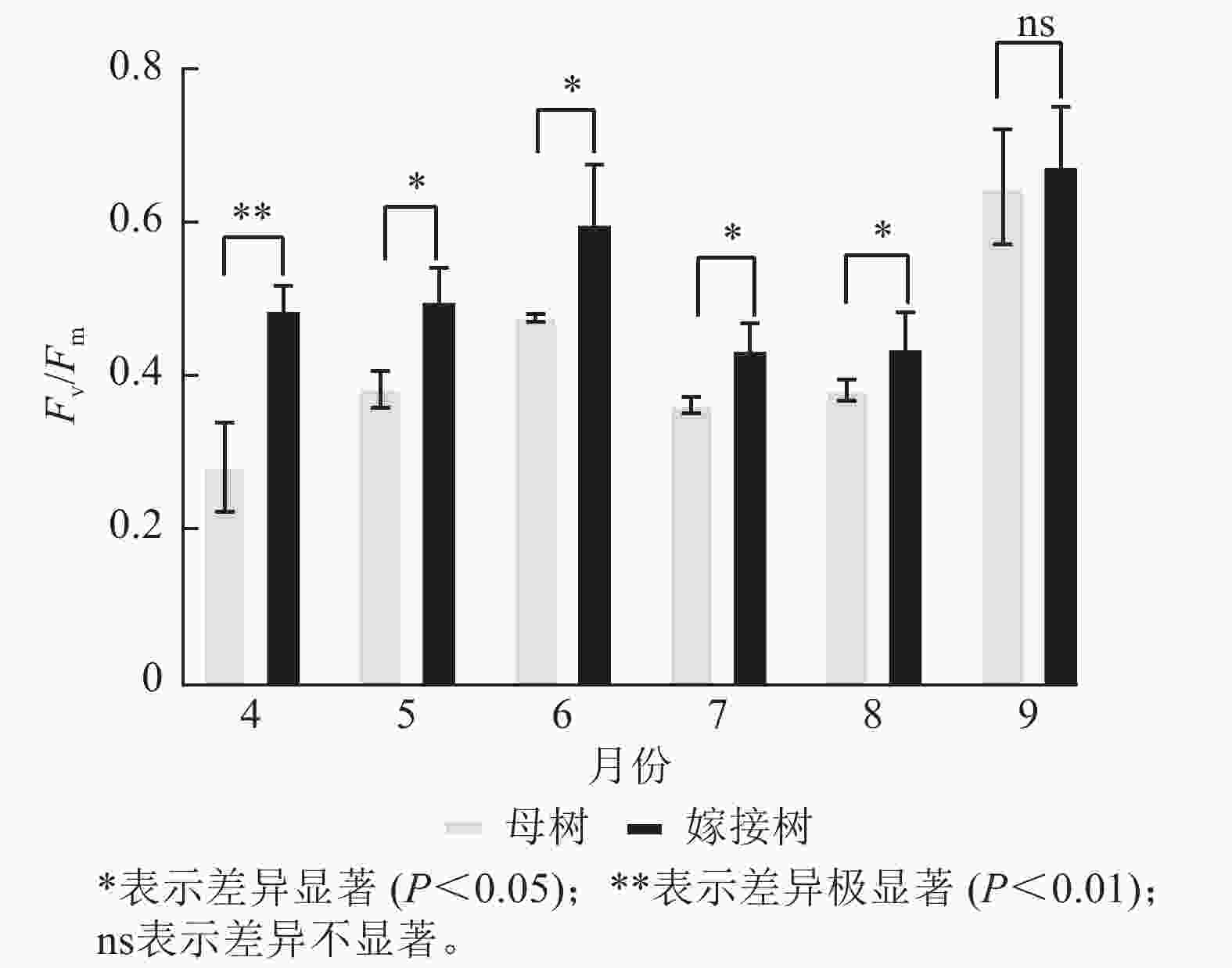
图 5 4—9月百山祖冷杉母树与嫁接树的最大光化学量子产量(Fv/Fm)比较
Figure 5. Comparison of the maximum photochemical quantum yield (Fv/Fm) between the original mother and grafted trees of A. beshanzuensis from April to September
-
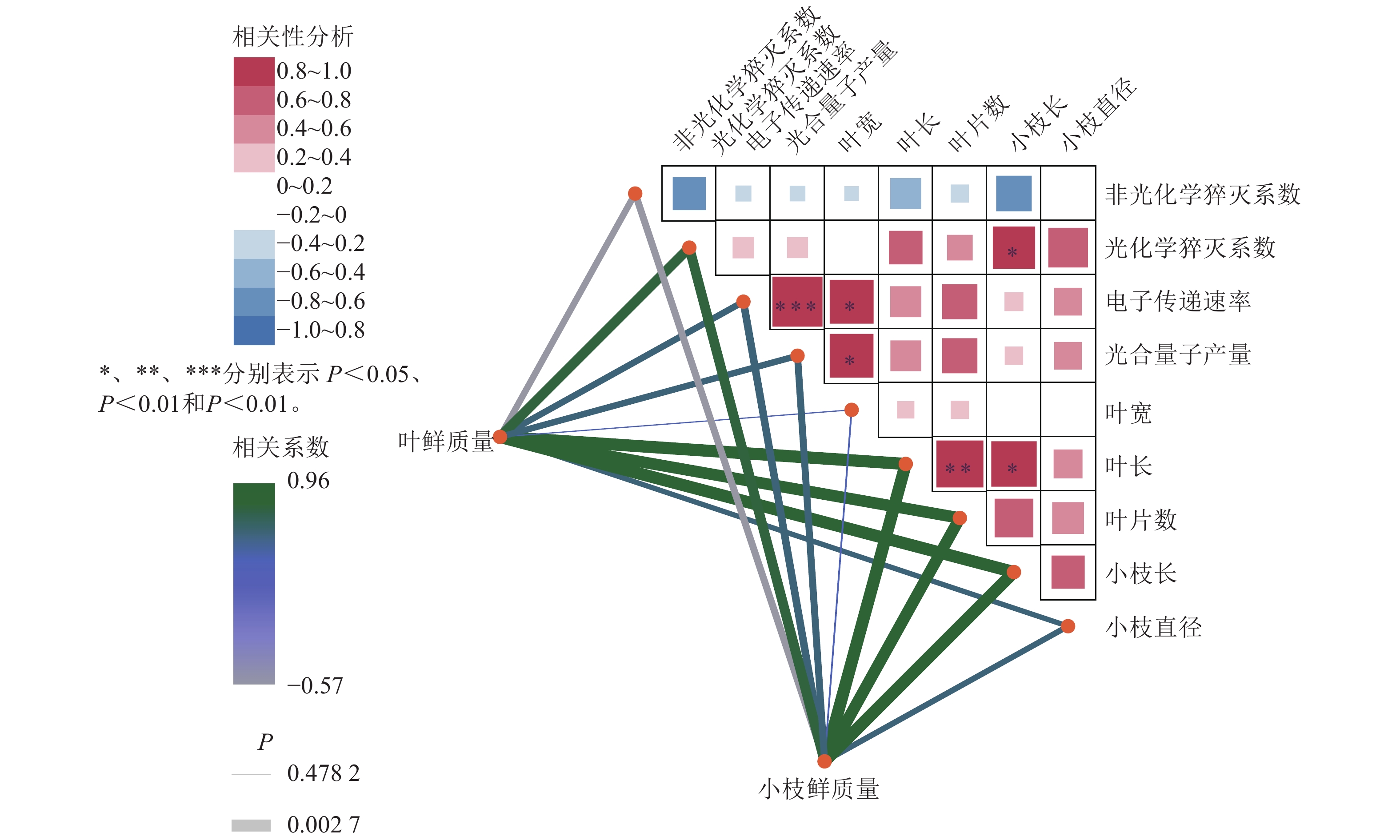
如图6所示:在光合参数之间,电子传递速率与光合量子产量呈极显著正相关(R=0.96,P<0.001);非光化学猝灭系数与光化学猝灭系数、光合量子产量均呈负相关,说明热耗散增强时光化学效率下降,符合植物在逆境下的生理响应。在光合参数与形态指标的关系方面,光合量子产量与叶片宽度、叶片长度呈显著正相关,提示光合效率较高的植株叶片发育较好;非光化学猝灭系数则与多数形态指标呈负相关,可能与胁迫状态下热耗散增强、生长受限有关。在形态指标之间,叶鲜质量与叶长、叶片数、小枝鲜质量、小枝长度与小枝直径等呈显著正相关;叶长与叶片数、小枝长度等呈显著正相关;体现生物量分配的协调性。整体而言,光合效率光合量子产量、光化学猝灭系数与叶片形态正相关,而热耗散非光化学猝灭系数与生长指标负相关,反映植物在光合性能与形态建成上的协同关系,非光化学猝灭系数升高可能指示环境胁迫。
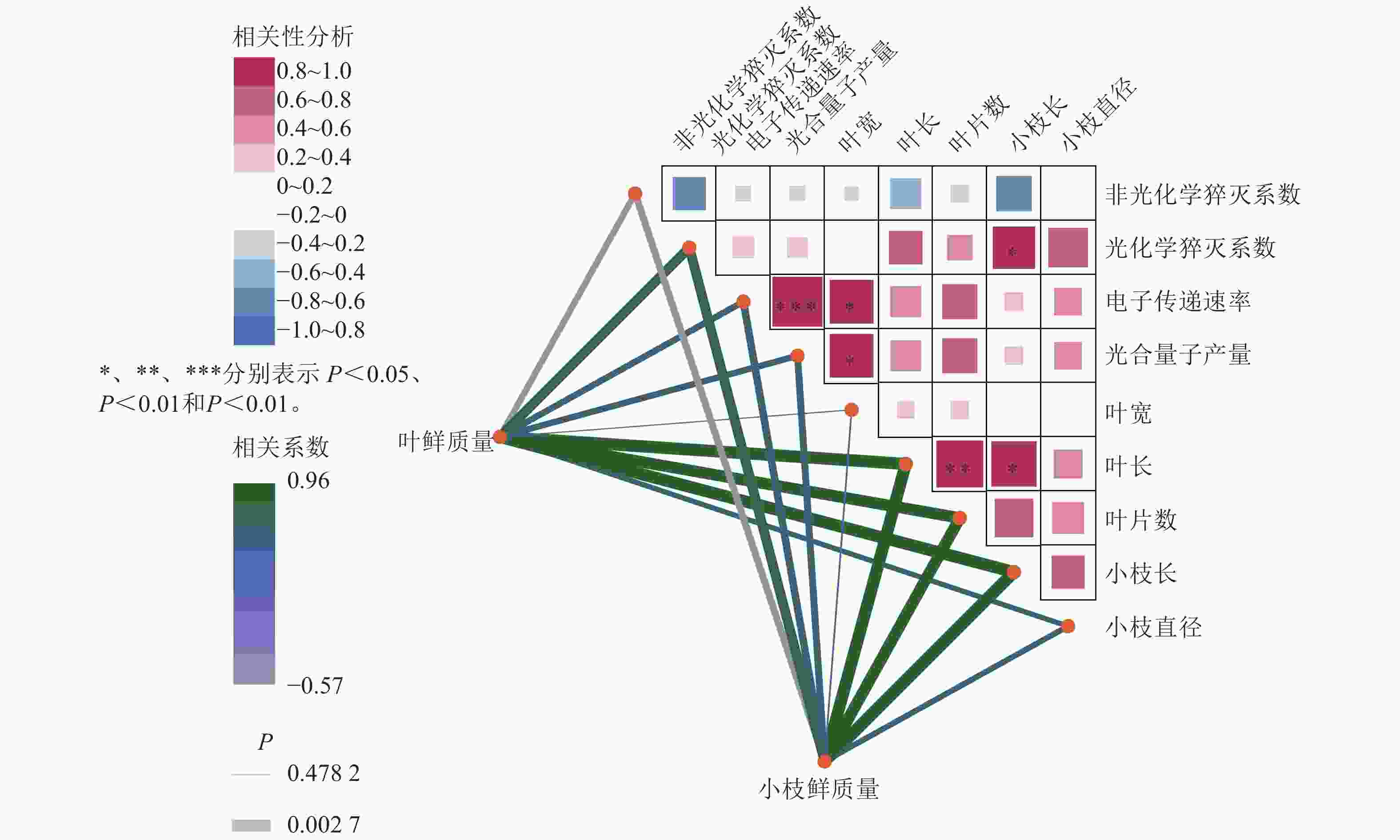
图 6 8月百山祖冷杉母树、嫁接树生长量与268 μmol·m−2·s−1光强下叶绿素荧光参数的相关性网络热图
Figure 6. heatmap of correlation networks between the growth parameters of A. beshanzuensis mother trees and grafted trees with chlorophyll fluorescence traits under 268 μmol·m−2·s−1 lightiintensity in August
-
本研究通过比较百山祖冷杉原生母树与嫁接树的生长和叶绿素荧光参数,揭示了嫁接树在光合效率和生长势上的显著优势。这一差异主要源于砧木效应、树龄及生境条件等因素的综合影响。
百山祖冷杉嫁接所用的日本冷杉砧木对其接穗生长可能起到了一定的积极作用。 砧木通常具有更强壮的根系,能更有效地吸收水分和养分[28]。在本研究中,嫁接树各项主要生长指标显著大于原生母树,很可能得益于砧木提供了更充足的资源供给。同时,优良的砧木还能通过调节激素平衡等方式改善接穗的生理状态[29],这为嫁接树拥有更高光化学猝灭系数和电子传递速率,从而将光能更高效地用于光合作用提供了生理学解释。值得注意的是,7月高温天气下,嫁接树的电子传递速率及光化学淬灭系数在>130 μmol m−2 s−1光强条件下等均显著或极显著高于原生母树,而非光化学淬灭系数在>130 μmol m−2 s−1光强条件下显著或极显著低于原生母树,表明嫁接树在高温胁迫下仍能维持较高的光能转化效率。这一差异可能与砧木效应和树龄密切相关。LI等[30]研究表明:耐热砧木可通过维持根系活力、诱导HSP70积累及增强抗氧化酶活性,有效保护PSⅡ免受高温损伤;杏树Armeniaca vulgaris嫁接研究也证实优良砧木能维持高温复合胁迫下更高的光合活性与恢复能力[31]。本研究中嫁接树所采用的日本冷杉砧木可能通过类似机制缓解了高温对光合机构的抑制。此外,树龄是影响植株长势的重要因素之一,高龄树木生理代谢活力普遍下降。王飞等[32]发现沙地云杉Picea mongolica随树龄增长光合性能发生显著下降。百山祖冷杉原生母树树龄约170 a,高龄导致的生理衰退使其对高温胁迫更为敏感。百山祖冷杉嫁接树(42 a)的1年生枝叶生长量和Fv/Fm均大于原生母树,与柑橘‘黄果柑’Citrus ‘Huangguogan’大龄树(280 a)的最大光化学效率和累计光合作用产物低于8 a和30 a植株的研究结果一致[33]。因此,嫁接树凭借砧木保护与树龄优势在夏季高温期表现出更优的光合生理性能,而原生母树受年龄与海拔双重因素制约,光合抑制更为严重。
本研究嫁接树的生长量和叶绿素荧光参数大于原生母树的研究结果,与西滨木槿Hibiscus hamabo根小枝嫁接提高盐胁迫下西洋木槿Hibiscus syriacus生长量、PSⅡ最大光化学量子产量、实际光化学量子产量以及表观电子传递速率的研究结果一致[34];与紫榆Ulmus glabra嫁接树与微繁殖树在高度、胸径和叶片形态方面等研究结果一致,而与其微繁殖榆树的PSⅡ最大光化学量子产量、潜在电子容量和可变初始荧光比较高的结果不同[35],这可能与不同植物物种和不同材料来源有关。
另外,原生母树所处高海拔生境(约1 750 m)是其生长和生理表现较弱的重要原因。高海拔生境导致树体长期面临更强的光辐射等环境胁迫,这与遮阴处理提高了百山祖冷杉光合作用能力的研究结果一致[2]。本研究中原生母树持续较低的最大光化学效率和实际光化学量子产量,以及显著升高的非光化学猝灭系数,是光抑制症状的典型表现[36]。较高的非光化学猝灭系数表明原生母树对比嫁接树启动了强烈的热耗散机制以保护光合机构免受强光破坏,但这不可避免地减少了用于光合碳同化的光能比例,导致光合效率低下[37]。因此,原生母树的表现是其在严峻生境下的一种生存适应,其代价是长势衰退。
本研究在对百山祖冷杉原生母树与嫁接树叶绿素荧光参数的连续6个月监测中发现,其最大光化学量子产量并未随着月份的增加而呈现单一的线性趋势,而是具有先上升、后下降、再上升的变化,夏季7—8月,最大光化学量子产量有明显下降,这可能是因为当气温上升到一定程度, 山顶的气温也不能满足百山祖冷杉对低温条件的需求[38],与百山祖冷杉苗木对高温胁迫的生理响应研究结果一致[10];对高温胁迫下百山祖冷杉的转录组研究中揭示了其相关分子机制[26],提示气候变化下的高温也是其面临的威胁和濒危的重要原因之一。
-
百山祖冷杉嫁接树的1年生小枝长、小枝直径和小枝鲜质量、叶长、叶片数、叶鲜质量等生长指标均显著大于原生母树,表明嫁接树生长量均优于原生母树。通过 4—9月的叶绿素荧光参数测定,嫁接树的光合量子产量、电子传递速率、光化学猝灭系数、最大光化学量子产量均大于原生母树。表明嫁接树的光能捕获效率、电子传递速率、光能转化效率均优于原生母树。嫁接树的非光化学猝灭系数均小于原生母树,表明原生母树的光能热耗散大于嫁接树。同样表明嫁接树的生长状态优于原生母树。百山祖冷杉嫁接树在光合效率和生长势上有显著优势,不仅体现了异砧嫁接在改善接穗生理状态方面的积极作用,也可能得益于其相对优越的较低海拔生境条件。 因此,在制定百山祖冷杉原生母树的精准保护策略时,除考虑嫁接繁殖作为有效的迁地保护手段外,同时应重视通过改善其原生境,如通过适度遮阴和喷雾降低高光强和高温胁迫,从而提升其光合效能、促进其生长和生存能力。
Comparative of growth and chlorophyll fluorescence parameters between original mothers and grafted trees of Abies beshanzuensis
-
摘要:
目的 探明濒危植物百山祖冷杉Abies beshanzuensis原生母树与1982年嫁接的嫁接树生长状况的差异,为百山祖冷杉原生母树精准保护提供科学依据。 方法 2023年4—9月,以百山祖冷杉原生母树和嫁接树1年生枝叶为材料,测量1年生小枝上叶的生长指标(叶长、叶宽、叶鲜质量、叶片数)和1年生小枝的生长指标(小枝长度、小枝直径、小枝鲜质量)等及其叶片的叶绿素荧光参数。 结果 8月,百山祖冷杉嫁接树的叶长(45.85 mm±0.38 mm)、叶片数(79.33片±2.40片)、叶鲜质量(4.93 g±0.21 g)、小枝长(78.66 mm±5.87 mm)、小枝直径(3.71 mm±0.21 mm)、小枝鲜质量(0.69 g±0.02 g)等生长指标均显著(P<0.05)大于原生母树的叶长(33.76 mm±2.56 mm)、叶片数(67.33片±2.91片)、叶鲜质量(2.32 g±0.25 g)、小枝长(56.39 mm±2.66 mm)、小枝直径(3.20 mm±0.15 mm)、小枝鲜质量(0.36 g±0.03 g),而叶宽在原生母树(3.49 mm±0.22 mm)和嫁接树(3.93 mm±0.45 mm)之间无显著差异。4—9月,百山祖冷杉嫁接树的光合量子产量、电子传递速率、光化学猝灭系数和最大光化学量子产量等叶绿素荧光参数均大于原生母树,而嫁接树的非光化学猝灭系数均小于原生母树。 结论 原生母树的光能利用效率较低,嫁接树生长状况优于原生母树。应加强原生母树的保护措施,通过改善生境条件提高原生母树光能利用效率,促进其生长。图6表1参38 Abstract:Objective Abies beshanzuensis is a critically endangered conifer species endemic to China. This study compares the difference in the growth status between the original mother trees and the grafted trees (grafted in 1982), in order to precisely protect the original mother trees. Method The study was conducted from April to September of 2023. Current-year branches and leaves were collected from the surviving original mother trees (located at 1 750 m altitude) and grafted trees (located at 1 550 m altitude) within Qianjiangyuan-Baishanzu National Park. In August, key growth indices were measured, including branch length, branch diameter, branch fresh weight, number of leaves, leaf length, leaf width, and leaf fresh weight. From April to September, chlorophyll fluorescence parameters, including actual photochemical efficiency (Y), electron transport rate (RET), photochemical quenching (CPQ), non-photochemical quenching (CNPQ), and the maximum photochemical efficiency of PSⅡ (Fv/Fm), were monitored monthly using a portable Mini-PAM Chlorophyll fluorometer. Dark-adapted leaves were exposed to a gradient of photosynthetic active radiation (PAR) from 0 to 1190 μmol·m−2·s−1 for measurements. Data was statistically analyzed using independent sample t-tests.Result The results showed that in August, the growth indicators of the grafted trees of A. beshanzuensis, including leaf length (45.85 mm±0.38 mm), number of leaves (79.33±2.40), leaf fresh weight (4.93 g±0.21 g), branch length (78.66 mm±5.87 mm), branch diameter (3.71 mm±0.21 mm), and branch fresh weight (0.69 g±0.02 g), were all significantly greater than those of the original mother trees, which had a leaf length of (33.76 mm±2.56 mm), number of leaves of (67.33±2.91), leaf fresh weight of (2.32 g±0.25 g), branch length of (56.39 mm±2.66 mm), branch diameter of (3.20 mm±0.15 mm), and branch fresh weight of (0.36 g±0.03 g) (P<0.05), respectively. However, there was no significant difference in leaf width between the original mother trees (3.49 mm±0.22 mm) and the grafted trees (3.93 mm±0.45 mm). From April to September, the chlorophyll fluorescence parameters of the grafted trees of A. beshanzuensis, such as Y value, RET, CPQ value, and Fv/Fm value, were all higher than those of the original mother trees, respectively, while the CNPQ value of the grafted trees was lower than that of the original mother trees. Conclusion The growth condition of the grafted trees is superior to that of the original mother trees, indicating a lower light energy utilization efficiency in the original mother trees. It is necessary to strengthen the conservation measures to improve habitat conditions and promote their growth. [Ch, 6 fig. 1 tab. 38 ref.] -
图 1 4—9月百山祖冷杉原生母树与嫁接树的光合量子产量(Y)对光强(PAR)的响应比较
Figure 1 Comparison of the response curves of photosynthetic quantum yield (Y) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
图 2 4—9月百山祖冷杉原生母树与嫁接树的电子传递速率(RET)对光强(PAR)的响应比较
Figure 2 Comparison of the response curves of electron transport rate (RET) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
图 3 4—9月百山祖冷杉原生母树与嫁接树的光化学猝灭系数(CPQ)和对光强(PAR)的响应比较
Figure 3 Comparison of the response curves of photochemical quenching coefficent (CPQ) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
图 4 4—9月百山祖冷杉原生母树与嫁接树的非光化学猝灭系数(CNPQ)和对光强(PAR)的响应比较
Figure 4 Comparison of the response curves of photochemical quenching coefficent (CNPQ) to photosynthetically active radiation (PAR) between the original mother and grafted trees of A. beshanzuensis from April to September
图 5 4—9月百山祖冷杉母树与嫁接树的最大光化学量子产量(Fv/Fm)比较
Figure 5 Comparison of the maximum photochemical quantum yield (Fv/Fm) between the original mother and grafted trees of A. beshanzuensis from April to September
图 6 8月百山祖冷杉母树、嫁接树生长量与268 μmol·m−2·s−1光强下叶绿素荧光参数的相关性网络热图
Figure 6 heatmap of correlation networks between the growth parameters of A. beshanzuensis mother trees and grafted trees with chlorophyll fluorescence traits under 268 μmol·m−2·s−1 lightiintensity in August
表 1 百山祖冷杉原生母树与嫁接树1年生枝叶的生长量
Table 1. Growth of current-year branches and leaves of the original mother and grafted trees of A. beshanzuensis
样树 小枝长/mm 小枝直径/mm 叶片数/片 叶长/mm 叶宽/mm 叶鲜质量/g 小枝鲜质量/g 原生母树 56.39±2.66 b 3.20±0.15 b 67.33±2.91 b 33.76±2.56 b 3.49±0.22 a 2.32±0.25 b 0.36±0.03 b 嫁接树 78.66±5.87 a 3.71±0.21 a 79.33±2.40 a 45.85±0.38 a 3.93±0.45 a 4.93±0.21 a 0.69±0.02 a 说明:数据为平均值±标准误,同列不同小写字母表示差异显著(P<0.05,独立样本t检验)。  下载: 导出CSV
下载: 导出CSV
-
[1] 杨玲, 梁思琪, 潘佳明, 等. 濒危植物百山祖冷杉和资源冷杉的物种划分及其遗传资源的保护[J]. 植物生态学报, 2023, 47(12): 1629−1645. YANG Ling, LIANG Siqi, PAN Jiaming, et al. Species delimitation and genetic conservation of the endangered firs Abies beshanzuensis and A. ziyuanensis[J]. Chinese Journal of Plant Ecology, 2023, 47(12): 1629−1645. DOI: 10.17521/cjpe.2022.0295. [2] 陈德良, 陶月良, 吴友贵, 等. 遮荫对百山祖冷杉光合特性和叶绿素荧光参数的影响[J]. 核农学报, 2016, 30(10): 2056−2064. CHEN Deliang, TAO Yueliang, WU Yougui, et al. Effect of shade on the photosynthetic characteristics and chlorophyll fluorescence parameters of Abies beshanzuensis M. H. Wu[J]. Journal of Nuclear Agricultural Sciences, 2016, 30(10): 2056−2064. DOI: 10.11869/j.issn.100-8551.2016.10.2056. [3] CHEN Haowei, JIANG Chunxiao, LI Jiyang, et al. Beshanzoides A-D, unprecedented cycloheptanone-containing polyketides from Penicillium commune P-4-1, an endophytic fungus of the endangered conifer Abies beshanzuensis[J]. RSC Advances, 2021, 11(63): 39781−39789. DOI: 10.1039/D1RA08377E. [4] 哀建国, 邱英雄, 余久华, 等. 百山祖冷杉的ISSR分析优化和遗传多样性初步研究[J]. 浙江大学学报(农业与生命科学版), 2005, 31(3): 277−282. AI Jianguo, QIU Yingxiong, YU Jiuhua, et al. Optimization of inter simple sequence repeats (ISSR) analysis as applied to preliminary study of genetic variation in Abies beshanzuensis M. H. Wu[J]. Journal of Zhejiang University (Agriculture & Life Sciences), 2005, 31(3): 277−282. DOI: 10.3321/j.issn:1008-9209.2005.03.009. [5] 哀建国. 百山祖冷杉濒危机制与保护对策研究[D]. 杭州: 浙江大学, 2005. AI Jianguo. Studies on the Endangerment Mechanism and Conservation Strategies for Abies beshanzuensis[D]. Hangzhou: Zhejiang University, 2005. [6] GUO Yi, WANG Quanjiu, ZHAO Xue, et al. Field irrigation using magnetized brackish water affects the growth and water consumption of Haloxylon ammodendron seedlings in an arid area[J]. Frontiers in Plant Science, 2022, 13: 929021. DOI: 10.3389/fpls.2022.929021. [7] WANG Wenyue, HUA Jinguo, ZHANG Zhen, et al. Effects of phosphorus supply levels on the growth and nutrient utilization of Pinus massoniana lamb hybrid saplings[J]. Frontiers in Plant Science, 2025, 16: 1606643. DOI: 10.3389/fpls.2025.1606643. [8] ZHENG Yueping, NITIN M, WANG Lanlan, et al. Early detection of squash silverleaf and chemical control of Bemisia tabaciusing imidacloprid[J]. International Journal of Pest Management, 2013, 59(1): 10−19. DOI: 10.1080/09670874.2012.738837. [9] QI Zixuan, YE Yuchen, SUN Lian, et al. Development of an indicator system for solar-induced chlorophyll fluorescence monitoring to enhance early warning of flash drought[J]. Agricultural Water Management, 2025, 312: 109397. DOI: 10.1016/j.agwat.2025.109397. [10] 庞振. 百山祖冷杉苗木对不同海拔高度和高温胁迫的生理响应及转录组分析[D]. 杭州: 浙江理工大学, 2023. PANG Zhen. Physiological Response and Transcriptome Analysis of Abies beshanzunesis M. H. Wu Seedlings to Different Altitudes and High Temperature Stress[D]. Hangzhou: Zhejiang Sci-Tech University, 2023. DOI: 10.27786/d.cnki.gzjlg.2023.000742. [11] YANG Haolin, WANG Lin, ZHANG Xiaolei, et al. Exploring optimal soil moisture for seedling tomatoes using thermal infrared imaging and chlorophyll fluorescence techniques[J]. Scientia Horticulturae, 2025, 339: 113846. DOI: 10.1016/j.scienta.2024.113846. [12] GONG Jiru, ZHANG Zihe, ZHANG Chunlai, et al. Ecophysiological responses of three tree species to a high-altitude environment in the Southeastern Tibetan Plateau[J]. Forests, 2018, 9(2): 48. DOI: 10.3390/f9020048. [13] 靳百慧, 孙婷, 潘磊, 等. 海拔变化对元阳梯田水稻叶片结构及叶绿素荧光特征的影响[J]. 分子植物育种, 2019, 17(22): 7467−7475. JIN Baihui, SUN Ting, PAN Lei, et al. Effects of elevation changes on chlorophyll fluorescence characteristics of different rice varieties in Yuanyang Terraces[J]. Molecular Plant Breeding, 2019, 17(22): 7467−7475. DOI: 10.13271/j.mpb.017.007467. [14] LIU Yuanhuan, LIU Fangli, LONG Bo, et al. Chlorophyll fluorescence characteristics and rapid light response curves of Alpine Rhododendron species across elevation gradients[J]. Horticultural Science and Technology, 2019, 37(4): 463−472. DOI: 10.7235/hort.20190047. [15] 郭源上, 何明珠, 韩国君, 等. 宁夏干旱区石灰岩矿山废弃地土壤改良及修复植物优选[J]. 植物资源与环境学报, 2024, 33(6): 44−55. GUO Yuanshang, HE Mingzhu, HAN Guojun, et al. Soil amelioration and restorative plant optimization for wasteland of limestone mine sites in arid region of Ningxia[J]. Journal of Plant Resources and Environment, 2024, 33(6): 44−55. DOI: 10.3969/j.issn.1674-7895.2024.06.05. [16] 张牡丹, 冯媛, 石珍珍, 等. 干旱和低温胁迫下细胞外ATP对当归幼苗叶绿素含量及其荧光特性的调节[J]. 中国中药杂志, 2019, 44(7): 1305−1313. ZHANG Mudan, FENG Yuan, SHI Zhengzheng, et al. Regulation of extracellular ATP on chlorophyll content and fluorescence characteristics of Angelica sinensis seedlings under drought and low temperature stress[J]. China Journal of Chinese Materia Medica, 2019, 44(7): 1305−1313. DOI: 10.19540/j.cnki.cjcmm.20190319.101. [17] MONTANARO G, CARLOMAGNO A, GIORIO P, et al. Are leaf chlorophyll fluorescence and Dark Green stressor-specific fingerprints in grapevine under drought or salt stress? A reanalysis study[J]. Plant Stress, 2025, 17: 100948. DOI: 10.1016/j.stress.2025.100948. [18] 邓施琴, 美朵卓嘎, 张欢, 等. 遮光对两种兰科植物叶绿素荧光参数的影响[J]. 北方园艺, 2024(22): 38−47. DENG Shiqin, Meiduozhuoga, ZHANG Huan, et al. Effects of shading on chlorophyll fluorescence parameters of two Orchidaceae species[J]. Northern Horticulture, 2024(22): 38−47. DOI: 10.11937/bfyy.20242317. [19] 姜霞, 周炳煌, 袁丛军, 等. 马尾松林下3种药用植物光合和叶绿素荧光特征[J]. 贵州林业科技, 2024, 52(4): 38−44. JIANG Xia, ZHOU Binghuang, YUAN Congjun, et al. Photosynthesis and chlorophyll fluorescence of three medicinal plants under Pinus massoniana Lamb[J]. Guizhou Forestry Science and Technology, 2024, 52(4): 38−44. DOI: 10.16709/j.cnki.gzlykj.2024.04.001. [20] LI Jiaqi, QIAO Zhensheng, ZONG Dan, et al. Effects of different light qualities on the growth characteristics of Populus trinervis[J]. Phyton, 2024, 93(5): 1043−1056. DOI: 10.32604/phyton.2024.050637. [21] HUANG D, WU L, CHEN J R, et al. Morphological plasticity, photosynthesis and chlorophyll fluorescence of Athyrium pachyphlebium at different shade levels[J]. Photosynthetica, 2011, 49(4): 611−618. DOI: 10.1007/s11099-011-0076-1. [22] 刘建锋, 杨文娟, 江泽平, 等. 遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响[J]. 生态学报, 2011, 31(20): 5999−6004. LIU Jianfeng, YANG Wenjuan, JIANG Zeping, et al. Effects of shading on photosynthetic characteristics and chlorophyll fluorescence parameters in leaves of the endangered plant Thuja sutchuenensis[J]. Acta Ecologica Sinica, 2011, 31(20): 5999−6004. DOI: 10.20103/j.stxb.2011.20.016. [23] CHAI Shengfeng, TANG Jianmin, MALLIK A, et al. Eco-physiological basis of shade adaptation of Camellia nitidissima, a rare and endangered forest understory plant of Southeast Asia[J]. BMC Ecology, 2018, 18: 5. DOI: 10.1186/s12898-018-0159-y. [24] 陈超, 金则新, 袁梦, 等. 不同光照强度下濒危植物景宁木兰幼苗光合特性的季节变化[J]. 浙江农林大学学报, 2022, 39(5): 950−959. CHEN Chao, JIN Zexin, YUAN Meng, et al. Seasonal changes of photosynthetic characteristics of seedlings of Magnolia sinostellata under different light intensities[J]. Journal of Zhejiang A&F University, 2022, 39(5): 950−959. DOI: 10.11833/j.issn.2095-0756.20210814. [25] LIU Bin, WANG Tingjin, LIU Lingjuan, et al. MYB6/bHLH13-AbSUS2 involved in sugar metabolism regulates root hair initiation of Abies beshanzuensis[J]. New Phytologist, 2023, 240(6): 2386−2403. DOI: 10.1111/nph.19301. [26] ZHAO Likang, LI Tao, CHEN Xiaorong, et al. Resistance mechanism of Abies beshanzuensis under heat stress was elucidated through the integration of physiological and transcriptomic analyses[J]. BMC Plant Biololy, 2025, 25(1): 621−635. DOI: 10.1186/s12870-025-06641-4. [27] 沈宗根, 陈翠琴, 王岚岚, 等. 3种石斛光合作用和叶绿素荧光特性的比较研究[J]. 西北植物学报, 2010, 30(10): 2067−2073. SHEN Zonggen, CHEN Cuiqin, WANG Lanlan, et al. Photosynthesis and chlorophyll fluorescence characteristics of three Dendrobium species[J]. Acta Botanica Boreali-Occidentalia Sinica, 2010, 30(10): 2067−2073. [28] HE Zhilong, ZHANG Ying, XUN Chengfeng, et al. Drought resistance evaluation of Camellia oleifera var. ‘Xianglin 210’ grafted onto different rootstocks[J]. Plants, 2025, 14(16): 2568. DOI: 10.3390/plants14162568. [29] RASOOL A, MANSOOR S, BHAT K M, et al. Mechanisms underlying graft union formation and rootstock scion interaction in horticultural plants[J]. Frontiers in Plant Science, 2020, 11: 590847. DOI: 10.3389/fpls.2020.590847. [30] LI Hao, AHAMMED G J, ZHOU Guona, et al. Unraveling main limiting sites of photosynthesis under below- and above-ground heat stress in cucumber and the alleviatory role of Luffa rootstock[J]. Frontiers in Plant Science, 2016, 7: 746. DOI: 10.3389/fpls.2016.00746. [31] DOGAN M, BOLAT I, TURAN M, et al. Differential rootstock-mediated regulation of physiological and hormonal responses enhances apricot resilience to combined drought and heat stress[J]. Physiologia Plantarum, 2025, 177(4): e70423. DOI: 10.1111/ppl.70423. [32] 王飞, 刘世增, 李得禄. 不同龄沙地云杉光合荧光生理特征的比较[J]. 中国农学通报, 2016, 32(28): 7−10. WANG Fei, LIU Shizeng, LI Delu. Photosynthetic and fluorescence physiological characteristics of different aged Picea mongolica[J]. Chinese Agricultural Science Bulletin, 2016, 32(28): 7−10. [33] XIONG B, QIU X, HUANG S J, et al. Comparative analysis of leaf photosynthetic characteristics and fruit sugar content in trees of Citrus cultivar ‘Huangguogan’ of different ages[J]. Photosynthetica, 2020, 58(4): 902−910. DOI: 10.32615/ps.2020.042. [34] ZHANG Shuqing, YU Wanwen, LU Zhiguo, et al. Hibiscus hamabo rootstock-grafting improves photosynthetic capacity of Hibiscus syriacus under salt stress[J]. Forests, 2023, 14(6): 1226. DOI: 10.3390/f14061226. [35] ĎURKOVIČ J, ČAŇOVÁ I, PRIWITZER T, et al. Field assessment of photosynthetic characteristics in micropropagated and grafted wych elm (Ulmus glabra Huds.) trees[J]. Plant Cell, Tissue and Organ Culture (PCTOC), 2010, 101(2): 221−228. DOI: 10.1007/s11240-010-9680-1. [36] BROETTO F, DUARTE HM, LÜTTGE U. Responses of chlorophyll fluorescence parameters of the facultative halophyte and C3-CAM intermediate species Mesembryanthemum crystallinum to salinity and high irradiance stress[J]. Journal of Plant Physiology, 2007, 164(7): 904−912. DOI: 10.1016/j.jplph.2006.04.010. [37] MAO Haotian, PANG Xuan, LI Teng, et al. Chlorophyllbis essential for the growth, photoprotection, and photosystem I assembly in wheat[J]. The Plant Journal, 2025, 123(4): e70442. DOI: 10.1111/tpj.70442. [38] 李晓笑, 陶翠, 王清春, 等. 中国亚热带地区4种极危冷杉属植物的地理分布特征及其与气候的关系[J]. 植物生态学报, 2012, 36(11): 1154−1164. LI Xiaoxiao, TAO Cui, WANG Qingchun, et al. Characteristics of geographic distribution of four critically endangered species of Abies in subtropical China and its relationship with climate[J]. Chinese Journal of Plant Ecology, 2012, 36(11): 1154−1164. DOI: 10.3724/sp.j.1258.2012.01154. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250414
 点击查看大图
点击查看大图
计量
- 文章访问数: 163
- HTML全文浏览量: 38
- PDF下载量: 47
- 被引次数: 0
