





-
水分和养分是影响植物生长发育的两大重要因素。水分是植株输送营养的必要条件,养分只有溶解在水中才能被植物吸收,水分状况在很大程度上决定着肥料的有效性、吸收量和利用率[1-3]。在农林生产中,经常通过灌溉或施肥来改善植物的生长状况,但施用过多会造成水资源和肥料浪费,严重时还会导致水体富营养化、土壤盐渍化等环境问题[4]。水肥耦合可以提高水分和肥料的利用效率,还能调节苗木的生理生长过程,达到节水节肥、促进苗木生长的目的[5]。对花椒Zanthoxylum bungeanum[6]和楸树Catalpa bungei[7]进行水肥研究发现:适宜的水肥耦合条件能够促进幼苗生长和养分吸收利用,能显著提高苗木生物量。栓皮栎Quercus variabilis是壳斗科Fagaceae栎属Quercus落叶乔木,是中国温带、暖温带和亚热带地区森林植被的优势树种,是生产木材、软木、栲胶、薪炭、食用菌等的主要原料,也是发展燃料乙醇重要的“非粮”原料[8],具有很高的经济价值。作为提供大径级无节良材的珍贵乡土树种,栓皮栎在中国水源涵养、水土保持、改善生态环境方面发挥重要的作用[9-10]。栓皮栎在天然林中种群更新困难,育苗成活率低,苗木质量不高,因此深入开展栓皮栎水肥需求规律的研究,对于提高出圃栓皮栎苗木质量具有重要意义。目前,关于栓皮栎苗期水肥需求的研究,主要涉及水分和氮肥单因素及交互效应对苗木生长和养分状况的影响[11-13]。磷、钾肥对植物的生长同样具有重要作用,是植物必需的营养元素之一[7],但磷、钾元素如何影响栓皮栎苗木生长的研究尚未有相关报道,且有关水肥交互效应如何影响苗高、地径、根系生物量等重要苗木质量指标的研究较少,无法制定精准的灌溉施肥制度。本研究以栓皮栎当年生幼苗为研究对象,采用4因素5水平二次回归通用旋转组合设计进行盆栽试验,通过水肥耦合处理对苗木生长指标的测定,建立其与土壤水分、施氮量、施磷量、施钾量的回归模型,探讨栓皮栎生长对水肥耦合的响应规律,重点探究各生长指标的水肥耦合交互效应,确立最佳水肥组合,以期为栓皮栎苗期的水分管理和合理施肥提供理论依据。
-
于2018年4−11月在中国林业科学研究院科研温室(40°00′10″N,116°14′38″E)中进行试验。海拔61 m,研究期间平均气温为24.0 ℃,夜间气温不低于10.0 ℃,白天气温不高于30.0 ℃,光合光子照度为200~1 000 μmol·m−2·s−1,湿度为60%~70%。
-
2017年9−10月,于河南林州随机选取50株健康栓皮栎母树,母树间距不小于20 m,采集到的种子用60 ℃温水持续浸泡2 h杀虫处理后置于阴凉处阴干。对种子进行筛选、消毒,将成熟种子进行沙藏储存。2018年4月挑选已发芽的栓皮栎种子断根并播种,种子播在花盆(内径10 cm,高20 cm)内,培养基质为V(泥炭土)∶V(珍珠岩)=5∶3,基质容重为0.28 g·cm−3,田间持水量18.5%,pH 6.05,速效氮、速效磷、速效钾质量分数分别为74.67、27.17、11.63 mg·kg−1。
种子播种后到幼苗出苗整齐,统一采用上方喷灌,每天喷水1次,使基质保持湿润。出苗后每隔1周喷洒质量浓度为0.1%的50%多菌灵可湿性粉剂。在培育过程中,采用自然光,所有苗木每周进行1次位置轮换,以减少边缘效应。待5月中旬出苗整齐后开始进行试验,试验处理前测定苗木平均高度和地径分别为16.8 cm和2.09 mm。肥料选用氮肥(尿素,含质量分数为46%的氮)、磷肥(磷酸二氢钠,含质量分数为26%磷)、钾肥(硫酸钾,含质量分数为50%钾)。
-
采用4因素5水平二次回归通用旋转设计(RCCD),共30个处理。各试验因素和水平编码值见表1,试验结构矩阵见表2。土壤含水率上限设为田间持水量。采用FOM/mts便携式土壤湿度、温度和盐度计测量土壤水分,同时结合称量法进行校正。3 d测定1次土壤含水率,测定时将传感器的探针插入基质中进行测定,得到体积含水量,根据土壤容重,经换算得到质量含水率,测得土壤含水率(占田间持水量的比例)低于下限则灌水。灌水量(mL)=[(田间持水量−土壤含水率)/容重]×盆土质量/水的密度。于5月底到7月底,7 d施氮肥1次,共10次,15 d施1次磷肥和钾肥,共5次[13]。每次施肥量固定,对苗木进行水分补充时将肥料溶于水中施入。此期间为栓皮栎苗木的速生期,水溶肥少量多施,符合植物根系不间断吸收养分的特点,以减少一次性大量施肥造成肥料损失。
表 1 试验因素和水平编码值
Table 1. Coded and physical values of experimental factors
编码水平 土壤含水
率/%施氮量/
(mg·株−1)施磷量/
(mg·株−1)施钾量/
(mg·株−1)−2.0 40 0 0 0 −1.0 50 75.0 15.0 15.0 0 60 150.0 30.0 30.0 1.0 70 225.0 45.0 45.0 2.0 80 300.0 60.0 60.0 表 2 结构矩阵和各处理栓皮栎苗木生长情况
Table 2. Experimental design matrix and growth of Q. variabilis seedlings under different treatments
处理 x1 x2 x3 x4 苗高/cm 地径/mm 根干质量/g 生物量/g 叶面积/cm2 1 −1.0 −1.0 −1.0 −1.0 31.97±0.56 h 3.46±0.11 cd 3.45±0.03 k 6.69±0.18 de 291.93±3.41 g 2 1.0 −1.0 −1.0 −1.0 38.80±0.85 cd 3.82±0.20 bc 4.57±0.02 e 8.53±0.25 bc 364.24±7.36 de 3 −1.0 1.0 −1.0 −1.0 30.07±0.75 i 3.92±0.10 bc 3.84±0.03 i 6.90±0.23 de 274.35±4.79 h 4 1.0 1.0 −1.0 −1.0 40.20±0.77 c 4.15±0.17 ab 5.21±0.04 b 9.71±0.12 b 397.33±10.49 c 5 −1.0 −1.0 1.0 −1.0 29.47±0.73 i 3.42±0.13 d 3.66±0.02 j 6.47±0.41 de 275.37±5.81 h 6 1.0 −1.0 1.0 −1.0 39.03±0.92 cd 3.93±0.19 bc 4.74±0.05 d 9.18±0.10 bc 370.65±7.67 de 7 −1.0 1.0 1.0 −1.0 30.92±0.64 hi 3.83±0.16 bc 4.10±0.03 h 7.33±0.18 cd 298.80±10.18 g 8 1.0 1.0 1.0 −1.0 42.27±0.98 b 4.24±0.20 ab 5.81±0.04 a 10.79±0.11a 426.75±13.37 b 9 −1.0 −1.0 −1.0 1.0 28.90±0.76 i 3.72±0.10 c 3.48±0.03 k 6.18±0.17 de 259.70±11.61 i 10 1.0 −1.0 −1.0 1.0 37.00±0.91 e 4.10±0.09 ab 4.56±0.05 e 8.65±0.11 bc 360.34±5.49 e 11 −1.0 1.0 −1.0 1.0 31.30±0.54 hi 3.66±0.13 cd 3.80±0.03 i 6.82±0.16 de 276.32±9.52 h 12 1.0 1.0 −1.0 1.0 39.33±0.85 cd 4.03±0.14 b 5.29±0.02 b 9.95±0.18 ab 390.31±6.27 cd 13 −1.0 −1.0 1.0 1.0 30.77±0.66 hi 3.61±0.11 cd 3.20±0.02 l 5.88±0.39 e 249.13±6.11 i 14 1.0 −1.0 1.0 1.0 38.97±0.82 cd 4.16±0.17 ab 4.67±0.04 d 8.94±0.11 bc 367.71±11.04 de 15 −1.0 1.0 1.0 1.0 30.77±0.60 hi 3.45±0.12 cd 3.78±0.01 i 6.96±0.45 de 294.75±8.29 g 16 1.0 1.0 1.0 1.0 44.57±0.95 a 4.04±0.20 b 5.75±0.03 a 10.99±0.12 a 445.03±13.33 a 17 −2.0 0 0 0 26.03±0.67 j 3.44±0.14 cd 2.60±0.06 m 4.85±0.35 e 245.46±5.69 i 18 2.0 0 0 0 43.13±0.99 b 4.34±0.26 a 5.72±0.05 a 10.69±0.35 ab 439.87±11.94 ab 19 0 −2.0 0 0 33.27±0.74 g 3.61±0.17 cd 3.77±0.04 i 7.10±0.19 c 272.22±7.48 hi 20 0 2.0 0 0 36.37±0.70 ef 3.82±0.15 bc 4.87±0.04 c 8.85±0.18 bc 339.81±8.12 f 21 0 0 −2.0 0 31.53±0.64 h 3.88±0.16 bc 4.13±0.04 h 7.48±0.18 c 327.63±4.80 f 22 0 0 2.0 0 33.97±0.77 g 3.91±0.19 bc 4.50±0.03 ef 8.30±0.20 bc 334.72±10.71 f 23 0 0 0 −2.0 35.50±0.81 f 3.92±0.17 bc 4.42±0.04 f 8.07±0.19 bc 335.99±10.88 f 24 0 0 0 2.0 36.33±0.58 ef 3.88±0.17 bc 4.30±0.03 g 7.95±0.31 c 330.40±12.97 f 25 0 0 0 0 37.87±0.89 de 4.00±0.22 bc 4.67±0.03 de 8.84±0.18 bc 376.50±9.18 d 26 0 0 0 0 36.77±0.69 ef 4.10±0.15 ab 4.45±0.04 f 8.44±0.20 c 356.43±4.59 e 27 0 0 0 0 38.53±0.78 d 4.05±0.12 b 4.80±0.03 cd 9.05±0.19 bc 397.25±4.30 c 28 0 0 0 0 39.33±0.82 cd 4.00±0.16 bc 4.70±0.03 d 8.82±0.19 bc 394.06±12.09 c 29 0 0 0 0 37.80±0.70 de 3.90±0.13 bc 4.87±0.04 c 9.09±0.21 bc 386.09±9.43 cd 30 0 0 0 0 39.77±0.81 cd 3.91±0.09 bc 4.70±0.04 d 8.85±0.17 bc 394.45±6.87 c 说明:x1、x2、x3、x4分别表示土壤含水率、施氮量、施磷量和施钾量的水平编码值;表中数值为平均值±标准误;不同小写字母代 表各处理间苗木苗高、地径和生物量在0.05水平上差异显著 -
2018年10月测量每处理苗木的苗高、地径,并对苗木进行破坏性取样。使用LI-3000(LI-COR Inc.,美国)测定叶面积,每处理测15株。随后将待测植株分成根、茎、叶,置于烘箱105 ℃杀青30 min,80 ℃烘干至恒量,称量。用Excel 365和SPSS 23.0进行数据整理,采用Design Expert 8.0.6与Matlab 2018建立模型并进行回归分析和回归系数显著性检验。各指标与土壤含水率和施肥量的回归分析采用如下模型:
$$\begin{split} y{\rm{ = }}&{b_0}{\rm{ + }}{b_1}{x_{\rm{1}}} + {b_2}{x_2} + {b_3}{x_3}{\rm{ + }}{b_4}{x_4} + {b_1}{b_2}{x_1}{x_2} + {b_1}{b_3}{x_1}{x_3}{\rm{ + }}{b_1}{b_4}{x_1}{x_4} + {b_2}{b_3}{x_2}{x_3} + \\ & {b_2}{b_4}{x_2}{x_4} + {b_3}{b_4}{x_3}{x_4} + {b_{11}}{x_1}^2 + {b_{22}}{x_2}^2 + {b_{33}}{x_3}^2 + {b_{44}}{x_4}^2 \text{。} \end{split} $$ 式(1)中:y表示响应变量,x1、x2、x3、x4分别表示土壤含水率、施氮量、施磷量和施钾量的水平编码值,b表示回归系数[14]。
-
如表2所示:各处理苗木的苗高、地径的趋势总体一致,且存在显著差异(P<0.05)。低水、低肥处理生长量相对较低,其中处理17苗高、地径、单株叶面积数值最低。高水、高氮处理生长量较大,其中处理8、处理16、处理18苗高为40.20~44.57 cm,处理4、处理8、处理18地径为4.15~4.34 mm。处理8、处理16、处理18的叶面积为426.75~445.03 cm2。总的来说,不同水肥配比对各指标的影响有一定差异,适宜的水肥配比可以促进苗木生长。
-
如表2所示:各处理苗木总生物量间存在显著差异,低水、低氮肥的处理生物量较低,其中处理17生物量最低。高水、高氮肥处理生物量较大,其中处理12、处理8、处理16、处理18生物量为9.95~10.99 g,处理8、处理16、处理18的根系生物量为5.72~5.81 g。适宜的灌溉和施肥量对苗木生物量有促进作用,土壤水分含量过低,施肥水平过高或过低都可能抑制苗木生长。
-
以栓皮栎苗木苗高、地径、叶面积、根干质量、总生物量为目标函数,土壤含水率、施氮量、施磷量和施钾量为自变量,运用方差分析和F-test对方程的可靠性和匹配程度进行检验,回归方程均为显著(P<0.05),R2为0.950 1~0.981 7,因此,模拟结果与实际情况拟合较好。剔除不显著的回归系数后,模型表达如下:
$${y_{\text{苗高}}} {\text{=}} 38{\rm{.34 \,+\, 4}}{\rm{.59}}{x_1} \,+\, 0.82{x_2} \,+\, 0.59{x_3} \,+\, 0.66{x_1}{x_2} + 0.61{x_1}{x_3} \,-\, 0.83{x_1}^2 \,-\, 0.70{x_2}^2 \,-\, 1.28{x_3}^2 - 0.49{x_4}^2\text{;}$$ (2) $${y_{\text{地径}}} {\text{=}} 3{\rm{.990 \,+\, 0}}{\rm{.220}}{x_{\rm{1}}} \,+\, 0.063{x_2}{\rm{ \,+\, }}0.046{x_1}{x_3} - 0.120{x_2}{x_4} \,-\, 0.026{x_1}^2 \,-\, 0.070{x_2}^2 \,-\, 0.026{x_3}^2 - 0.024{x_4}^2\text{;}$$ (3) $$ \begin{split} \!\!\!\!\!\!{y_{\text{叶面积}}}\,{\text{=}}\, &{\rm{ 384}}{\rm{.13 \,+\, 53}}{\rm{.78}}{x_1} \,+\, 16.66{x_2} \,+\, 5.33{x_3} \,+\, 8.03{x_1}{x_2} \,+\, 8.77{x_2}{x_3}\, -\, 9.42{x_1}^2 \,-\, 18.59{x_2}^2 \,-\, 12.30{x_3}^2 - \\ & 11.79{x_4}^2\text{;} \end{split} $$ (4) $$ \begin{split} {y_{\text{根干质量}}} {\text{=}} & 4{\rm{.700 + 0}}{\rm{.730}}{x_1} + 0.310{x_2} + 0.094{x_3} + 0.110{x_1}{x_2} + 0.074{x_1}{x_3} + 0.067{x_2}{x_3} - 0.120{x_1}^2 - 0.081{x_2}^2 - \\ & 0.083{x_3}^2 - 0.071{x_4}^2\text{;} \end{split} $$ (5) $$ \begin{split} {y_{\text{生物量}}} {\text{=}} & 8{\rm{.85 + 1}}{\rm{.47}}{x_1} \,+\, 0.52{x_2} \,+\, 0.20{x_3} \,+ \,0.21{x_1}{x_2}\, +\, 0.19{x_1}{x_3} + 0.12{x_1}{x_4} + 0.14{x_2}{x_3} - 0.23{x_1}^2 - 0.18{x_2}^2 -\\ & 0.20{x_3}^2 - 0.17{x_4}^2\text{。} \end{split} $$ (6) 式(2)~(6)中,y表示响应变量,x1、x2、x3、x4分别表示土壤含水率、施氮量、施磷量和施钾量的水平编码值。
-
土壤含水率、施氮量、施磷量对苗高、单株叶面积、根干质量、生物量有显著正效应,效应从高到低依次为土壤含水率、施氮量、施磷量;土壤含水率、施氮量对地径有显著正效应,且土壤含水率效应高于施氮量。施钾量对所有指标的效应不显著,施磷对地径无显著效应。此外,土壤含水率×施氮量、土壤含水率×施磷量对苗高有显著正交互效应,且后者大于前者;土壤含水率×施磷量对地径有显著正交互效应,施氮量×施钾量有显著负交互效应,且施氮量×施钾量的负效应大于土壤含水率×施磷量的正效应;土壤含水率×施氮量、施氮量×施磷量对单株叶面积有显著正交互效应,且后者大于前者;土壤含水率×施氮量、土壤含水率×施磷量、施氮量×施磷量、土壤含水率×施钾量对生物量有显著正交互效应,且从高到低依次为土壤含水率×施氮量、土壤含水率×施磷量、施氮量×施磷量、土壤含水率×施钾量;土壤含水率×施氮量、土壤含水率×施磷量、施氮量×施磷量根系生物量有显著正交互效应,且从高到低依次为土壤含水率×施氮量、土壤含水率×施磷量、施氮量×施磷量。土壤含水率和施氮量的交互效应对植物的生长影响更大。
-
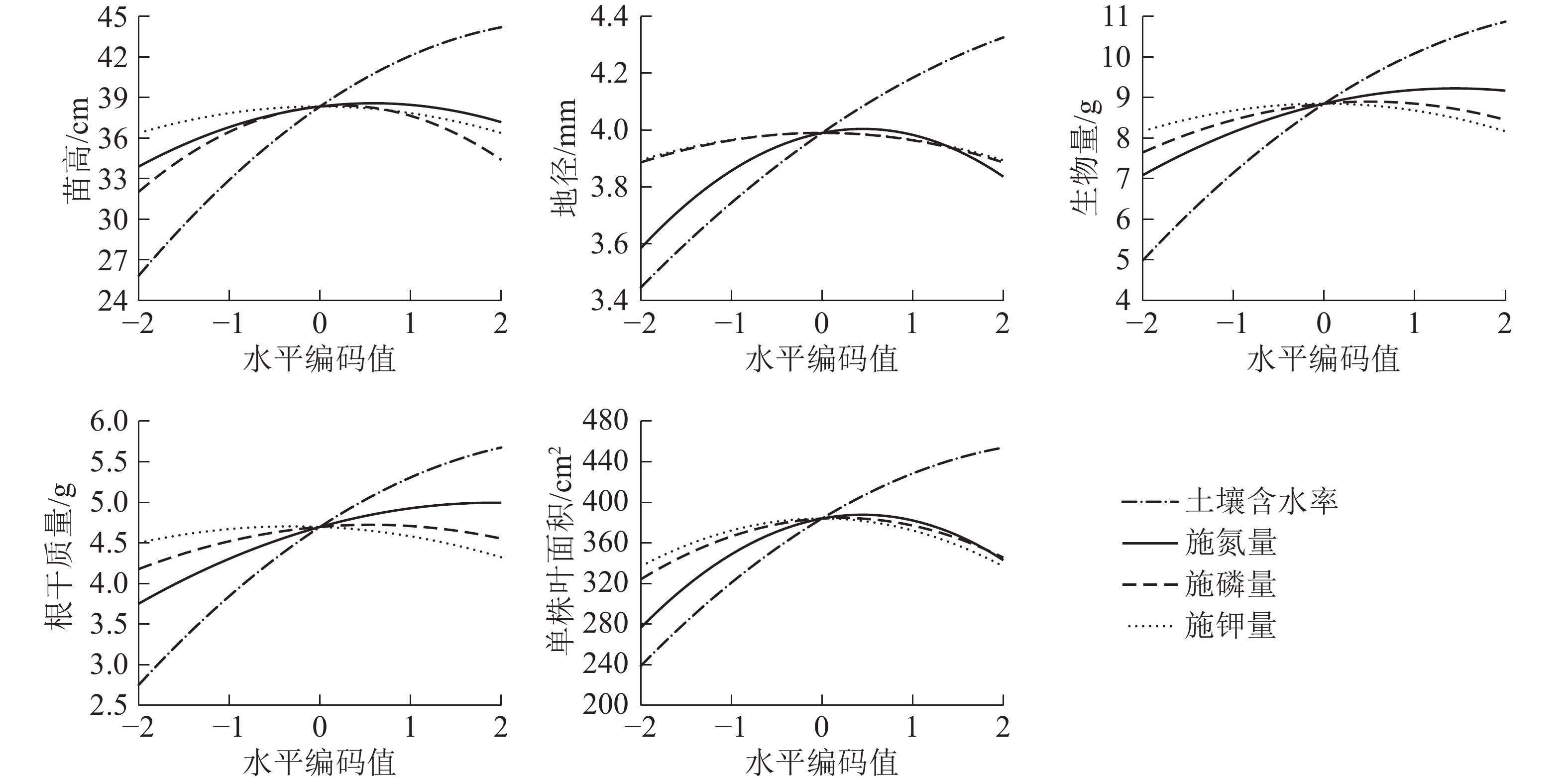
为了更直观地分析水肥因素对生长的影响效应,对回归模型进行降维处理,分析各个因素对苗高、地径、单株叶面积、根干质量、生物量的单独效应。如图1所示:氮、磷和钾效应对各指标的影响均为抛物线形状,呈现出先增加后减小的变化趋势,符合“报酬递减”的规律,但各指标达到峰值时对应的土壤水分、施肥量因子不同。当土壤含水率达到一定程度(x1=1.0)时,增长速率减缓。5个指标均随着施氮量的增加而显著增加,当施氮量达到一定水平(x2=1.0)时,苗木的生长会受到抑制,开始出现下降的趋势。磷肥对各指标的影响趋势为先促进后抑制,x3=0.5时,开始对根系生长和总生物量产生抑制作用。在较低水平(x4=−0.5)的钾肥下5个指标均可达到最高值,且x4=2.0时比不施钾肥的苗木生长量和生物量更小。土壤水分对幼苗的生长影响更大,其次为氮肥。增加土壤水分和施肥量可以显著促进幼苗的生长和生物量的积累,但过度灌溉和施肥会限制幼苗生长。
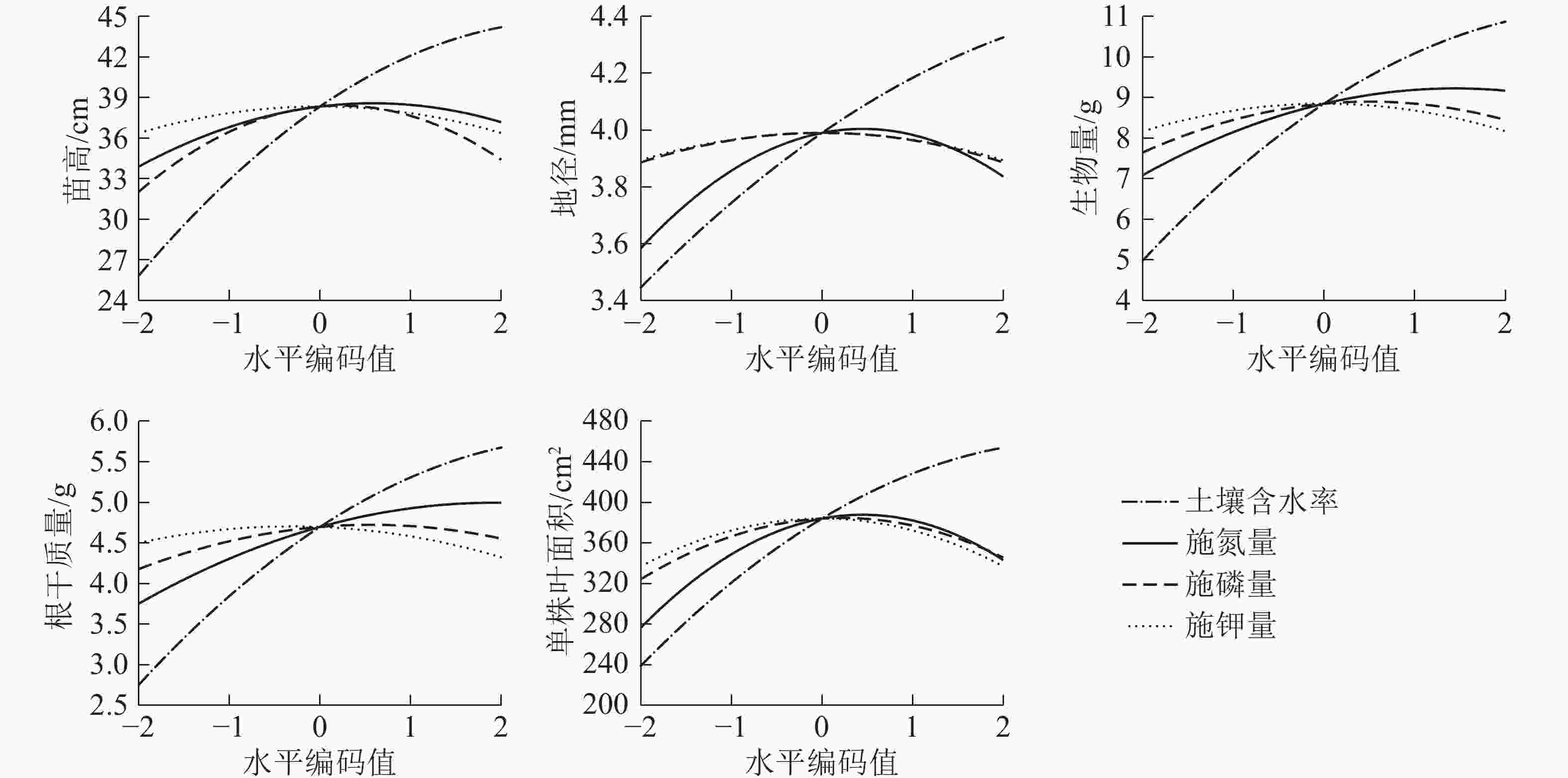
图 1 土壤含水率、施氮量、施磷量、施钾量对栓皮栎苗高、地径、单株叶面积、根干质量、生物量单因素效应
Figure 1. Monofactor effects of soil water content, N, P and K fertilizers on stem height, ground diameter, root weight, biomass, leaf area of Q. variabilis seedlings
-
根据各指标的回归方程,对其中效应显著的交互项做耦合效应分析。如图2所示:土壤含水率×施氮量、土壤含水率×施磷量对苗高有显著正效应。苗高随着土壤含水率和施氮量的增加而显著增加,当x1=−2.0、x2=1.5时,苗高达到最大,为45.79 cm。低水分条件下,低磷肥和高磷肥均不利于苗高生长,苗高随着水分和磷肥同时增加而增大,达到一定程度时增加速率减缓,当x1=2.0、x3=0.7时,苗高最大可达44.84 cm。土壤含水率×施磷量对地径有显著正效应。水分较低时,磷肥过高或者过低都会抑制地径生长。水分较高时,添加适量磷肥有利于地径生长,当x1=2.0、x3=1.6,地径可达4.40 mm。施氮量×施钾量对地径有显著负效应,氮肥和钾肥都较低时地径生长受到抑制,但随着氮肥的增加,添加少量钾肥可以促进地径的增加,随后继续添加钾肥会对地径产生显著的抑制作用。
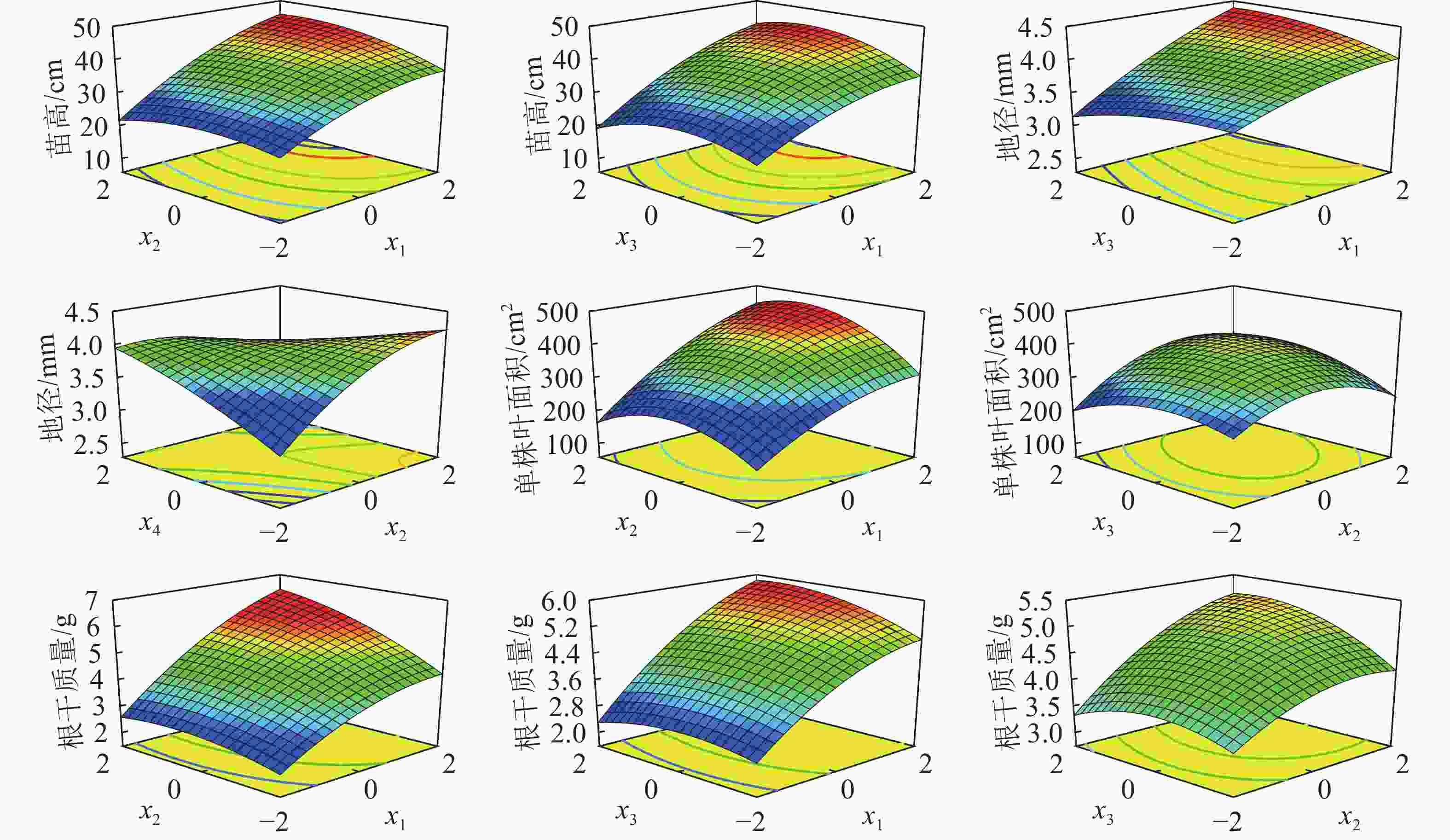
图 2 水肥耦合对栓皮栎苗木苗高、地径、叶面积、根干质量的影响
Figure 2. Coupling effects of water and fertilizeron stem height, ground diameter, leaf area, root weight of Q. variabilis seedlings
如图2所示:土壤含水率×施氮量、施氮量×施磷量对叶面积有显著正效应。单株叶面积随着水分和氮肥的增加而逐渐增大,当x1=2.0、x2=0.9时,单株叶面积达到最大为467.00 cm2。氮肥和磷肥的交互效应相比水氮交互效应小,且随着氮肥和磷肥的增加,单株叶面积先增加后降低,当x2=0.5、x3=0.4时,单株叶面积最大达到390.00 cm2。
如图2所示:土壤含水率×施氮量、土壤含水率×施磷量、施氮量×施磷量对根干质量有显著正效应。随着水分和氮肥的同时增加,根干质量逐渐增大,水分较氮肥的影响更大,当x1=2.0、x2=2.0时,根干质量达到最大为6.40 g。水磷交互作用较水氮交互作用小,随着磷肥和水分的增加,根干质量逐渐增大,当x1=2.0、x3=1.2时,根干质量可达最大,为5.80 g。肥料之间的交互作用较水分和肥料的交互作用小,氮肥和磷肥均处于较低水平时根干质量较小,随着施肥量的增加,根干质量呈现出先增大后减小的趋势,当x2=1.3、x3=1.0时,根干质量可达最大为5.10 g。
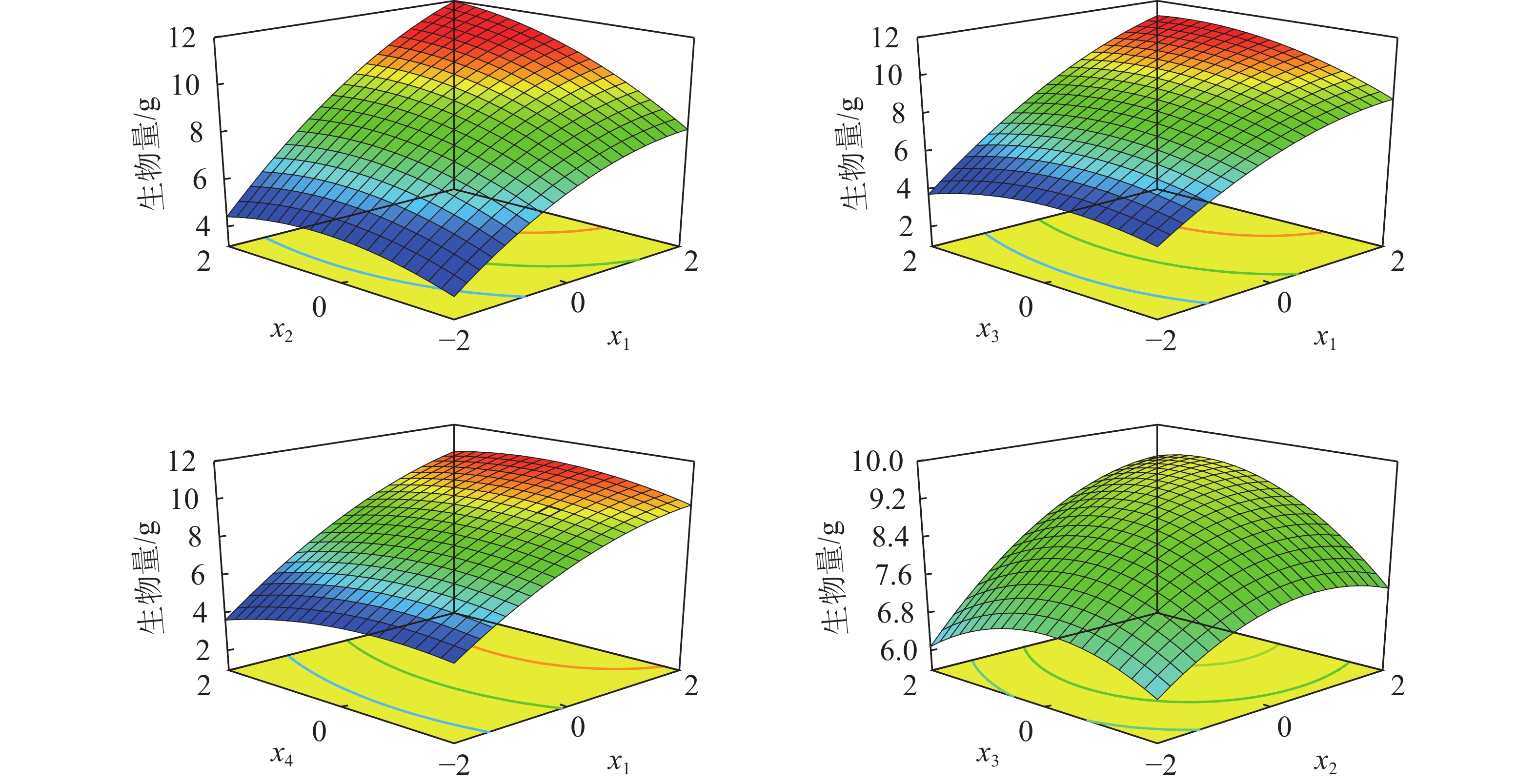
如图3所示:土壤含水率×施氮量、土壤含水率×施磷量、土壤含水率×施钾量、施氮量×施磷量对苗木生物量有显著正效应。低水分和氮肥苗木的生物量较小,随着水分和氮肥的增加,生物量逐渐增大,当x1=2.0、x2=2.0时,植株生物量可达到11.95 g。低磷肥和高磷肥均能抑制苗木生长,随着水分和磷肥的增加,生物量呈现先增加后减小的变化趋势,直到x1=1.9、x3=1.4,生物量可以达到11.20 g。低钾肥和高钾肥均会抑制苗木生长,且在低水和高钾肥的条件下生物量较小,当x1=2.0、x4=0.5时,生物量最高可达10.88 g。氮和磷肥的交互作用相对水肥交互作用的影响小,且生物量随施氮量和施磷量的增加先增大后减小;在低水分条件下,过高的磷、钾肥比低磷、钾肥更易对生物量产生抑制作用,当x2=1.7、x3=1.0时,生物量达到最大为9.43 g。适宜的水肥比有利于苗木生长和生物量的积累。
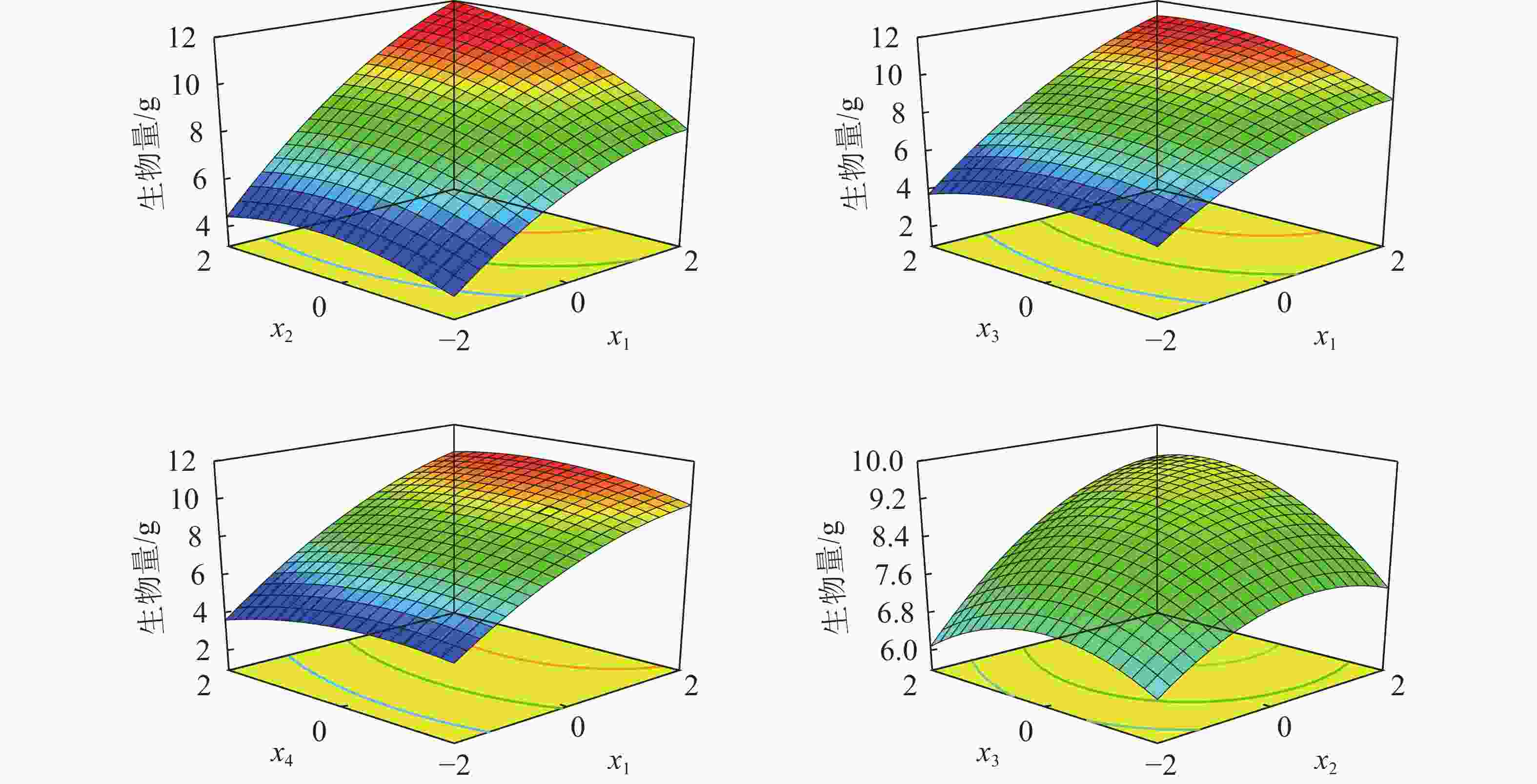
图 3 水肥耦合对栓皮栎苗木生物量的影响
Figure 3. Coupling effects of water and fertilizer on biomass of Q. variabilis seedlings
-
综合各个指标进行多目标决策模型分析,在各因素取−2.0≤x≤2.0时,用计算机对模型进行模拟寻优,以寻求可促进栓皮栎生长量最大化的最佳水肥条件组合。得出当x1=1.9,x2=1.3,x3=0.8,x4=−0.9,即土壤含水率为79%,施氮量为215.3 mg·株−1,施磷量46.0 mg·株−1,施钾量为18.1 mg·株−1,苗木生长可达最优,苗高可达45.14 cm,地径达4.40 mm,根干质量达6.30 g,生物量达11.70 g,单株叶面积可达460.83 cm2。
-
土壤水分和养分(氮、磷、钾)是影响植物生长的重要因素且发挥着不同的作用[15-16],并与植物生理生长紧密相关,对植物的生物量积累、光合作用、营养分配和根系生长有着显著影响[17]。本研究中水分、氮肥和磷肥对苗木生长有着显著影响,并且水分大于施氮量和施磷量的影响,这与之前研究结论类似[18-20]。本研究中测定的5个指标均随着土壤含水率的增加而快速增长,当土壤含水率达到一定程度(70%)时,增长速率减缓。5个指标均随着施氮量的增加而显著增加,当施氮量达到一定水平时(大于225.0 mg·株−1),苗木生长减缓,各指标开始出现下降的趋势。磷肥大于45.0 mg·株−1,开始对苗木生物量产生抑制作用。5个指标均在较低水平的钾肥(22.5 mg·株−1)下达到最高值,但钾肥单因素影响较其他3个因素的影响不显著。这说明合理增施肥料可促进栓皮栎苗木生长发育,施肥量过高会导致土壤水分亏缺,此时再施加肥料会产生拮抗效应,引起水分胁迫,进而使苗木生长受阻。
-
水分与肥料之间的互作效应对栓皮栎苗木的生长影响显著。在一定范围内,土壤含水率×施氮量、土壤含水率×施磷量能产生正向协同效应。高水分和高氮肥有利于栓皮栎的生长,这与杨自立[13]的研究结果不同,可能是因为样地条件和种源地不同而导致的苗木间的差异。然而,水氮交互能够显著影响其他树种的生长状况,如土壤含水率×施氮量对楸树苗高、地径、叶面积、生物量有显著正效应,土壤含水率×施磷量对楸树苗高有显著正效应[7];土壤含水率×施氮量对尾巨桉Eucalyptus urophylla × E. grandis生物量具有显著正效应[21]。干旱环境中土壤养分会对植物生长产生互作影响,在水分亏缺的条件下,施肥可以提高植物的耐旱能力[22],如通过氮磷等养分的施入可以补偿小麦Triticum aestivum水分亏缺产生的不良反应,使生长抑制现象得到一定缓解[23]。但也有研究表明,施肥会加剧植物生长后期的水分胁迫[24]。本研究中,土壤含水率×施磷量除叶面积外,对其他4个指标均有显著交互正效应,且在低水分条件下,添加高磷肥比低磷肥更能抑制苗木生长。土壤含水率×施钾量对生物量有正向协同效应,低水分条件下施加氮肥可促进生物量积累,过量施加钾肥会明显产生拮抗作用。较高水分条件下,施加高氮肥、磷肥可促进苗木生长、根生物量和总生物量增加,低钾肥可促进地径增加和生物量的积累。原因是土壤水分增加时,土壤中的有机氮会矿化为更利于植物吸收的铵态氮和硝态氮,让有机态磷矿化为植物可以吸收的无机态磷,并增强钾元素的移动性。而土壤水分缺乏时,各营养元素进入根部会受到影响,各组织中的元素比例会失调,进而植物生长受到抑制[25]。随着水分的增加,适宜的施肥量能够很快促进苗木生长,而养分过量和亏缺都会对植株生长造成伤害。MARIS等[26]研究表明:土壤水分能促进植物对养分的吸收,且在很大程度上决定着养分的吸收,土壤中的养分只有溶解在水中才能被植物吸收,充足的灌溉能显著提高养分吸收的有效性。
-
除不同肥料与水分之间存在交互效应外,不同肥料之间也存在交互效应。水曲柳Fraxinus mandschurica苗木在氮、磷供给失衡的情况下,不能调整氮磷营养资源,单株总生物量显著下降[27]。枣树Ziziphus jujuba施肥试验也表明:钾肥可促进氮肥的增产,但两者之间存在拮抗效应[28]。此外,在低氮肥条件下,少量的磷肥可使叶面积、根干质量和总生物量达到最大,再继续添加磷肥会对苗木生长产生抑制作用,这与徐有明等[29]对湿地松Pinus elliottii幼林研究的结论类似。在较高水平的氮肥下,适当添加磷肥可促进叶面积、根干质量和总生物量的增加。钾肥与氮肥交互对地径有负效应,即随着氮肥的增加,继续添加钾肥会对地径产生显著抑制效应。原因可能是栓皮栎幼苗对钾肥需求量较小,栓皮栎种子中所含的营养元素已经能够满足其当年生长过程对钾的需求,因此再施用钾肥不会对地径生长产生促进作用。有研究发现:钾肥能缓解氮肥过量所引起的有害作用[30],这也可能是钾肥产生负效应的原因,但具体原因还有待进一步研究。由此可见,在栓皮栎苗期,应当给予苗木充足的水分,适宜的氮肥和磷肥,在此基础上施加少量钾肥即可满足苗木生长的需求。单一种类的肥料并不能满足苗木生长的需求,需要各种肥料混合以适当的配比进行使用,并供给苗期充足的水分,从而促进植物更好地吸收养分,提高苗木的生长量。
-
土壤水分和氮肥是影响栓皮栎苗木的主要因子,而土壤水分是限制苗木生长的最主要因素。高水和高氮肥可以促进苗木吸收磷和钾,总体而言钾肥对苗木的增产效应较小。因此,本研究综合考虑单因素以及因素之间的耦合效应,对模型进行多目标决策分析得到:栓皮栎苗木苗高、地径、单株叶面积、根干质量和生物量的最大值出现在高水(田间最大持水量的79%),高氮肥(215.3 mg·株−1),高磷肥(46.0 mg·株−1),低钾肥(18.1 mg·株−1)的条件下。
Coupling effects of soil water and fertilizer application on the growth of Quercus variabilis seedlings
-
摘要:
目的 探讨栓皮栎Quercus variabilis当年生苗木的苗高、地径、单株叶面积、根干质量、生物量积累对水肥耦合的响应规律,并确立最佳的灌溉施肥组合。 方法 采用4因素5水平二次回归通用旋转组合设计(RCCD),建立各指标与土壤含水率、施氮量、施磷量和施钾量回归模型,分析各因子的主效应、单因素和耦合效应对生长的影响。 结果 ①对苗高、地径、单株叶面积、根干质量、生物量的方差分析表明,各指标的不同处理间存在显著差异(P<0.05),处理8、处理16、处理18苗木生长情况较好。②模型检验结果表明:土壤含水率和施氮量对5个指标均有显著正效应,施磷量对地径无显著效应,施钾量对各指标均无显著效应,主效应从大到小依次为土壤含水率、施氮量、施磷量和施钾量。③单因素效应表明:各指标随着施肥量的增加均呈现出类似“抛物线”的变化趋势,生长速率随着土壤含水率的增加而增加,到一定程度时速率减缓。④耦合效应表明:土壤含水率×施氮量对苗高、生物量、叶面积、根干质量有显著正效应,土壤含水率×施磷量对苗高、地径、根干质量、生物量有显著正效应,土壤含水率×施钾量对生物量有显著正效应,施氮量×施磷量对叶面积、根干质量、生物量有显著正效应,施氮量×施钾量对地径有显著负效应,水肥间的耦合效应大于肥料间的耦合效应。 结论 栓皮栎苗期水肥需求量从大到小依次为土壤含水率、施氮量、施磷量和施钾量,适宜的水分和施肥配比可促进苗木生长。高水、高氮、高磷、低钾条件下苗木生长情况更好。水肥调控的最佳组合为:土壤含水率为田间最大持水量的79%,氮、磷、钾用量分别为215.3、46.0、18.1 mg·株−1,苗木的苗高可达到45.14 cm,地径达到4.40 mm,根干质量达到6.30 g,生物量达到11.70 g,单株叶面积可达460.83 cm2。图3表2参30 Abstract:Objective The aim is to investigate the response of stem height, ground diameter, leaf area, root weight and biomass accumulation of the seedlings of Quercus variabilis to the coupling of water and fertilizer, and to establish optimal combination of irrigation and fertilization. Method The four-factor-five-level quadratic regression general rotatable central composite design(RCCD) was employed to establish the regression model of each index and soil moisture (W), nitrogen (N), phosphorus (P) and potassium (K), with an analysis conducted of the main effect, monofactor and coupling effect of each factor on growth. Result (1)With the variance analysis of the stem height, ground diameter, leaf area, root weight and biomass, it was found that there is a significant divergence in the treatment of various factors (P<0.05) and the growth of seedling 8,16 and18 is the most favorable. (2) The model test results show that W and N have significant positive effect on all 5 indexes, P has no significant effect on ground diameter, K has no significant effect on any indexes, and there is a decrease of the main effect: W>N>P>K. (3) In terms of monofactor effect, it was shown that all the indexes presented a trend of first increase and then decrease with the increase of fertilizer application, in the path of a parabola. There was a gradual growth of the seedling with the increase of W before the a drop occurred in the growth rate. (4) With the anlaysis of the coupling effect, it was found that W×N has significant positive effects on stem height, root weight, biomass and leaf area, W×P has significant positive effects on stem height, ground diameter, root weight and biomass, W×K has significant positive effects on biomass, and N×P has significant positive effects on root weight, leaf area and biomass, N×K has significant negative effects on the ground diameter and the coupling effect between water and fertilizer is greater than that between fertilizers. Conclusion In conclusion, the water and fertilizer demand in the seedling stage of Q. variabilis is W>N>P>K, and suitable water and fertilizer ratio can promote the growth of seedlings. Seedlings grow better with conditions of high water, high nitrogen, high phosphorus and low potassium fertilizer. The optimal combination of water and fertilizer regulation was as follows: soil water content was 79% of the maximum field moisture capacity, nitrogen application was 215.3 mg·plant−1, phosphorus application was 46.0 mg·plant−1, potassium fertilizer application was 18.1 mg·plant−1, with seedling height reaching 45.14 cm, ground diameter reaching 4.40 mm, root weight reaching 6.30 g·plant−1,biomass reaching 11.70 g· plant−1, and leaf area reaching 460.83 cm2. [Ch, 3 fig. 2 tab. 30 ref.] -
图 1 土壤含水率、施氮量、施磷量、施钾量对栓皮栎苗高、地径、单株叶面积、根干质量、生物量单因素效应
Figure 1 Monofactor effects of soil water content, N, P and K fertilizers on stem height, ground diameter, root weight, biomass, leaf area of Q. variabilis seedlings
图 2 水肥耦合对栓皮栎苗木苗高、地径、叶面积、根干质量的影响
Figure 2 Coupling effects of water and fertilizeron stem height, ground diameter, leaf area, root weight of Q. variabilis seedlings
图 3 水肥耦合对栓皮栎苗木生物量的影响
Figure 3 Coupling effects of water and fertilizer on biomass of Q. variabilis seedlings
表 1 试验因素和水平编码值
Table 1. Coded and physical values of experimental factors
编码水平 土壤含水
率/%施氮量/
(mg·株−1)施磷量/
(mg·株−1)施钾量/
(mg·株−1)−2.0 40 0 0 0 −1.0 50 75.0 15.0 15.0 0 60 150.0 30.0 30.0 1.0 70 225.0 45.0 45.0 2.0 80 300.0 60.0 60.0  下载: 导出CSV
下载: 导出CSV
表 2 结构矩阵和各处理栓皮栎苗木生长情况
Table 2. Experimental design matrix and growth of Q. variabilis seedlings under different treatments
处理 x1 x2 x3 x4 苗高/cm 地径/mm 根干质量/g 生物量/g 叶面积/cm2 1 −1.0 −1.0 −1.0 −1.0 31.97±0.56 h 3.46±0.11 cd 3.45±0.03 k 6.69±0.18 de 291.93±3.41 g 2 1.0 −1.0 −1.0 −1.0 38.80±0.85 cd 3.82±0.20 bc 4.57±0.02 e 8.53±0.25 bc 364.24±7.36 de 3 −1.0 1.0 −1.0 −1.0 30.07±0.75 i 3.92±0.10 bc 3.84±0.03 i 6.90±0.23 de 274.35±4.79 h 4 1.0 1.0 −1.0 −1.0 40.20±0.77 c 4.15±0.17 ab 5.21±0.04 b 9.71±0.12 b 397.33±10.49 c 5 −1.0 −1.0 1.0 −1.0 29.47±0.73 i 3.42±0.13 d 3.66±0.02 j 6.47±0.41 de 275.37±5.81 h 6 1.0 −1.0 1.0 −1.0 39.03±0.92 cd 3.93±0.19 bc 4.74±0.05 d 9.18±0.10 bc 370.65±7.67 de 7 −1.0 1.0 1.0 −1.0 30.92±0.64 hi 3.83±0.16 bc 4.10±0.03 h 7.33±0.18 cd 298.80±10.18 g 8 1.0 1.0 1.0 −1.0 42.27±0.98 b 4.24±0.20 ab 5.81±0.04 a 10.79±0.11a 426.75±13.37 b 9 −1.0 −1.0 −1.0 1.0 28.90±0.76 i 3.72±0.10 c 3.48±0.03 k 6.18±0.17 de 259.70±11.61 i 10 1.0 −1.0 −1.0 1.0 37.00±0.91 e 4.10±0.09 ab 4.56±0.05 e 8.65±0.11 bc 360.34±5.49 e 11 −1.0 1.0 −1.0 1.0 31.30±0.54 hi 3.66±0.13 cd 3.80±0.03 i 6.82±0.16 de 276.32±9.52 h 12 1.0 1.0 −1.0 1.0 39.33±0.85 cd 4.03±0.14 b 5.29±0.02 b 9.95±0.18 ab 390.31±6.27 cd 13 −1.0 −1.0 1.0 1.0 30.77±0.66 hi 3.61±0.11 cd 3.20±0.02 l 5.88±0.39 e 249.13±6.11 i 14 1.0 −1.0 1.0 1.0 38.97±0.82 cd 4.16±0.17 ab 4.67±0.04 d 8.94±0.11 bc 367.71±11.04 de 15 −1.0 1.0 1.0 1.0 30.77±0.60 hi 3.45±0.12 cd 3.78±0.01 i 6.96±0.45 de 294.75±8.29 g 16 1.0 1.0 1.0 1.0 44.57±0.95 a 4.04±0.20 b 5.75±0.03 a 10.99±0.12 a 445.03±13.33 a 17 −2.0 0 0 0 26.03±0.67 j 3.44±0.14 cd 2.60±0.06 m 4.85±0.35 e 245.46±5.69 i 18 2.0 0 0 0 43.13±0.99 b 4.34±0.26 a 5.72±0.05 a 10.69±0.35 ab 439.87±11.94 ab 19 0 −2.0 0 0 33.27±0.74 g 3.61±0.17 cd 3.77±0.04 i 7.10±0.19 c 272.22±7.48 hi 20 0 2.0 0 0 36.37±0.70 ef 3.82±0.15 bc 4.87±0.04 c 8.85±0.18 bc 339.81±8.12 f 21 0 0 −2.0 0 31.53±0.64 h 3.88±0.16 bc 4.13±0.04 h 7.48±0.18 c 327.63±4.80 f 22 0 0 2.0 0 33.97±0.77 g 3.91±0.19 bc 4.50±0.03 ef 8.30±0.20 bc 334.72±10.71 f 23 0 0 0 −2.0 35.50±0.81 f 3.92±0.17 bc 4.42±0.04 f 8.07±0.19 bc 335.99±10.88 f 24 0 0 0 2.0 36.33±0.58 ef 3.88±0.17 bc 4.30±0.03 g 7.95±0.31 c 330.40±12.97 f 25 0 0 0 0 37.87±0.89 de 4.00±0.22 bc 4.67±0.03 de 8.84±0.18 bc 376.50±9.18 d 26 0 0 0 0 36.77±0.69 ef 4.10±0.15 ab 4.45±0.04 f 8.44±0.20 c 356.43±4.59 e 27 0 0 0 0 38.53±0.78 d 4.05±0.12 b 4.80±0.03 cd 9.05±0.19 bc 397.25±4.30 c 28 0 0 0 0 39.33±0.82 cd 4.00±0.16 bc 4.70±0.03 d 8.82±0.19 bc 394.06±12.09 c 29 0 0 0 0 37.80±0.70 de 3.90±0.13 bc 4.87±0.04 c 9.09±0.21 bc 386.09±9.43 cd 30 0 0 0 0 39.77±0.81 cd 3.91±0.09 bc 4.70±0.04 d 8.85±0.17 bc 394.45±6.87 c 说明:x1、x2、x3、x4分别表示土壤含水率、施氮量、施磷量和施钾量的水平编码值;表中数值为平均值±标准误;不同小写字母代 表各处理间苗木苗高、地径和生物量在0.05水平上差异显著
下载: 导出CSV
-
[1] WANG Yu, FU Delong, PAN Lulu, et al. The coupling effect of water and fertilizer on the growth of tea plants[Camellia sinensis (L.) O. Kuntz] [J]. J Plant Nutr, 2016, 39(5): 620 − 627. [2] HARTMANN A A, BARNARD R L, MARHAN S, et al. Effects of drought and N-fertilization on N cycling in two grassland soils [J]. Oecologia, 2013, 171(3): 705 − 717. [3] RAHBARI A, SINAKI J M, ZAREI M. Phosphate fertilizer and drought effects on growth components and elements of forage millet varieties [J]. Int J Agron Plant Prod, 2013, 4(2): 292 − 299. [4] DENIEL F C, GOMES D, DIONIZIO H, et al. Carrot yield and water-use efficiency under different mulching, organic fertilization and irrigation levels [J]. Revista Brasileira de Engenharia Agrícolae Ambiental, 2018, 22(7): 445 − 450. [5] 张小朋, 殷有, 于立忠, 等. 土壤水分与养分对树木细根生物量及生产力的影响[J]. 浙江农林大学学报, 2010, 27(4): 606 − 613. ZHANG Xiaopeng, YIN You, YU Lizhong, et al. Influence of water and soil nutrients on biomass and productivity of fine tree roots: a review [J]. J Zhejiang A&F Univ, 2010, 27(4): 606 − 613. [6] 王景燕, 唐海龙, 龚伟, 等. 水肥耦合对汉源花椒幼苗生长、养分吸收和肥料利用的影响[J]. 南京林业大学学报(自然科学版), 2016, 40(3): 33 − 40. WANG Jinyan, TANG Hailong, GONG Wei, et al. Effects of water and fertilizer coupling on growth, nutrients absorption and fertilizer use of Zanthoxylum bungeanum Maxim ‘Hanyuan’ seedling [J]. J Nanjing For Univ Nat Sci Ed, 2016, 40(3): 33 − 40. [7] QIU Quan, LI Jiyue, WANG Junhui, et al. Coupling effects of water and fertilizer on the growth characteristics of Catalpa bungei seedlings [J]. Pakistan J Bot, 2015, 47(3): 889 − 896. [8] 李迎超, 于海燕, 付甜, 等. 中国栓皮栎资源生产燃料乙醇的潜力及其空间分布[J]. 林业科学, 2013, 49(11): 129 − 134. LI Yinchao, YU Haiyan, FU Tian, et al. Spatial distribution and bio-ethanol potential of Quercus variabilis in China [J]. Sci Silv Sin, 2013, 49(11): 129 − 134. [9] 李志萍, 张文辉. NaCl胁迫对栓皮栎幼苗生长及其生理响应[J]. 西北植物学报, 2013, 33(8): 1630 − 1637. LI Zhiping, ZHANG Wenhui. Growth and physiological response of Quercus variabilis seedlings under NaCl stress [J]. Acta Bot Boreali-Occident Sin, 2013, 33(8): 1630 − 1637. [10] SUN Qiaoyu, DUNROESE R, LIU Yong. Container volume and subirrigation schedule influence, Quercus variabilis seedling growth and nutrient status in the nursery and field [J]. Scandinavian J For Res, 2018, 7: 1 − 8. [11] 李国雷, 祝燕, 蒋乐, 等. 指数施肥对栓皮栎容器苗生长和氮积累的影响[J]. 东北林业大学学报, 2012, 40(11): 6 − 9. LI Guolei, ZHU Yan, JIANG Le, et al. Effect of exponential fertilization on growth and nitrogen storage of containerized Quercus variabilis seedlings [J]. J Northeast For Univ, 2012, 40(11): 6 − 9. [12] 陈闯, 刘勇, 李国雷, 等. 底部渗灌灌水梯度对栓皮栎容器苗生长和养分状况的影响[J]. 林业科学, 2015, 51(7): 21 − 27. CHEN Chuang, LIU Yong, LI Guolei, et al. Effects of sub-irrigation gradients on growth and nutrient status of containerized seedlings of Quercus variabilis [J]. Sci Silv Sin, 2015, 51(7): 21 − 27. [13] 杨自立. 栓皮栎播种苗水氮需求规律研究[D]. 北京: 北京林业大学, 2011. YANG Zili. Study on the Water and Nitrogen Demand Characteristics of Quercus variabilis Seedlings[D]. Beijing: Beijing Forestry University, 2011. [14] 邱权, 李吉跃, 王军辉, 等. 水肥耦合效应对楸树苗期叶片净光合速率和SPAD值的影响[J]. 生态学报, 2016, 36(11): 3459 − 3468. QIU Quan, LI Jiyue, WANG Junhui, et al. Interactive effects of soil water and fertilizer application on leaf netphotosynthetic rate and SPAD readings of Catalpa bungei seedlings [J]. Acta Ecol Sin, 2016, 36(11): 3459 − 3468. [15] ABDELHALIM E, SERRET M D, ARAUS J L. Interactive effect of water and nitrogen regimes on plant growth, root traits and water status of old and modern durum wheat genotypes [J]. Planta, 2016, 244(1): 125 − 144. [16] GHASSEMI G K, RAEI N, RAEI Y. Effects of water deficit and nitrogen levels on grain yield and oil and protein contents of maize [J]. Azarian J Agric, 2015, 2: 46 − 50. [17] DONG Lei, LI Jiyue, WANG Junhui, et al. Effects of drought stress on osmotic regulation substances of five Catalpa bungei clones [J]. Agric Sci Technol, 2013, 14(9): 1335 − 1343. [18] 孙巧玉, 刘勇. 控释肥和灌溉方式对栓皮栎容器苗苗木质量及造林效果的影响[J]. 林业科学研究, 2018, 31(5): 137 − 144. SUN Qiaoyu, LIU Yong. Effect of controlled-release fertilizer and irrigation method on seedlingquality and outplanting performance of Quercus variabilis [J]. Forest Res, 2018, 31(5): 137 − 144. [19] 程中倩, 李国雷. 氮肥和容器深度对栓皮栎容器苗生长、根系结构及养分贮存的影响[J]. 林业科学, 2016, 52(4): 21 − 29. CHENG Zhongqian, LI Guolei. Effects of nitrogen supply and container size on seedling growth root characteristics and nutrient status in Quercus variabilis container seedlings [J]. Sci Silv Sin, 2016, 52(4): 21 − 29. [20] 屈明华, 俞元春, 李生, 等. 丛枝菌根真菌对矿质养分活化作用研究进展[J]. 浙江农林大学学报, 2019, 36(2): 394 − 405. QU Minghua, YU Yuanchun, LI Sheng, et al. Advances in research on activation of mineral nutrients by arbuscular mycorrhizal fungi [J]. J Zhejiang A&F Univ, 2019, 36(2): 394 − 405. [21] 林国祚, 谢耀坚, 彭彦, 等. 水肥耦合对尾巨桉苗木生物量的影响[J]. 桉树科技, 2013, 30(2): 1 − 8. LIN Guozuo, XIE Yaojian, PENG Yan, et al. Couple effects of water and fertilizer on the biomass of Eucalyptus urophylla×E. grandis seedlings [J]. Eucalypt Sci Technolo, 2013, 30(2): 1 − 8. [22] SINGH B, SINGH G. Effects of controlled irrigation on water potential, nitrogen uptake and biomass production in Dalbergia sissoo seedlings [J]. Environ Exp Bot, 2006, 55(1/2): 209 − 219. [23] GOWING J W, ROSE D A, GHAMARNIA H. The effect of salinity on water productivity of wheat under deficit irrigation above shallow groundwater [J]. Agric Water Manage, 2009, 96(3): 517 − 524. [24] CABELLO M J, CASTELLANOS M T, ROMOJARO F, et al. Yield and quality of melon grown under different irrigation and nitrogen rates [J]. Agric Water Manage, 2009, 96(5): 866 − 874. [25] 王梓, 马履一, 贾忠奎, 等. 滴灌下欧美107杨苗高水肥耦合效应[J]. 浙江农林大学学报, 2012, 29(1): 29 − 34. WANG Zi, MA Lüyi, JIA Zhongkui, et al. Interaction of water and fertilizer on seedling height for first-year growth Populus×euramericana ‘74/76’ with drip irrigation [J]. J Zhejiang A&F Univ, 2012, 29(1): 29 − 34. [26] MARIS S C, TERIA-ESMATGES M R, ARBONES A, et al. Effects of irrigation, nitrogen application, and a nitrification inhibitor on nitrous oxide, carbon dioxide and methane emissions from an olive (Olea europaea L.) orchard [J]. Sci Total Environ, 2015, 538(15): 966 − 978. [27] 吴楚, 王政权, 范志强. 氮素形态处理下水曲柳幼苗养分吸收利用与生长及养分分配与生物量分配的关系[J]. 生态学报, 2005, 25(6): 1282 − 1290. WU Chu, WANG Zhenquan, FAN Zhiqiang. Relationships between nutrient uptake, use efficiency and growth, and between nutrient partitioning patterns and biomass partitioning patterns in Fraxinus mandshurica seedlings supplied with different ratios of nitrogen forms [J]. Acta Ecol Sin, 2005, 25(6): 1282 − 1290. [28] 柴仲平, 王雪梅, 孙霞, 等. 不同氮磷钾配比滴灌对灰枣产量与品质的影响[J]. 果树学报, 2011, 28(2): 229 − 233. CHAI Zhongping, WANG Xuemei, SUN Xia, et al. Influence of N, P, K with drip irrigation on yield and fruit quality of Huizao jujube [J]. J Fruit Sci, 2011, 28(2): 229 − 233. [29] 徐有明, 林汉, 洪院生, 等. 施肥对湿地松幼林生长和木材物理力学性质的影响[J]. 林业科学, 2002, 38(4): 125 − 133. XU Youming, LIN Han, HONG Yuansheng, et al. Effects of fertilization on growth increments and wood physical mechanical properties of young slash pine [J]. Sci Silv Sin, 2002, 38(4): 125 − 133. [30] YAN Xiaoli, JIA Liming, DAI Tengfei, et al. Effects of water and nitrogen coupling under drip irrigation on tree growth and soil nitrogen content of Populus×euramericana cv. ‘Guariento’ [J]. Chin J Appl Ecol, 2018, 29(7): 2159 − 2202. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190456
 点击查看大图
点击查看大图
计量
- 文章访问数: 2162
- HTML全文浏览量: 589
- PDF下载量: 67
- 被引次数: 0






