




-
小贯小绿叶蝉 Empoasca onukii在中国茶园广泛分布,是对茶树Camellia sinensis危害最大的害虫之一[1]。小贯小绿叶蝉对茶树嫩茎、嫩叶的刺吸会导致茶叶焦枯,抑制茶树正常生长,严重影响茶叶的产量和品质[2−3]。游猎性蜘蛛(游猎蛛)作为一种捕食性天敌,捕食量大、行动活跃、捕食范围广,对防治害虫、调节生态系统平衡都有重要作用[4−6]。利用天敌与害虫之间的捕食关系对茶园害虫进行生物防治的方法已有诸多报道[7−16],这些研究大都直接讨论天敌与害虫之间的关联,对于不同种天敌之间的竞争作用却极少研究。但单一地增加茶园害虫的优势种天敌等势必会导致整个茶园生态系统的不稳定[17−18]。FAUST等[19]将2个物种之间的作用关系分为中性、竞争、偏害、寄生、捕食、偏利和互利共生等几种类型。天敌和害虫之间的关系分为捕食和寄生2种类型,天敌之间为了争夺资源多数是竞争关系[20]。除了天敌与害虫之间的捕食和被捕食关系外,各天敌之间的竞争关系也需要得到重视。因此,从生物多样性等多维度对茶园整个生态环境进行调控更有利于长期高效防治茶园害虫,以达到绿色环保的平衡调节作用[21]。
在昆虫种群数量生态学的研究中,对害虫及天敌数据的统计分析通常为抽样分析,采用的分级方式为连续等分分级,这会使得不同精度级宽下的结果具有主观性和偶然性。张书平等[22]将Fuzzy分级法与灰色关联度法相结合研究茶园与假眼小绿叶蝉 Empoasca vitis数量上关系密切的天敌种类,是一种较好的种间关系研究方法。多数学者对林木种间竞争的研究较多[23],未见对茶园各天敌种间竞争关系的报道,将数据按不同方式分级后再研究种间竞争关系的研究更少。为此,本研究运用灰色关联度法,并将Fuzzy分级法与竞争系数法相结合,研究‘农抗早’‘Nongkangzao’和‘平阳特早’‘Pingyangtezao’茶园游猎蛛取食小贯小绿叶蝉的种间竞争关系,并通过竞争强度指数验证所得结论,为茶园合理保护和利用种间竞争关系强的天敌,有效防治小贯小绿叶蝉提供科学依据。
-
在安徽农业大学科技示范园(31°56′N,117°12′E)中共调查了2种茶园,分别为‘农抗早’和‘平阳特早’茶园,面积均为0.2 hm2。调查时间为2021年3月25日至11月19日,隔15 d调查1次,共14次。茶园互不相连,按常规措施管理。在春茶采摘结束和秋末进行茶树修剪,并且在秋末进行茶园耕翻,加强秋冬季管理,及时除草、修剪茶树和采摘茶叶,诱杀和人工捕杀害虫,不施用化学农药。
-
采用平行跳跃法在茶园随机选取3行,每行茶树相隔1 m,在每行间隔1 m处设置1个2 m×1 m的样方,每行取10个样方,每个茶园取30个样方。用目测调查的方式在每个样方随机选取10片叶,调查一些不易振落的害虫及天敌的种类和数量,再用盘拍法对样方中所有枝条进行盘拍(盘拍所用搪瓷盘口长为40 cm,宽30 cm,搪瓷盘上喷洒稀释1 000倍的洗衣粉水溶液)。调查记载盘中害虫及天敌物种数和个体数,不能准确鉴定的物种样本编号保存,装瓶带回室内由专家鉴定。
-
将害虫数量(Yi)及其主要天敌数量(Xj)分别看作1个本征系统,害虫数量作为该系统的参照序列,天敌作为比较序列,并把不同样方的害虫及其主要天敌的数量作为该序列在第k个样方的效果白化值,进行双序列关系分析。数据经均值化后得:
$$ Y_i=\left\{Y_i(1),\; Y_i(2),\; \cdots ,\; Y_i(n)\right\},\; i=1; $$ (1) $$ X_j=\left\{X_j(1),\; X_j(2),\; \cdots,\; X_j(n)\right\}, \;j=1,\;2,\; \cdots,\; M。 $$ (2) 式(1)和式(2)中:n表示样方数,M表示天敌种类数。Yi与Xj在第k个样方上的关联系数rij为:
$$ {r}_{ij}=\frac{{\mathrm{min}}_{i}{\mathrm{min}}_{j}\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right|+\rho {\mathrm{max}}_{i}{\mathrm{max}}_{j}\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right|}{\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right|+\rho {\mathrm{max}}_{i}{\mathrm{max}}_{j}\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right|} 。 $$ (3) 式(3)中:ρ 为分辨系数,取值区间为[0, 1],一般取 ρ = 0.5,为扩大各物种之间关联度的差异,本研究取 ρ = 0.8。$|Y_{i}\left(k\right)-{X}_{j}\left(k\right) |$ 为序列Yi与Xj在第k点上差的绝对值;$ \mathrm{m}\mathrm{i}\mathrm{n}\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right| $为1级最小差,$ {\mathrm{min}}_{i}{\mathrm{min}}_{j}\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right| $ 为2级最小差。$ \mathrm{m}\mathrm{a}\mathrm{x}\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right| $与$ {\mathrm{max}}_{i}{\mathrm{max}}_{j}\left|{Y}_{i}\left(k\right)-{X}_{j}\left(k\right)\right| $ 分别为1级和2级最大差。利用该公式可求出第j种天敌(Xj)与害虫(Yi)数量间的关联度为$R({Y}_{i},{X}_{j})=1/n\displaystyle \sum _{k=1}^{M}{r}_{ij}\left(k\right)$,其大小反映害虫与天敌相互联系的紧密程度。天敌与害虫数量间关联度越大,表明天敌与害虫数量关系越密切[24]。本研究由数据处理系统软件(DPS系统)进行灰色关联度数据的运算。
-
在论域[A1, A2, A3$, \;\cdots , $ An$, \;\cdots , $ Am]上按所求解问题的性质和要求规定的一个隶属函数μi,叫作Fuzzy分级隶属函数。将通常连续等分分级的频数作为原始数据,设原始数据为[a1, a2, a3$,\; \cdots , $ an$, \;\cdots , $ am],称${\hat{{a}}}_{{i}}=\displaystyle \sum _{{n}={i}-5}^{{i}+5}{{\mu }}_{{i}}{{a}}_{{n}}$为第i个Fuzzy等级的Fuzzy频数。它在论域上的分布曲线叫Fuzzy频数曲线[25−26]。本研究规定Fuzzy分级隶属函数μi为:
$$ {\mu }_{i}=\left\{\begin{array}{l}1.0\;n=i\\ 0.8\;n=i+1,\;i-1\\ 0.6\;n=i+2,\;i-2\\ 0.4\;n=i+3,\;i-3\\ 0.2\;n=i+5,\;i-5\\ 0.1\;n=i+5,\;i-5\\ 0\;n > i+5,\;n < i-5\end{array}\right. 。 $$ (4) 式(4)中:n为原始数据项数,i为Fuzzy频数项数。则Fuzzy频数为$ {\hat{a}}_{i}=\displaystyle \sum _{n=i-5}^{i+5}{\mu }_{i}{a}_{n}={a}_{i}+ 0.8\left({a}_{i+1}+{a}_{i-1}\right)+0.6\left({a}_{i+2}+{a}_{i-2}\right)+0.4\left({a}_{i+3}+{a}_{i-3}\right)+0.2\left({a}_{i+4}+{a}_{i-4}\right)+0.1\left({a}_{i+5}+{a}_{i-5}\right) $。为了解茶园游猎蛛的自然种群动态,减少在对游猎蛛进行竞争关系分析时由种群数据导致的误差,本研究根据游猎蛛种群数量变化幅度,对游猎蛛的种群数据按照30个样方中5只游猎蛛的级宽进行等分分级统计(级宽不宜太宽也不宜太窄),算出各级出现的频数,再以30个样方中5只游猎蛛为级宽的频数作为原始数据即[a1, a2, a3$,\; \cdots , $ an$, \;\cdots , $ am]进行Fuzzy分级统计。由Excel 2019计算Fuzzy频数。
-
一般用Levins的生态重叠公式计算竞争系数[27]:
$$ {\alpha }_{ij}=\sum _{k=1}^{n}\left({P}_{ik}{P}_{jk}\right)/\sum _{k=1}^{n}{P}_{ik}^{2} 。 $$ (5) 式(5)中:$ {P}_{ik} $ 和$ {P}_{jk} $分别为种i和种j在第k个样方中的相对优势度,n为样地数。
使用重叠的方法计算竞争系数的公式[28]:
$$ {\alpha }_{ij}=\sum _{k=1}^{n}\left({P}_{ik}{P}_{jk}\right)/\sqrt{\sum _{k=1}^{n}{P}_{ik}^{2}\sum _{k=1}^{n}{P}_{jk}^{2}} 。 $$ (6) 式(6)中:αij为种j对种i的竞争系数,Pik和Pjk分别是第i个物种和第j个物种使用的第k个资源的比例。αij = αji,PIANKA[28]评价该式是对称的,并称其为重叠值。MAY[29−30]将该式与其他表达式进行比较,沿用了PIANKA的表达式,并将其作为按函数比例进行计算的竞争系数。本研究中所求得的竞争系数均按上述(6)式计算。
-
本研究将关联度与竞争系数相结合,引入竞争强度指数的概念。不同天敌与同一害虫之间关联度的比值称为相对密切度,该相对密切度与2种天敌之间竞争系数的乘积即为竞争强度指数,种i对种j的竞争强度指数(C)为:
$$ C={\alpha }_{ij}\left(\frac{{R}_{iy}}{{R}_{jy}}\right) 。 $$ (7) 式(7)引入了害虫与天敌关系因素,Riy、Rjy分别为天敌Xi、Xj与害虫Y数量上的关联度,αij为种j对种i的竞争系数。本研究中竞争系数和竞争强度指数均由 Excel 2019 对Fuzzy频数计算所得,再使用DPS数据处理系统软件用多重比较方法中的 Duncan 新复极差法进行竞争系数和竞争强度指数平均值和显著水平的分析[31]。
-
选取调查日期中2个茶园游猎蛛数量最多的7种蜘蛛作为主要天敌,分别为鞍型花蟹蛛Xysticus ephippiatus、三突花蟹蛛Ebrechtella tricuspidata、粽管巢蛛Clubiona japonicola、斑管巢蛛C. reichlini、斜纹猫蛛Oxyopes sertatus、黑色跳蛛Plexippus paykulli和条纹蝇虎P. setipes。它们的数量动态见表1。
表 1 2个茶园小贯小绿叶蝉与游猎蛛数量动态
Table 1. Population dynamics of E. onukii and wandering spiders in two tea plantations
茶树品种 日期
(月-日)害虫数量/头 游猎蛛数量/头 X1 X2 X3 X4 X5 X6 X7 ‘农抗早’ 03-25 3 10 16 3 2 32 7 7 04-16 11 2 7 1 0 7 0 19 05-07 42 2 6 0 0 2 2 0 05-23 30 1 5 0 0 5 12 1 06-04 222 7 1 5 0 1 5 0 06-20 175 20 5 5 5 0 5 0 07-08 594 14 65 51 88 6 38 0 08-10 209 27 23 15 60 20 26 24 08-22 429 23 19 29 59 33 25 6 09-06 92 19 9 19 23 57 18 6 09-17 30 17 17 13 10 31 13 19 10-13 1 8 1 2 0 45 4 5 11-01 52 4 2 9 9 39 4 19 11-19 47 1 1 3 6 15 3 5 合计 1 937 310 250 155 177 253 155 262 ‘平阳特早’ 03-25 9 2 3 0 0 13 17 4 04-16 11 0 5 0 0 1 6 2 05-07 31 1 6 0 0 0 6 0 05-23 57 3 4 1 0 0 5 0 06-04 251 1 5 0 3 0 11 0 06-20 139 15 8 18 2 0 10 1 07-08 674 26 46 24 34 22 7 0 08-10 318 17 3 17 42 33 13 0 08-22 534 8 18 21 48 31 10 29 09-06 160 29 12 14 18 39 16 10 09-17 20 13 12 12 19 37 12 9 10-13 4 7 2 0 0 20 11 2 11-01 23 5 2 8 3 2 2 3 11-19 108 1 2 3 5 26 2 4 合计 2 339 245 230 128 128 170 118 174 说明:X1~X7分别指鞍型花蟹蛛、三突花蟹蛛、粽管巢蛛、斑管巢蛛、斜纹猫蛛、黑色跳蛛和条纹蝇虎数量(头)。 用灰色关联度法求得小贯小绿叶蝉与游猎蛛数量之间的灰色关联度(表2)。由表2可知:与小贯小绿叶蝉在数量上关联度最大的前3位天敌,‘农抗早’茶园为斑管巢蛛、粽管巢蛛和黑色跳蛛;‘平阳特早’茶园为斑管巢蛛、粽管巢蛛和三突花蟹蛛。2个茶园均有斑管巢蛛和粽管巢蛛。
表 2 2个茶园小贯小绿叶蝉与游猎蛛间的灰色关联度
Table 2. Grey correlation between E. onukii and wandering spiders in two tea plantations
游猎蛛 ‘农抗早’ ‘平阳特早’ 游猎蛛 ‘农抗早’ ‘平阳特早’ 灰色关联度 排位 灰色关联度 排位 灰色关联度 排位 灰色关联度 排位 鞍型花蟹蛛 0.833 8 5 0.826 1 4 斜纹猫蛛 0.754 8 6 0.794 7 6 三突花蟹蛛 0.856 9 4 0.856 2 3 黑色跳蛛 0.857 0 3 0.802 6 5 粽管巢蛛 0.890 2 2 0.857 1 2 条纹蝇虎 0.744 2 7 0.763 4 7 斑管巢蛛 0.896 7 1 0.860 3 1 -
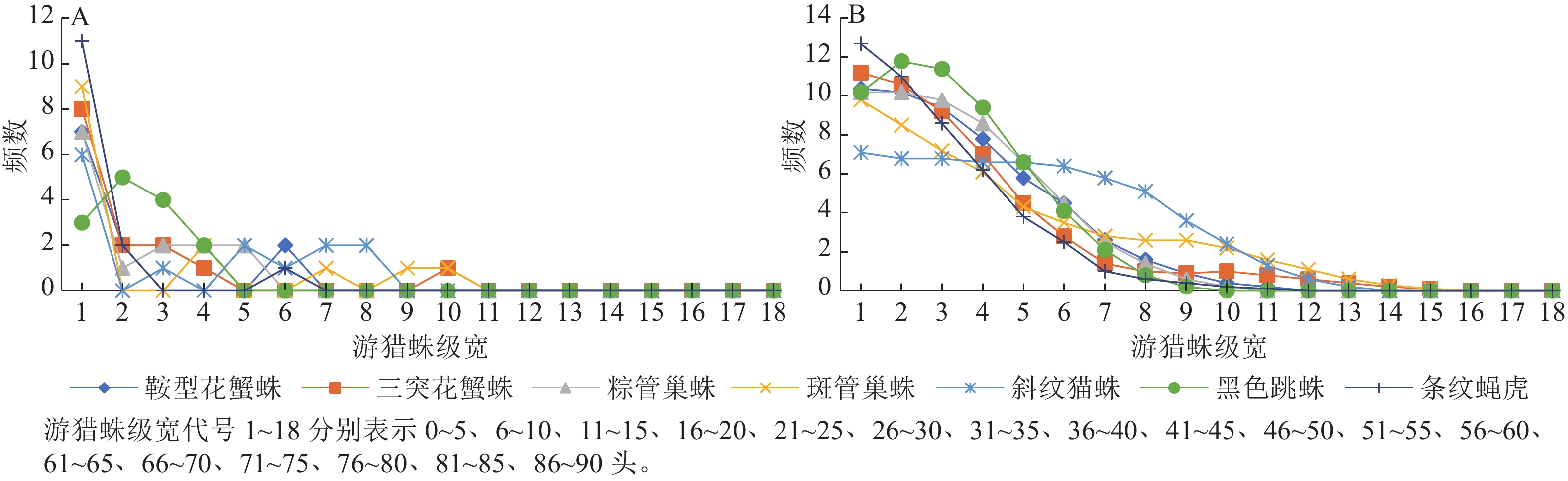
由图1和图2可看出:2个茶园Fuzzy分级频数集中性明显大于等分分级频数。图1A中在第4、7和12个级宽处出现3个明显的峰值,而图1B中 7条曲线均由高到低一致趋于平缓。图2A中从第11个级宽处多数曲线已贴近横轴,而图2B中的曲线从第15个级宽开始才趋于横轴。通过Fuzzy分级法得到的新数据明显增加了原始数据的有效区间,并且Fuzzy频数由原始数据通过Fuzzy隶属函数求得,每个原始数据都与几个等级发生联系,而等分分级里每个原始数据只与1个等级发生联系,因此Fuzzy频数无明显的分级界限。对2种频数进行t检验,‘农抗早’茶园中鞍型花蟹蛛、三突花蟹蛛、粽管巢蛛、斑管巢蛛、斜纹猫蛛、黑色跳蛛和条纹蝇虎这7种蜘蛛的t值依次为2.333、2.379、2.281、2.646、3.937、3.059、1.975,‘平阳特早’茶园这7种蜘蛛的$ t $值依次为 2.176、2.048、2.153、2.477、3.151、2.090、1.591。自由度为34时,t0.01=2.728,t0.05=2.032,t0.10=1.091,除条纹蝇虎外,2个茶园各游猎蛛2种频数间的差异都显著,并且斜纹猫蛛的2种频数间差异极显著。Fuzzy频数的直接图示是曲线,可保持原始数据的精度,故用此法分级的数据进行后续竞争关系的分析,其结果更接近实际。
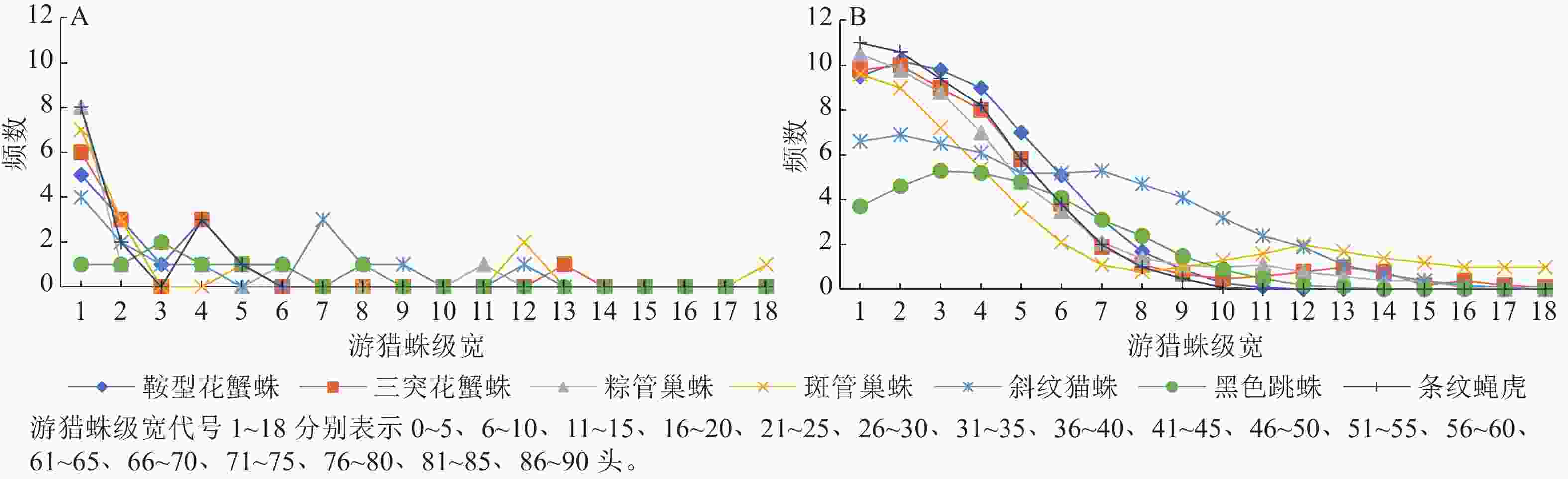
图 1 ‘农抗早’茶园游猎蛛种群数量等分分级频数曲线(A)和Fuzzy频数曲线(B)
Figure 1. Equally graded frequency curve (A) and Fuzzy frequency curve (B) of the number of wandering spiders in ‘Nongkangzao’ tea plantation
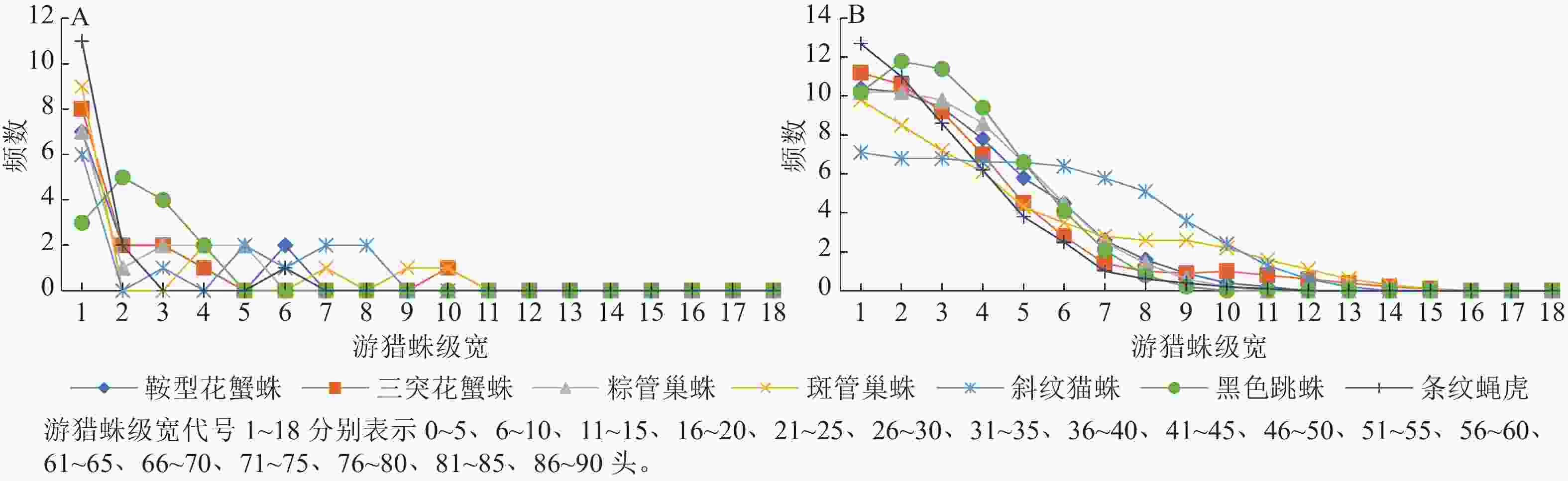
图 2 ‘平阳特早’茶园游猎蛛种群数量等分分级频数曲线(A)和Fuzzy频数曲线(B)
Figure 2. Equally graded frequency curve (A) and Fuzzy frequency curve (B) of the number of wandering spiders in ‘Pingyangtezao’ tea plantation
-
以Fuzzy频数作为原始数据,计算2个茶园各游猎蛛之间的竞争系数并将结果列于表3。对各游猎蛛及其竞争对手之间的竞争系数进行方差分析,再用新复极差法分析天敌之间竞争系数的差异性(表4)。所得结果中,当竞争对手分别为鞍型花蟹蛛、三突花蟹蛛、粽管巢蛛和条纹蝇虎时,游猎蛛间差异显著,此时不同竞争对手下竞争力最强的游猎蛛不同,故暂时无法得出茶园竞争力最强的蜘蛛类别,但除斑管巢蛛和黑色跳蛛外,无论竞争对手为哪种游猎蛛,所有游猎蛛都与斜纹猫蛛差异显著,且斜纹猫蛛的均值都最小。因此,可得出结论:除斑管巢蛛和黑色跳蛛外,5种游猎蛛里斜纹猫蛛竞争力最弱。
表 3 2个茶园各游猎蛛之间的竞争系数
Table 3. Competition coefficients among wandering spiders in two tea plantations
茶树品种 竞争对手 游猎蛛间竞争系数 鞍型花蟹蛛 三突花蟹蛛 粽管巢蛛 斑管巢蛛 斜纹猫蛛 黑色跳蛛 条纹蝇虎 ‘农抗早’ 鞍型花蟹蛛 1.000 0 0.991 5 0.983 7 0.939 8 0.906 3 0.959 2 0.991 0 三突花蟹蛛 0.991 5 1.000 0 0.995 2 0.973 3 0.893 8 0.928 2 0.995 3 粽管巢蛛 0.983 7 0.995 2 1.000 0 0.980 3 0.901 5 0.916 7 0.993 6 斑管巢蛛 0.939 8 0.973 3 0.980 3 1.000 0 0.872 9 0.856 8 0.961 4 斜纹猫蛛 0.906 3 0.893 8 0.901 5 0.872 9 1.000 0 0.959 8 0.871 7 黑色跳蛛 0.959 2 0.928 2 0.916 7 0.856 8 0.959 8 1.000 0 0.915 9 条纹蝇虎 0.991 0 0.995 3 0.993 6 0.961 4 0.871 7 0.915 9 1.000 0 ‘平阳特早’ 鞍型花蟹蛛 1.000 0 0.989 3 0.998 5 0.976 5 0.913 7 0.993 5 0.975 1 三突花蟹蛛 0.989 3 1.000 0 0.984 2 0.977 4 0.869 7 0.984 4 0.992 5 粽管巢蛛 0.998 5 0.984 2 1.000 0 0.967 4 0.909 3 0.996 3 0.967 4 斑管巢蛛 0.976 5 0.977 4 0.967 4 1.000 0 0.939 7 0.954 0 0.957 1 斜纹猫蛛 0.913 7 0.869 7 0.909 3 0.939 7 1.000 0 0.880 6 0.826 5 黑色跳蛛 0.993 5 0.984 4 0.996 3 0.954 0 0.880 6 1.000 0 0.967 7 条纹蝇虎 0.975 1 0.992 5 0.967 4 0.957 1 0.826 5 0.967 7 1.000 0 表 4 2个茶园各游猎蛛之间竞争系数新复极差法分析结果
Table 4. Results of the new multiple range test analysis of competition coefficients among various wandering spiders in two tea plantations
竞争对手 F 物种 2个茶园竞争
系数均值5%显著
水平1%极显著
水平竞争对手 F 物种 2个茶园竞争
系数均值5%显著
水平1%极显著
水平鞍型花蟹蛛 7.530 粽管巢蛛 0.991 1 a A 斜纹猫蛛 1.203 黑色跳蛛 0.920 2 a A 三突花蟹蛛 0.990 4 a A 鞍型花蟹蛛 0.910 0 a A 条纹蝇虎 0.983 0 a A 斑管巢蛛 0.906 3 a A 黑色跳蛛 0.976 4 a A 粽管巢蛛 0.905 4 a A 斑管巢蛛 0.958 2 a AB 三突花蟹蛛 0.881 7 a A 斜纹猫蛛 0.910 0 b B 条纹蝇虎 0.849 1 a A 三突花蟹蛛 11.356 条纹蝇虎 0.993 9 a A 黑色跳蛛 0.562 鞍型花蟹蛛 0.976 4 a A 鞍型花蟹蛛 0.990 4 a A 粽管巢蛛 0.956 5 a A 粽管巢蛛 0.989 7 a A 三突花蟹蛛 0.956 3 a A 斑管巢蛛 0.975 4 a A 条纹蝇虎 0.941 8 a A 黑色跳蛛 0.956 3 a A 斜纹猫蛛 0.920 2 a A 斜纹猫蛛 0.881 7 b B 斑管巢蛛 0.905 4 a A 粽管巢蛛 3.305 鞍型花蟹蛛 0.991 1 a A 条纹蝇虎 12.037 三突花蟹蛛 0.993 9 a A 三突花蟹蛛 0.989 7 a A 鞍型花蟹蛛 0.983 0 a A 条纹蝇虎 0.980 5 a A 粽管巢蛛 0.980 5 a A 斑管巢蛛 0.973 9 a A 斑管巢蛛 0.959 2 a A 黑色跳蛛 0.956 5 ab A 黑色跳蛛 0.941 8 a A 斜纹猫蛛 0.905 4 b A 斜纹猫蛛 0.849 1 b B 斑管巢蛛 1.610 三突花蟹蛛 0.975 3 a A 粽管巢蛛 0.973 9 a A 条纹蝇虎 0.959 2 a A 鞍型花蟹蛛 0.958 2 a A 斜纹猫蛛 0.906 3 a A 黑色跳蛛 0.905 4 a A 说明:计算不同竞争对手下各游猎蛛在2个茶园竞争系数的平均值,5%水平上均数最大的标记为a,1%水平上均数最大的标记为A。向下比较,与之差异性不显著的标记相同字母,差异性显著的标记不同字母。 -
结合关联度和竞争系数将所求的竞争强度指数列于表5。对竞争强度指数进行方差分析,用新复极差法进行比较(表6)。由表6可知:无论竞争对手为哪种游猎蛛,粽管巢蛛都与其他蜘蛛差异显著,其次为斑管巢蛛,且游猎蛛中斜纹猫蛛与两者差异极显著。因此,可得出结论:粽管巢蛛竞争力最强,斑管巢蛛次之,斜纹猫蛛竞争力最弱。
表 5 2个茶园各游猎蛛之间的竞争强度指数
Table 5. Competition intensity indices among wandering spiders in two tea plantations
茶树品种 竞争对手 游猎蛛间竞争强度指数 鞍型花蟹蛛 三突花蟹蛛 粽管巢蛛 斑管巢蛛 斜纹猫蛛 黑色跳蛛 条纹蝇虎 ‘农抗早’ 鞍型花蟹蛛 1.000 0 1.019 0 1.050 3 1.010 7 0.820 4 0.985 9 0.884 5 三突花蟹蛛 0.964 7 1.000 0 1.033 8 1.018 5 0.787 3 0.928 3 0.864 3 粽管巢蛛 0.921 4 0.958 0 1.000 0 0.987 5 0.764 4 0.882 5 0.830 6 斑管巢蛛 0.873 9 0.930 1 0.973 2 1.000 0 0.734 8 0.818 9 0.797 8 斜纹猫蛛 1.001 2 1.014 7 1.063 2 1.037 0 1.000 0 1.089 8 0.859 4 黑色跳蛛 0.933 2 0.928 1 0.952 2 0.896 5 0.845 3 1.000 0 0.795 3 条纹蝇虎 1.110 4 1.146 1 1.188 6 1.158 5 0.884 2 1.054 8 1.000 0 ‘平阳特早’ 鞍型花蟹蛛 1.000 0 1.025 4 1.036 1 1.017 0 0.879 0 0.965 3 0.9012 三突花蟹蛛 0.954 4 1.000 0 0.985 2 0.982 0 0.807 2 0.922 7 0.884 9 粽管巢蛛 0.962 3 0.983 2 1.000 0 0.971 0 0.843 1 0.932 9 0.861 7 斑管巢蛛 0.937 6 0.972 8 0.963 8 1.000 0 0.868 1 0.890 0 0.849 3 斜纹猫蛛 0.949 7 0.937 0 0.980 7 1.017 2 1.000 0 0.889 3 0.794 0 黑色跳蛛 1.022 6 1.050 2 1.064 0 1.022 6 0.872 0 1.000 0 0.920 5 条纹蝇虎 1.055 1 1.113 1 1.086 1 1.078 5 0.860 4 1.017 3 1.000 0 表 6 2个茶园各游猎蛛之间的竞争强度指数新复极差法分析结果
Table 6. Results of the new multiple range test analysis of competitive intensity indices among various wandering spiders in two tea plantations
竞争对手 F 物种 2个茶园竞争
强度指数均值5%显著
水平1%极显著
水平竞争对手 F 物种 2个茶园竞争
强度指数均值5%显著
水平1%极显著
水平鞍型花蟹蛛 32.992 粽管巢蛛 1.043 2 a A 斜纹猫蛛 2.142 斑管巢蛛 1.027 1 a A 三突花蟹蛛 1.022 2 ab A 粽管巢蛛 1.022 0 a A 斑管巢蛛 1.013 8 ab A 黑色跳蛛 0.989 5 ab A 黑色跳蛛 0.975 6 b A 三突花蟹蛛 0.975 8 ab A 条纹蝇虎 0.892 9 c B 鞍型花蟹蛛 0.975 4 ab A 斜纹猫蛛 0.849 7 c B 条纹蝇虎 0.826 7 b A 三突花蟹蛛 33.492 粽管巢蛛 1.009 5 a A 黑色跳蛛 1.575 粽管巢蛛 1.008 1 a A 斑管巢蛛 1.000 2 a AB 三突花蟹蛛 0.989 1 a A 鞍型花蟹蛛 0.960 9 ab AB 鞍型花蟹蛛 0.977 9 a A 黑色跳蛛 0.925 5 b BC 斑管巢蛛 0.959 6 a A 条纹蝇虎 0.874 6 c C 斜纹猫蛛 0.858 6 a A 斜纹猫蛛 0.797 3 d D 条纹蝇虎 0.857 9 a B 粽管巢蛛 9.706 斑管巢蛛 0.979 3 a A 条纹蝇虎 10.533 粽管巢蛛 1.137 3 a A 三突花蟹蛛 0.970 6 a AB 三突花蟹蛛 1.129 6 a A 鞍型花蟹蛛 0.941 9 a AB 斑管巢蛛 1.118 5 a A 黑色跳蛛 0.907 7 ab ABC 鞍型花蟹蛛 1.082 7 a A 条纹蝇虎 0.846 2 bc BC 黑色跳蛛 1.036 1 a A 斜纹猫蛛 0.803 8 c C 斜纹猫蛛 0.872 3 b B 斑管巢蛛 3.588 粽管巢蛛 0.968 5 a A 三突花蟹蛛 0.951 4 ab A 鞍型花蟹蛛 0.905 8 abc A 黑色跳蛛 0.854 4 abc A 条纹蝇虎 0.823 6 bc A 斜纹猫蛛 0.801 5 c A 说明:计算不同竞争对手下各游猎蛛在2个茶园竞争强度指数的平均值,5%水平上均数最大的标记为a,1%水平上均数最大的标记为A。向下比较,与之差异性不显著的标记相同字母,差异性显著的标记不同字母。 -
本研究首先通过灰色关联度分析初步得出2个茶园与小贯小绿叶蝉数量相关性最大的游猎蛛均为斑管巢蛛和粽管巢蛛,再对7种游猎蛛的种群数量进行Fuzzy分级统计,将得出的Fuzzy频数作为原始数据进行竞争系数分析,结果显示:除斑管巢蛛和黑色跳蛛外,5种游猎蛛中斜纹猫蛛竞争力最弱。为验证结果准确性,综合灰色关联度和竞争系数结果引入了竞争强度指数概念,得出7种游猎蛛中斜纹猫蛛竞争力最弱,并且在任何竞争对手下粽管巢蛛都与其他蜘蛛差异显著,其次为斑管巢蛛,即在取食茶园小贯小绿叶蝉时粽管巢蛛和斑管巢蛛竞争力最强,斜纹猫蛛竞争力最弱。
在进行Fuzzy分级统计时,并未对小贯小绿叶蝉数量进行分级,因为在进行竞争关系分析时,只需要7种游猎蛛的数量数据,故无需对小贯小绿叶蝉数量做同样的处理。在进行灰色关联度分析时,小贯小绿叶蝉和7种游猎蛛数量均未进行处理,原因是小贯小绿叶蝉数量与7种游猎蛛数量数据大小相差较大,选择过大的级宽会导致游猎蛛数量均处于第1级宽内,选择较小级宽会出现较多级层且在靠后多个级层里只有小贯小绿叶蝉数据,而游猎蛛数据均为0。对7种游猎蛛的种群数量进行Fuzzy分级,然后进行竞争关系的统计计算,使它们的数据集中性更加突出,弥补了抽样时造成的误差,是一种简洁有效的计算方法。
2个茶园竞争力最强和最弱的蜘蛛相同。本研究的2个茶园均按常规措施管理,不使用化学农药,且于冬季除草修剪,修剪会影响天敌的虫口基数,而‘农抗早’和‘平阳特早’抗逆性和抗寒性强[32],因此在相近的受害程度后,‘农抗早’和‘平阳特早’恢复时间均较短,恢复效果均较好,害虫所处环境变化的速度一致性可能是研究结果相同的原因之一。此外,茶园竞争力最强的是斑管巢蛛,这可能与斑管巢蛛的生活习性有关。斑管巢蛛定居且游猎于树冠上被害卷叶或枯叶等阴暗干燥处,白天基本不出行,黄昏时刻,蜘蛛开始活动,主动巡游猎取食,沿着枝、叶逐一搜索前进,几乎无遗漏之处。合理保护和利用斑管巢蛛这类竞争力强的蜘蛛可达到有效防治小贯小绿叶蝉的目的。
对茶园游猎蛛之间竞争作用的研究可以更好地理解游猎蛛之间的竞争如何影响害虫的数量和变化趋势以及如何影响生物防治的有效性[33]。至今利用害虫与天敌的种间关系对茶园害虫进行生物防治更多地还是停留在增加优势种天敌数量的方向上,分析天敌之间的竞争作用有利于选出最高效的天敌组合,在不破坏茶园原有生态环境的基础上高效防治害虫。
本研究中的7种游猎蛛都是广食性天敌,但为了研究方便,把它们作为只取食小贯小绿叶蝉一种食饵的单食性天敌,若进行深入研究就需要考虑多种猎物(害虫)共存时天敌对食物的嗜食性,在此基础上应用竞争关系分析方法就能更加真实地反映天敌之间的关系。另外,本研究是分析天敌两两之间的竞争关系,实际上,在食饵不足时,7种天敌之间也存在竞争关系,这种情况有待进一步研究。
Interspecific competition of wandering spiders feeding on Empoasca onukii in tea plantations based on Fuzzy analysis
-
摘要:
目的 探究茶园游猎蛛取食小贯小绿叶蝉Empoasca onukii的种间竞争作用,为合理保护和利用茶园游猎蛛防治小贯小绿叶蝉提供科学依据。 方法 运用灰色关联度法、Fuzzy分级法和竞争系数法对安徽农业大学科技示范园茶树Camellia sinensis‘农抗早’‘Nongkangzao’和‘平阳特早’‘Pingyangtezao’茶园小贯小绿叶蝉与游猎蛛的种群动态进行分析,研究不同游猎蛛之间的竞争关系,并得出竞争力最强的游猎蛛,再将灰色关联度与竞争系数相结合引申出竞争强度指数的概念以验证所得结果准确性。 结果 由灰色关联度结果可知:与小贯小绿叶蝉数量相关性最大的游猎蛛,‘农抗早’茶园为斑管巢蛛Clubiona reichlini(0.896 7)和粽管巢蛛Clubiona japonicola (0.890 2),‘平阳特早’茶园也为斑管巢蛛(0.860 3)和粽管巢蛛(0.857 1);根据竞争系数和竞争强度指数的方差分析结果可知:在取食茶园小贯小绿叶蝉时,无论竞争对手为何种游猎蛛,粽管巢蛛都与其他蜘蛛差异显著,其次为斑管巢蛛,且游猎蛛里斜纹猫蛛Oxyopes sertatus与两者差异极显著。 结论 取食茶园小贯小绿叶蝉时,粽管巢蛛和斑管巢蛛竞争力最强,斜纹猫蛛竞争力最弱。通过合理保护和利用粽管巢蛛和斑管巢蛛这类竞争力强的蜘蛛可达到有效防治小贯小绿叶蝉的目的。图2表6参33 Abstract:Objective The objective is to explore the interspecific competition of wandering spiders feeding on Empoasca onukii in tea plantations, so as to provide scientific basis for the rational conservation and control of wandering spiders against E. onukii. Method The population dynamics of E. onukii and wander spiders in ‘Nongkangzao’ and ‘Pingyangtezao’ tea plantations in Science and Technology Demonstration Garden of Anhui Agricultural University in 2021 were analyzed using grey correlation method, Fuzzy classification method and competition coefficient method. The competitive relationship between different wandering spiders was studied and the most competitive wandering spiders were identified. By combining grey correlation degree with competition coefficient, the concept of competition intensity index was derived to verify the accuracy of the obtained results. Result According to the grey correlation results, it could be seen that the wandering spiders most correlated with the number of E. onukii were Clubiona reichlini (0.896 7) and Clubiona japonicola (0.890 2) in ‘Nongkangzao’ tea plantation, and ‘Pingyangtezao’ tea plantation was also dominated by C. reichlini(0.860 3)and C. japonicola (0.857 1). Based on the variance analysis results of competition coefficient and competition intensity index, it was found that C. japonicola differed significantly from other spiders, followed by C. reichlini. Oxyopes sertatus differed very significantly from the two in feeding on E. onukii, regardless of the type of wandering spider in the competition. Conclusion When feeding on E. onukii in tea plantations, C. japonicola and C. reichlini are the most competitive, while O. sertatus is the least competitive. Effective control of E. onukii can be achieved by reasonably protecting and utilizing highly competitive spiders such as C. japonicola and C. reichlini. [Ch. 2 fig. 6 tab. 33 ref ] -
Key words:
- Empoasca onukii /
- wandering spiders /
- interspecific competition /
- Fuzzy analysis /
- biological control
-
图 1 ‘农抗早’茶园游猎蛛种群数量等分分级频数曲线(A)和Fuzzy频数曲线(B)
Figure 1 Equally graded frequency curve (A) and Fuzzy frequency curve (B) of the number of wandering spiders in ‘Nongkangzao’ tea plantation
图 2 ‘平阳特早’茶园游猎蛛种群数量等分分级频数曲线(A)和Fuzzy频数曲线(B)
Figure 2 Equally graded frequency curve (A) and Fuzzy frequency curve (B) of the number of wandering spiders in ‘Pingyangtezao’ tea plantation
表 1 2个茶园小贯小绿叶蝉与游猎蛛数量动态
Table 1. Population dynamics of E. onukii and wandering spiders in two tea plantations
茶树品种 日期
(月-日)害虫数量/头 游猎蛛数量/头 X1 X2 X3 X4 X5 X6 X7 ‘农抗早’ 03-25 3 10 16 3 2 32 7 7 04-16 11 2 7 1 0 7 0 19 05-07 42 2 6 0 0 2 2 0 05-23 30 1 5 0 0 5 12 1 06-04 222 7 1 5 0 1 5 0 06-20 175 20 5 5 5 0 5 0 07-08 594 14 65 51 88 6 38 0 08-10 209 27 23 15 60 20 26 24 08-22 429 23 19 29 59 33 25 6 09-06 92 19 9 19 23 57 18 6 09-17 30 17 17 13 10 31 13 19 10-13 1 8 1 2 0 45 4 5 11-01 52 4 2 9 9 39 4 19 11-19 47 1 1 3 6 15 3 5 合计 1 937 310 250 155 177 253 155 262 ‘平阳特早’ 03-25 9 2 3 0 0 13 17 4 04-16 11 0 5 0 0 1 6 2 05-07 31 1 6 0 0 0 6 0 05-23 57 3 4 1 0 0 5 0 06-04 251 1 5 0 3 0 11 0 06-20 139 15 8 18 2 0 10 1 07-08 674 26 46 24 34 22 7 0 08-10 318 17 3 17 42 33 13 0 08-22 534 8 18 21 48 31 10 29 09-06 160 29 12 14 18 39 16 10 09-17 20 13 12 12 19 37 12 9 10-13 4 7 2 0 0 20 11 2 11-01 23 5 2 8 3 2 2 3 11-19 108 1 2 3 5 26 2 4 合计 2 339 245 230 128 128 170 118 174 说明:X1~X7分别指鞍型花蟹蛛、三突花蟹蛛、粽管巢蛛、斑管巢蛛、斜纹猫蛛、黑色跳蛛和条纹蝇虎数量(头)。  下载: 导出CSV
下载: 导出CSV
表 2 2个茶园小贯小绿叶蝉与游猎蛛间的灰色关联度
Table 2. Grey correlation between E. onukii and wandering spiders in two tea plantations
游猎蛛 ‘农抗早’ ‘平阳特早’ 游猎蛛 ‘农抗早’ ‘平阳特早’ 灰色关联度 排位 灰色关联度 排位 灰色关联度 排位 灰色关联度 排位 鞍型花蟹蛛 0.833 8 5 0.826 1 4 斜纹猫蛛 0.754 8 6 0.794 7 6 三突花蟹蛛 0.856 9 4 0.856 2 3 黑色跳蛛 0.857 0 3 0.802 6 5 粽管巢蛛 0.890 2 2 0.857 1 2 条纹蝇虎 0.744 2 7 0.763 4 7 斑管巢蛛 0.896 7 1 0.860 3 1
下载: 导出CSV
表 3 2个茶园各游猎蛛之间的竞争系数
Table 3. Competition coefficients among wandering spiders in two tea plantations
茶树品种 竞争对手 游猎蛛间竞争系数 鞍型花蟹蛛 三突花蟹蛛 粽管巢蛛 斑管巢蛛 斜纹猫蛛 黑色跳蛛 条纹蝇虎 ‘农抗早’ 鞍型花蟹蛛 1.000 0 0.991 5 0.983 7 0.939 8 0.906 3 0.959 2 0.991 0 三突花蟹蛛 0.991 5 1.000 0 0.995 2 0.973 3 0.893 8 0.928 2 0.995 3 粽管巢蛛 0.983 7 0.995 2 1.000 0 0.980 3 0.901 5 0.916 7 0.993 6 斑管巢蛛 0.939 8 0.973 3 0.980 3 1.000 0 0.872 9 0.856 8 0.961 4 斜纹猫蛛 0.906 3 0.893 8 0.901 5 0.872 9 1.000 0 0.959 8 0.871 7 黑色跳蛛 0.959 2 0.928 2 0.916 7 0.856 8 0.959 8 1.000 0 0.915 9 条纹蝇虎 0.991 0 0.995 3 0.993 6 0.961 4 0.871 7 0.915 9 1.000 0 ‘平阳特早’ 鞍型花蟹蛛 1.000 0 0.989 3 0.998 5 0.976 5 0.913 7 0.993 5 0.975 1 三突花蟹蛛 0.989 3 1.000 0 0.984 2 0.977 4 0.869 7 0.984 4 0.992 5 粽管巢蛛 0.998 5 0.984 2 1.000 0 0.967 4 0.909 3 0.996 3 0.967 4 斑管巢蛛 0.976 5 0.977 4 0.967 4 1.000 0 0.939 7 0.954 0 0.957 1 斜纹猫蛛 0.913 7 0.869 7 0.909 3 0.939 7 1.000 0 0.880 6 0.826 5 黑色跳蛛 0.993 5 0.984 4 0.996 3 0.954 0 0.880 6 1.000 0 0.967 7 条纹蝇虎 0.975 1 0.992 5 0.967 4 0.957 1 0.826 5 0.967 7 1.000 0
下载: 导出CSV
表 4 2个茶园各游猎蛛之间竞争系数新复极差法分析结果
Table 4. Results of the new multiple range test analysis of competition coefficients among various wandering spiders in two tea plantations
竞争对手 F 物种 2个茶园竞争
系数均值5%显著
水平1%极显著
水平竞争对手 F 物种 2个茶园竞争
系数均值5%显著
水平1%极显著
水平鞍型花蟹蛛 7.530 粽管巢蛛 0.991 1 a A 斜纹猫蛛 1.203 黑色跳蛛 0.920 2 a A 三突花蟹蛛 0.990 4 a A 鞍型花蟹蛛 0.910 0 a A 条纹蝇虎 0.983 0 a A 斑管巢蛛 0.906 3 a A 黑色跳蛛 0.976 4 a A 粽管巢蛛 0.905 4 a A 斑管巢蛛 0.958 2 a AB 三突花蟹蛛 0.881 7 a A 斜纹猫蛛 0.910 0 b B 条纹蝇虎 0.849 1 a A 三突花蟹蛛 11.356 条纹蝇虎 0.993 9 a A 黑色跳蛛 0.562 鞍型花蟹蛛 0.976 4 a A 鞍型花蟹蛛 0.990 4 a A 粽管巢蛛 0.956 5 a A 粽管巢蛛 0.989 7 a A 三突花蟹蛛 0.956 3 a A 斑管巢蛛 0.975 4 a A 条纹蝇虎 0.941 8 a A 黑色跳蛛 0.956 3 a A 斜纹猫蛛 0.920 2 a A 斜纹猫蛛 0.881 7 b B 斑管巢蛛 0.905 4 a A 粽管巢蛛 3.305 鞍型花蟹蛛 0.991 1 a A 条纹蝇虎 12.037 三突花蟹蛛 0.993 9 a A 三突花蟹蛛 0.989 7 a A 鞍型花蟹蛛 0.983 0 a A 条纹蝇虎 0.980 5 a A 粽管巢蛛 0.980 5 a A 斑管巢蛛 0.973 9 a A 斑管巢蛛 0.959 2 a A 黑色跳蛛 0.956 5 ab A 黑色跳蛛 0.941 8 a A 斜纹猫蛛 0.905 4 b A 斜纹猫蛛 0.849 1 b B 斑管巢蛛 1.610 三突花蟹蛛 0.975 3 a A 粽管巢蛛 0.973 9 a A 条纹蝇虎 0.959 2 a A 鞍型花蟹蛛 0.958 2 a A 斜纹猫蛛 0.906 3 a A 黑色跳蛛 0.905 4 a A 说明:计算不同竞争对手下各游猎蛛在2个茶园竞争系数的平均值,5%水平上均数最大的标记为a,1%水平上均数最大的标记为A。向下比较,与之差异性不显著的标记相同字母,差异性显著的标记不同字母。
下载: 导出CSV
表 5 2个茶园各游猎蛛之间的竞争强度指数
Table 5. Competition intensity indices among wandering spiders in two tea plantations
茶树品种 竞争对手 游猎蛛间竞争强度指数 鞍型花蟹蛛 三突花蟹蛛 粽管巢蛛 斑管巢蛛 斜纹猫蛛 黑色跳蛛 条纹蝇虎 ‘农抗早’ 鞍型花蟹蛛 1.000 0 1.019 0 1.050 3 1.010 7 0.820 4 0.985 9 0.884 5 三突花蟹蛛 0.964 7 1.000 0 1.033 8 1.018 5 0.787 3 0.928 3 0.864 3 粽管巢蛛 0.921 4 0.958 0 1.000 0 0.987 5 0.764 4 0.882 5 0.830 6 斑管巢蛛 0.873 9 0.930 1 0.973 2 1.000 0 0.734 8 0.818 9 0.797 8 斜纹猫蛛 1.001 2 1.014 7 1.063 2 1.037 0 1.000 0 1.089 8 0.859 4 黑色跳蛛 0.933 2 0.928 1 0.952 2 0.896 5 0.845 3 1.000 0 0.795 3 条纹蝇虎 1.110 4 1.146 1 1.188 6 1.158 5 0.884 2 1.054 8 1.000 0 ‘平阳特早’ 鞍型花蟹蛛 1.000 0 1.025 4 1.036 1 1.017 0 0.879 0 0.965 3 0.9012 三突花蟹蛛 0.954 4 1.000 0 0.985 2 0.982 0 0.807 2 0.922 7 0.884 9 粽管巢蛛 0.962 3 0.983 2 1.000 0 0.971 0 0.843 1 0.932 9 0.861 7 斑管巢蛛 0.937 6 0.972 8 0.963 8 1.000 0 0.868 1 0.890 0 0.849 3 斜纹猫蛛 0.949 7 0.937 0 0.980 7 1.017 2 1.000 0 0.889 3 0.794 0 黑色跳蛛 1.022 6 1.050 2 1.064 0 1.022 6 0.872 0 1.000 0 0.920 5 条纹蝇虎 1.055 1 1.113 1 1.086 1 1.078 5 0.860 4 1.017 3 1.000 0
下载: 导出CSV
表 6 2个茶园各游猎蛛之间的竞争强度指数新复极差法分析结果
Table 6. Results of the new multiple range test analysis of competitive intensity indices among various wandering spiders in two tea plantations
竞争对手 F 物种 2个茶园竞争
强度指数均值5%显著
水平1%极显著
水平竞争对手 F 物种 2个茶园竞争
强度指数均值5%显著
水平1%极显著
水平鞍型花蟹蛛 32.992 粽管巢蛛 1.043 2 a A 斜纹猫蛛 2.142 斑管巢蛛 1.027 1 a A 三突花蟹蛛 1.022 2 ab A 粽管巢蛛 1.022 0 a A 斑管巢蛛 1.013 8 ab A 黑色跳蛛 0.989 5 ab A 黑色跳蛛 0.975 6 b A 三突花蟹蛛 0.975 8 ab A 条纹蝇虎 0.892 9 c B 鞍型花蟹蛛 0.975 4 ab A 斜纹猫蛛 0.849 7 c B 条纹蝇虎 0.826 7 b A 三突花蟹蛛 33.492 粽管巢蛛 1.009 5 a A 黑色跳蛛 1.575 粽管巢蛛 1.008 1 a A 斑管巢蛛 1.000 2 a AB 三突花蟹蛛 0.989 1 a A 鞍型花蟹蛛 0.960 9 ab AB 鞍型花蟹蛛 0.977 9 a A 黑色跳蛛 0.925 5 b BC 斑管巢蛛 0.959 6 a A 条纹蝇虎 0.874 6 c C 斜纹猫蛛 0.858 6 a A 斜纹猫蛛 0.797 3 d D 条纹蝇虎 0.857 9 a B 粽管巢蛛 9.706 斑管巢蛛 0.979 3 a A 条纹蝇虎 10.533 粽管巢蛛 1.137 3 a A 三突花蟹蛛 0.970 6 a AB 三突花蟹蛛 1.129 6 a A 鞍型花蟹蛛 0.941 9 a AB 斑管巢蛛 1.118 5 a A 黑色跳蛛 0.907 7 ab ABC 鞍型花蟹蛛 1.082 7 a A 条纹蝇虎 0.846 2 bc BC 黑色跳蛛 1.036 1 a A 斜纹猫蛛 0.803 8 c C 斜纹猫蛛 0.872 3 b B 斑管巢蛛 3.588 粽管巢蛛 0.968 5 a A 三突花蟹蛛 0.951 4 ab A 鞍型花蟹蛛 0.905 8 abc A 黑色跳蛛 0.854 4 abc A 条纹蝇虎 0.823 6 bc A 斜纹猫蛛 0.801 5 c A 说明:计算不同竞争对手下各游猎蛛在2个茶园竞争强度指数的平均值,5%水平上均数最大的标记为a,1%水平上均数最大的标记为A。向下比较,与之差异性不显著的标记相同字母,差异性显著的标记不同字母。
下载: 导出CSV
-
[1] ZHAO Qian, SHI Longqing, HE Weiyi, et al. Genomic variation in the tea leafhopper reveals the basis of adaptive evolution [J/OL]. Genomics, Proteomics & Bioinformatics, 2022[2022-11-06]. doi: 10.1016/j.gpb.2022.05.011. [2] YAO Qi, WANG Manqun, CHEN Zongmao. The relative preference of Empoasca onukii (Hemiptera: Cicadellidae) for oviposition on twenty-four tea cultivars [J]. Journal of Economic Entomology, 2022, 115(5): 1521 − 1530. [3] 郭明明, 李兆群, 刘岩, 等. 双丙环虫酯对小贯小绿叶蝉的防治效果及残留评价[J]. 茶叶科学, 2022, 42(3): 358 − 366. GUO Mingming, LI Zhaoqun, LIU Yan, et al. The control efficiency of afidopyropen to tea green leafhoppers and evaluation of residue in tea [J]. Journal of Tea Science, 2022, 42(3): 358 − 366. [4] HAZZI N A, HORMIGA G. Molecular phylogeny of the tropical wandering spiders (Araneae, Ctenidae) and the evolution of eye conformation in the RTA clade [J]. Cladistics, 2023, 39(1): 18 − 42. [5] MONSIMET J, COLINET H, DEVINEAU O, et al. Biogeographic position and body size jointly set lower thermal limits of wandering spiders [J]. Ecology and Evolution, 2021, 11(7): 3347 − 3356. [6] LAPINSKI W, TSCHAPKA M. Vertical distribution of wandering spiders in central America [J]. The Journal of Arachnology, 2018, 46(1): 13 − 20. [7] CHEN Shiyan, CHENG Honghao, WU Xiaomeng, et al. The dominant species of natural enemies of Pentatomidae in the peak period [J]. Entomological Research, 2023, 53(3): 95 − 105. [8] 孙佳照, 吴筱萌, 徐悦, 等. 小贯小绿叶蝉与其主要天敌空间关系研究方法的比较[J]. 植物保护, 2021, 47(5): 190 − 197. SUN Jiazhao, WU Xiaomeng, XU Yue, et al. A comparison of methods for investigating the spatial relationships between Empoasca onukii Matsuda and its main natural enemies [J]. Plant Protection, 2021, 47(5): 190 − 197. [9] 吴筱萌, 徐悦, 张淋, 等. 4种茶园花蓟马与其天敌的空间关系及差异[J]. 应用昆虫学报, 2022, 59(4): 805 − 814. WU Xiaomeng, XU Yue, ZHANG Lin, et al. Spatial relationships between Frankliniella intonsa and its natural enemies in four tea gardens [J]. Chinese Bulletin of Entomology, 2022, 59(4): 805 − 814. [10] CHENG Honghao, CHEN Shiyan, WU Xiaomeng, et al. Analysis of the following effect of the natural enemies with Frankliniella intonsa in tea garden [J]. Entomological Research, 2022, 52(8): 356 − 366. [11] ZHANG Lin, SUN Jiazhao, WU Xiaoment, et al. Comprehensive evaluation of natural enemy dominant species of Breuipalpus oboyats in tea garden in autumn and winter [J]. Entomological Research, 2021, 51(12): 650 − 657. [12] CHENG Xian, ZHANG Lin, WU Xiaomeng, et al. Differences in the closeness of spatial relationship between Ricanidae in their prime and natural enemies in five kinds of tea gardens [J]. Entomological Research, 2022, 52(8): 367 − 375. [13] 徐悦, 吴筱萌, 陈诗燕, 等. 茶园蜘蛛与广翅蜡蝉的时间和空间关系[J]. 浙江农林大学学报, 2022, 39(5): 1067 − 1079. XUE Yue, WU Xiaomeng, CHEN Shiyan, et al. Temporal and spatial relationship between spiders and Ricanidae in tea gardens [J]. Journal of Zhejiang A&F University, 2022, 39(5): 1067 − 1079. [14] BI Shoudong, QIAN Guangjing, SONG Xueyu, et al. Studies on dominant natural enemies of Ceroplastes rubens Maskell in tea gardens at different altitudes [J]. International Journal of Tropical Insect Science, 2022, 42(4): 2845 − 2852. [15] 李兆群. 我国茶园有害生物生物防治技术研究及应用[J]. 中国茶叶, 2022, 44(5): 8 − 12. LI Zhaoqun. Research and application of tea pest control technology in China [J]. China Tea, 2022, 44(5): 8 − 12. [16] 黄焯钦. 生物防治技术在茶园病虫害绿色防控中的应用研究[J]. 种子科技, 2022, 40(5): 103 − 105. HUANG Zhuoqin. Study on the application of biological control technology in the green control of diseases and pests in tea garden [J]. Seed Scicence &Technology, 2022, 40(5): 103 − 105. [17] SENTIS A, HEMPTINNE J L, MAGRO A, et al. Biological control needs evolutionary perspectives of ecological interactions [J]. Evolutionary Applications, 2022, 15(10): 1537 − 1554. [18] 徐学农, 吕佳乐, 王恩东. 国际捕食螨研发与应用的热点问题及启示[J]. 中国生物防治学报, 2013, 29(2): 163 − 174. XU Xuenong, LÜ Jiale, WANG Endong. Hot spots in international predatory mite studies and lessons to us [J]. Chinese Journal of Biological Control, 2013, 29(2): 163 − 174. [19] FAUST K, RAES J. Microbial interactions: from networks to models [J]. Nature Reviews Microbiology, 2012, 10(8): 538 − 550. [20] ROSALIA S, YONARIZA Y, SYAHRAWATI M. Effect of farmer’s behavior in cocoa management on insect diversity in Salayo Cocoa Plantation, West Sumatra, Indonesia [J]. Biodiversitas Journal of Biological Diversity, 2022, 23(10): 5064 − 5073. [21] ZOU Yan, SHEN Fangyuan, ZHONG Yanni, et al. Impacts of intercropped maize ecological shading on tea foliar and functional components, insect pest diversity and soil microbes [J/OL]. Plants, 2022, 11(14): 1883[2022-11-06]. doi: 10.3390/plants11141883. [22] 张书平, 钱广晶, 毕守东, 等. 基于Fuzzy分析的假眼小绿叶蝉与天敌蜘蛛的数量关系[J]. 四川农业大学学报, 2019, 37(5): 653 − 662. ZHANG Shuping, QIAN Guangjing, BI Shoudong, et al. Quantitative relationship between Empoasca vitis and spiders of natural enemies based on Fuzzy analysis [J]. Journal of Sichuan Agricultural University, 2019, 37(5): 653 − 662. [23] 汪清, 潘萍, 欧阳勋志, 等. 马尾松-木荷不同比例混交林种内和种间竞争强度[J]. 生态学杂志, 2021, 40(1): 49 − 57. WANG Qing, PAN Ping, OUYANG Xunzhi, et al. Intraspecific and interspecific competition intensity in mixed plantation with different proportion of Pinus massoniana and Schima superba [J]. Chinese Journal of Ecology, 2021, 40(1): 49 − 57. [24] 邓聚龙. 灰色系统理论教程[M]. 武汉: 华中理工大学出版社, 1990: 33 − 84. DENG Julong. Grey System Theory Tutorial [M]. Wuhan: Huazhong University of Science and Technology Press, 1990: 33 − 84. [25] 贺仲雄. 模糊数学及其应用[M]. 天津: 天津科学技术出版社, 1984. HE Zhongxiong. Fuzzy Mathematics and Its Application [M]. Tianjin: Tianjin Science and Technology Press, 1984. [26] 邹运鼎. 模糊分级法在害虫分析中的应用[J]. 昆虫知识, 1986, 23(3): 138 − 140. ZOU Yunding. Application of fuzzy classification method in pest analysis [J]. Chinese Bulletin of Entomology, 1986, 23(3): 138 − 140. [27] 宋丁全, 姜志林, 郑作孟, 等. 光皮桦林优势树种间的竞争[J]. 南京林业大学学报, 2000, 24(4): 26 − 28. SONG Dingquan, JIANG Zhilin, ZHENG Zuomeng, et al. Competition among dominant species in forest of Betula luminifera [J]. Journal of Nanjing Forestry University, 2000, 24(4): 26 − 28. [28] PIANKA E R. The structure of lizard communities [J]. Annual Review of Ecology and Systematics, 1973, 4(1): 53 − 74. [29] MAY R M. Stability and Complexity in Model Ecosystems [M]. Princeton: Princeton University Press, 2019. [30] MAY R M. Some notes on estimating the competition matrix, α [J]. Ecology, 1975, 56(3): 737 − 741. [31] 李永宏, 黄清臻. 新复极差法在生物统计中的应用[J]. 医学动物防制, 2002, 18(5): 270 − 272. LI Yonghong, HUANG Qingzhen. Application of new complex range method in biostatistics [J]. Journal of Medical Pest Control, 2002, 18(5): 270 − 272. [32] 张翠玲, 侯君合, 王芝云, 等. 青岛地区茶树无性系品种抗寒性比较试验[J]. 山东林业科技, 2008, 38(2): 30 − 31. ZHANG Cuiling, HOU Junhe, WANG Zhiyun, et al. The comparison of the cold-resistance of the tea clone varieties in Qingdao district [J]. Journal of Shandong Forestry Science and Technology, 2008, 38(2): 30 − 31. [33] CUSUMANO A, PERI E, BRADLEIGH VINSON S, et al. Interspecific extrinsic and intrinsic competitive interactions in egg parasitoids [J]. BioControl, 2012, 57(6): 719 − 734. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220753
 点击查看大图
点击查看大图
计量
- 文章访问数: 1399
- HTML全文浏览量: 430
- PDF下载量: 16
- 被引次数: 0
