




-
竹类植物因其形态多样性和生态适应性在景观营造中具有独特价值,是园林植物中的重要观赏类群。花叶矢竹Pseudosasa japonica f. akebonosuji是原产于日本的珍稀观赏竹矢竹P. japonica的一个变型,为混生竹种。叶片分为白叶、绿叶以及白绿色条纹叶,具有较高的观赏价值,其叶色存在不规则的变异,部分花叶叶片白色部位会逐渐恢复绿色(复绿现象)。
关于花叶竹类的叶色变异规律,目前已开展了一些研究[1−2]。陈凌艳等[3]对银丝竹Bambusa multiplex‘Silverstripe’研究发现:造成叶片白化现象的原因是白叶叶片在叶绿素生物合成过程中明显受阻,导致叶绿体类囊体的形成受到抑制,进而影响了叶绿体的正常发育。徐薪璐等[4]对靓竹Sasaella glabra‘Albostriata’不同发育时期的叶片研究发现:黄区相比于绿区,存在指状臂细胞层数少、叶绿体发育异常以及叶绿素合成受阻的现象。同时,植物叶色突变机制受外部环境影响,温度是其中重要的影响因子[5]。目前,温度对植物叶色变化的研究集中在水稻Oryza sativa、小麦Triticum aestivum、辣椒Capsicum annuum、黄瓜Cucumis sativus以及羽衣甘蓝Brassica oleracea var. acephala等植物中[6−10]。高温胁迫下,水稻在幼苗期会缺失TCM5基因,影响叶绿体发育,导致叶色白化[11]。LIU等[12]对高温处理下中华金叶榆Ulmus pumila‘jinye’的研究发现:金叶榆叶片在高温胁迫下光合色素含量降低、叶绿体结构遭到破坏以及丧失了光合能力,其叶片颜色从金黄色转变为白色。这些研究都证明植物叶色受到温度的影响,但温度对竹类植物叶色变化的影响尚缺乏报道。因此,本研究以花叶矢竹白叶为研究对象,以自然温度下新生白叶为对照,通过3个恒定温度(15、25、35 ℃)处理,研究其叶片光合色素水平、叶绿素合成前体物质相对含量、光系统(PSⅠ、PSⅡ)特性以及叶绿体超微结构,探讨温度对花叶矢竹白叶发育生理特性的影响,为进一步开展竹子叶色变异分子机制研究提供参考。
-
试验地点为浙江省杭州市临安区浙江农林大学校园内,试验材料为浙江农林大学智能温室的花叶矢竹花叶株的盆苗(育苗盆口径为25 cm),选取有新展开白叶的花叶株(生长健壮、高度相差不大),标记同一时期、同一位置展开的白叶,分别放置于15、25和35 ℃的人工培养箱(白天16 h,黑夜8 h,光照为180 μmol·m−2·s−1,空气相对湿度为70%),以自然温度下生长的花叶株白叶为对照组(ck),每处理6盆,共24盆。培养30 d后分别取样,每处理取18片叶子。试验时间为2024年4月(日最高和最低温度分别为21和12 ℃,平均温度为16.5 ℃)、12月(日最高和最低温度分别为18和−3 ℃,平均温度为11 ℃),2个时间段均为花叶株白叶伸展的时期,且没有出现复绿现象。后续测定结果均取2次试验的平均值。
-
花叶矢竹白叶光合色素测定参考李合生[13]的方法:称取0.5 g叶片,去中脉并剪碎后,置于带盖试管中,加入5.00 mL叶绿素提取液[V(丙酮)∶V(无水乙醇)∶V(纯水)=4.5∶4.5∶1.0],室温下遮光萃取12 h,以提取液为空白对照,在波长663、645和470 nm处测定吸光度,并计算叶绿素a (Chl a),叶绿素b (Chl b)及类胡萝卜素的质量分数,重复3次。
-
δ-氨基乙酰丙酸(ALA)测定参照DEI[14]的方法:称取0.5 g叶片,在液氮中研磨成粉末,加入质量分数为4%的三氯乙酸溶液,将总体积调整至20 mL。接着,将混合液在12 000 r·min−1条件下离心15 min后,取5.00 mL上清液,加入0.15 mL乙酰丙酮和2.35 mL醋酸钠溶液。将混合物置于沸水浴中加热10 min,之后冷却至室温。取2.00 mL处理后的液体,加入2.00 mL Ehrlich-Hg试剂,在避光条件下显色15 min。最后,在553 nm波长处测量吸光度。
胆色素原(PBG)测定参照BOGORAD[15]的方法:称取0.5 g叶片,在液氮中充分研磨后,加入5.00 mL提取缓冲液(含0.6 mol·L−1的Tris,0.1 mol·L−1的EDTA,pH为5.8)。将混合液在12 000 r·min−1条件下离心15 min后,取2.00 mL上清液,加入2.00 mL Ehrlich-Hg试剂,在避光环境中显色15 min。最后,在553 nm波长处测定吸光度。
尿卟啉原Ⅲ (Urogen Ⅲ)和粪卟啉原Ⅲ (Coprogen Ⅲ)测定参照BOGORAD[15]的方法:称取1.0 g叶片,在液氮中研磨后,加入10.00 mL提取缓冲液(0.067 mol·L−1,pH为6.8的磷酸缓冲液),混合液离心后,取5.00 mL上清液,加入0.25 mL质量分数为1%的亚硫酸钠溶液,剧烈震荡后用强光照射20 min。随后,用冰醋酸将溶液pH调至3.5,并用10.00 mL的乙醚萃取3次,测定水相在405.5 nm处的吸光度。最后,将上述乙醚萃取液用0.1 mol·L−1的盐酸萃取3次,测定盐酸相在399.5 nm处的吸光度。
原卟啉Ⅸ (Proto Ⅸ)、镁原卟啉Ⅸ (Mg-Proto Ⅸ)和原脱植基叶绿素(Pchlide)测定参照杨海芸[16]的方法:称取1.0 g叶片,液氮研磨后加入25.00 mL质量分数为80%的丙酮,离心后取上清液,分别在波长575、590和628 nm处测定吸光度。
-
取处理后的花叶矢竹白叶,避开叶脉,用刀片割成1 mm×3 mm×1 mm大小,在质量分数为2.5%的戊二醛中固定,抽真空至样品沉底。在4 ℃固定一晚后,倾倒该固定液,用磷酸缓冲液冲洗。接着,将样品用质量分数为1%的锇酸溶液固定后,用磷酸缓冲液清洗。在此基础上,采用不同浓度的乙醇进行脱水处理,再分别用质量分数为90%和95%的丙酮溶液处理,最后经过2次纯丙酮处理。将样品使用包埋剂与丙酮的混合液(体积比为1∶1)处理,再用其混合液(体积比为3∶1)处理;置于纯包埋剂中一夜,浸渍后在70 ℃过夜处理,经切片染色,用Hitachi H-7650型透射电镜观察[17]。
-
参照王妮等[18]的方法:花叶矢竹叶片经过暗适应15 min后,使用M-PEA多功能植物效率分析仪测量叶绿素荧光诱导动力学曲线,计算PSⅠ和PSⅡ中的各项指标。
-
用Excel处理数据,数据均为平均值±标准差;用SPSS 27进行方差分析,显著水平为0.05;利用Origin 2022作图。
-
由图1可知:花叶矢竹花叶在不同温度下培养30 d后,叶片呈现出不同的复绿程度。25 ℃处理下,花叶矢竹花叶株全盆白叶叶片复绿,新展开叶从叶脉中部开始出现复绿现象(图1 C),直至全株复绿;而15 ℃处理下全盆白叶叶片基部呈淡绿色,35 ℃处理下花叶株白叶维持白色。

图 1 不同温度处理前后花叶矢竹白叶的表型特征
Figure 1. Morphological characteristics of white leaves of P. japonica f. akebonosuji before and after different temperature treatments
-
由表1可知:不同温度下花叶矢竹白叶光合色素存在差异。15和25 ℃处理下白叶Chl a、Chl b和总叶绿素的质量分数均显著高于ck (P<0.05),而35 ℃处理低于ck。其中,25 ℃处理下Chl a、Chl b、总叶绿素的质量分数以及Chl a/b最高,分别是ck的3.8、2.6、3.4及1.5倍,是15 ℃处理的3.2、2.0、2.7及1.6倍;15和35 ℃处理的类胡萝卜素质量分数显著低于ck (P<0.05),而25 ℃处理仍高于ck,达到最大,是ck的1.7倍。总之,不同处理叶绿素类质量分数从大到小依次为:25 ℃、15 ℃、ck、35 ℃处理,而类胡萝卜素质量分数从大到小依次为:25 ℃、ck、15 ℃、35 ℃处理。这表明25 ℃处理促进花叶矢竹白叶复绿,而35 ℃处理抑制花叶矢竹白叶复绿,15 ℃处理则延缓复绿。
表 1 不同温度处理下花叶矢竹白叶的光合色素
Table 1. Photosynthetic pigment in white leaves of P. japonica f. akebonosuji under different temperatures
温度/℃ Chl a/(µg·g−1) Chl b/(µg·g−1) 总叶绿素/(µg·g−1) 类胡萝卜素/(µg·g−1) Chl a/b ck 62.41±0.16 c 39.66±0.29 c 102.07±0.25 c 57.35±0.07 b 1.57±0.01 b 15 75.60±0.14 b 51.44±0.49 b 127.05±0.63 b 29.81±0.10 c 1.47±0.01 b 25 239.10±0.61 a 103.30±0.67 a 342.48±1.29 a 95.67±0.13 a 2.31±0.01 a 35 31.48±1.24 d 33.96±2.05 d 65.44±1.71 d 24.04±0.44 d 0.92±0.07 c 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。 -
由表2可知:不同温度下花叶矢竹白叶叶绿素合成前体物质的质量分数存在显著差异(P<0.05)。与ck相比,虽然不同温度下δ-氨基乙酰丙酸、尿卟啉原Ⅲ、粪卟啉原Ⅲ的质量分数均有显著差异(P<0.05),但其倍数均在3倍以内。而25 ℃处理下,白叶原卟啉Ⅸ、镁原卟啉Ⅸ、原脱植基叶绿素的质量分数分别是ck的4.7、6.1、11.2倍。结果表明:花叶矢竹白叶25 ℃处理下出现复绿,可能与原卟啉Ⅸ、镁原卟啉Ⅸ、原脱植基叶绿素的增加有关。
表 2 不同温度处理下花叶矢竹白叶的叶绿素合成前体物质
Table 2. Chlorophyll synthesis precursors in white leaves of P. japonica f. akebonosuji under different temperatures
温度/℃ δ-氨基乙酰丙酸/
(µmol·g−1)胆色素原/
(µmol·g−1)尿卟啉原Ⅲ/
(mol·g−1)粪卟啉原Ⅲ/
(mol·g−1)原卟啉Ⅸ/
(µmol·g−1)镁原卟啉Ⅸ/
(µmol·g−1)原脱植基叶绿素/
(µmol·g−1)ck 1.36±0.02 b 2.11±0.05 c 28.58±0.09 d 63.39±0.33 b 44.20±0.47 d 18.47±0.23 d 7.77±0.32 d 15 1.18±0.04 d 4.67±0.03 a 31.63±0.22 c 62.03±0.45 c 53.21±0.87 c 23.65±0.14 c 11.31±0.33 c 25 1.27±0.09 c 3.69±0.02 b 32.24±0.24 b 59.99±0.00 d 205.84±0.43 a 113.41±0.12 a 86.70±0.12 a 35 1.46±0.04 a 3.69±0.01 b 35.28±0.16 a 66.81±0.17 a 96.15±1.01 b 44.36±0.18 b 23.83±0.31 b 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。 -
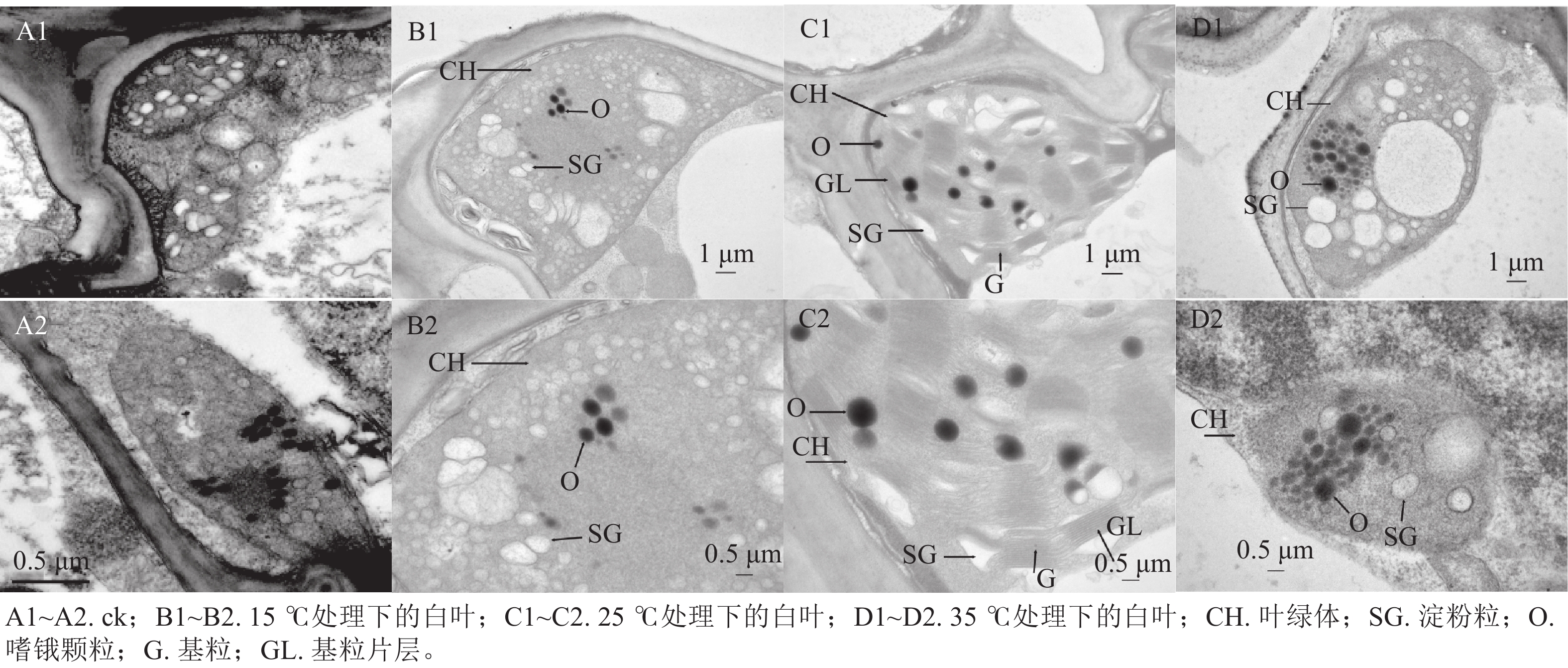
通过对花叶矢竹白叶的叶绿体结构进行观察发现,与ck(图2A1~A2)相比,15 ℃处理下的白叶叶绿体内,淀粉粒垛叠在一起但未形成类囊体(图2B1~B2);25 ℃处理下的白叶叶绿体结构中,类囊体片层结构垛叠紧实,基粒结构清晰,嗜锇颗粒零散分布在叶绿体内(图2C1~C2);35 ℃处理下的白叶叶绿体内,淀粉粒仍分散分布,且未见明显的垛叠(图2D1~D2)。由此可知,25 ℃处理下花叶矢竹白叶叶绿体形成类囊体;15 ℃处理延缓了花叶矢竹白叶叶绿体类囊体的垛叠,而35 ℃处理能抑制淀粉粒垛叠形成类囊体,15和35 ℃处理都造成了叶绿体发育异常,使花叶矢竹白叶没有出现复绿现象。
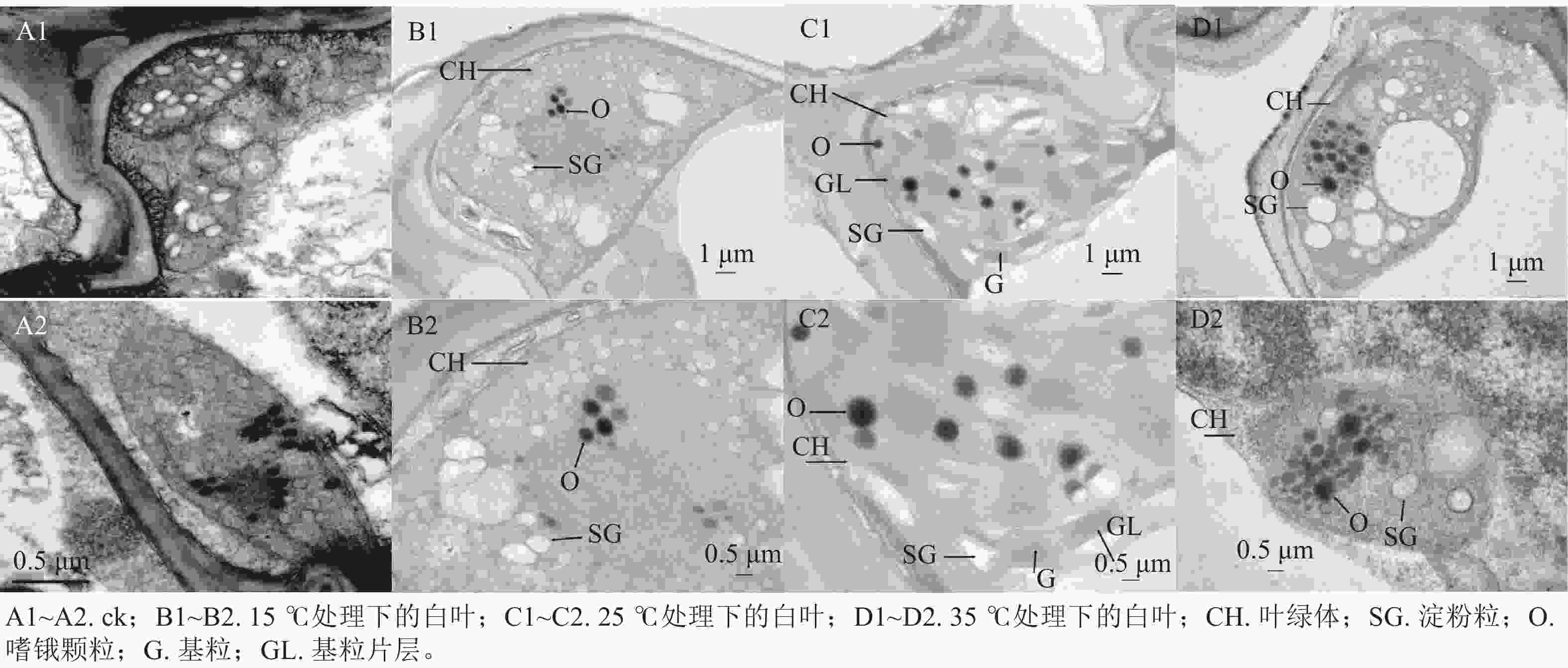
图 2 不同温度处理下花叶矢竹白叶的超微结构
Figure 2. Ultrastructure of white leaves of P. japonica f. akebonosuji under different temperature treatments
-
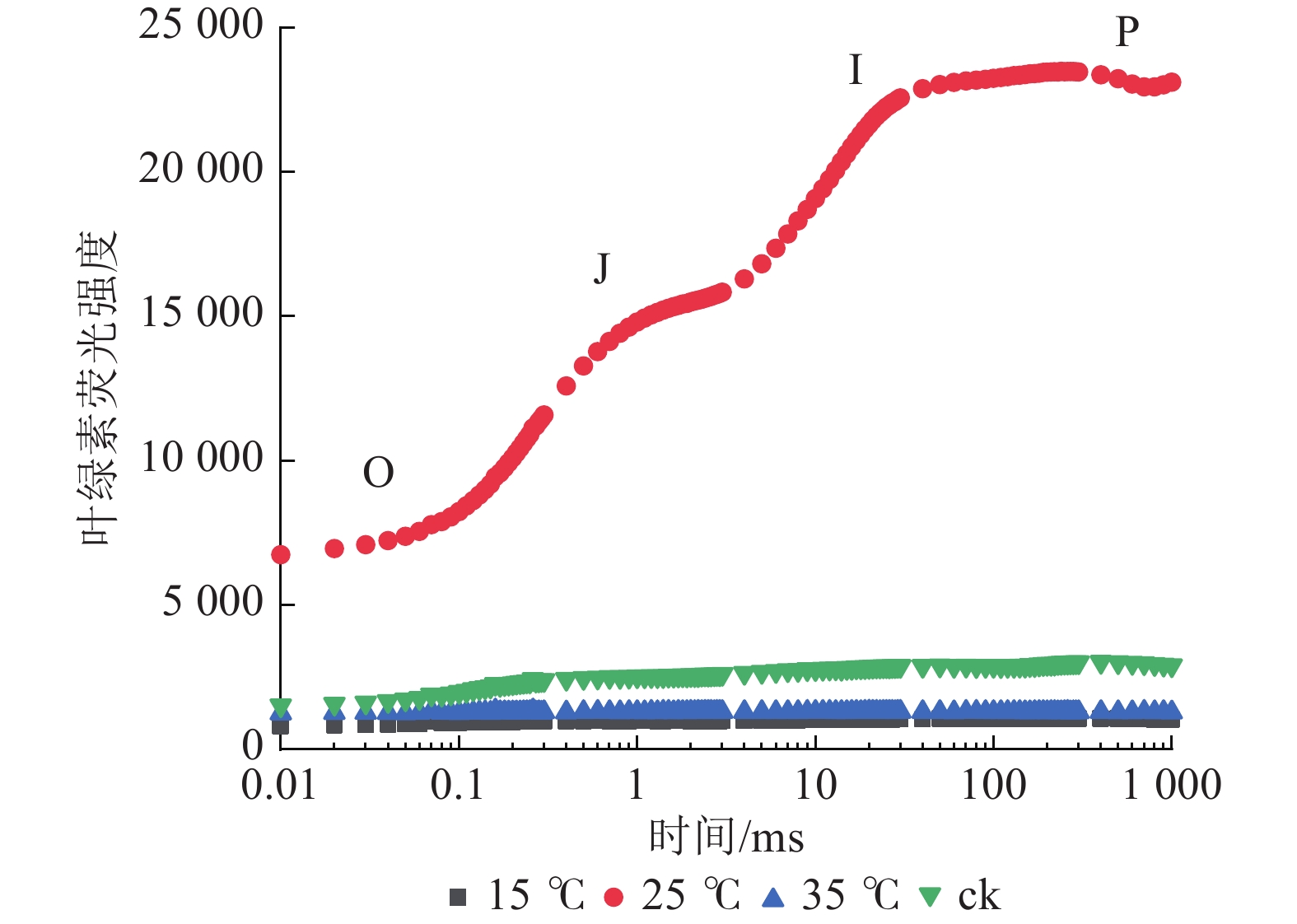
由图3可知:25 ℃处理下的花叶矢竹白叶与其他白叶的叶绿素荧光诱导曲线有明显差异。与ck相比,25 ℃处理下的白叶经过暗适应后,曝光的荧光强度高,且呈现O、J、I、P相,并伴随着明显的荧光上升和逐渐平缓的趋势;而15和35 ℃处理下的白叶经暗适应后,曝光的荧光强度未见明显上升。这表明花叶矢竹白叶经25 ℃处理后,其PSⅡ活性得以恢复;而15和35 ℃处理下,其PSⅡ活性受到抑制,与ck相近。
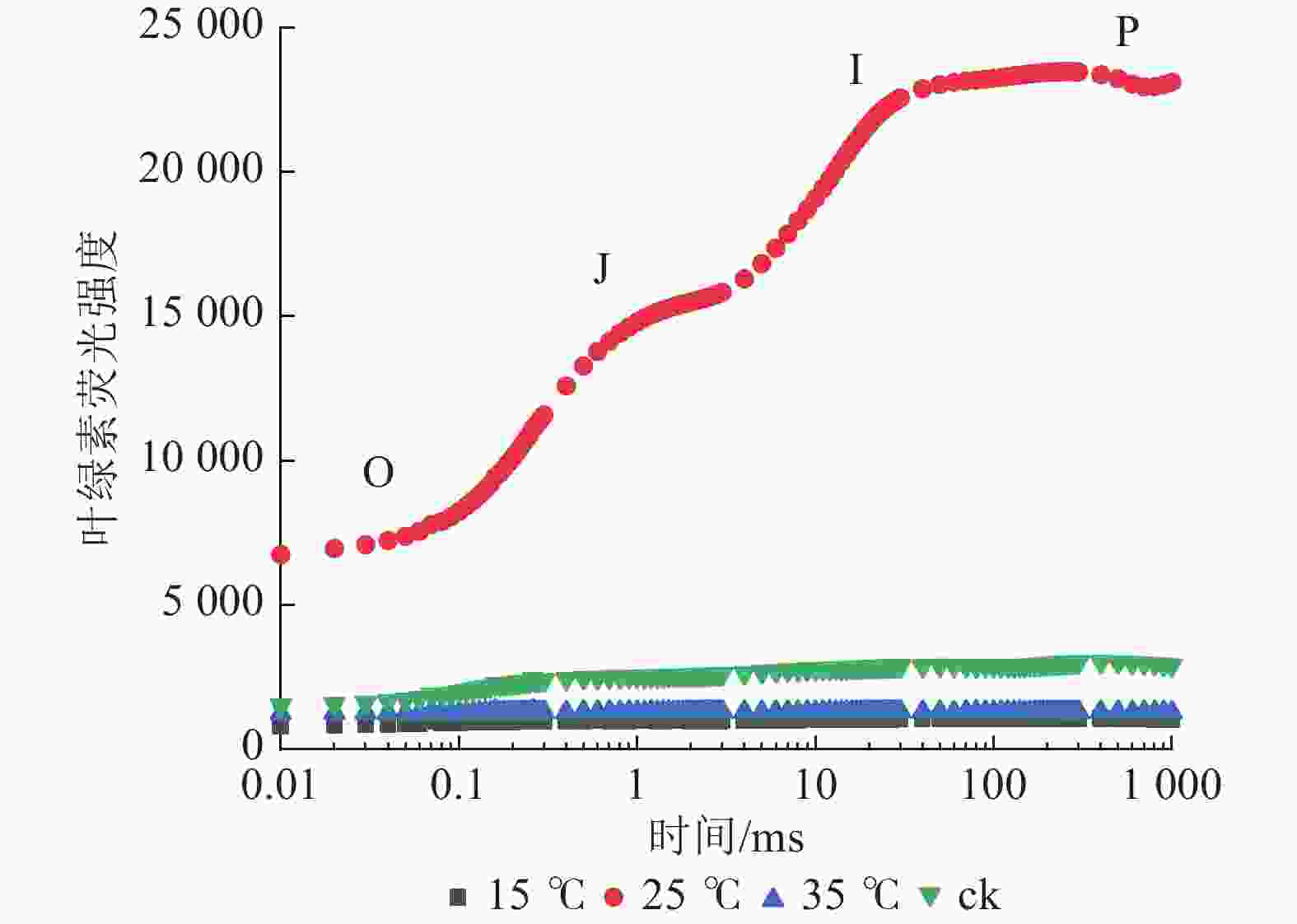
图 3 不同温度处理下花叶矢竹白叶之间的叶绿素荧光诱导动力学曲线
Figure 3. Dynamic curves for chlorophyll a fluorescein of white leaves of P. japonica f. akebonosuji under different temperature treatments
-
由表3可知:与ck相比,25 ℃处理下的花叶矢竹白叶反应中心密度(RC/CSO)、量子产额、能量分配比率以及性能指数中的各项指标都显著提高(P<0.05)。但在比活性参数中,单位反应中心吸收的光量(ABS/RC)、单位反应中心热耗散的能量(DIO/RC)均低于ck,分别是ck的7.3%和2.3%;15 ℃处理的ABS/RC、DIO/RC也低于ck,是ck的42.1%和38.1%;35 ℃处理与ck无显著差异。与ck相比,3个温度下被单位反应中心捕获的光量(TRO/RC)和单位反应中心用于电子传递的能量(ETO/RC)均无显著差异,25 ℃处理的RC/CSO增长了150倍,量子产额中最大PSⅡ的光能转换效率[φPO(Fv/Fm)]增长了10.4倍,PSⅡ吸收光能后进行电子转移的量子产额(φEO)增长了15倍,能量分配比率中反应中心向下传递电子的能力(ψO)提高了44.8%,从光子的吸收至电子传递链的还原性能指数(PIABS)从0提升至0.66。
表 3 不同温度处理下花叶矢竹白叶叶绿素荧光参数的变化
Table 3. Changes in chlorophyll fluorescence parameters of the white leaves of P. japonica f. akebonosuji under different temperatures
温度/℃ ABS/RC DIO/RC TRO/RC ETO/RC RC/CSO ck 41.23±2.19 a 37.42±3.15 a 3.41±0.96 a 2.32±0.26 a 12.5±3.91 b 15 17.35±5.49 b 14.27±5.49 bc 3.13±0.07 a 0.95±0.22 a 53.2±29.0 b 25 3.02±0.30 c 0.85±0.24 c 2.18±0.06 a 0.91±0.08 a 1876.9 ±438.6 a35 30.21±5.84 ab 28.13±6.54 ab 2.07±0.71 a 0.15±0.01 a 18.70±6.80 b 温度/℃ φPO(Fv/Fm) φEO ψO PIABS ck 0.07±0.04 b 0.02±0.01 b 0.29±0.11 a 0.00±0.00 b 15 0.17±0.09 b 0.05±0.02 b 0.27±0.06 a 0.00±0.00 b 25 0.73±0.05 a 0.30±0.02 a 0.42±0.03 a 0.66±0.18 a 35 0.08±0.02 b 0.01±0.00 b 0.04±0.00 b 0.00±0.00 b 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。 -
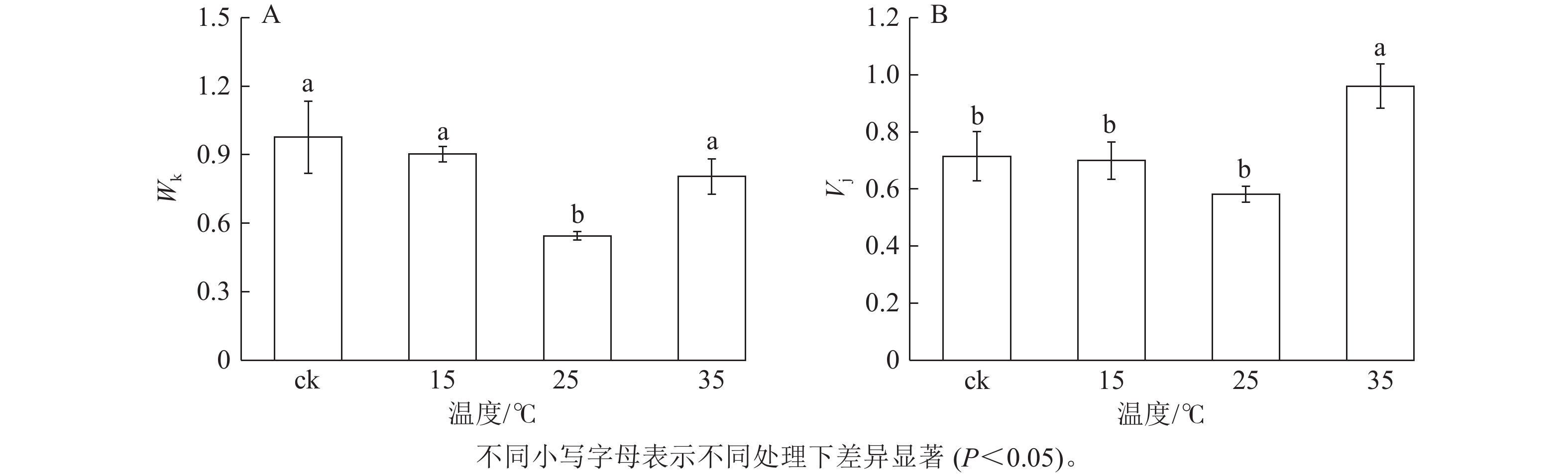
K点的荧光强度变化(Wk)反映PSⅡ供体侧的变化,J点的可变荧光(Vj)反映受体侧的变化。由图4A可知:与ck相比,25 ℃处理下Wk显著降低(P<0.05),下降了44.2%;15和35 ℃处理白叶与ck无显著差异。这表明25 ℃处理后的白叶PSⅡ供体侧放氧复合体之后的电子传递链得到恢复,而15和35 ℃处理的白叶PSⅡ供体侧仍受到抑制。由图4B可知:与ck相比,35 ℃处理下Vj显著提升(P<0.05),而25 ℃处理的白叶Vj虽无显著差异,但与ck相比仍下降了18.5%,表明35 ℃处理的白叶PSⅡ受体侧性能受到严重抑制,而25 ℃处理白叶PSⅡ受体侧性能开始恢复。
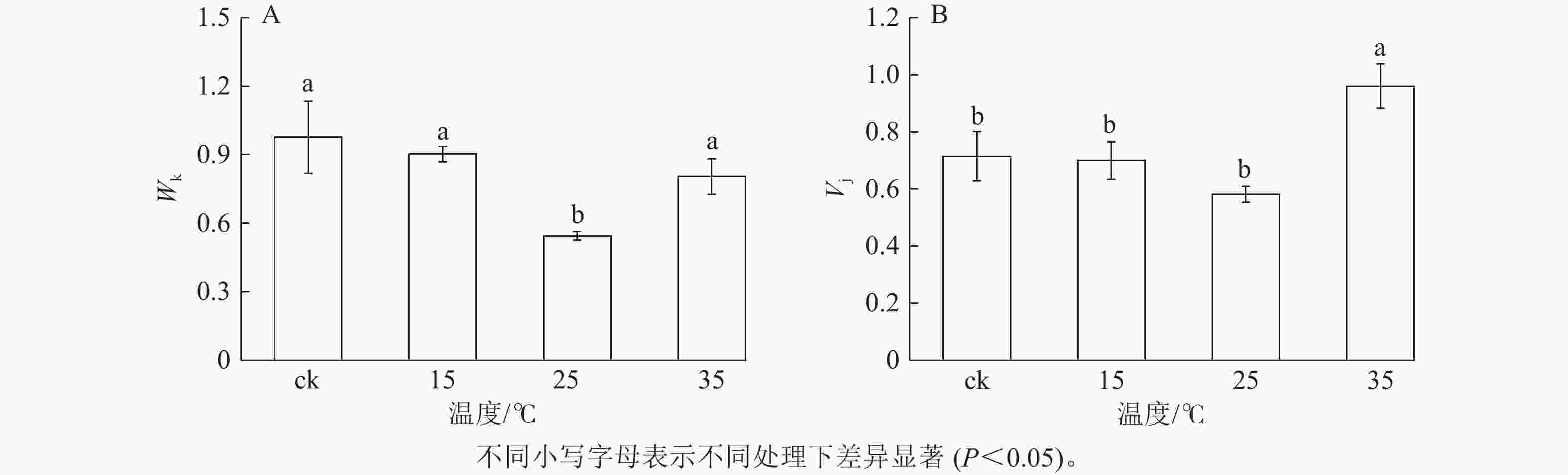
图 4 不同温度处理下花叶矢竹白叶K点的荧光强度变化(Wk)和J点的可变荧光(Vj)
Figure 4. Changes in fluorescence intensity at K point (Wk) and variable fluorescence at J point (Vj) of white leaves of P. japonica f. akebonosuji under different temperatures
-
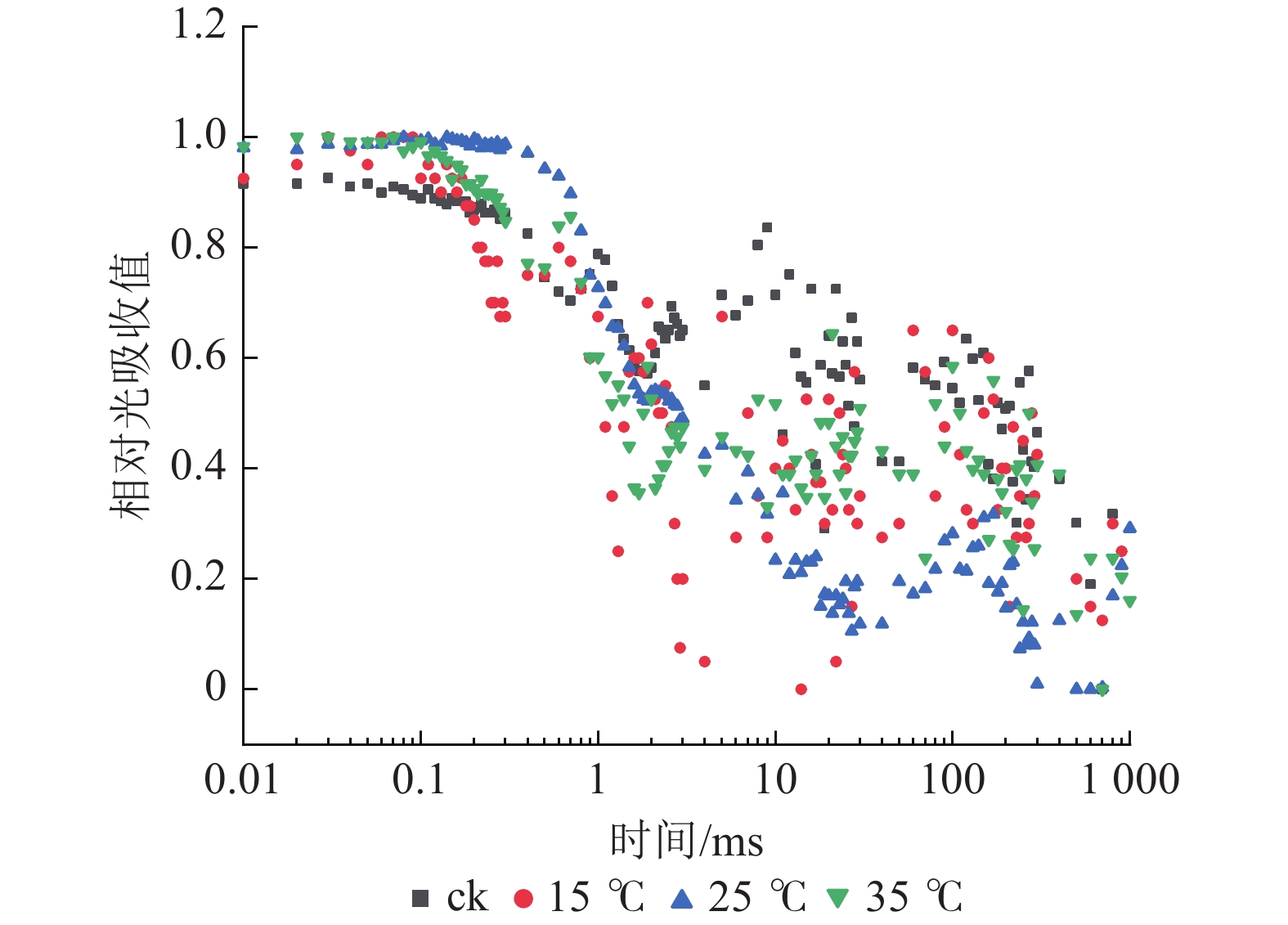
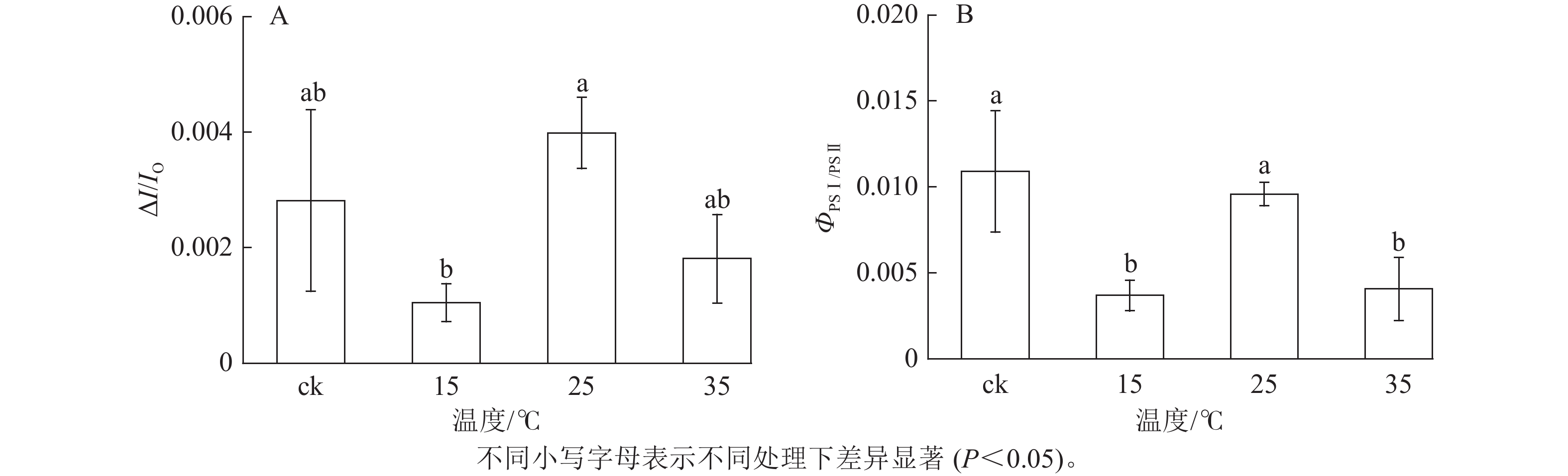
如图5所示:与ck相比,不同温度下白叶叶片PSⅠ对820 nm处的光吸收量差异较小。PSⅠ的最大氧化还原活性(ΔI/IO)是对其性能的综合评价。由图6A可知:与ck相比,15 ℃处理下ΔI/IO显著下降(P<0.05),降幅达62.8%,而25和35 ℃处理均无明显差异,表明15 ℃处理下白叶PSⅠ的最大氧化还原活性受到显著抑制(P<0.05)。
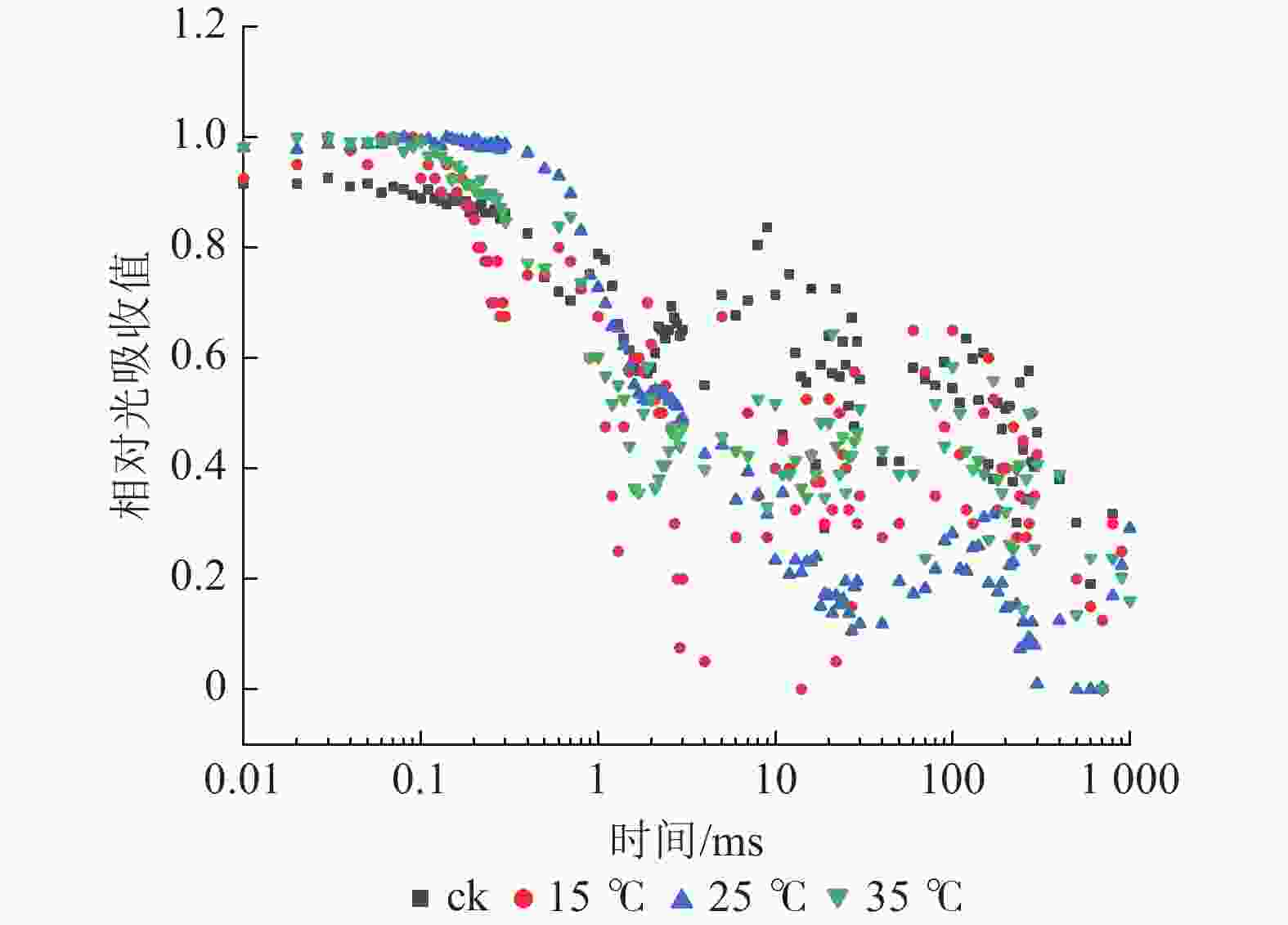
图 5 花叶矢竹白叶不同温度处理间的PSⅠ 820 nm相对光吸收值变化
Figure 5. Changes of relative absorbtion of PSⅠ 820 nm white leaves of P. japonica f. akebonosuji under different temperature treatments
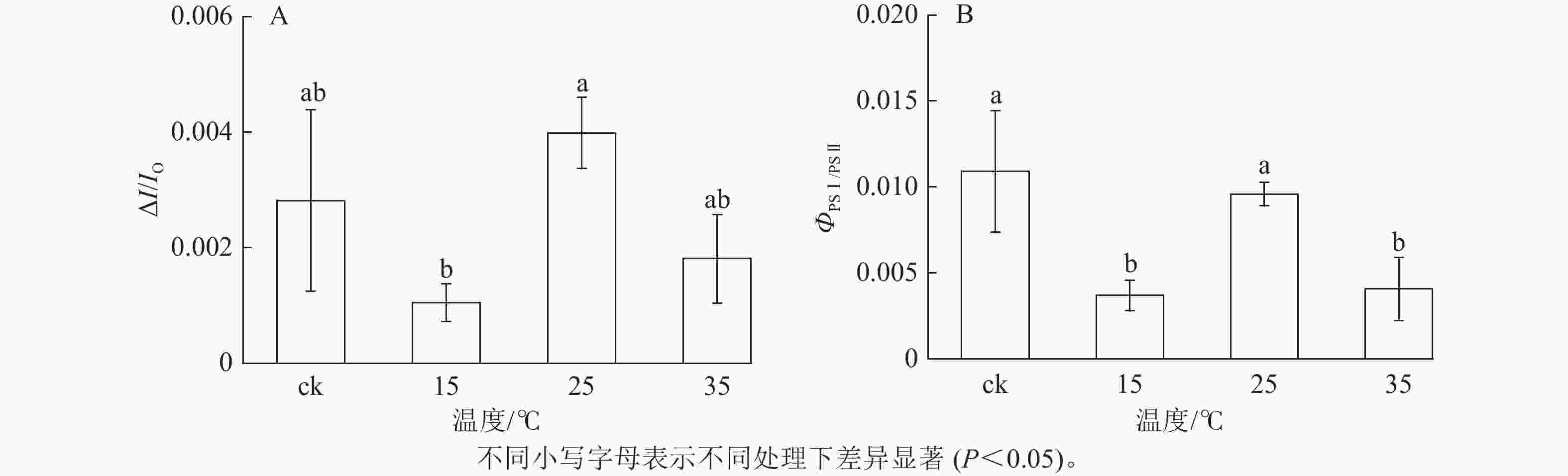
图 6 花叶矢竹白叶不同温度处理间的PSⅠ最大氧化还原能力(ΔI/IO)及PSⅠ与PSⅡ之间的协调性变化(ΦPSⅠ/PSⅡ)
Figure 6. PSⅠ maximum ability of oxido-reduction (ΔI/IO) and coordination between PSⅠ and PSⅡ (ΦPSⅠ/PSⅡ) of the white leaves of P. japonica f. akebonosuji under different temperature treatments
-
如图6B所示:与ck相比,25 ℃处理的白叶叶片PSⅠ与PSⅡ间协调性未发生显著变化,但15和35 ℃处理出现显著差异(P<0.05),分别下降了66.1%和62.7%。结果表明:15和35 ℃处理后的白叶PSⅡ与PSⅠ间协调性受到抑制。
-
高等植物叶色突变的主要原因是叶片中叶绿素含量的增加或减少[19],叶绿素含量变化受叶绿素合成前体物质的影响。本研究中,不同温度处理下花叶矢竹白叶叶绿素合成前体物质差异显著,变化位点位于粪卟啉原Ⅲ合成原卟啉Ⅸ;与ck相比,25 ℃处理后的白叶从原卟啉Ⅸ到原脱植基叶绿素的相对含量显著增加。毛晶晶等[20]在分析低温胁迫下玉米幼苗转绿过程中叶绿素生物合成中间产物时发现:低温通过影响δ-氨基乙酰丙酸到胆色素原和镁原卟啉Ⅸ向镁原卟啉甲酯的转化过程,抑制了玉米的转绿过程。在水稻突变体W1的研究中发现:低温下W1叶片叶绿素生物合成过程中δ-氨基乙酰丙酸和胆色素原大量积累,而原卟啉Ⅸ、镁原卟啉Ⅸ和原脱植基叶绿素含量减少,表明低温使胆色素原转化原卟啉Ⅸ受阻[21]。CHEN等[22]对不同叶片表型的花叶唐竹Sinobambusa tootsik‘Huayetangzhu’研究中发现:叶色突变受叶绿素合成受阻影响,受阻位点位于粪卟啉原Ⅲ到原卟啉Ⅸ的转化。虽然不同植物叶绿素合成受阻点不同,但都导致叶绿素缺失,引起叶色突变。因此,花叶矢竹白叶在不同温度处理30 d后,叶绿素合成过程中原卟啉Ⅸ到原脱植基叶绿素质量分数的变化是导致叶色变化的直接原因。
-
温度作为重要的环境因素,能够影响并调节植物的光合作用。而叶绿素荧光参数是衡量光能吸收、利用、耗散和分配的重要指标,可通过叶绿素荧光参数研究PSⅡ的功能变化,其中,利用植物对820 nm光的吸收,可以分析PSⅠ的功能变化[23]。耿仕丽等[24]在对紫果西番莲叶Passiflora edulis进行低温胁迫后发现:随着低温胁迫时间的增加,紫果西番莲植株的叶绿素荧光参数各项指标都表现出下降趋势。夏溪等[25]研究了杜鹃花Rhododendron simsii在高温胁迫下的叶绿素荧光响应,发现叶绿素荧光参数下降的幅度随高温胁迫程度的加强而增加。PSⅡ中,Wk是反映PSⅡ供体侧伤害程度的指标,Vj代表受体侧QA之后电子传递链的受损程度,PIABS能够衡量PSⅡ整体性能,ΔI/IO则反映PSⅠ的最大氧化还原能力,表示PSⅠ的活性[26]。本研究中,25 ℃处理后白叶的PIABS显著上升,Wk、Vj下降,说明25 ℃处理促进了花叶矢竹白叶PSⅡ整体性能的提升,同时ΔI/IO与ΦPSⅠ/PSⅡ均未出现显著变化。15和35 ℃处理下的白叶PIABS和Wk均无显著差异,而Vj在35 ℃处理下显著提高,说明15和35 ℃处理下的花叶矢竹白叶PSⅡ的整体性能无显著提升,但35 ℃处理下的白叶PSⅡ受体侧受伤,同时ΦPSⅠ/PSⅡ均出现显著降低,而只有15 ℃处理下的ΔI/IO显著下降。因此,15 ℃处理下白叶是PSⅠ本身受到损伤,导致协调性降低,而35 ℃处理下白叶是由于PSⅡ受体侧受损,导致协调性下降。
-
植物叶绿体发育异常是导致叶色变化的主要原因之一。ZHANG等[27]在对海棠Malus spectabilis叶色突变体研究时发现:黄化叶片的叶绿体内包含不规则排列的小泡和更多的嗜锇颗粒,且类囊体层结构被破坏。在水稻突变体tcd9的研究中发现:20 ℃低温下的水稻叶绿体结构存在基粒片层堆叠不紧密以及缺乏结构良好的类囊体膜的问题[28]。GAO等[29]在番茄Solanum lycopersicum突变体wv中发现:相比于成熟叶,低温处理下wv幼叶叶片叶绿体内缺乏规整的类囊体膜。不同植株叶色突变体的叶绿体结构存在差异,但都导致叶绿体发育异常。本研究中,与ck相比,25 ℃处理下的白叶叶绿体内具备完整的类囊体片层,且基粒结构清晰;而在15 ℃处理下的白叶叶绿体内开始出现淀粉粒的垛叠,但未见明显的类囊体;35 ℃处理下的白叶叶绿体淀粉粒仍保持分散状态。因此,不同温度对花叶矢竹白叶叶绿体中类囊体垛叠的影响,阻碍了叶绿体的发育。
-
花叶矢竹白叶发育与温度有着密切联系。25 ℃处理后的白叶叶绿素合成过程中,从原卟啉Ⅸ到原脱植基叶绿素的相对含量显著增加,致使光合色素显著增加,叶绿体中出现完整的类囊体片层,PSⅡ供体侧和受体侧以及PSⅠ的ΔI/IO特性得到修复,PSⅡ和PSⅠ协调性未明显降低,光合性能得到提升;15 ℃处理下白叶Chl a和Chl b质量分数显著提升,叶绿体内出现淀粉粒的垛叠,但PSⅡ和PSⅠ协调性显著降低;35 ℃处理下白叶光和色素显著降低,叶绿体内淀粉粒仍保持分散状态,且协调性显著降低。因此,25 ℃处理下花叶矢竹白叶出现复绿现象,15 ℃处理延缓了白叶复绿现象,35 ℃处理下白叶复绿现象受到了抑制。
Physiological responses of Pseudosasa japonica f. akebonosuji leaf color changes to temperature
-
摘要:
目的 温度对植物叶色变化有重要影响,花叶矢竹Pseudosasa japonica f. akebonosuji观赏价值高,但叶色变化明显,探讨温度对花叶矢竹白叶转绿的生理影响,可为竹子叶色变异机制研究提供参考。 方法 将花叶矢竹花叶株分别置于不同温度(15、25和35 ℃)处理30 d,以自然温度为对照(ck),分析温度对花叶矢竹白叶颜色、光合色素、叶绿素前体物质、叶绿体超微结构和光系统性能的影响。 结果 25 ℃处理下,白叶全部复绿,光合色素质量分数显著高于其他处理(P<0.05);叶绿素合成前体物质中,25 ℃处理下的白叶从粪卟啉原Ⅲ(Coprogen Ⅲ)到原卟啉Ⅸ(Proto Ⅸ)相对含量迅速提升,提升的幅度显著高于ck (P<0.05),且质量分数从大到小依次为:25、35、15 ℃处理;25 ℃处理下,白叶的叶绿体基粒和片层类囊体清晰,而ck和15 ℃处理下均未形成正常的基粒类囊体,35 ℃处理下类囊体降解多杂质;25 ℃处理下叶片在光诱导后出现O、J、I、P相,叶片单位反应中心密度(RC/CSO)、量子产额(φPO、φEO)和性能指数(PIABS)均显著高于ck (P<0.05),从高到低依次为:25、15、35 ℃处理,光系统Ⅱ(PSⅡ)中K点的荧光强度变化(Wk)和J点的可变荧光(Vj)均下降,白叶25 ℃处理下光系统Ⅱ的整体性能提升;同时25 ℃处理下白叶光系统Ⅰ (PSⅠ)最大氧化还原能力(ΔI/IO)和光系统间协调性(ΦPSⅠ/PSⅡ)均高于15和35 ℃处理,PSⅠ活性提升。 结论 温度通过影响花叶矢竹的叶绿素前体物质造成光合色素累积水平的差异,导致PSⅠ和PSⅡ性能的变化,进而影响叶绿体基粒类囊体的组装,综合表现为25 ℃处理促进白叶复绿,35 ℃处理抑制白叶复绿。图6表3参29 Abstract:Objective Temperature has a significant impact on the color changes of plant leaves. Pseudosasa japonica f. akebonosuji has high ornamental value, but its leaf color changes significantly. Exploring the physiological effects of temperature on greening of white leaves in P. japonica f. akebonosuji can provide a reference for the research on mechanism of leaf color variation in bamboo. Method P. japonica f. akebonosuji plants with variegated leaves were placed in constant temperature environments of 15, 25 and 35 ℃ for 30 days, with natural temperature as the control (ck). The effects of temperature on color of white leaves, content of photosynthetic pigments, content of chlorophyll precursor substances, ultrastructure of chloroplasts, and performance of photosystems were analyzed. Result Under 25 ℃ treatment, all the white leaves turned green again, and the content of photosynthetic pigments was significantly higher than the other treatments (P<0.05). Among chlorophyll synthesis precursor substances, the relative content of coproporphyrinogen Ⅲ to protoporphyrin Ⅸ in white leaves under 25 ℃ treatment increased rapidly, with a significantly higher increase than ck (P<0.05), and the quality scores were ranked from highest to lowest as follows: 25, 35, 15 ℃ treatments. Under 25 ℃ treatment, the grana and lamellar thylakoids of chloroplasts in white leaves were clear, while no normal grana thylakoids were formed under ck and 15 ℃ treatment, and the thylakoids in 35 ℃ treatment were degraded with many impurities. After light induction, the white leaves under 25 ℃ treatment showed O, J, I, and P phases. Density of reaction centers per unit area (RC/CSO), quantum yield (φPO, φEO), and performance index (PIABS) were significantly higher than ck (P<0.05), from highest to lowest: 25, 15, 35 ℃ treatments. Fluorescence intensity change at point K (Wk) and variable fluorescence at point J (Vj) in photosystem (PS)Ⅱ both decreased, indicating an overall improvement in the performance of PSⅡ under 25 ℃ treatment with white leaves. At the same time, ΔI/IO and ΦPSⅠ/PSⅡ values of white leaves under 25 ℃ treatment were higher than those under 15 and 35 ℃, and activity of PSⅠ increased. Conclusion Temperature affects the content of chlorophyll precursor substances in leaves of P. japonica f. akebonosuji, leading to differences in accumulation level of photosynthetic pigments, which in turn causes changes in performance of PSⅠ and PSⅡ, and affects the assembly of grana thylakoids in chloroplasts. The overall effect is that 25 ℃ treatment promotes the greening of white leaves, while 35 ℃ treatment inhibits it. [Ch, 6 fig. 3 tab. 29 ref.] -
图 1 不同温度处理前后花叶矢竹白叶的表型特征
Figure 1 Morphological characteristics of white leaves of P. japonica f. akebonosuji before and after different temperature treatments
图 2 不同温度处理下花叶矢竹白叶的超微结构
Figure 2 Ultrastructure of white leaves of P. japonica f. akebonosuji under different temperature treatments
图 3 不同温度处理下花叶矢竹白叶之间的叶绿素荧光诱导动力学曲线
Figure 3 Dynamic curves for chlorophyll a fluorescein of white leaves of P. japonica f. akebonosuji under different temperature treatments
图 4 不同温度处理下花叶矢竹白叶K点的荧光强度变化(Wk)和J点的可变荧光(Vj)
Figure 4 Changes in fluorescence intensity at K point (Wk) and variable fluorescence at J point (Vj) of white leaves of P. japonica f. akebonosuji under different temperatures
图 5 花叶矢竹白叶不同温度处理间的PSⅠ 820 nm相对光吸收值变化
Figure 5 Changes of relative absorbtion of PSⅠ 820 nm white leaves of P. japonica f. akebonosuji under different temperature treatments
图 6 花叶矢竹白叶不同温度处理间的PSⅠ最大氧化还原能力(ΔI/IO)及PSⅠ与PSⅡ之间的协调性变化(ΦPSⅠ/PSⅡ)
Figure 6 PSⅠ maximum ability of oxido-reduction (ΔI/IO) and coordination between PSⅠ and PSⅡ (ΦPSⅠ/PSⅡ) of the white leaves of P. japonica f. akebonosuji under different temperature treatments
表 1 不同温度处理下花叶矢竹白叶的光合色素
Table 1. Photosynthetic pigment in white leaves of P. japonica f. akebonosuji under different temperatures
温度/℃ Chl a/(µg·g−1) Chl b/(µg·g−1) 总叶绿素/(µg·g−1) 类胡萝卜素/(µg·g−1) Chl a/b ck 62.41±0.16 c 39.66±0.29 c 102.07±0.25 c 57.35±0.07 b 1.57±0.01 b 15 75.60±0.14 b 51.44±0.49 b 127.05±0.63 b 29.81±0.10 c 1.47±0.01 b 25 239.10±0.61 a 103.30±0.67 a 342.48±1.29 a 95.67±0.13 a 2.31±0.01 a 35 31.48±1.24 d 33.96±2.05 d 65.44±1.71 d 24.04±0.44 d 0.92±0.07 c 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 不同温度处理下花叶矢竹白叶的叶绿素合成前体物质
Table 2. Chlorophyll synthesis precursors in white leaves of P. japonica f. akebonosuji under different temperatures
温度/℃ δ-氨基乙酰丙酸/
(µmol·g−1)胆色素原/
(µmol·g−1)尿卟啉原Ⅲ/
(mol·g−1)粪卟啉原Ⅲ/
(mol·g−1)原卟啉Ⅸ/
(µmol·g−1)镁原卟啉Ⅸ/
(µmol·g−1)原脱植基叶绿素/
(µmol·g−1)ck 1.36±0.02 b 2.11±0.05 c 28.58±0.09 d 63.39±0.33 b 44.20±0.47 d 18.47±0.23 d 7.77±0.32 d 15 1.18±0.04 d 4.67±0.03 a 31.63±0.22 c 62.03±0.45 c 53.21±0.87 c 23.65±0.14 c 11.31±0.33 c 25 1.27±0.09 c 3.69±0.02 b 32.24±0.24 b 59.99±0.00 d 205.84±0.43 a 113.41±0.12 a 86.70±0.12 a 35 1.46±0.04 a 3.69±0.01 b 35.28±0.16 a 66.81±0.17 a 96.15±1.01 b 44.36±0.18 b 23.83±0.31 b 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。
下载: 导出CSV
表 3 不同温度处理下花叶矢竹白叶叶绿素荧光参数的变化
Table 3. Changes in chlorophyll fluorescence parameters of the white leaves of P. japonica f. akebonosuji under different temperatures
温度/℃ ABS/RC DIO/RC TRO/RC ETO/RC RC/CSO ck 41.23±2.19 a 37.42±3.15 a 3.41±0.96 a 2.32±0.26 a 12.5±3.91 b 15 17.35±5.49 b 14.27±5.49 bc 3.13±0.07 a 0.95±0.22 a 53.2±29.0 b 25 3.02±0.30 c 0.85±0.24 c 2.18±0.06 a 0.91±0.08 a 1876.9 ±438.6 a35 30.21±5.84 ab 28.13±6.54 ab 2.07±0.71 a 0.15±0.01 a 18.70±6.80 b 温度/℃ φPO(Fv/Fm) φEO ψO PIABS ck 0.07±0.04 b 0.02±0.01 b 0.29±0.11 a 0.00±0.00 b 15 0.17±0.09 b 0.05±0.02 b 0.27±0.06 a 0.00±0.00 b 25 0.73±0.05 a 0.30±0.02 a 0.42±0.03 a 0.66±0.18 a 35 0.08±0.02 b 0.01±0.00 b 0.04±0.00 b 0.00±0.00 b 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。
下载: 导出CSV
-
[1] 安苗苗, 刘静, 郦元, 等. 花叶矢竹转录组中的转座子表达分析[J]. 浙江农林大学学报, 2016, 33(6): 935−943. AN Miaomiao, LIU Jing, LI Yuan, et al. Transposons expression analysis of transcriptome in Pseudosasa japonica f. akebonosuji[J]. Journal of Zhejiang A&F University, 2016, 33(6): 935−943. DOI: 10.11833/j.issn.2095-0756.2016.06.003. [2] 许冰清, 安苗苗, 姜可以, 等. 花叶矢竹叶绿体psbD基因的克隆与功能分析[J]. 浙江农林大学学报, 2015, 32(4): 557−565. XU Bingqing, AN Miaomiao, JIANG Keyi, et al. Cloning and functional analysis of the psbD gene in Pseudosasa japonica f. akebonosuji[J]. Journal of Zhejiang A&F University, 2015, 32(4): 557−565. DOI: 10.11833/j.issn.2095-0756.2015.04.010. [3] 陈凌艳, 何丽婷, 赖金莉, 等. 银丝竹不同叶色叶绿素合成及叶结构差异[J]. 森林与环境学报, 2017, 37(4): 385−391. CHEN Lingyan, HE Liting, LAI Jinli, et al. The variation of chlorophyll biosynthesis and the structure in different color leaves of Bambusa multiplex ‘Silverstripe’[J]. Journal of Forest and Environment, 2017, 37(4): 385−391. DOI: 10.13324/j.cnki.jfcf.2017.04.001. [4] 徐薪璐, 蔡鸥, 赵婉琪, 等. 靓竹彩叶形成的细胞学以及光合色素[J]. 林业科学, 2023, 59(8): 40−51. XU Xinlu, CAI Ou, ZHAO Wanqi, et al. Changes in cytology and photosynthetic pigments during the formation of colored leaves of Sasaella glabra ‘Albostriata’[J]. Scientia Silvae Sinicae, 2023, 59(8): 40−51. DOI: 10.11707/j.1001-7488.LYKX20220802. [5] 徐薪璐, 蔡鸥, 秦敏, 等. 植物叶色变异研究进展[J/OL]. 分子植物育种, 2022-05-17. https://kns.cnki.net/kcms/detail/46.1068.S.20220517.1326.020.html. XU Xinglu, CAI Ou, QIN Min, et al. Research progress on leaf color variation in plants[J/OL]. Molecular Plant Breeding, 2022-05-17. https://kns.cnki.net/kcms/detail/46.1068.S.20220517.1326.020.html. [6] GONG Xiaodi, JIANG Quan, XU Jianlong, et al. Disruption of the rice plastid ribosomal protein S20 leads to chloroplast developmental defects and seedling lethality[J]. G3 Genes|Genomes|Genetics, 2013, 3(10): 1769−1777. DOI: 10.1534/g3.113.007856. [7] DU Jingjing, WANG Junwei, SHAN Sicong, et al. Low-temperature-mediated promoter methylation relates to the expression of TaPOR2D, affecting the level of chlorophyll accumulation in albino wheat (Triticum aestivum L.)[J]. International Journal of Molecular Sciences, 2023, 24(19): 14697. DOI: 10.3390/ijms241914697. [8] SUN Ying, MA Qilong, MAO Lianzhen, et al. Integrated transcriptome and metabolome analysis reveals the mechanism of carotenoid regulation in the yellowing-leaf mutant of pepper (Capsicum annuum L.) in response to different temperatures[J]. Scientia Horticulturae, 2024, 323: 112530. DOI: 10.1016/j.scienta.2023.112530. [9] LIU Weikang, CHEN Guangling, HE Mingming, et al. ABI5 promotes heat stress-induced chlorophyll degradation by modulating the stability of MYB44 in cucumber[J]. Horticulture Research., 2023, 10(6): uhad089. DOI: 10.1093/hr/uhad089. [10] ZHANG Yuting, FENG Xin, LIU Yang, et al. A single-base insertion in BoDFR1 results in loss of anthocyanins in green-leaved ornamental kale[J]. Theoretical and Applied Genetics, 2022, 135(6): 1855−1865. DOI: 10.1007/s00122-022-04079-y. [11] ZHENG Kailun, ZHAO Jian, LIN Dongzhi, et al. The rice TCM5 gene encoding a novel deg protease protein is essential for chloroplast development under high temperatures[J]. Rice, 2016, 9(1): 13. DOI: 10.1186/s12284-016-0086-5. [12] LIU Yichao, WANG Shijie, FENG Shuxiang, et al. Combined whole transcriptome analysis and physical-chemical reveals the leaf color change mechanism of Ulmus pumila under heat stress[J]. Environmental and Experimental Botany, 2023, 210: 105347. DOI: 10.1016/j.envexpbot.2023.105347. [13] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000. LI Hesheng. Principles and Techniques of Plant Physiological Biochemical Experiment[M]. Beijing: Higher Education Press, 2000. [14] DEI M. Benzyladenine-induced stimulation of two components of chlorophyII formation in etiolated cucumber cotyledons[J]. Physiologia Plantarum, 1984, 62(4): 521−526. DOI: 10.1111/j.1399-3054.1984.tb02793.x. [15] BOGORAD L. Porphyrin synthesis[J]. Methods in Enzymology, 1962, 5: 885−895. [16] 杨海芸. 花叶矢竹叶色变异机理研究[D]. 北京: 北京林业大学, 2015. YANG Haiyun. Study on Mechanism of Leaf Color Variation of Pseudosasa japonica f. akebonosuj[D]. Beijing: Beijing Forestry University, 2015. [17] 汪瑛琦. 遮荫和温度对光敏白化茶树‘黄金芽’叶色影响及其潜在机制[D]. 杭州: 浙江大学, 2023. WANG Yingqi. Effect of Shading and Temperature on the Leaf Color of the Light-sensetive Albino Tea Plant ‘Huangjinya’ and Its Potential Mechanism[D]. Hangzhou: Zhejiang University, 2023. [18] 王妮, 李朝娜, 郑旭理, 等. 花叶矢竹叶片色素合成和光合特性[J]. 植物生态学报, 2024, 48(11): 1536−1546. WANG Ni, LI Zhaona, ZHENG Xuli, et al. Pigment synthesis and photosynthetic characteristics of leaves in Pseudosasa japonica f. akebonosuji[J]. Chinese Journal of Plant Ecology, 2024, 48(11): 1536−1546. DOI: 10.17521/cjpe.2023.0258. [19] ZHAO Minghui, LI Xiang, ZHANG Xinxin, et al. Mutation mechanism of leaf color in plants: a review[J]. Forests, 2020, 11(8): 851. DOI: 10.3390/f11080851. [20] 毛晶晶, 李泽娇, 赵雨晴, 等. 低温胁迫对玉米转绿过程中叶绿素生物合成的影响[J]. 四川农业大学学报, 2019, 37(5): 617−622. MAO Jingjing, LI Zejiao, ZHAO Yuqing, et al. The effects of low temperature on chlorophyll synthesis during greening of maize[J]. Journal of Sichuan Agricultural University, 2019, 37(5): 617−622. DOI: 10.16036/j.issn.1000-2650.2019.05.005. [21] 崔海瑞, 夏英武, 高明尉. 温度对水稻突变体W1叶色及叶绿素生物合成的影响[J]. 核农学报, 2001, 15(5): 269−273. CUI Hairui, XIA Yingwu, GAO Mingwei. Effects of temperature on leaf color and chlorophyll biosynthesis of rice mutant W1[J]. Acta Agriculturae Nucleatae Sinica, 2001, 15(5): 269−273. DOI: 10.3969/j.issn.1000-8551.2001.05.003. [22] CHEN Lingyan, LAI Jinli, HE Tianyou, et al. Differences in photosynthesis of variegated temple bamboo leaves with various levels of variegation are related to chlorophyll biosynthesis and chloroplast development[J]. Journal of the American Society for Horticultural Science, 2018, 143(2): 144−153. DOI: 10.21273/jashs04359-18. [23] 李璐瑶, 张君, 应学兵, 等. 红膜对3种蔬菜光合性能与果实品质的影响[J]. 浙江农林大学学报, 2025, 42(3): 564−571. LI Luyao, ZHANG Jun, YING Xuebing, et al. Effects of red film on photosynthetic abilities and fruit qualities in 3 vegetables[J]. Journal of Zhejiang A&F University, 2025, 42(3): 564−571. DOI: 10.11833/j.issn.2095-0756.20240552. [24] 耿仕丽, 王雪, 陈诗, 等. 低温胁迫下不同倍性紫果西番莲叶绿素荧光参数差异分析[J]. 中国南方果树, 2026, 55(2): 65−70. GENG Shili, WANG Xue, CHEN Shi, et al. Differential analysis of chlorophyll fluorescence parameters in different ploidy Passiflora edulis under low temperature stress[J]. South China Fruits, 2026, 55(2): 65−70. DOI: 10.13938/j.issn.1007-1431.20240634. [25] 夏溪, 龚睿, 冷寒冰, 等. 高温胁迫下杜鹃花叶绿素荧光响应及耐热性评价[J]. 北方园艺, 2025(6): 82−89. XIA Xi, GONG Rui, LENG Hanbing, et al. Comprehensive evaluation of chlorophyll fluorescence response and heat tolerance of different Azalea cultivars to high-temperature stress[J]. Northern Horticulture, 2025(6): 82−89. DOI: 10.11937/bfyy.20243338. [26] 王仁杰, 朱凡, 梁惠子, 等. 重金属Mn对苦楝叶片光系统性能的影响[J]. 生态学报, 2020, 40(6): 2019−2027. WANG Renjie, ZHU Fan, LIANG Huizi, et al. Effects of Manganese (Mn) on the performances of photosystems Ⅰ and Ⅱ in Melia azedarach young plant[J]. Acta Ecologica Sinica, 2020, 40(6): 2019−2027. DOI: 10.5846/stxb201810282325. [27] ZHANG Lulu, ZHANG Junkang, MAO Yunfei, et al. Physiological analysis and transcriptome sequencing of a delayed-green leaf mutant ‘Duojiao’ of ornamental crabapple (Malus sp.)[J]. Physiology and Molecular Biology of Plants, 2022, 28(10): 1833−1848. DOI: 10.1007/s12298-022-01248-7. [28] JIANG Quan, MEI Jie, GONG Xiaodi, et al. Importance of the rice TCD9 encoding α subunit of chaperonin protein 60 (Cpn60α) for the chloroplast development during the early leaf stage[J]. Plant Science. 2014, 215/216: 172−179. DOI: 10.1016/j.plantsci.2013.11.003. [29] GAO Shenghua, GAO Wenjing, LIAO Xiaoli, et al. The tomato WV gene encoding a thioredoxin protein is essential for chloroplast development at low temperature and high light intensity[J]. BMC Plant Biology, 2019, 19(1): 265. DOI: 10.1186/s12870-019-1829-4. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250294
 点击查看大图
点击查看大图

计量
- 文章访问数: 89
- HTML全文浏览量: 24
- PDF下载量: 19
- 被引次数: 0
