





-
城市热岛效应成为困扰城市发展的主要障碍和难点,如何通过生态途径协调城市发展与环境问题之间的关系,成为当前研究的热点。自然山水架构在调和城市中的人工—自然矛盾体之间发挥了重要的生态效能,为解决城镇化进程中出现的“城市病”提供了最基本的生态解决途径之一。绿色廊道将城市公园、街头绿地、自然保护地、农田、滨水湿地和山地等串联构成具有自我维持能力的动态绿色景观结构体系[1]。作为城市最重要的生态载体之一,绿色廊道的空间分布和结构特征对改善城市热环境具有重要的生态效益。不同的绿地类型、形态、结构和植物群落等构成的绿色廊道对降低城市热岛强度的生态调节功能和效益各不相同。绿色廊道按空间结构特征分为:线状绿色廊道、带状绿色廊道和河流型绿色廊道等[2]。按功能类型分为:生态型绿色廊道、文化与历史遗产型绿色廊道、休闲游憩型绿色廊道等[3]。
绿色廊道的路径选线直接影响其空间属性特征,因此,通过优化绿色廊道的景观格局来降低城市的热岛强度是行之有效的生态途径和方法。根据不同的研究目标,研究人员基于兴趣点( point of interest,POI)、选线适宜性评价、生态适宜性分析、多源数据与深度学习、层次分析法、重力模型、最小累积阻力模型(minimum cumulative model,MCR)、环境公平视角等方法对绿色廊道的选线和评价进行了深入探讨[4-9]。有研究表明:当城镇绿化覆盖率达到50%时,地表温度下降约13 ℃,城市热岛效应可基本消除[10]。本研究以改善城市热环境为目标,基于MCR模型与水文流域模型,对福建省福州市绿色廊道的景观格局进行优化分析,为城市规划和建设提供参考。
-
福州市位于中国东南沿海(25°15′~26°39′N,118°08′~120°31′E),属于中、南亚热带海洋性季风气候区,常年雨量充沛,夏季高温湿热,逆温频率高,持续时间长,城市热岛效应明显。福州市位于河口盆地中心,主城区被海拔600~1 000 m的莲花山、鼓山、五虎山和旗山等群山环绕,形成“枕山、襟江、面海”的自然地理环境特征。为更全面地分析热岛效应与绿色廊道的空间关系,研究范围选取鼓楼区、台江区、晋安区、仓山区、马尾区、长乐新区及闽侯县等区域,总面积约1 759.95 km²。
根据《福州市中心城区绿地系统总体规划(2011—2020年)》《福建省绿道网总体规划纲要(2012—2020)》和《福州市绿道网总体规划》,福州市共规划省级绿道3 119.0 km,其中,6条省级绿道、3条支线和2条连接线。福州市以“沿江、沿河、环湖、达山、通公园”为目标,整合城市自然生态和历史人文资源,在省级绿道总体规划框架内,共规划市级绿道1 238.0 km,其中,2条省级绿道、36条市级绿道、94条社区级绿道。目前,已建16条山地丘陵型绿道,86条环湖滨河型绿道,5条滨江面海型绿道,15条绿色通勤型绿道,10条历史文化型绿道。截至2020年,全市新建168个串珠公园绿地,中心城区的绿地率达42%,绿化覆盖率达48%,人均绿地面积超过15 m2。
-
研究数据主要包括:土地利用类型数据根据2019年福州市谷歌(Google)地图2.5 m高精度影像(谷歌地图资源共享平台下载),结合GB/T 721010—2017《土地利用现状分类》分为林地、建设用地、道路、水体、农田、草地、未利用地,分类精度为89.84%,Kappa系数0.85。地表温度数据由2019年9月22日Landsat 8 OLI遥感影像(美国地质调查局网站下载)根据辐射传输方程法反演获取[11]。2019年福州市建筑矢量数据、《福州市城市总体规划(2010—2020年)》《福州新区总体规划(2015—2020年)》《福州市中心城区绿地系统总体规划(2011—2020年)》《福州市中心城区综合交通总体规划(2011—2020年)》等来源于福州市规划局网站。福州市90 m分辨率数字高程模型(DEM)遥感影像来源于中国地理空间数据云网站。影像噪声低、几何形变小。研究区范围云量低于2%,满足研究需求。
-
根据土地利用分类数据,通过形态学空间格局模型(morphological spatial pattern analysis,MSPA)开闭运算数学方法[12],对栅格图像进行度量、识别及数据分割等操作,分别提取MSPA模型的前景数据和背景数据,得出像元层面的景观生态斑块,将其进行二值化栅格处理,运用ArcGIS软件中的Guidos Toolbox分析工具,采用八邻域图像细化分析方法[13],提取像素目标和矢量化跟踪,得到互不重叠的7种景观类型和结构:核心区、孤岛、孔隙、边缘、环道、桥接、支线,将结果按面积大小提取核心区斑块和生态源地[14]。
景观连通性指数是反映景观连通度和斑块重要性的主要依据[15-16]。本研究选取斑块整体连通性指数(IIC)和斑块面积比指数(DA)对源斑块的重要性进行评价。通过ArcGIS平台,设定源斑块之间连通的距离阀值。如果距离小于阈值,则视为连通;如果距离大于阈值,则视为不连通[17]。IIC指数通过计算2个斑块之间最短路径来确定,结果为0~1,当值为0时,说明各斑块间互不连通;当值为1时,说明整个景观单元都是生境斑块。利用Conefor Inputs for ArcGIS插件,计算关键源点间的阻力距离值[18]。将IIC指数和DA指数的结果输入Conefor Sensinode 2.6软件,设置阈值距离等参数,计算绿地斑块重要性指数(dIIC)。dIIC指数的值越高,表示各斑块在绿地系统中的重要性越高。根据dIIC指数对斑块重要性进行评价,由此划分生态源斑块和绿色廊道的等级。
-
构建绿色廊道最常用的方法是MCR模型[19],表示生态流克服生态阻力面,从一个生态源地到另一个生态源地的最小累积距离,可确定生态源地起始点和目标点之间的最短费用路径[20]。通过阻力面赋值和权重生成最佳路径,进而模拟生物运动的潜在趋势与景观格局改变之间的关系[21]。MCR模型是计算生态流克服不同土地利用类型阻力进行移动的过程,需考虑生态源点、路径距离和阻力因子,对生态敏感性影响因子进行赋值和权重计算[22]。权重值通过序关系法获取[23]。由此构建生态综合阻力面来表征景观类型对生态流动的阻力影响。利用ArcGIS插件中的Build Network and Map Linkages工具计算各生态源地到各斑块之间的费用距离栅格(距离测算)和成本回溯链接栅格数据(费用路径),利用最短路径函数计算并生成“源”到各“网络中心”耗费最少、流动性成本最小的潜在绿色廊道。公式为:
$$ {C}_{i}=\sum _{j=1}^n({D}_{i}\times {F}_{j}) 。$$ 其中:Ci表示从景观单元i (i=1,2,3
$,\cdots, $ m)到生态源的累积耗费距离值,即最小成本的计算结果;Di表示景观单元i到生态源的间隔距离;Fj表示空间中景观单元j (j=1,2,3$,\cdots, $ n)对物种迁移的阻抗值。 -
基于DEM数据,根据水文流域模型对水的径流方向及路径进行模拟和定量化分析[24]。同理,在ArcGIS平台中,将地表反演温度和绿色廊道生态综合阻力面的值等同于DEM数据,利用栅格计算器将各因子叠加的适宜性图取反后,整个区间为负数,再将取反的适宜性图的最小值取绝对值,使最小值为0,得到一张“反适宜性”图,即取值高的地方不适宜,取值低的地方适宜。通过对“反适宜性”图进行填洼—流向—流量等步骤处理后,利用栅格计算器设定阈值。借助Flow Accumulation工具,按场地特征不断实验阈值来提取温度分布域和绿色廊道分布域,采用Shreve河网分级工具对两者进行分级。
将温度分布域和绿色廊道分布域相叠加,两者的交点视为重要降温生态节点,对降低热岛强度具有重要作用。结合绿色廊道内部廊道之间的交点,与地表温度相叠加,提取未被绿色廊道覆盖的“热点”区域,进而优化绿色廊道的空间分布格局。具体流程如图1。
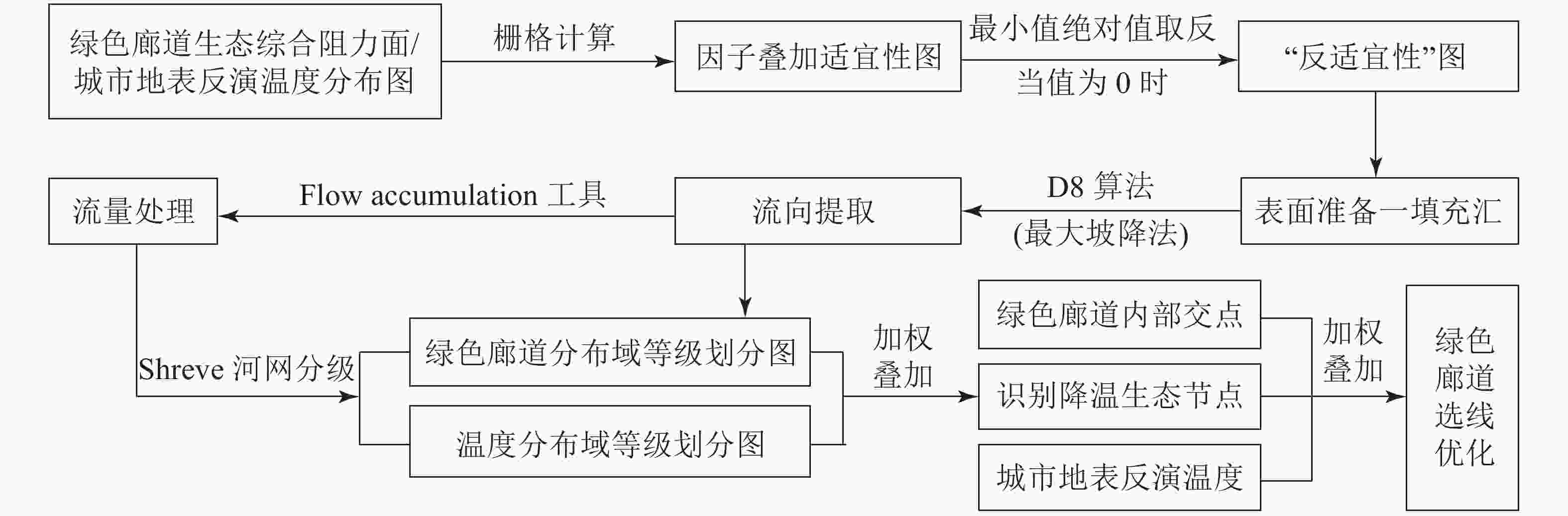
图 1 基于水文流域模型的绿色廊道优化流程图
Figure 1. Greenway optimization flow chart based on hydrological basin model
-
利用ArcGIS平台,将福州市土地分类数据的林地、草地和农田设为MSPA模型的前景数据,建设用地、道路、水体和未利用地设为背景数据,基于MSPA模型对提取的7种景观类型按面积从大到小排序:核心区、边缘区、桥接区、孤岛、支线、环道区、孔隙,从中筛选出面积排名前30位的核心区斑块作为生态源斑块,并进行编号(表1)。
表 1 福州市重要生态源地统计表
Table 1. Overview of important ecological sources in Fuzhou City
编号 面积/km² 区位 编号 面积/km² 区位 编号 面积/km² 区位 1 126.77 闽侯县、晋安区 11 3.24 长乐新区 21 2.96 长乐新区 2 1.98 晋安区 12 5.35 长乐新区 22 2.87 长乐新区 3 2.03 闽侯县、鼓楼区 13 1.74 仓山区 23 2.07 长乐新区 4 1.65 晋安区 14 3.42 长乐新区 24 20.97 长乐新区 5 13.65 马尾区 15 1.65 闽侯县 25 3.16 长乐新区 6 2.17 马尾区 16 1.90 长乐新区 26 1.62 长乐新区 7 1.56 马尾区 17 1.63 闽侯县 27 158.00 闽侯县 8 3.85 马尾区 18 73.07 闽侯县 28 31.08 长乐新区 9 17.68 闽侯县 19 2.53 长乐新区 29 5.86 长乐新区 10 140.47 马尾区 20 4.06 闽侯县 30 100.87 闽侯县 将源斑块间连通距离的阈值设为2 km。当距离小于2 km时,2个斑块间视为可连通;当距离大于2 km时,则视为不连通。利用自然断点法对30个生态源斑块的重要性进行评价,共分为4级:一级最重要源斑块2个,dIIC值为25.780~39.620;二级非常重要源斑块3个,dIIC为7.610~25.780;三级很重要源斑块3个,dIIC为1.980~7.610;四级一般重要源斑块22个,dIIC为0.002~1.980。源斑块重要性分布如表2。
表 2 源斑块重要性分级统计
Table 2. Statistical graph of the importance classification to source plaques
斑块重要性 斑块等级 dIIC区间值 斑块数/个 面积/km2 比例/% 斑块编号 区位说明 最重要 一级 25.780~39.620 2 298.47 40.34 10、27 包括旗山和鼓山自然保护区、风景名胜区、国家森
林公园、重要水源及水库保护区等非常重要 二级 7.610~25.780 3 300.71 40.64 1、18、30 包括莲花山、五虎山和天台山自然林地、森林公
园、重要水源及水库保护区、大面积山地公
园、水源涵养保护区等很重要 三级 1.980~7.610 3 52.61 7.11 8、9、28 包括马尾琅岐旗山公园、闽侯六路山和长乐新区寨
顶山自然林地、大型山地公园、重要湿地等一般重要 四级 0.002~1.980 22 88.08 11.91 剩余斑块 包括大型山地公园、基本农田、大型水库、重要湿
地等 -
结合福州市河口型盆地特征,按生态适应性分析法[25],选取土地利用类型、地形因子、生境因子、景观类型、生态系统服务等5个生态敏感性因子,通过计算赋值和权重值,得到各类景观单元的生态景观阻力面分布特征。由此可知:生态景观阻力值较高的区域集中于鼓楼区、台江区、仓山区、马尾区和晋安区等主城区。其中,长乐新区生态景观阻力值较高的区域集中于老城区、营前新区、各乡镇建成区及机场等,闽侯县集中于大学城、高新区、甘蔗县城、青口镇区、南屿镇区和南通镇区等。闽江流域因对物种迁移和扩散构成较大阻力,生态景观阻力值较高;环绕主城区的五虎山、旗山、莲花山、鼓山和天台山等大面积自然林地,生态景观阻力值较小,构成生境斑块的基底。
-
将5个生态敏感性因子的景观生态阻力面进行加权叠加,得到综合生态阻力面。基于MCR最小累积阻力模型的最小路径工具计算起点与终点的最小费用路径,模拟源斑块间相互连接的潜在绿色廊道。
将筛选出的30个重要生态源地与城市道路矢量数据、城市水系矢量数据、潜在绿色廊道等进行加权叠加,获得福州市潜在绿色廊道合成图。由此可知:潜在绿色廊道共74条,在主城区鼓楼区、台江区、仓山区中北部、晋安区主城区、马尾区主城区、闽侯上街和高新区等分布均比较少,主要因为这些区域建设密度大,土地利用集约化程度高,地形因子影响大,综合生态阻力高。在仓山区东部、马尾琅岐、长乐新区滨海新城、闽侯南屿镇至青口区域、晋安区北部等分布比较多,说明这些区域生态阻力小,重要斑块分布多,景观破碎度低,连通性高,利于绿色廊道生态路径的形成。
结合源斑块重要性等级划分结果,将起点或终点为一级源斑块的廊道定义为一级廊道,将起点或终点为二级源斑块的廊道定义为二级廊道,其他等级以此类推。因此,如表3可见:福州市绿色廊道分为4级,共74条,总长度918.11 km。其中,一级9条,132.88 km,占14.47%;二级14条,207.48 km,占22.60%;三级14条,153.57 km,占16.73%;四级37条,424.18 km,占46.20%。
表 3 福州市绿色廊道分级概况
Table 3. Classification of greenway in Fuzhou City
廊道类型 数量/条 长度/km 绿色廊道路径 一级 9 132.87 ①斑块27(旗山国家森林公园)—斑块9(闽侯六路山);②斑块27—斑块15(闽侯官塘山);③斑块
27—斑块30(五虎山);④斑块10(鼓山风景名胜区)—斑块2(鼓岭风景名胜区);⑤斑块10—斑
块5(马尾琅岐南山尾);⑥斑块10—斑块8(马尾琅岐旗山);⑦斑块10—斑块13(城门山);
⑧斑块10—斑块17(南山公园);⑨斑块10—斑块18(天台山)二级 14 207.48 ①斑块1(莲花山)—斑块3(金鸡山公园);②斑块1—斑块4(鼓岭);③斑块1—斑块9(闽侯六路
山);④斑块18(天台山)—斑块6(牛岭山);⑤斑块18—斑块8(琅岐旗山);⑥斑块18—斑块
10(鼓山风景名胜区);⑦斑块18—斑块12(火焰山);⑧斑块18—斑块14(长乐烟台山);⑨斑块
18—斑块17(南山公园);⑩斑块18—斑块19(龙角峰公园);⑪斑块18—斑块23(长乐莲花山);
⑫斑块30(五虎山)—斑块15(闽侯官塘山);⑬斑块30—斑块20(祥谦林森公园);⑭斑块30—斑
块27(旗山国家森林公园)三级 14 153.57 ①斑块9(六路山)—斑块1(莲花山);②斑块9—斑块3(金鸡山公园);③斑块9—斑块15(闽侯官塘
山);④斑块9—斑块27(旗山国家森林公园);⑤斑块8(马尾琅岐旗山)—斑块5(马尾琅岐南山
尾);⑥斑块8—斑块6(牛岭山);⑦斑块8—斑块10(鼓山风景名胜区);⑧斑块8—斑块12(火焰
山);⑨斑块8—斑块18(天台山);⑩斑块28(寨顶山)—斑块17(南山公园);⑪斑块28—斑块
23(长乐莲花山);⑫斑块28—斑块24(塔山);⑬斑块28—斑块26(古槐镇农保地);⑭斑块28—
斑块29(三溪水库)四级 37 424.18 主要由城市大型山地公园、农保地、大型水库、河流湿地等四级斑块之间相互连接,构成建成
区的主要绿色廊道 -
绿色廊道各路径之间汇聚相交的点是重要的生态节点,对降低城市热岛强度具有重要的作用。利用ArcGIS平台,识别绿色廊道内部的生态“交点”。由此:共生成80个生态“交点”,主要分布于城区周边的自然林地生态斑块,以及城区内部的大型城市绿地和山体公园,中心主城区因绿地斑块破碎化严重,连通性较差,内部生态“交点”稀少。其中,鼓楼区2个,仓山区3个,晋安区6个,台江区无生态“交点”。马尾区和闽侯县因分布数量较多的自然生态林地和森林公园等,分别有生态“交点”13个和14个,长乐新区因分布大面积的农田、山体公园、湿地保护区等,生态“交点”高达42个。
根据水文流域模型,将绿色廊道分布域图和温度分布域图叠加,共识别降温生态节点176个。由图2可知:鼓楼区有降温生态节点8个,仓山区21个,晋安区14个,台江区1个,马尾区27个,长乐新区63个,闽侯县42个。中心城区生态节点分布较少,说明绿色廊道对主城区热岛效应的缓解作用较弱,需对其数量和结构进行优化,增加主城区的降温生态节点,提升城区夏季的热舒适度,增强绿色廊道的生态效益。
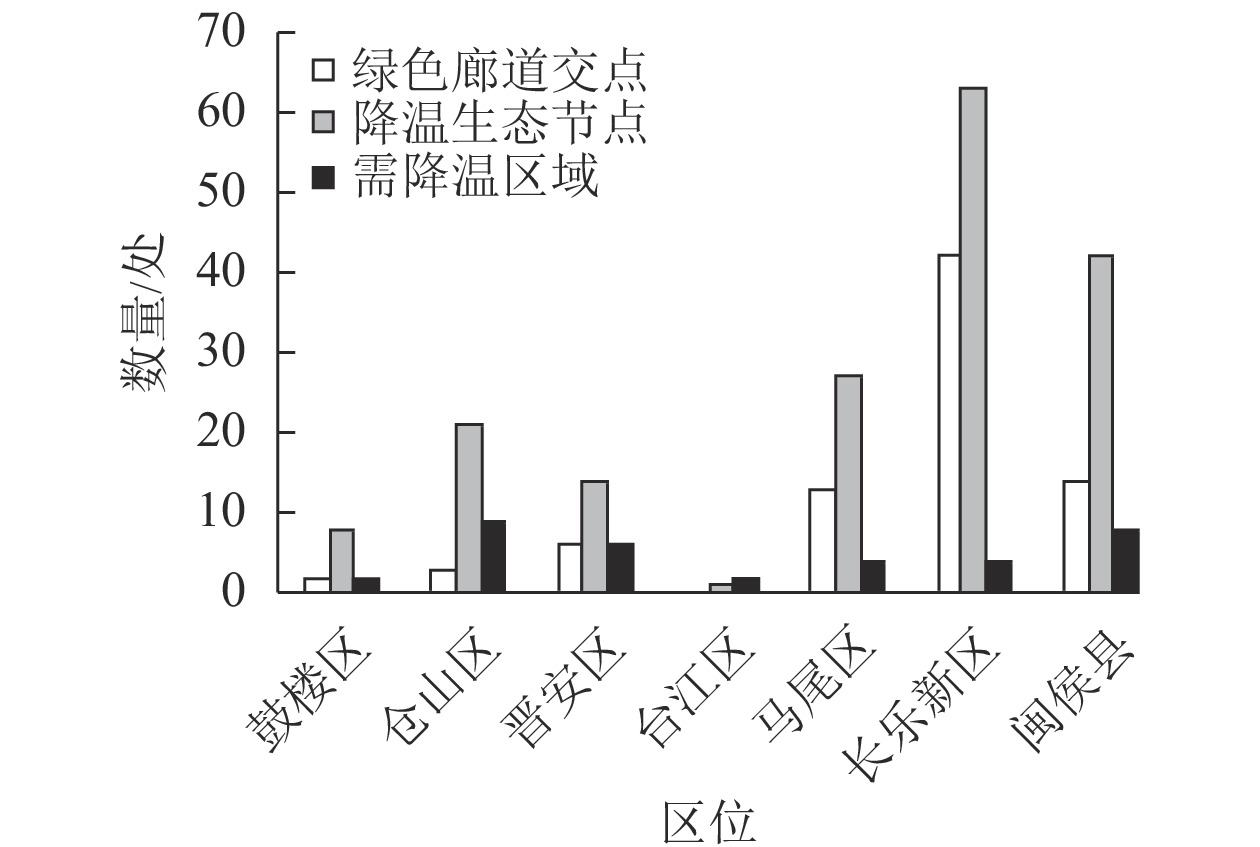
图 2 绿色廊道降温生态节点和需降温区域分布
Figure 2. Distribution of cooling ecological nodes and cooling areas in greenway
-
将福州市绿色廊道与地表温度叠加分析,由此识别出35处需降温区域(图2),其中,鼓楼区2处、仓山区9处、晋安区6处、台江区2处、马尾区4处、长乐新区4处、闽侯县8处。以识别的降温区域为导向,基于缓解城市热岛效应为目标,将福州市绿色廊道与城市用地分类矢量数据、绿色廊道生态“交点”、降温生态节点、地表温度等叠加后,在原有74条绿色廊道的基础上,新增绿色廊道14条,主要包括道路型绿色廊道和滨水型绿色廊道2种类型(表4)。其中,道路型绿色廊道8条,主要由城市主干道路构成;滨水型绿色廊道6条,主要由闽江和乌龙江流域,以及城市内河构成。
表 4 福州市优化新增绿色廊道统计
Table 4. Statistical table of optimized new greenways in Fuzhou City
廊道类型 数量/条 廊道路径 道路型
绿色廊道8 ①五四北泰禾广场—福州北绕城高速—福山郊野公园—软件园—铜盘路—西湖公园—东街口—白马河—中
亭街—长安山公园—旧师大—高盖山公园—义序机场—三环路等
②闽侯六路山—国宾大道—永嘉天地—洪塘大桥—金牛山公园—梅峰路—西湖公园—华林路—牛岗山公园— 东三环路—盛丰物流园区—上洋工业区—鼓山等
③狮子峰—牛岗山公园—福光路—盛丰物流园区—上洋工业区—光明港公园等
④东街口—东大路—东二环泰禾广场等
⑤旗山国家森林公园—大学城—建平路—橘园洲大桥—金山大道—金山工业园区—金洲南路—飞凤山公园— 海峡奥林匹克体育中心—高盖山公园—白湖亭—南二环路—会展中心等
⑥浦上大桥—浦上大道—仓山万达—闽江大道—旧师大—长安山公园—世欧上江城等
⑦永嘉天地—旗山大道—大学城—高新区—南屿镇区—117县道—南港大桥—南通镇区等
⑧五虎山—奔驰大道—青口镇区—104国道—闽江绿道等、长乐洋屿工业园区—洞江—朝阳路—南山公园— 首占镇区—大象山滨水型
绿色廊道6 ①亭江镇区—马尾主城区—马尾工业园区—台江金融街万达—中亭街—闽侯荆溪永丰—甘蔗城区等沿闽江
北岸构成
②火车南站—会展中心—世欧上江城—闽江公园—淮安大桥—三环快速路乌龙江段—火车南站等形成的环
仓山区南台岛分布
③闽侯候官村—洪塘大桥—乌龙江大道—南屿镇区—203省道—福昆线—长乐洋屿工业园区—马尾琅岐镇区
等沿乌龙江及闽江构成
④长乐旗山—两港工业区—滨海新城—龙角峰公园等沿海岸线构成
⑤五四北泰禾广场—新店溪及秀峰路—火车北站—晋安河—世欧王庄—光明港—闽江等沿内河构成
⑥世欧王庄—光明港—上洋工业区—鼓岭等沿光明港构成结合《福州市城市总体规划(2010—2020年)》《福州新区总体规划(2015—2020年)》《福州市中心城区绿地系统总体规划(2011—2020年)》《福州市中心城区综合交通总体规划(2011—2020年)》等,对新增绿色廊道的路径选线进行合理性验证和分析,研究结果基本满足要求。优化后的绿色廊道共88条,廊道数量和类型更丰富,景观格局空间分布和结构特征更合理。绿色廊道对城市热点区域的覆盖更全面,对改善城市热岛具有更明显的生态调节功能。在福州市大山水格局的生态框架下,结合城市总体规划和用地布局,对福州市绿色廊道的宽度、走向、植物配置形式等方面进行科学规划和全局统筹,并通过制定行之有效的保护措施和法律法规,提升绿色廊道的空间结构分布特征,发挥绿色廊道的生态效益和功能。
-
本研究选取具有河口型盆地地貌特征的福州市为对象,以改善城市热环境为目标,基于MCR模型和水文流域模型,对福州市绿色廊道的景观格局进行优化和分析。主要结论如下:①根据MSPA模型和源斑块重要性评价,将提取的30个生态源斑块分为4级:一级最重要斑块(2个)、二级非常重要斑块(3个)、三级很重要斑块(3个)、四级一般重要斑块(22个)。结合生态适应性分析法,由土地利用类型、地形因子、生境因子、景观类型、生态系统服务等5个生态敏感性因子构建综合生态阻力面。②通过MCR模型计算综合生态阻力面的最小成本路径,由此生成74条绿色廊道,总长度918.11 km:一级9条,132.88 km;二级14条,207.48 km;三级14条,153.57 km;四级37条,424.18 km。将绿色廊道与城市地表温度叠加,共识别80个生态“交点”、35处未覆盖需降温区域,表明绿色廊道的降温效果不够明显,景观格局需进一步优化提升。③利用水文流域模型,将绿色廊道分布域与温度分布域叠加生成176个降温生态节点。据此新增14条绿色廊道:8条道路型绿色廊道和6条滨水型绿色廊道。优化后的福州市绿色廊道共计88条,廊道路径与城市上位规划相符合,整体空间分布更合理,对城市热点区域覆盖更全面,降温效果更明显,缓解城市热岛效应的生态调节功能更突出。
Optimization of greenways in Fuzhou based on heat island effect
-
摘要:
目的 绿色廊道的空间分布和结构特征对改善城市热环境具有重要的生态效益,绿色廊道的路径选线直接影响其属性特征,因此,通过优化绿色廊道的景观格局来降低城市的热岛强度是行之有效的生态途径和方法。 方法 选取福建省福州市为研究对象,以缓解热岛效应为目标,基于最小累积阻力模型(minimum cumulative model,MCR)和水文流域模型,对福州市绿色廊道的景观格局进行优化布局。 结果 ①通过形态学空间格局模型(morphological spatial pattern analysis,MSPA),用提取的30个生态源斑块构建综合生态阻力面,利用MCR模型将其生成福州市绿色廊道,共74条,总长度918.11 km。根据源斑块重要性评价分为4级:一级9条,132.88 km;二级14条,207.48 km;三级14条,153.57 km;四级37条,424.18 km;②通过水文流域模型,将74条绿色廊道与福州市地表温度叠加分析,共提取80个生态“交点”、176个降温生态节点、35处需降温区域。 结论 据此福州市新增14条绿色廊道,其中道路型绿色廊道8条,滨水型绿色廊道6条。优化后的绿色廊道对福州市的热点区域覆盖更全面,对改善城市热岛问题具有更明显的生态调节功能。图2表4参25 Abstract:Objective The spatial distribution and structural characteristics of greenway have important ecological benefits for improving urban thermal environment. The route selection of greenway directly affects its attribute characteristics. Therefore, it is an effective ecological way and method to reduce the intensity of urban heat island by optimizing the landscape pattern of the greenway. Method Taking Fuzhou City, Fujian Province as the research object, aiming at alleviating the heat island effect, the landscape pattern of Fuzhou greenway was optimized based on the minimum cumulative model(MCR) and hydrological basin model. Result (1) Through the morphological spatial pattern analysis (MSPA), 30 ecological source patches were extracted to construct a comprehensive ecological resistance surface, and MCR model was used to generate 74 greenways with a total length of 918.11 km in Fuzhou, which, according to the importance evaluation of source patches, was divided into 4 levels: 9 pieces of 1st level totaling 132.88 km, 14 of 2nd level totaling 207.48 km, 14 of 3rd level totaling 153.57 km and 37 of 4th level totaling 424.18 km. (2) Through the hydrological basin model, 74 greenways were superimposed with the surface temperature of Fuzhou, and a total of 80 ecological ‘intersections’, 176 cooling ecological nodes and 35 cooling areas were extracted. Conclusion Accordingly, 14 new greenways are added in Fuzhou, including 8 road-based greenways and 6 waterfront greenways. The optimized greenways have a more comprehensive coverage of hot spots and a more obvious ecological regulation function for improving the urban heat island problem. [Ch, 2 fig. 4 tab. 25 ref.] -
图 1 基于水文流域模型的绿色廊道优化流程图
Figure 1 Greenway optimization flow chart based on hydrological basin model
图 2 绿色廊道降温生态节点和需降温区域分布
Figure 2 Distribution of cooling ecological nodes and cooling areas in greenway
表 1 福州市重要生态源地统计表
Table 1. Overview of important ecological sources in Fuzhou City
编号 面积/km² 区位 编号 面积/km² 区位 编号 面积/km² 区位 1 126.77 闽侯县、晋安区 11 3.24 长乐新区 21 2.96 长乐新区 2 1.98 晋安区 12 5.35 长乐新区 22 2.87 长乐新区 3 2.03 闽侯县、鼓楼区 13 1.74 仓山区 23 2.07 长乐新区 4 1.65 晋安区 14 3.42 长乐新区 24 20.97 长乐新区 5 13.65 马尾区 15 1.65 闽侯县 25 3.16 长乐新区 6 2.17 马尾区 16 1.90 长乐新区 26 1.62 长乐新区 7 1.56 马尾区 17 1.63 闽侯县 27 158.00 闽侯县 8 3.85 马尾区 18 73.07 闽侯县 28 31.08 长乐新区 9 17.68 闽侯县 19 2.53 长乐新区 29 5.86 长乐新区 10 140.47 马尾区 20 4.06 闽侯县 30 100.87 闽侯县  下载: 导出CSV
下载: 导出CSV
表 2 源斑块重要性分级统计
Table 2. Statistical graph of the importance classification to source plaques
斑块重要性 斑块等级 dIIC区间值 斑块数/个 面积/km2 比例/% 斑块编号 区位说明 最重要 一级 25.780~39.620 2 298.47 40.34 10、27 包括旗山和鼓山自然保护区、风景名胜区、国家森
林公园、重要水源及水库保护区等非常重要 二级 7.610~25.780 3 300.71 40.64 1、18、30 包括莲花山、五虎山和天台山自然林地、森林公
园、重要水源及水库保护区、大面积山地公
园、水源涵养保护区等很重要 三级 1.980~7.610 3 52.61 7.11 8、9、28 包括马尾琅岐旗山公园、闽侯六路山和长乐新区寨
顶山自然林地、大型山地公园、重要湿地等一般重要 四级 0.002~1.980 22 88.08 11.91 剩余斑块 包括大型山地公园、基本农田、大型水库、重要湿
地等
下载: 导出CSV
表 3 福州市绿色廊道分级概况
Table 3. Classification of greenway in Fuzhou City
廊道类型 数量/条 长度/km 绿色廊道路径 一级 9 132.87 ①斑块27(旗山国家森林公园)—斑块9(闽侯六路山);②斑块27—斑块15(闽侯官塘山);③斑块
27—斑块30(五虎山);④斑块10(鼓山风景名胜区)—斑块2(鼓岭风景名胜区);⑤斑块10—斑
块5(马尾琅岐南山尾);⑥斑块10—斑块8(马尾琅岐旗山);⑦斑块10—斑块13(城门山);
⑧斑块10—斑块17(南山公园);⑨斑块10—斑块18(天台山)二级 14 207.48 ①斑块1(莲花山)—斑块3(金鸡山公园);②斑块1—斑块4(鼓岭);③斑块1—斑块9(闽侯六路
山);④斑块18(天台山)—斑块6(牛岭山);⑤斑块18—斑块8(琅岐旗山);⑥斑块18—斑块
10(鼓山风景名胜区);⑦斑块18—斑块12(火焰山);⑧斑块18—斑块14(长乐烟台山);⑨斑块
18—斑块17(南山公园);⑩斑块18—斑块19(龙角峰公园);⑪斑块18—斑块23(长乐莲花山);
⑫斑块30(五虎山)—斑块15(闽侯官塘山);⑬斑块30—斑块20(祥谦林森公园);⑭斑块30—斑
块27(旗山国家森林公园)三级 14 153.57 ①斑块9(六路山)—斑块1(莲花山);②斑块9—斑块3(金鸡山公园);③斑块9—斑块15(闽侯官塘
山);④斑块9—斑块27(旗山国家森林公园);⑤斑块8(马尾琅岐旗山)—斑块5(马尾琅岐南山
尾);⑥斑块8—斑块6(牛岭山);⑦斑块8—斑块10(鼓山风景名胜区);⑧斑块8—斑块12(火焰
山);⑨斑块8—斑块18(天台山);⑩斑块28(寨顶山)—斑块17(南山公园);⑪斑块28—斑块
23(长乐莲花山);⑫斑块28—斑块24(塔山);⑬斑块28—斑块26(古槐镇农保地);⑭斑块28—
斑块29(三溪水库)四级 37 424.18 主要由城市大型山地公园、农保地、大型水库、河流湿地等四级斑块之间相互连接,构成建成
区的主要绿色廊道
下载: 导出CSV
表 4 福州市优化新增绿色廊道统计
Table 4. Statistical table of optimized new greenways in Fuzhou City
廊道类型 数量/条 廊道路径 道路型
绿色廊道8 ①五四北泰禾广场—福州北绕城高速—福山郊野公园—软件园—铜盘路—西湖公园—东街口—白马河—中
亭街—长安山公园—旧师大—高盖山公园—义序机场—三环路等
②闽侯六路山—国宾大道—永嘉天地—洪塘大桥—金牛山公园—梅峰路—西湖公园—华林路—牛岗山公园— 东三环路—盛丰物流园区—上洋工业区—鼓山等
③狮子峰—牛岗山公园—福光路—盛丰物流园区—上洋工业区—光明港公园等
④东街口—东大路—东二环泰禾广场等
⑤旗山国家森林公园—大学城—建平路—橘园洲大桥—金山大道—金山工业园区—金洲南路—飞凤山公园— 海峡奥林匹克体育中心—高盖山公园—白湖亭—南二环路—会展中心等
⑥浦上大桥—浦上大道—仓山万达—闽江大道—旧师大—长安山公园—世欧上江城等
⑦永嘉天地—旗山大道—大学城—高新区—南屿镇区—117县道—南港大桥—南通镇区等
⑧五虎山—奔驰大道—青口镇区—104国道—闽江绿道等、长乐洋屿工业园区—洞江—朝阳路—南山公园— 首占镇区—大象山滨水型
绿色廊道6 ①亭江镇区—马尾主城区—马尾工业园区—台江金融街万达—中亭街—闽侯荆溪永丰—甘蔗城区等沿闽江
北岸构成
②火车南站—会展中心—世欧上江城—闽江公园—淮安大桥—三环快速路乌龙江段—火车南站等形成的环
仓山区南台岛分布
③闽侯候官村—洪塘大桥—乌龙江大道—南屿镇区—203省道—福昆线—长乐洋屿工业园区—马尾琅岐镇区
等沿乌龙江及闽江构成
④长乐旗山—两港工业区—滨海新城—龙角峰公园等沿海岸线构成
⑤五四北泰禾广场—新店溪及秀峰路—火车北站—晋安河—世欧王庄—光明港—闽江等沿内河构成
⑥世欧王庄—光明港—上洋工业区—鼓岭等沿光明港构成
下载: 导出CSV
-
[1] 侍昊. 基于RS和GIS的城市绿地生态网络构建技术研究: 以扬州市为例[D]. 南京: 南京林业大学, 2010. SHI Hao. Study on City Greenspace Ecological Network Based on RS and GIS: A Case Study in Yangzhou[D]. Nanjing: Nanjing Forestry University, 2010. [2] FORMAN R T T, GODRON M. Landscpae Ecology[M]. New York: Wiley, 1986: 121 − 155. [3] 周年兴, 俞孔坚, 黄震方. 绿道及其研究进展[J]. 生态学报, 2006, 26(9): 3108 − 3115. ZHOU Nianxing, YU Kongjian, HUANG Zhenfang. Perspectives on greenway development [J]. Acta Ecol Sin, 2006, 26(9): 3108 − 3115. [4] 戴菲, 杨超, 徐亚如, 等. 基于POI点数据的武汉汉阳区绿道线路选择[J]. 中国城市林业, 2020, 18(6): 26 − 31. DAI Fei, YANG Chao, XU Yaru, et al. Greenway route selection in Hanyang district of Wuhan based on POI date [J]. J Chin Urban For, 2020, 18(6): 26 − 31. [5] 王春晓, 黄佳雯, 林广思. 基于选线适宜性评价的城镇型绿道规划方法研究[J]. 风景园林, 2020, 27(7): 108 − 113. WANG Chunxiao, HUANG Jiawen, LIN Guangsi. Research on urban greenway planning methods based on suitability evaluation [J]. Landscape Archit, 2020, 27(7): 108 − 113. [6] 赵晨晓, 刘春卉, 魏家星. 缓解城市热岛效应的南京市绿色基础设施网络构建方法[J]. 浙江农林大学学报, 2021, 38(6): 1127 − 1135. ZHAO Chenxiao, LIU Chunhui, WEI Jiaxing. Green infrastructure network construction method for mitigating urban heat island effect in Nanjing [J]. J Zhejiang A&F Univ, 2021, 38(6): 1127 − 1135. [7] 周媛. 多元目标导向下的成都中心城区绿地生态网络构建[J]. 浙江农林大学学报, 2019, 36(2): 359 − 365. ZHOU Yuan. Developing urban greenspace ecological network in Chengdu City center based on multiple objectives [J]. J Zhejiang A&F Univ, 2019, 36(2): 359 − 365. [8] 杨光旭. 缓解热岛效应的潜在生态网络分析及优化[D]. 北京: 中国地质大学, 2016. YANG Guangxu. Potential Ecological Network Analysis and Optimization for Relieving Heat Island Effect[D]. Beijing: China University of Geosciences, 2016. [9] 刘雅心, 顾阳, 胡一可. 环境公平视角下的社区绿道选线评价与导控策略[J]. 景观设计, 2021(3): 30 − 5. LIU Yaxin, GU Yang, HU Yike. Evaluation and control strategy of community greenway route selection from the perspective of environmental justice [J]. Landscape Des, 2021(3): 30 − 5. [10] 李磊, 宋伟. 园林绿地对改善城市人居环境的作用[J]. 天津科技, 2012, 39(4): 31 − 32. LI Lei, SONG Wei. Effect of garden green space on improving urban human settlements [J]. Tianjin Sci Technol, 2012, 39(4): 31 − 32. [11] 张爱因, 张晓丽. Landsat-8地表温度反演及其与MODIS温度产品的对比分析[J]. 北京林业大学学报, 2019, 41(3): 1 − 13. ZHANG Aiying, ZHANG Xiaoli. Land surface temperature retrieved from Landsat-8 and comparison with MODIS temperature product [J]. J Beijing For Univ, 2019, 41(3): 1 − 13. [12] SAURA S, ESTREGUIL C, MOUTON C, et al. Network analysis to assess landscape connectivity trends: application to European forests (1990−2000) [J]. Ecol Indic, 2011, 11(2): 407 − 416. [13] 崔凤奎, 王晓强, 张丰收, 等. 二值图像细化算法的比较与改进[J]. 洛阳工学院学报, 1997, 18(4): 50 − 54. CUI Fengkui, WANG Xiaoqiang, ZHANG Fengshou, et al. The comparision and betterment to thinning algorithm of two tone image [J]. Luoyang Inst Sci Technol, 1997, 18(4): 50 − 54. [14] VOGT P, RIITTERS K. Guidos Toolbox: universal digital image object analysis[J]. Eur J Remote Sensing, 2017, 2017, 50(1): 352-361. [15] GONZALEZ J R, BARRIO G D, DUGUY B. Assessing functional landscape connectivity for disturbance propagation on regional scales cost-: a cost-surface model approach applied to surface fire spread [J]. Ecol Modelling, 2008, 211(1/2): 121 − 141. [16] RUBIO S. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape RID A-9994-2008 [J]. Ecography, 2010, 33(3): 523 − 537. [17] 熊春妮, 魏虹, 兰明娟. 重庆市都市区绿地景观的连通性[J]. 生态学报, 2008, 28(5): 2237 − 2244. XIONG Chunni, WEI Hong, LAN Mingjuan. Analysis of connectivity on greenland landscape in metropolitan region of Chongqing City [J]. Acta Ecol Sin, 2008, 28(5): 2237 − 2244. [18] 吴金凤, 王秀红, 汪然. 中国东部沿海地区耕地和建设用地连通性变化: 以山东省平度市为例[J]. 水土保持通报, 2015, 35(5): 251 − 256. WU Jinfeng, WANG Xiuhong, WANG Ran. Connectivity change of cultivated land and construction land in eastern coastal China: a case study in Pingdu City in Shandong Province [J]. Bull Soil Water Conserv, 2015, 35(5): 251 − 256. [19] 张玮. 基于最小累积阻力模型的绿道选线研究: 以五台县绿道为例[D]. 太原: 山西农业大学, 2017. ZHANG Wei. Study of Greenway Site Selection Based on Minimum Cumulative Resistance Model: with Wutai Greenway for Example[D]. Taiyuan: Shanxi Agricultural University, 2017. [20] 许峰, 尹海伟, 孔繁花, 等. 基于MSPA与最小路径方法的巴中西部新城生态网络构建[J]. 生态学报, 2015, 35(19): 6425 − 6434. XU Feng, YIN Haiwei, KONG Fanhua, et al. Developing ecological networks based on MSPA and least-cost path method: a case study in Bazhong western new district [J]. Acta Ecol Sin, 2015, 35(19): 6425 − 6434. [21] 郭家新, 胡振琪, 李海霞, 等. 基于MCR模型的市域生态空间网络构建: 以唐山市为例[J]. 农业机械学报, 2021, 52(3): 275 − 284. GUO Jiaxin, HU Zhenqi, LI Haixia, et al. Construction of municipal ecological space network based on MCR model: a case study of Tangshan City [J]. Trans Chin Soc Agric Mach, 2021, 52(3): 275 − 284. [22] 徐文彬, 尹海伟, 孔繁花. 基于生态安全格局的南京都市区生态控制边界划定[J]. 生态学报, 2017, 37(12): 4019 − 4028. XU Wenbin, YIN Haiwei, KONG Fanhua. Development of ecological control boundaries in Nanjing metropoliis district based on ecological security patterns [J]. Acta Ecol Sin, 2017, 37(12): 4019 − 4028. [23] 张发明. 区间标度群组序关系评价法及其运用[J]. 系统工程理论与实践, 2013, 33(3): 720 − 725. ZHAN Faming. Interval scales rank correlation analysis group evaluation method and its application [J]. Syst Eng Theor Prac, 2013, 33(3): 720 − 725. [24] 陆明, 柳清. 基于Archydro水文分析模型的城市水生态网络识别研究: 以“海绵城市”试点济南市为例[J]. 城市发展研究, 2016, 23(8): 26 − 32. LU Ming, LIU Qing. Study on the identification of urban aquatic ecological network based on archydro hydrological model: taking “sponge city” Ji’nan as an example [J]. Urban Studies, 2016, 23(8): 26 − 32. [25] 王瑞燕, 赵庚星, 周伟, 等. 县域生态环境脆弱性评价及其动态分析: 以黄河三角洲垦利县为例[J]. 生态学报, 2009, 29(7): 3790 − 3799. WANG Ruiyan, ZHAO Gengxing, ZHOU Wei, et al. Assessment and dynamic changes of environment vulnerability at county level: a case study in Kenli County of the Yellow River Delta, China [J]. Acta Ecol Sin, 2009, 29(7): 3790 − 3799. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210672
 点击查看大图
点击查看大图

计量
- 文章访问数: 1878
- HTML全文浏览量: 378
- PDF下载量: 31
- 被引次数: 0



