




 下载:
下载:

-
机械损伤是植物生长过程中所面临最为普遍的胁迫之一[1]。植物不能通过躲避来免受机械损伤以及食叶昆虫和大型草食动物啃食的伤害。植物为了能够生存,在面对胁迫时通过启动体内防御反应信号系统应答外界的伤害,产生相应的防御反应[1],诱导植物体提高抵抗能力[2],从而获得系统免疫性,以提高植物的整体防御能力[3-4]。在植物受到损伤后,非结构性碳水化合物代谢会通过自身的改变来适应外界条件的变化。轻度刈割可以提高冷蒿Artemisia frigida可溶性碳水化合物含量和增加生物量[5],马尾松Pinus massoniana针叶被损伤后可溶性糖先降低,后逐渐恢复到原来水平[6]。冷蒿是菊科Asteraceae蒿属Artemisia呈半匍匐状或直立状生长的多年生小半灌木。已对冷蒿的生态学特性[7]、抗性生理特性[8-9]以及不同放牧条件下抗氧化防御系统[10]和次生代谢产物[11]等变化进行了研究。张汝民等[12]和左照江等[13]对冷蒿挥发物和化感作用等方面的研究,也更加明确了冷蒿在草原退化中的重要地位与作用。本研究以内蒙古草原主要建群植物冷蒿为对象,通过不同强度机械损伤处理冷蒿叶片和枝条(以剪刀损伤和揉捏方式),模拟牲畜对冷蒿枝叶的采食和践踏,从冷蒿体内非结构性碳水化合物代谢入手,研究机械损伤处理后冷蒿叶片和根部蔗糖、葡糖糖、果糖、淀粉质量分数的变化,探讨冷蒿体内非结构性碳水化合物代谢对机械损伤的响应机制。
-
供试材料冷蒿采自内蒙古自治区锡林浩特毛登牧场内蒙古大学草地生态学研究基地,地理位置为44°10′02′′ N,116°28′56′′ E,海拔1 160 m。全年平均气温为-0.4 ℃,1月最冷平均温度-22.3 ℃,7月最热平均气温18.8 ℃,≥0 ℃年积温为2 410.0 ℃,≥10 ℃积温为1 597.9°C,无霜期91.0 d,草原植物生长期为150 d左右。年降水量为365.6 mm,降水多集中在6-8月,占年降水量的80%左右。土壤为栗钙土。本研究区域主要草原植物为羊草Leymus chinensis,糙隐子草Cleistogenes squarrosa,克氏针茅Stipa krylovii,大针茅S. grandis,防风Saposhnikovia divaricata,冷蒿,瓣蕊唐松草Thalictrum petaloideum和阿尔泰狗哇花Heteropappus altaicus等。
-
2014年6月中旬在采样地选取生长健壮的冷蒿植株,栽植于盛有采样地原土的花盆中(直径18 cm,高20 cm),1丛·盆-1,苗高20 cm,15~20小枝·丛-1。盆栽苗置于浙江农林大学实验室温室中,自然光照,相对湿度为(30 ± 2)%,白天气温为(25.0 ± 2.0) ℃,夜晚温度为(20.0 ± 2.0) ℃。缓苗生长20 d后进行实验处理。选取株高一致、生长良好、无病虫害冷蒿20盆,随机分为4组,以剪刀损伤冷蒿叶片,损伤1/4叶片为轻度、损伤1/3叶片为中度、损伤1/2叶片为重度、不作处理为对照。3盆·处理-1,每盆为1个独立重复。分别在处理前(0 h),处理6,12和24 h后,对冷蒿叶片和根系分别取样,样品用液氮速冻后放置于-80 ℃低温冰箱内保存。
-
可溶性糖提取:称取植物样品0.2 g,液氮研磨匀浆,加蒸馏水5 mL,80 ℃恒温水浴30 min,不断搅拌,冷却,4 000 r·min-1离心5 min,沉淀用2 mL蒸馏水重复提取2次,合并上清液,定容至10 mL。用于可溶性糖测定。
淀粉水解:可溶性糖提取后的沉淀,加入2 mL蒸馏水,2 mL冷的6 mol·L-1盐酸,沸水浴30 min,4 000 r·min-1离心10 min,沉淀用3 mol·L-1盐酸 2 mL,沸水浴30 min重复提取2次;水浴时取出1滴提取液置白瓷板上,加1滴碘-碘化钾检查淀粉是否水解完全;合并上清液,冷却至室温加1滴酚酞,以6 mol·L-1氢氧化钠中和至溶液呈微红色。定容10 mL,用于淀粉质量分数测定。
-
葡萄糖、果糖和蔗糖使用葡萄糖试剂盒(上海荣盛生物药业有限公司生产)、果糖和蔗糖试剂盒(南京建成生物工程研究所南京建成科技有限公司生产)测定。具体方法按说明书操作。
-
参照参考文献[13]的方法,采用3,5-二硝基水杨酸比色法测定淀粉质量分数。
-
采用Origin 8软件(美国OriginLab公司)对实验数据进行统计分析和作图。统计方法采用One-Way ANOVA进行检验,并进行Tukey多重比较(P<0.05)。每个变量数值表示为平均值+标准误差。采用独立样本t检验进行冷蒿组织和机械损伤与对照组之间的差异。
-
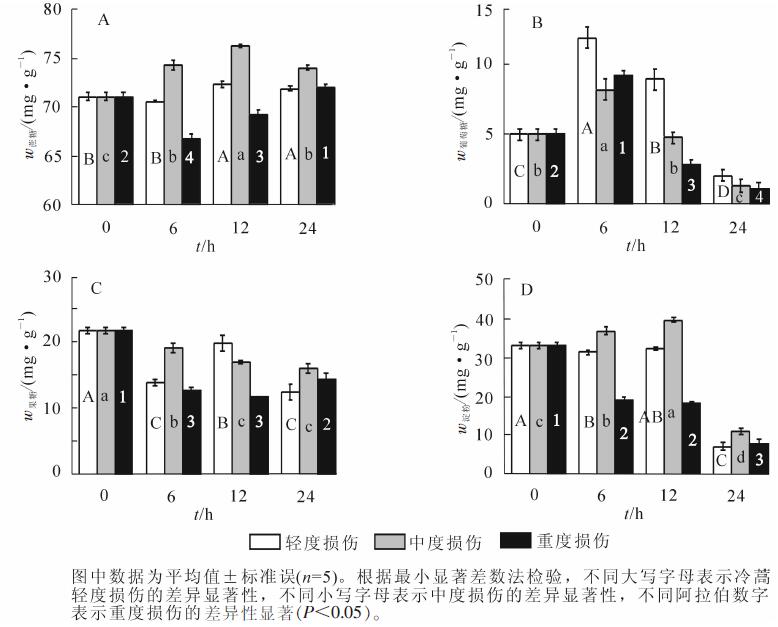
从图 1A中可以看出,机械损伤程度为轻度、中度、重度处理后,在24 h内冷蒿叶片蔗糖质量分数变化有明显差别。在轻度机械损伤后,叶片蔗糖随着处理时间的延长而增加,与对照相比,12和24 h分别增加了1.8%和1.1%(P<0.05);在中度机械损伤后,冷蒿叶片内蔗糖随着处理时间的延长继续增加,6,12和24 h分别比对照增加了4.5%,7.3%和4.1%(P<0.05),表明此刻冷蒿叶片合成蔗糖的能力较强;经过重度损伤后,冷蒿叶片蔗糖质量分数开始下降,6和12 h分别比对照降低了6.1%和2.6%(P<0.05),到24 h时,合成能力恢复,比对照增加了1.4%。冷蒿经过轻度、中度和重度损伤后,在6和12 h时,中度处理后蔗糖质量分数显著高于重度损伤,到24 h,三者的质量分数基本相同,说明蔗糖的合成能力在损伤冷蒿体内较强。

图 1 机械损伤对冷蒿叶片非结构性碳质量分数的影响
Figure 1. Effects of mechanical damage on non-structural carbohydrate content in leaves of Artemisia figida
-
从图 1B中可以看出:分别进行轻度、中度、重度机械损伤处理的冷蒿,6 h葡萄糖质量分数较高,分别比对照增加了140.0%,64.8%和84.9%(P<0.05);到12 h,轻度处理葡萄糖质量分数增加了80.0%,中度和重度处理葡萄糖质量分数下降了5.1%和44.5%(P<0.05);到24 h,葡萄糖质量分数分别比对照下降了60.0%,74.6%和80.1%(P<0.05)。冷蒿经过损伤处理后,6 h可显著增加体内葡萄糖质量分数,延长到24 h,体内葡萄糖显著下降,轻度、中度和重度处理后比6 h下降了83.3%,84.9%和89.2%(P<0.05)。
-
从图 1C中可以看出:冷蒿叶片经过轻度、中度和重度损伤处理后,果糖质量分数都发生下降,中度处理后,下降的程度最小。
-
从图 1D中可以看出:轻度处理冷蒿后,6 h淀粉质量分数开始下降,24 h比对照降低了78.9%(P<0.05);中度处理后,淀粉开始上升,6和12 h分别比对照上升了11.4%和20.2%(P<0.05),24 h比对照下降了67.1%(P<0.05);重度处理后,淀粉质量分数开始显著下降,分别比对照下降了43.4%,45.2%和77.2%(P<0.05)。经过损伤处理后,到6 h,轻度、中度和重度处理之间,淀粉质量分数差异显著;到12 h,轻度、中度和重度处理之间,淀粉质量分数都有显著差异变化。
-
从图 2A中可以看出:在轻度机械损伤后,根部蔗糖质量分数在6 h比对照下降了5.0%(P<0.05),到12和24 h与对照相比分别增加了6.9%(P<0.05)和0.7%,表明轻度损伤处理对冷蒿根部蔗糖合成的影响较小;在中度机械损伤处理后,冷蒿根部蔗糖质量分数随着处理时间的延长而明显下降,6,12和24 h分别比对照下降了37.4%,14.4%和33.9%(P<0.05);经过重度损伤后,冷蒿根部蔗糖质量分数6,12和24 h分别比对照下降了52.7%,50.6%和33.2%(P<0.05)。冷蒿根部经过轻度、中度和重度损伤后,在6和12 h,轻度处理的蔗糖质量分数明显高于中度损伤,而中度损伤处理的蔗糖质量分数显著高于重度损伤;到24 h后,轻度损伤处理的蔗糖含量仍高于中度和重度损伤。

图 2 机械损伤对冷蒿根系非结构性碳质量分数的影响
Figure 2. Effects of mechanical damage on non-structural carbohydrate content in roots of Artemisia frigida
-
从图 2B中可以看出:6 h葡萄糖质量分数较高,分别比对照增加了62.9%,94.3%和34.3%(P<0.05);到12 h,轻度损伤处理冷蒿根部葡萄糖质量分数回到对照水平,中度和重度处理葡萄糖质量分数下降了17.1%和28.6%(P<0.05);到24 h,轻度和重度处理葡萄糖质量分数比对照下降了2.9%和8.6%,而中度处理的葡萄糖质量分数与对照相比增加了2.9%。冷蒿经过损伤处理后,6 h根部葡萄糖质量分数显著增加,到24 h后,三者的合成能力基本相同且与对照相接近。
-
从图 2C中可以看出:6和12 h轻度损伤处理后的冷蒿根部果糖质量分数较高,分别比对照增加了28.5%和41.0%(P<0.05),到24 h,轻度处理果糖质量分数增加了8.2%(P<0.05);中度损伤处理的冷蒿根部果糖质量分数在6,12 h与对照相比,下降了1.0%和4.8%,到24 h果糖质量分数比对照增加了8.2%(P<0.05);经过重度损伤后,6,12和24 h冷蒿根部果糖质量分数比对照下降了0.4%,13.5%和1.7%。轻度处理后,根部果糖质量分数增加最多。
-
从图 2D中可以看出:冷蒿轻度处理后,6 h根部淀粉质量分数开始下降,则到12和24 h淀粉质量分数上升,比对照增加了2.5%和5.3%(P<0.05);中度处理后,根部淀粉质量分数开始上升,6和12 h分别比对照上升了1.4%和4.9%,24 h比对照下降了12.7%(P<0.05);重度处理后,淀粉质量分数开始下降,分别比对照下降了2.3%,3.9%和21.9%(P<0.05)。经过损伤处理后,在6和12 h,中度处理淀粉质量分数较高,高于轻度和重度损伤处理;到24 h,轻度、中度和重度处理之间,根部淀粉质量分数差异显著。
-
糖是生物大分子物质。糖类是光合作用的产物,又是呼吸作用的底物,它为植物的生长发育提供碳骨架和能量,并能增强植物抗逆性。糖代谢是植物的基础代谢之一,它的中间产物能为蛋白质代谢、脂类代谢、核酸代谢及次生代谢提供原料。
植物体内的糖代谢属于基础代谢,各种可溶性糖和储藏的淀粉处于不断合成与分解的动态变化中,这是正常的生理代谢过程。植物光合产物主要是糖类,包括:葡萄糖、果糖、蔗糖和淀粉,糖类是光合作用的唯一直接产物。从研究结果图 1可看出:冷蒿叶片通过光合作用在体内积累的糖主要是蔗糖和淀粉,其质量分数较高;而葡萄糖和果糖合成了蔗糖,导致其在体内积累量较少,特别是葡萄糖,它既是合成淀粉的供体,又是合成蔗糖的供体。
当植物受到损伤后,特别是进行光合作用的叶片被损伤,其光合能力下降,形成的糖类质量分数发生变化,有些糖类将转化成抵抗损伤的应急产物。在损伤初期,6和12 h冷蒿受到轻度和中度机械损伤后,体内蔗糖质量分数较高,在12 h与对照相比轻度和中度处理蔗糖质量分数分别增加了1.8%和7.3%,可能是葡萄糖和果糖结合形成。从图 1可见:葡萄糖和果糖质量分数这时较高,在6 h轻度、中度和重度机械损伤处理后葡萄糖质量分数分别比对照增加了140.0%,64.8%和84.9%,说明冷蒿有能力继续合成蔗糖,在体内进行积累。这时轻度和中度处理的冷蒿叶片淀粉质量分数变化不大,说明淀粉的合成与分解未受到损伤的影响,处于动态平衡过程,而重度机械损伤后的淀粉质量分数与对照相比下降67.1%,说明淀粉进行分解反应,形成葡萄糖。到损伤后期,图 1D表明,3种损伤处理的淀粉质量分数都出现明显下降,说明淀粉进行分解形成葡萄糖。此时,葡萄糖一方面与果糖结合,形成蔗糖,导致蔗糖质量分数保持在较高水平,进行应激反应;另一方面葡萄糖参与应激反应,可能转化成其他中间代谢产物,为抵抗损伤提供应激物质和能量,因为葡萄糖分子在分解氧化过程中可以释放大量的能量。从图 1可说明:冷蒿受到损伤后,体内淀粉、蔗糖、葡萄糖质量分数发生变化,非结构性碳水化合物进行转化,参加应激反应,作为能量和物质的提供者。这些物质不是持续无控的而是快速间断和被调节的,反映了冷蒿的应激反应过程是一种时态的变化。受害程度的强弱和时间长短对冷蒿非结构碳水化合物代谢的变化有一定的影响。 王燕等[15]研究了马尾松Pinus massoniana受到不同程度的损伤后,其体内可溶性糖发生变化后逐渐恢复到原来水平。李镇宇等[16]研究发现:受赤松毛虫Dendrolimus spectabilis危害的油松松针内粗脂肪、单宁、生物碱含量增加,可溶性糖含量下降,其抗虫性增强;而油松Pinus tabulaeformis新长出的1年生针叶,水溶性总糖、粗脂肪、生物碱含量及总糖/氨基酸比例上升[17];当小油松受害后2年生针叶内单宁、生物碱含量增加,水溶性糖、还原糖、糖/氨基酸比例下降[18]。
植物被刈割后,叶面积大幅度降低,其净光合速率也下降,由于地上部分的营养体减少,地下资源过剩促使植物产生更多的生活物质,补充形成应激反应所要的原料和能量[19]。因此,通过增加可溶性碳水化合物进行调节,DONAGHY等[20]在研究多年生黑麦草Lolium perenne刈割后发现,刈割损伤可以增加植物体内的非结构性碳化合物含量。王静等[5]对不同强度刈割后的冷蒿体内碳水化合物进行研究,发现冷蒿经过适度的刈割干扰,有利于体内碳水化合物和生物量的积累,其生长的可利用资源空间越多,可溶性碳水化合物含量增高。本研究表明:轻度和中度损伤冷蒿后,初期体内蔗糖、葡萄糖和淀粉质量分数上升,到后期葡萄糖和淀粉质量分数下降,可能它们形成了应激反应所需要的原料和能量,与前人的研究结果一致。
蔗糖是碳水化合物的主要运输形式,在植物叶片形成后,可直接运输到根部,为根生长、有机物代谢提供碳源和能量。从图 2可知:轻度损伤后,根内蔗糖质量分数较高,可能一部分来自葡萄糖和果糖结合而成,另一部分是由地上部分运输而来。经过重度损伤后,由于体内缺乏能量,葡萄糖和果糖未能合成蔗糖,使其质量分数不能升高。淀粉是根内积累的碳水化合物,冷蒿经过不同程度的损伤后,对其质量分数影响较小,但在24 h重度损伤后,其质量分数明显下降,可能是进行分解,形成葡萄糖为根系维持基础代谢提供能量。从根系结果也可看出,蔗糖和淀粉是冷蒿根内积累的2种主要非结构性碳水化合物。
Responses of non-structural carbohydrate metabolism to mechanical damage in Artemisia frigida
-
摘要: 为了探讨放牧过程牲畜采食和践踏损伤对冷蒿Artemisia frigida体内非结构性碳水化合物代谢的影响,对盆栽冷蒿枝叶采用不同程度(轻度、中度、重度)机械损伤的方式模拟放牧,测定了冷蒿叶片和根部蔗糖、葡萄糖、果糖和淀粉质量分数的变化。结果表明:在损伤初期,轻度和中度处理后冷蒿叶片中蔗糖、葡萄糖和淀粉质量分数显著增加(P<0.05);到损伤24 h,3种处理中蔗糖质量分数基本恢复到对照水平,而葡萄糖和淀粉质量分数显著下降,与对照相比,葡萄糖质量分数降了60.0%,74.6%和80.1%,淀粉质量分数下降了43.4%,45.2%和77.2%。葡萄糖和果糖结合成蔗糖,使其质量分数较高,在冷蒿体内积累。冷蒿根部非结构性碳水化合物质量分数变化与叶片相比是有所不同的。损伤初期,冷蒿根部葡萄糖质量分数上升,3种处理与对照相比分别增加了62.9%,94.3%和34.3%,果糖在轻度机械损伤处理后明显上升;到损伤后期,葡萄糖和果糖恢复到对照水平,蔗糖和淀粉随着损伤加强而下降。根部积累的主要是蔗糖和淀粉。冷蒿受损伤后,体内淀粉、蔗糖、葡萄糖质量分数发生变化,参加应急反应,同时轻度损伤可以增加冷蒿体内非结构性碳水化合物。Abstract: To determine the effect of grazing and trampling damage of livestock on the metabolism of non-structural carbohydrates in Artemisia frigida, mechanical damage at different degrees (light, moderate, and severe) was applied to simulate grazing. The content of starch, fructose, glucose, and sucrose in leaves and roots were determined. Results showed that in the early stage with light and moderate mechanical damage, sucrose, glucose, and starch content of A. frigida leaves increased significantly (P<0.05). Sucrose content under all the three treatments restored to the level of the control in 24 h; whereas glucose decreased by 60.0%, 74.6%, and 80.1%(P<0.05), respectively, under light, moderate, and high treatments, whereas starch content decreased by 43.4%, 45.2%, and 77.2% (P<0.05), respectively, under light, moderate, and heavy treatments. The variation pattern of non-structural carbohydrates in roots differed that in leaves. At early stage glucose content increased, by 62.9%, 94.3%, and 34.3% (P<0.05), respectively, under light, moderate, and heavy treatments. Fructose content increased significantly at early stage of light treatment. At late stages of three treatments, glucose and fructose contents were restored to the control level; however, sucrose and starch contents decreased with increasing damage. Sucrose and starch were accumulated mainly in roots. Thus, after treatments, the content of starch, sucrose, and glucose in A. frigida changed in response to emergency while mild damage increased the content of non-structural carbohydrates.
-
Key words:
- botany /
- Artemisia frigida /
- mechanical damage /
- non-structural carbohydrate
-
图 1 机械损伤对冷蒿叶片非结构性碳质量分数的影响
Figure 1 Effects of mechanical damage on non-structural carbohydrate content in leaves of Artemisia figida
-
[1] 宋恒,王长泉. 机械伤害诱导的植物防御机制和信号转导[J]. 植物学报, 2013, 48(4):461-469. SONG Heng, WANG Changquan. Mechanism of wound-induced defense response and signal transduction in plants[J]. Chin Bull Bot, 2013, 48(4):461-469. [2] LEÓN J, ROJO E, SÁNCHEZ-SERRANO J J. Wound signalling in Plants[J]. J Exp Bot, 2001, 52(354):1-9. [3] DOMBROWSKI J E, HIND S R, MARTIN R C, et al. Wounding systemically activates a mitogenactivated protein kinase in forage and turf grasses[J]. Plant Sci, 2011, 180(5):686-693. [4] 冯远娇,金琼,王建武. 机械损伤对Bt玉米化学防御的系统诱导效应[J]. 植物生态学报,2010,34(6):695-703. FENG Yuanjiao, JIN Qiong, WANG Jianwu. Systemic induced effects of mechanical wounding on the chemical defense of Bt corn (Zea mays)[J]. Chin J Plant Ecol, 2010, 34(6):695-703. [5] 王静,杨持,韩文权,等. 刈割强度对冷蒿可溶性碳水化合物的影响[J]. 生态学报,2003,23(5):908-913. WANG Jing, YANG Chi, HAN Wenquan, et al. Effects on water-soluble carbohydrate of Artemisia frigida under different defoliation intensities[J]. Acta Ecol Sin, 2003, 23(5):908-913. [6] 王燕,戈峰, 李镇宇. 马尾松诱导化学物质变化的时空动态[J]. 生态学报,2001,21(8):1256-1261. WANG Yan, GE Feng, LI Zhenyu. Spatial-tempetial trends of induced chemical change in pine Pinus massoniana[J]. Acta Ecol Sin, 2001, 21(8):1256-1261. [7] 占布拉,陈世璜,张昊,等. 冷蒿的特性和生态地理分布的研究[J]. 内蒙古农牧学院学报,1999,20(1):1-7. Zhanbula, CHEN Shihuang, ZHANG Hao, et al. The characteristics ecological and geographical distribution of Artemisia feigida[J]. J Inner Mongolia Inst Agric Anim Husb, 1999, 20(1):1-7. [8] 吴建华,张汝民,高岩.干旱胁迫对冷蒿保护酶活性及膜脂过氧化作用的影响[J]. 浙江林学院学报,2010,27(3):329-333. WU Jianhua, ZHANG Rumin, GAO Yan. Membrane lipid peroxidation and protective enzyme systems with drought stressed Artemisia frigida leaves[J]. J Zhejiang For Coll, 2010, 27(3):329-333. [9] 周海燕. 水分胁迫对冷蒿和差不嘎蒿体内水分状况的影响[J]. 中国沙漠,1999,19(增刊1):26-29. ZHOU Haiyan. Water state of Artemisia Feigida and Artemisia halodendron under fifferent water stress conditions[J]. J Desert Res, 1999, 19(supp 1):26-29. [10] 贾丽,刘盟盟,张洪芹,等. 冷蒿抗氧化防御系统对机械损伤的响应[J]. 浙江农林大学学报,2016,33(3):462-470. JIA Li, LIU Mengmeng, ZHANG Hongqin, et al. Antioxidant defense system responses of Artemisia frigida to mechanical damage[J]. J Zhejiang A & F Univ, 2016, 33(3):462-470. [11] 刘盟盟,贾丽,张洪芹,等. 机械损伤对冷蒿叶片次生代谢产物的影响[J]. 浙江农林大学学报,2015,32(6):845-852. LIU Mengmeng, JIA Li, ZHANG Hongqin, et al. Mechanical damage on secondary metabolites from Artemisia frigida leaves[J]. J Zhejiang A & F Univ, 2015, 32(6):845-852. [12] 张汝民, 王玉芝, 侯平, 等. 几种牧草幼苗对冷蒿茎叶水浸提液的化感作用的生理响应[J]. 生态学报, 2010, 30(8):2197-2220. ZHANG Rumin,WANG Yuzhi,HOU Ping,et al. Physiological responses to allelopathy of aquatic stem and leaf extract of Artemisia frigida in seedling of several pasture plants[J]. Acta Ecol Sin, 2010, 30(8):2197-2220. [13] 左照江,张汝民,王勇,等. 损伤冷蒿挥发性有机化合物(VOCs)成分分析及其对牧草根系生长发育的影响[J]. 生态学报,2010,30(19):5131-5139. ZUO ZhaoJiang, ZHANG RuMin, WANG Yong, et al. The composition of volatile organic compounds (VOCs) emitted from damaged Artemisia frigida Willd. plants and their effects on root growth and development of pasture plants[J]. Acta Ecol Sin, 2010, 30(19):5131-5139. [14] 张治安,张美善,蔚荣海. 植物生理学实验指导[M]. 北京:中国农业科学技术出版社,2004. [15] 王燕,李镇宇,戈峰. 马尾松受害诱导的化学物质滞后变化[J]. 昆虫学报,2000,43(3):291-296. WANG Yan, LI Zhenyu, GE Feng. Lag-change of chemical components in needles of injured pine, Pinus massoniana[J]. Acta Entomol Sin, 2000, 43(3):291-296. [16] 李镇宇,陈华盛,袁小环,等. 油松对赤松毛虫的诱导化学防御[J]. 林业科学,1998,34(2):43-50. LI Zhenyu, CHEN Huasheng, YUAN Xiaohuan, et al. Induced chemical defenses of Pinus tabulaeformis Carr. to Dendrolmus spectabilis Butler[J]. Sci Silv Sin, 1998, 34(2):43-50. [17] 李镇宇,王燕,陈华盛,等. 油松对赤松毛虫的诱导化学防御及滞后诱导抗性[J]. 林业科学,2000,36(1):66-70. LI Zhenyu, WANG Yan, CHEN Huasheng, et al. Induced chemical defenses and delayed induced resistance of Pinus tabulaeformis Carr. to Dendrolimus spectabilis Butler[J]. Sci Silv Sin, 2000, 36(1):66-70. [18] 李镇宇,王燕,陈华盛,等. 赤松毛虫的危害对小油松针叶内物质含量的影响[J]. 北京林业大学学报,1999,21(5):41-45. LI Zhenyu, WANG Yan, CHEN Huasheng, et al. Affection of contents of some substances in needle of Pinus tabulaeformis Carr. induced by Dendrolimus spectabilis Butler[J]. J Beijing For Univ, 1999, 21(5):41-45. [19] 张鹏翀,胡增辉,沈应柏,等. 不同损伤对合作杨光合特性的影响[J]. 北京林业大学学报,2010,32(1):46-52. ZHANG Pengchong, HU Zenghui, SHEN Yingbai, et al. Effects of three types of wound on photosynthetic activity of Populus simonii×P. pyramidalis ‘Opera 8277’ seedlings[J]. J Beijing For Univ, 2010, 32(1):46-52. [20] DONAGHY D J, FULKERSON W J. Priority for allocation of water-soluble carbohydrate reserves during regrowth of Lolium perenne[J]. Grass Forage Sci, 1998, 53(3):211-218. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2016.04.011
 点击查看大图
点击查看大图
图(2)
计量
- 文章访问数: 3196
- HTML全文浏览量: 620
- PDF下载量: 512
- 被引次数: 0
