




-
过去20 a间,全球气候发生了巨大变化,对生态系统的结构和功能产生了深远影响[1]。气候改变会导致植物体内非结构性碳水化合物(NSC)的变化。有研究表明:气候变暖显著降低了陆地植被的NSC、可溶性糖和淀粉质量分数[2]。NSC主要由可溶性糖和淀粉组成,参与植物细胞在逆境下的渗透调节过程[3]。NSC在植物体内的累积量一定程度上可以反映植物体内碳的供需平衡关系,对维持植物体渗透调节、水力传输和生长发育,缓冲树木在年际、季节和器官间碳供需关系至关重要[4]。NSC质量分数是反映植物生存策略的重要指标[5−7],在改善植物对干旱环境的适应性、保持植物不利条件下的生长和提高植物存活率方面发挥着关键作用[8]。
降水格局变化是全球气候变化的研究热点[9],深刻影响着区域干旱[10−11]、地表径流[12−14]、河流水量[14−15]等,同时也间接作用于生态系统的弹性与安全[16]。降水是植物获取水分的重要来源,是植物生长的关键因素[17]。降水量减少会导致树木死亡,严重损害森林生态系统的固碳能力[18−19]。目前,有关降水格局变化对植物NSC的影响仅在少数植物,如红砂Reaumuria soongarica[20]和马尾松Pinus massoniana[21]等上有研究。其中,红砂幼苗NSC质量分数在干旱胁迫下随胁迫时间的增加而增加[20],持续隔离降水导致马尾松针叶NSC质量分数先显著增加后减少[21]。这说明不同树种在面临干旱条件时可能会采取不同方式调节各器官的NSC质量分数来适应干旱,但是目前对南方造林面积最大的树种杉木Cunninghamia lanceolata鲜有报道。因此,研究中国亚热带地区降水格局变化下先锋树种杉木的生理生态特性,对预测未来气候条件下基于碳水化合物调节的杉木适应降水变化的生理机制极具价值。
杉木作为一种重要的速生针叶木材种类,以其迅速的生长速度、笔直的干形和上乘的材料而著称[22],在中国人工林木材资源中占比很大[23−24]。但是,季节性干旱事件的频繁发生,对杉木人工林生产能力带来的负面影响逐渐加剧。杉木的树高与胸径比值较高,使其更容易受到极端气候的影响[25]。然而关于杉木幼苗不同器官NSC及其相关组分对降水格局变化的响应策略以及在降水格局变化时杉木人工林应制定的管理策略尚不清楚。本研究通过分析杉木幼苗不同器官NSC及其相关组分(可溶性糖、淀粉)在不同降水量和不同降水间隔的变化与分配情况,旨在全面和深入掌握杉木生长季中不同器官NSC对不同降水格局变化的响应,为亚热带地区杉木人工林的科学抚育管理提供理论指导和科学依据。
-
研究区位于江西省南昌市(28°10′~29°11′N,115°27′~116°35′E),属亚热带湿润季风气候,极高温为40.9 ℃,极低温为−15.2 ℃,年平均气温为17.0 ℃,年平均降水量为1 600.0~1 700.0 mm,年平均相对湿度为78.5%。森林覆盖率为21.3%,主要植被类型为常绿阔叶林,主要土壤类型为红壤[26]。
-
试验开展于江西农业大学中药园。2021年4月,将2年生杉木幼苗置于40 cm×40 cm×50 cm的长方体花盆中开始缓苗,8月开始控制试验,7:00—8:00对幼苗进行降水,其余生态因子均保持一致,2022年1月结束试验。杉木幼苗胸径为1.5 cm,树高为1.0~1.3 m。以南昌1955—2018年旱季的月平均降水量(69.12 mm)为对照组(ck),分别设置减少30%、50%、80%降水量的处理组(W-30%、W-50%、W-80%)。在此基础上,参考南昌1955—2018年实测降水间隔数据,设置降水间隔5和10 d (T5和T10) 2个处理。因此,降水格局共8个处理组,每处理5次重复,共40盆。
-
样品收集过程中,先从盆里完整地取出杉木幼苗,再用保鲜膜包裹根系,确保根系完整性的同时防止水分流失,并迅速将样品带回实验室。将杉木幼苗分为叶、枝条、枝干、运输根、吸收根5个部分,因幼苗较小,枝干为地上部分最粗壮的主干,枝条为除去主干后的其余地上分支[27]。放入105 ℃的烘箱中烘烤30 min,然后在60 ℃下烘干至恒量,研磨粉碎过0.25 mm筛,测定NSC及其相关组分质量分数。可溶性糖质量分数采用蒽酮比色法测定,淀粉质量分数采用稀酸水解法测定,NSC质量分数为可溶性糖和淀粉质量分数的总和[28−29]。其中,根系按照欧阳园丽等[30]的根序级别划分方式分级。
-
所有数值以平均值±标准差表示。数据分析和绘图采用R 4.3.0进行。采用线性混合模型分析降水格局对杉木幼苗不同器官NSC及其相关组分的影响;单因素方差(one-way ANOVA)分析不同降水量同一降水间隔不同器官NSC及其相关组分的差异。
-
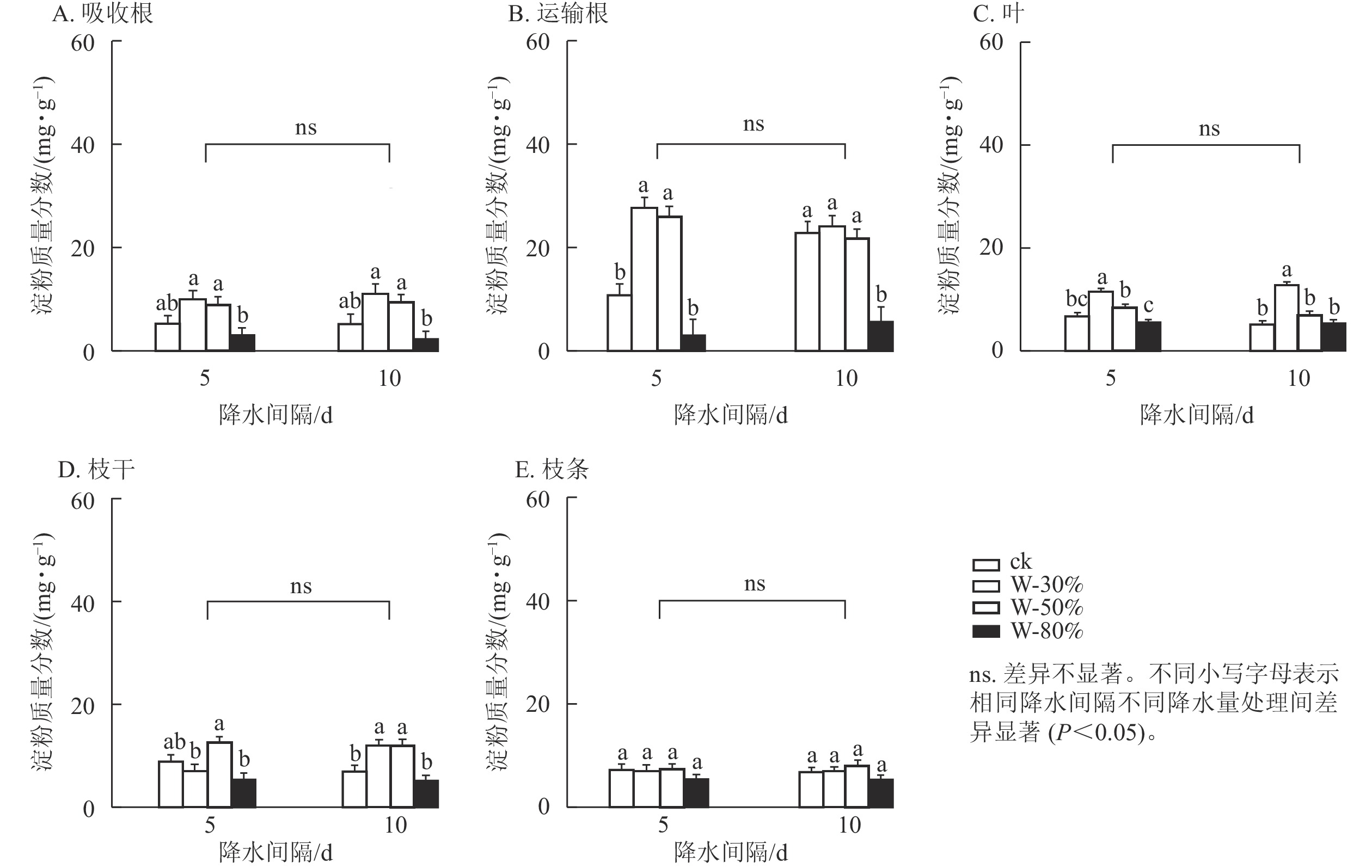
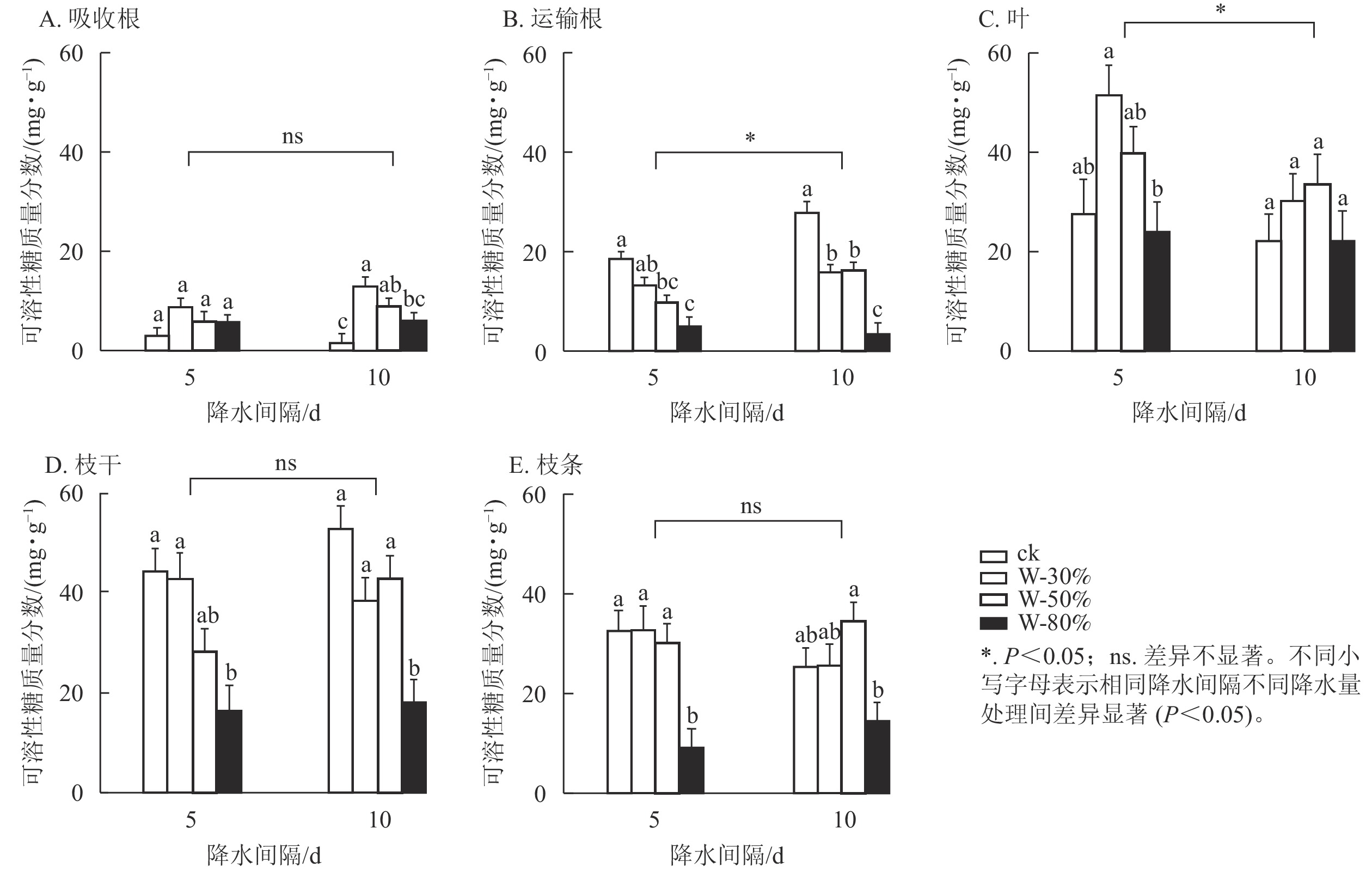
降水量对杉木幼苗不同器官可溶性糖质量分数具有极显著影响(P<0.01),降水间隔则整体无显著影响(表1)。由图1可知:降水间隔对运输根和叶的可溶性糖质量分数有显著影响(P<0.05),对吸收根、枝干和枝条无显著影响。T5处理下,随着降水量的减少,可溶性糖质量分数在吸收根、叶和枝条中呈先增加后减少的趋势,其中,枝条可溶性糖质量分数在W-80%处理下最小,显著低于ck (P<0.05),比ck降低了71.9%;可溶性糖质量分数在运输根和枝干中表现为持续减少,均在W-80%处理下最小,显著低于ck (P<0.05),分别降低了72.8%和63.1%。T10处理下,随着降水量减少,可溶性糖质量分数在吸收根、叶和枝条中呈先增加后减少的趋势,其中,吸收根的可溶性糖质量分数在W-30%处理下最大,显著高于ck (P<0.05),为ck的9.6倍;在运输根和枝干中可溶性糖质量分数表现为先减少后增加再减少,均在W-80%处理下最小,显著低于ck (P<0.05),分别降低了87.9%和65.8%。
表 1 降水格局对杉木幼苗不同器官可溶性糖、淀粉、NSC质量分数影响的线性混合模型分析
Table 1. Effect of precipitation patterns on soluble sugar, starch and NSC content of different organs of C. lanceolata seedlings based on analysis of linear mixed-effects models
影响因素 可溶性糖 淀粉 NSC F P F P F P W 29.06 <0.001 87.72 <0.001 129.55 <0.001 T 0.98 0.320 0.20 0.660 2.73 0.100 G 66.30 <0.001 119.74 <0.001 98.46 <0.001 W×T 2.56 <0.100 2.18 <0.100 9.28 <0.001 W×G 5.42 <0.001 11.79 <0.001 20.13 <0.001 T×G 3.83 <0.010 0.38 0.820 3.98 <0.010 W×T×G 0.96 0.490 4.57 <0.001 4.03 <0.001 说明:W. 不同降水量处理;T. 不同降水间隔处理;G. 杉木幼苗不同器官;×表示交互作用。P<0.01表示差异极显著。 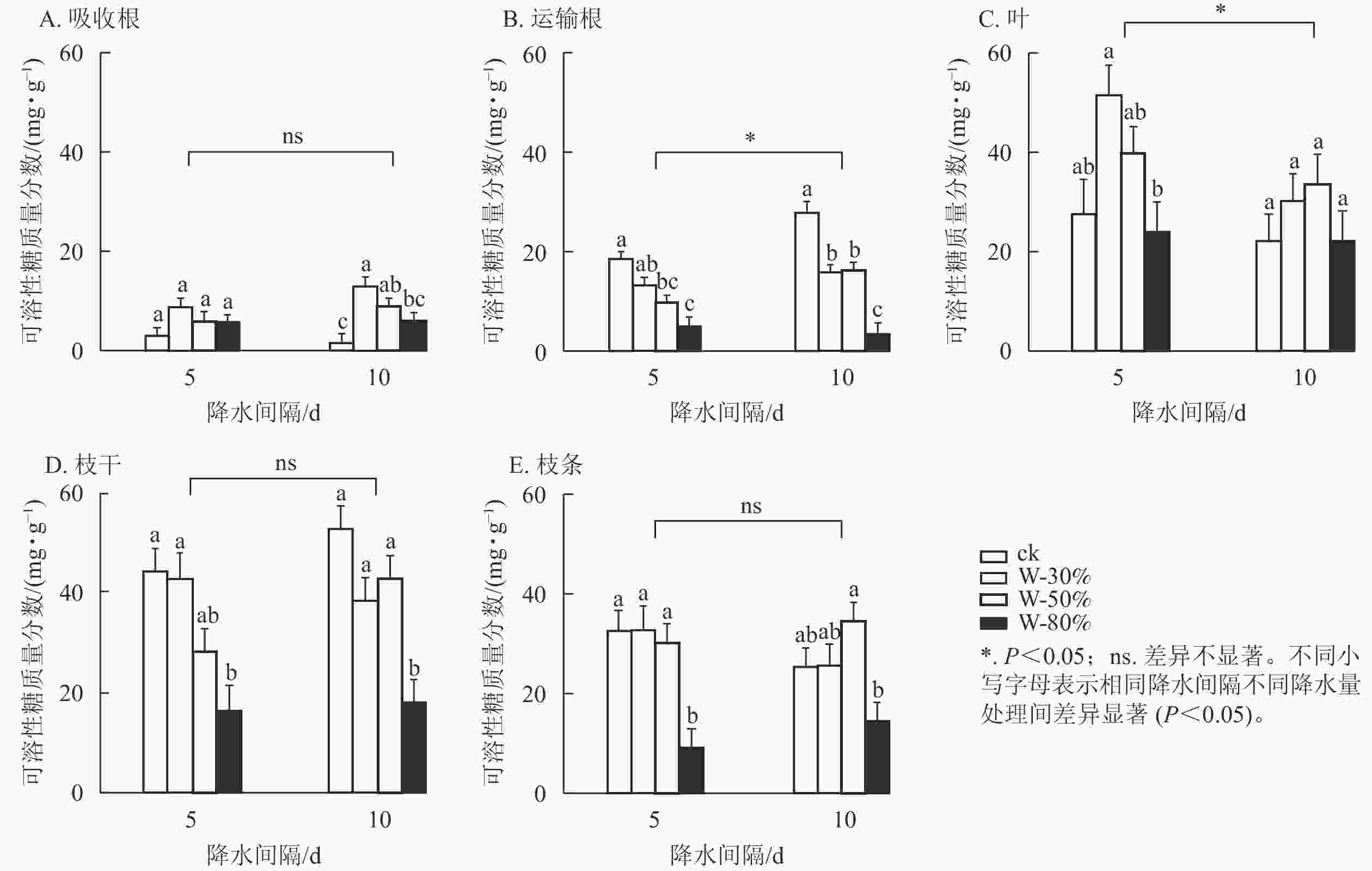
图 1 不同降水量和降水间隔对杉木幼苗各器官可溶性糖质量分数的影响
Figure 1. Effects of different precipitation amount and intervals on soluble sugar content in different organs of C. lanceolata seedlings
-
降水量对杉木幼苗不同器官淀粉质量分数具有极显著影响(P<0.01),降水间隔则整体无显著影响(表1)。由图2可知:T5处理下,随着降水量的减少,淀粉质量分数在吸收根、运输根和叶中呈先增加后减少的趋势,其中,运输根和叶的淀粉质量分数在W-30%处理下最大,均显著高于ck (P<0.05),分别为ck的2.6和1.9倍。T10处理下,随着降水量的减少,淀粉质量分数在吸收根、运输根、叶、枝干和枝条中均呈先增加后减少的趋势,其中,运输根的淀粉质量分数在W-80%处理下最小,显著低于ck (P<0.05),比ck降低了78.0%;叶的淀粉质量分数在W-30%处理下最大,显著高于ck (P<0.05),为ck的2.5倍;枝干的淀粉质量分数在W-50%处理下最大,显著高于ck (P<0.05),为ck的1.8倍。
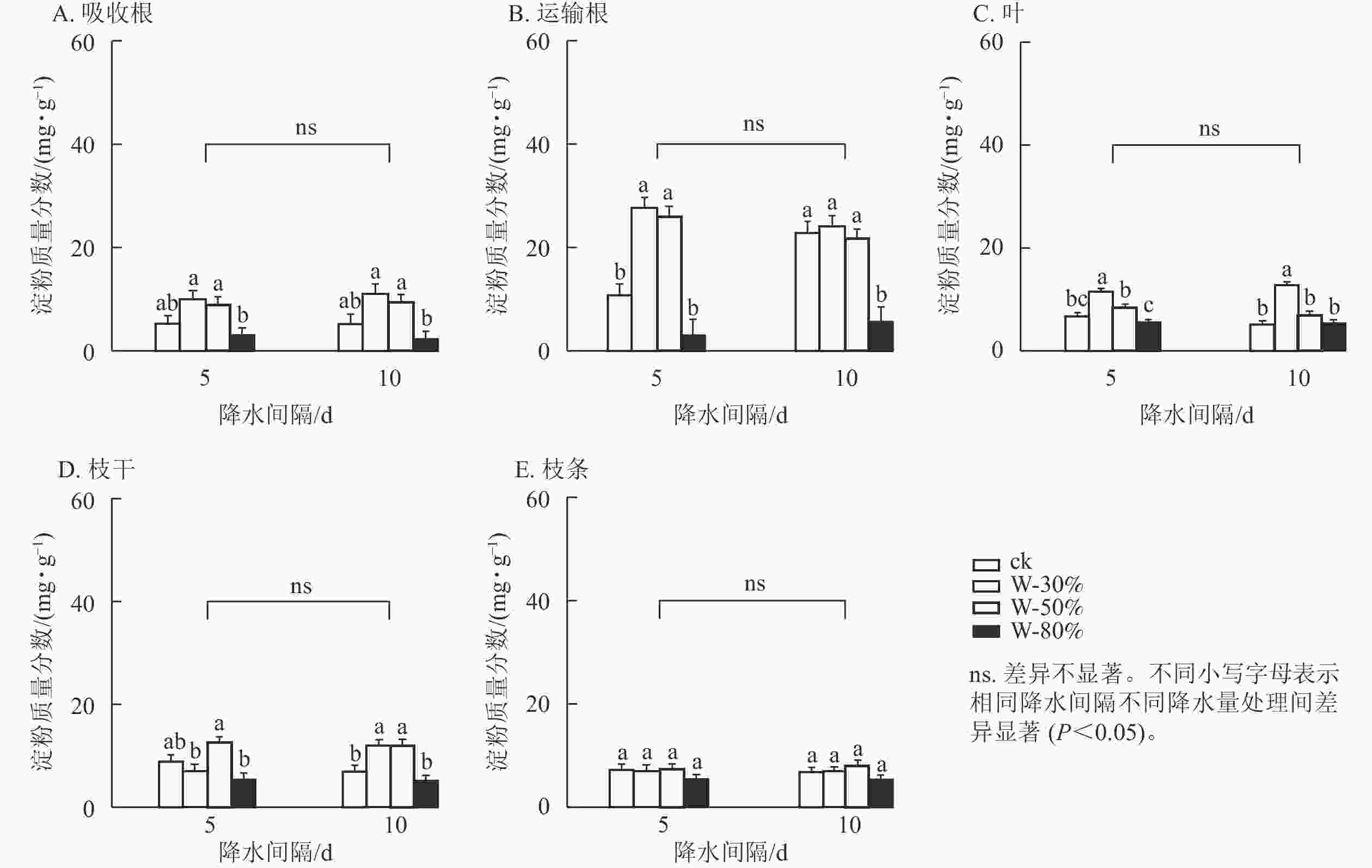
图 2 不同降水量和降水间隔对杉木幼苗各器官淀粉质量分数的影响
Figure 2. Effects of different precipitation amount and intervals on starch content in different organs of C. lanceolata seedlings
-
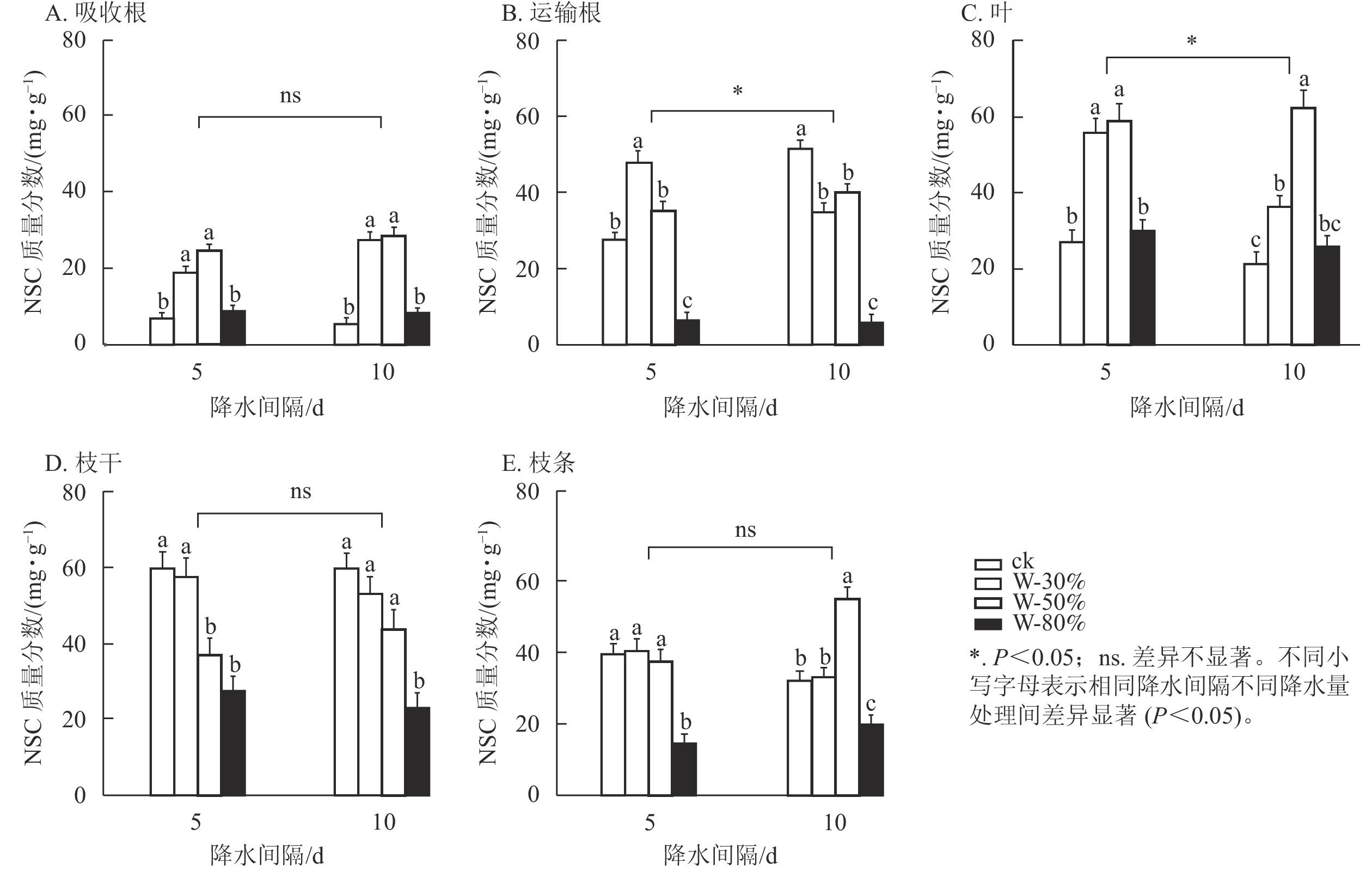
降水量对杉木幼苗不同器官NSC质量分数具有极显著影响(P<0.01),降水量和降水间隔的共同作用对NSC质量分数整体有极显著影响(P<0.01),降水间隔则整体无显著影响(表1)。T5处理下,随着降水量的减少,NSC质量分数在吸收根、运输根、叶和枝条中呈先增加后减少的趋势(图3),其中,吸收根和叶的NSC质量分数在W-50%处理下最大,均显著高于ck (P<0.05),分别为ck的3.6和2.2倍;运输根和枝条的NSC质量分数在W-80%处理下最小,均显著低于ck (P<0.05),分别降低了79.5%和63.3%;NSC质量分数在枝干中表现为持续减少,在W-80%处理下最小,显著低于ck (P<0.05),比ck降低了53.9%。T10处理下,随着降水量的减少,NSC质量分数在吸收根、叶和枝条中呈先增加后减少的趋势,且吸收根、叶和枝条的NSC质量分数均在W-50%处理下最大,显著高于ck (P<0.05),分别为ck的5.3、3.0和1.7倍;NSC质量分数在运输根中表现为先减少后增加再减少,在枝干中表现为持续减少,均在W-80%处理下最小,显著低于ck (P<0.05),分别降低了88.7%和61.5%。
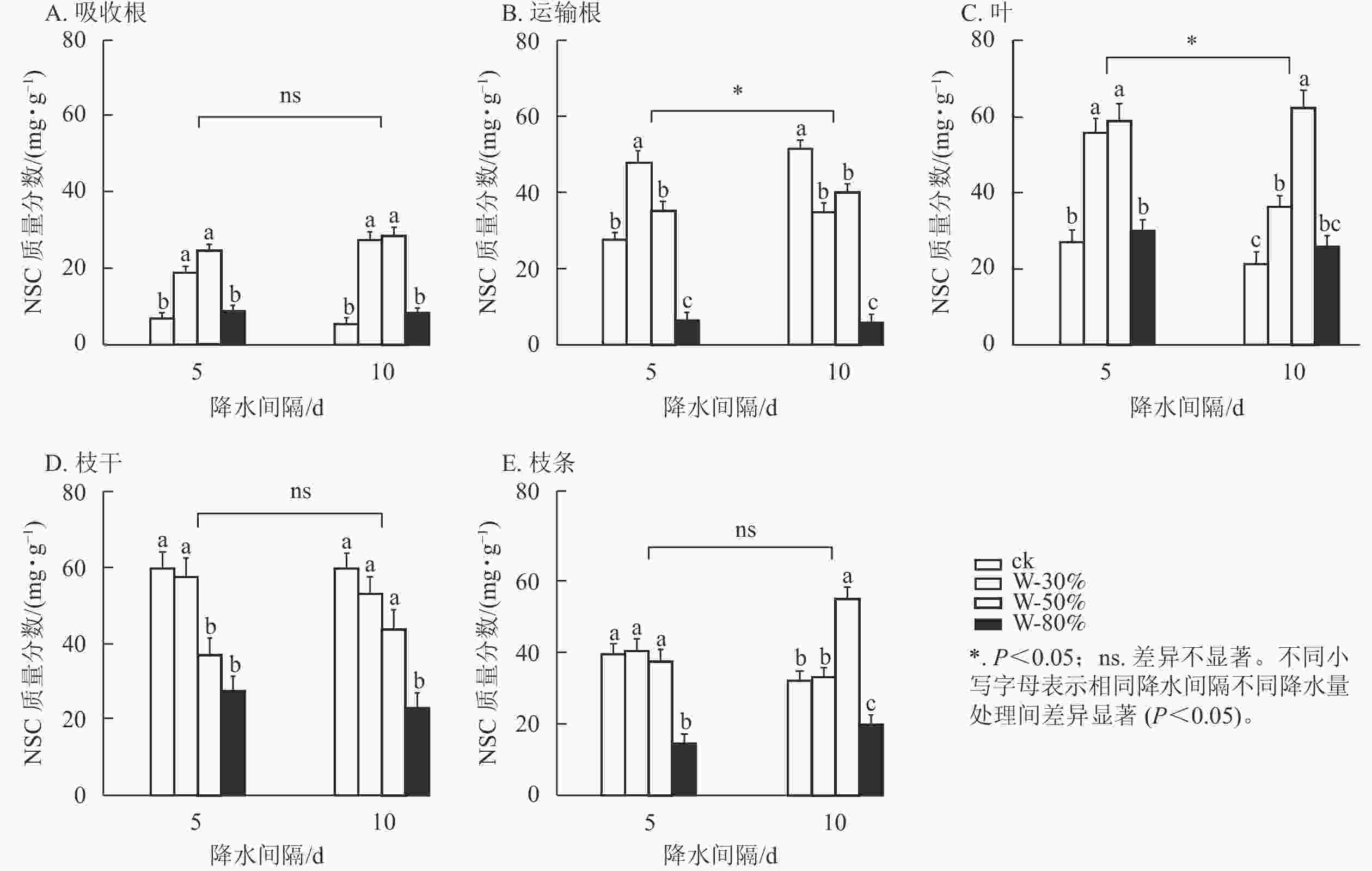
图 3 不同降水量和降水间隔对杉木幼苗各器官NSC质量分数的影响
Figure 3. Effects of different precipitation amount and intervals on NSC content in different organs of C. lanceolata seedlings
-
NSC在树木生长和新陈代谢过程中发挥着重要作用,其质量分数和分布可用于推断植物碳储量状况以及对不同环境的生长适应性[31−32]。本研究发现:杉木幼苗不同器官可溶性糖、淀粉和NSC质量分数对降水量变化具有显著响应。植物体内NSC积累可减少基本代谢活动,延长存活时间[31];可溶性糖的增加可降低植物水势,维持细胞膨胀压力,增加植物的吸水量[33];淀粉的增加可促进植物抗性的提高,在逆境条件下为植物提供更多的能量[34]。当降水量减少时,杉木幼苗可溶性糖主要分布于叶、枝条和枝干等地上部分,这可能是因为树干、枝条、叶中的淀粉水解转化为可溶性糖,有利于提高渗透势,促进植物吸水,最终增强植物光合固碳的过程并促进植物生长[35],且树干和枝条中更多的NSC也有利于提高杉木幼苗的抗逆性[36]。这说明可溶性糖直接参与植物的生理活动,其渗透调节功能是植物应对干旱胁迫的重要方式[37],而叶、枝条和枝干是树木的碳源器官[38],树木代谢活动的中间产物暂时储存在叶片中,降水量减少情况下需要大量的可溶性糖来帮助其维持正常的细胞张力[39]。植物体内可溶性糖和淀粉成分的比例和动态变化,即NSC的储存和转化机制,在缺水条件下可以维持植物生长、呼吸、繁殖等功能[40]。一定程度的干旱会促进植物体淀粉转化为可溶性糖,维持细胞膨压、抵御并适应干旱环境[41]。而淀粉作为植物体内重要的能量储存物质,其质量分数高低代表了植物耐受力的强弱。当降水量减少到一定程度时(W-30%、W-50%),淀粉主要存在于运输根和枝干等运输器官中。植物具有固碳能力,杉木幼苗通过存储更多的淀粉来应对缺水环境,以保持其新陈代谢的稳定[42];但降水量过少时(W-80%),树木会加快分解淀粉[43],导致淀粉质量分数先增后降。因此,降水量减少到一定程度时,会促进杉木幼苗的生长,导致NSC及其组分在植物体内形成和积累;杉木幼苗对降水量变化的响应是增加运输根、吸收根、叶和枝条的可溶性糖和淀粉,同时将吸收根中的淀粉水解为可溶性糖,以调节细胞内的水势,从而应对降水格局的变化。当降水量过少时,杉木幼苗通过减少运输根、枝干和枝条的NSC来应对降水格局的变化,各器官内的可溶性糖和淀粉质量分数并未发生转化,这是因为降水量过少时杉木会消耗体内的NSC及其组分,优先供给吸收根的生存需求。这与云南松Pinus yunnanensis[44]及樟子松Pinus sylvestris var. mongolica[45]的研究结果相似。
-
降水的频率是影响植物生存、生长、物种组成及结构的重要因素[46],降水间隔的变化会影响植物遭受干旱的持续时间,而植物正常的生理代谢活动如生长、光合作用、呼吸作用等会受到干旱胁迫的影响,破坏植物体内碳供需的平衡,碳的转化和储存之间的关系也会随之改变,此时植物会调动储存在机体内的NSC,使其发生转移或转化,引起不同干旱时间下NSC的动态变化[47]。本研究发现:降水间隔和杉木幼苗不同器官之间的交互作用显著影响可溶性糖和NSC质量分数。随着降水间隔的增加,杉木幼苗各器官NSC及其组分质量分数呈波动性变化,运输根和叶的可溶性糖有显著差异,与T5处理相比,T10处理下叶的可溶性糖质量分数显著下降,运输根的可溶性糖质量分数显著上升;运输根和叶的NSC质量分数有显著差异,与T5处理相比,延长降水间隔导致叶和运输根的NSC质量分数显著上升。可能是因为延长降水间隔时,杉木新吸收的碳水化合物主要流向运输根,以维持资源供应;可溶性糖在叶片中转化为淀粉,增加细胞膨胀压力,提高杉木的抗旱能力。在延长降水间隔的条件下,杉木选择将生物量更多地分配给能够获取资源的器官(根和叶),而非主要起运输和支持作用的器官(茎),尽可能增大叶片面积来增加叶片碳水化合物的供应,从而维持自身的光合和生长过程,这与郭旭曼等[48]在桢楠Phoebe zhennan上的研究相似。杉木幼苗各器官的NSC并没有全部随着时间的推移而减少,这可能是由于在长期无降水的情况下,植物体内储存的碳未被充分利用或碳储量的利用率有限,因此即使植物死亡也不会发生碳耗竭[49−50]。由于杉木林深层土壤含水量较低,干旱下幼苗可通过降低生长,将NSC及其相关组分供给根系等获取水分的器官,但仍需根据当地气候条件来调整浇水间隔,高降水量条件下适当延长降水间隔更有利于植物利用地表和深层土壤水分[51],使幼苗更好地成长。建议维持土壤含水量不低于当地多年旱季月平均降水量的50%,以保证杉木林中萌发的幼苗能够存活,完成天然更新。在经济条件较好的地区,可以实行定期喷灌的管理,保持土壤含水量在当地多年旱季月平均降水量的70%以上,同时根据当地气候条件可以适当延长浇水间隔,提高杉木幼苗存活率。
-
为适应环境的变化,不同降水格局下杉木幼苗NSC及其相关组分会发生不同的转化和增减。当降水量减少时,杉木幼苗叶、枝条和枝干等地上器官NSC中可溶性糖的转化比例增加,储藏在根系,由运输根转移至吸收根,促进植物生长;延长降水间隔能促使杉木幼苗叶中的NSC转移到根系,提高根系水分获取能力,防止植物因缺水而死亡。为提高杉木人工林在降水格局变化下的存活率,土壤含水量应保持在当地多年旱季月平均降水量的50%以上,降水量较高地区可以适当延长浇水间隔。
Effects of precipitation pattern change on non-structural carbohydrates in different organs of Cunninghamia lanceolata seedlings
-
摘要:
目的 全球气温上升和降水格局变化导致的频繁干旱对森林生态系统造成了重大影响,因此,研究植物干旱前后水分利用和适应的机制至关重要。 方法 以杉木Cunninghamia lanceolata幼苗为研究对象,设置4个降水量梯度和2个降水间隔的双因素控制试验,研究杉木幼苗不同器官中非结构性碳水化合物(NSC)及其相关组分质量分数对降水量和降水间隔变化的响应。 结果 随着降水量减少,杉木幼苗的NSC质量分数在吸收根、运输根、枝条和叶中均呈先增加后减少的趋势,在枝干中则持续下降,杉木幼苗通过增加运输根、吸收根、叶和枝条的NSC质量分数,同时将吸收根的淀粉水解转化成可溶性糖,来调节细胞内的水势;可溶性糖质量分数在运输根和枝干中均呈持续减少的趋势,在吸收根、叶和枝条中先增加后减少;淀粉质量分数在所有器官中均呈先增加后减少的趋势;当降水量过少时,杉木幼苗会消耗体内的NSC,优先供给吸收根的生存需求,导致运输根、枝条和枝干的NSC质量分数显著下降(P<0.05)。延长降水间隔,杉木幼苗叶和运输根的NSC质量分数显著上升(P<0.05);叶的可溶性糖质量分数显著下降(P<0.05),运输根的可溶性糖质量分数显著上升(P<0.05)。 结论 土壤含水量应保持在当地多年旱季月平均降水量的50%以上,可提高杉木人工林在降水格局变化下的存活率,降水量较高的地区可以适当延长浇水间隔。图3表1参51 -
关键词:
- 降水格局 /
- 杉木 /
- 不同器官 /
- 非结构性碳水化合物(NSC)
Abstract:Objective Frequent droughts caused by global temperature rise and precipitation pattern change have significant impacts on forest ecosystems, so information of plant water utilization and adaptation during and after droughts is crucial. Method The seedlings of Cunninghamia lanceolata were used as test material. A two-factor controlled experiment with 4 precipitation amount and 2 precipitation intervals gradients was set to investigate the response of non-structural carbohydrates (NSC) and related fractions content in different organs that suffer different precipitation amount and intervals. Result With the decrease of precipitation, NSC content of the seedlings showed an overall trend of firstly increasing and then decreasing in absorbing roots, transport roots, branches and leaves, and continuous decreasing in branches and trunks. C. lanceolata seedlings increased NSC content in transport roots, absorbing roots, leaves and branches, while hydrolyzed and converted the starch from the absorbing roots to soluble sugars to regulate intracellular water potentials; soluble sugars content showed an overall trend of continuous decreasing in transport roots and trunks, and an overall trend of firstly increasing and then decreasing in absorbing roots, leaves and branches; starch content of all organs showed an overall trend of firstly increasing and then decreasing. When precipitation was too low, C. lanceolata seedlings would consume NSC to preferentially meet the survival needs of absorbing roots, which led to a significant decrease of NSC content in transport roots, branches and twigs (P<0.05). When precipitation intervals increased, NSC content in leaves and transport roots of the seedlings increased significantly (P<0.05); soluble sugar content of leaves decreased significantly (P<0.05), and soluble sugar content of transported roots increased significantly (P<0.05). Conclusion To improve the survival of C. lanceolata plantation forests under changes in precipitation patterns, soil water content should be maintained at more than 50% of the average monthly precipitation in the local multi-year dry season, and watering intervals can be extended in areas with higher precipitation. [Ch, 3 fig. 1 tab. 51 ref.] -
图 1 不同降水量和降水间隔对杉木幼苗各器官可溶性糖质量分数的影响
Figure 1 Effects of different precipitation amount and intervals on soluble sugar content in different organs of C. lanceolata seedlings
图 2 不同降水量和降水间隔对杉木幼苗各器官淀粉质量分数的影响
Figure 2 Effects of different precipitation amount and intervals on starch content in different organs of C. lanceolata seedlings
图 3 不同降水量和降水间隔对杉木幼苗各器官NSC质量分数的影响
Figure 3 Effects of different precipitation amount and intervals on NSC content in different organs of C. lanceolata seedlings
表 1 降水格局对杉木幼苗不同器官可溶性糖、淀粉、NSC质量分数影响的线性混合模型分析
Table 1. Effect of precipitation patterns on soluble sugar, starch and NSC content of different organs of C. lanceolata seedlings based on analysis of linear mixed-effects models
影响因素 可溶性糖 淀粉 NSC F P F P F P W 29.06 <0.001 87.72 <0.001 129.55 <0.001 T 0.98 0.320 0.20 0.660 2.73 0.100 G 66.30 <0.001 119.74 <0.001 98.46 <0.001 W×T 2.56 <0.100 2.18 <0.100 9.28 <0.001 W×G 5.42 <0.001 11.79 <0.001 20.13 <0.001 T×G 3.83 <0.010 0.38 0.820 3.98 <0.010 W×T×G 0.96 0.490 4.57 <0.001 4.03 <0.001 说明:W. 不同降水量处理;T. 不同降水间隔处理;G. 杉木幼苗不同器官;×表示交互作用。P<0.01表示差异极显著。  下载: 导出CSV
下载: 导出CSV
-
[1] 岳永寰, 靳瑰丽, 宫珂, 等. 人工模拟降水格局变化对醉马草种子萌发和幼苗生长的影响[J]. 生态学杂志, 2020, 39(3): 838 − 846. YUE Yonghuan, JIN Guili, GONG Ke, et al. Effects of simulated precipitation pattern changes on seed germination and seedling growth of Achnatherum inebrians [J]. Chinese Journal of Ecology, 2020, 39(3): 838 − 846. [2] DU Ying, LU Ruiling, XIA Jianyang. Impacts of global environmental change drivers on non-structural carbohydrates in terrestrial plants [J]. Functional Ecology, 2020, 34(8): 1525 − 1536. [3] OHTO M, ONAI K, FURUKAWA Y, et al. Effects of sugar on vegetative development and floral transition in Arabidopsis [J]. Plant Physiology, 2001, 127(1): 252 − 261. [4] 董涵君, 王兴昌, 苑丹阳, 等. 温带不同材性树种树干非结构性碳水化合物的径向分配差异[J]. 植物生态学报, 2022, 46(6): 722 − 734. DONG Hanjun, WANG Xingchang, YUAN Danyang, et al. Radial distribution differences of non-structural carbohydrates in stems of tree species of different wood in a temperate forest [J]. Chinese Journal of Plant Ecology, 2022, 46(6): 722 − 734. [5] MYERS J A, KITAJIMA K. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest [J]. Journal of Ecology, 2007, 95(2): 383 − 395. [6] KÖRNER C. Carbon limitation in trees [J]. Journal of Ecology, 2003, 91(1): 4 − 17. [7] WILEY E, HELLIKER B. A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth [J]. New Phytologist, 2012, 195(2): 285 − 289. [8] 雷虹, 王凯, 田浩, 等. 小叶锦鸡儿幼苗非结构性碳水化合物积累及分配对干旱胁迫的响应[J]. 生态学杂志, 2017, 36(11): 3168 − 3175. LEI Hong, WANG Kai, TIAN Hao, et al. Responses of non-structural carbohydrates accumulation and distribution of Caragana microphylla seedlings to drought stress [J]. Chinese Journal of Ecology, 2017, 36(11): 3168 − 3175. [9] 刘志林, 丁银平, 角媛梅. 中国西南—东南季风交汇区降水时空格局变化及其对食物产量的影响[J]. 地理学报, 2021, 76(9): 2297 − 2311. LIU Zhilin, DING Yinping, JIAO Yuanmei. Spatiotemporal patterns of precipitation changes and their impacts on food supply in southwest China from 1988 to 2018: a case study in Yunnan Province [J]. Acta Geographica Sinica, 2021, 76(9): 2297 − 2311. [10] AULT T R. On the essentials of drought in a changing climate [J]. Science, 2020, 368(6488): 256 − 260. [11] HU Tian, van DIJK A I J M, RENZULLO L J, et al. On agricultural drought monitoring in Australia using Himawari-8 geostationary thermal infrared observations [J/OL]. International Journal of Applied Earth Observation and Geoinformation, 2020, 91: 102153[2024-03-19]. doi: 10.1016/j.jag.2020.102153. [12] CHEN S A, MICHAELIDES K, GRIEVE S W D, et al. Aridity is expressed in river topography globally [J]. Nature, 2019, 573(7775): 573 − 577. [13] KONAPALA G, MISHRA A K, WADA Y, et al. Climate change will affect global water availability through compounding changes in seasonal precipitation and evaporation [J/OL]. Nature Communications, 2020, 11(1): 3044 [2024-03-19]. doi: 10.1038/s41467-020-16757-w. [14] VALMASSOI A, DUDHIA J, DI SABATINO S, et al. Irrigation impact on precipitation during a heatwave event using WRF-ARW: the summer 2015 Po Valley case [J/OL]. Atmospheric Research, 2020, 241: 104951[2024-03-19]. doi: 10.1016/j.atmosres.2020.104951. [15] BLÖSCHL G, HALL J, VIGLIONE A, et al. Changing climate both increases and decreases European river floods [J]. Nature, 2019, 573(7772): 108 − 111. [16] NYSTRÖM M, JOUFFRAY J B, NORSTRÖM A V, et al. Anatomy and resilience of the global production ecosystem [J]. Nature, 2019, 575(7781): 98 − 108. [17] LIU Ziqiang, YU Xinxiao, JIA Guodong, et al. Water uptake by coniferous and broad-leaved forest in a rocky mountainous area of northern China [J]. Agricultural and Forest Meteorology, 2019, 265: 381 − 389. [18] MURRAY R B, THRALL H P, GILL M A, et al. How plant life-history and ecological traits relate to species rarity and commonness at varying spatial scales [J]. Austral Ecology, 2002, 27(3): 291 − 310. [19] 张蕊, 赵学勇, 左小安, 等. 荒漠草原沙生针茅(Stipa glareosa)群落物种多样性和地上生物量对降雨量的响应[J]. 中国沙漠, 2019, 39(2): 45 − 52. ZHANG Rui, ZHAO Xueyong, ZUO Xiao’an, et al. Responses of the Stipa glareosa community species diversity and above-ground biomass to precipitation in the desert-steppe region in northern China [J]. Journal of Desert Research, 2019, 39(2): 45 − 52. [20] 王雲霞, 单立山, 解婷婷, 等. 干旱-复水对红砂幼苗各器官非结构性碳水化合物的影响[J]. 生态学杂志, 2024, 43(2): 383 − 394. WANG Yunxia, SHAN Lishan, XIE Tingting, et al. The effects of drought-rehydration on n on-structural carbohydrates in Reaumuria soongorica seedlings [J]. Chinese Journal of Ecology, 2024, 43(2): 383 − 394. [21] 林恬, 郑怀舟, 朱锦懋. 隔离降雨对马尾松针叶非结构性碳水化合物和碳、氮、磷的影响[J]. 生态学报, 2022, 42(18): 7641 − 7651. LIN Tian, ZHENG Huaizhou, ZHU Jinmao. Influence of rainfall exclusion on the needle non-structural carbohydrates and C, N, P concentrations in Pinus massoniana [J]. Acta Ecologica Sinica, 2022, 42(18): 7641 − 7651. [22] 李秉钧, 颜耀, 张辉, 等. 不同种源杉木细根根序及碳氮计量的比较分析[J]. 森林与环境学报, 2019, 39(6): 561 − 567. LI Bingjun, YAN Yao, ZHANG Hui, et al. Comparison of fine root morphology and carbon and nitrogen content in Chinese fir from different provenances [J]. Journal of Forest and Environment, 2019, 39(6): 561 − 567. [23] 陈炎根, 胡艳静, 黄莎, 等. 不同间伐强度对杉木人工林土壤呼吸速率的短期影响[J]. 浙江农林大学学报, 2023, 40(5): 1054 − 1062. CHEN Yan’gen, HU Yanjing, HUANG Sha, et al. Short-term effects of different thinning intensities on soil respiration rate in the Cunninghamia lanceolata plantation [J]. Journal of Zhejiang A&F University, 2023, 40(5): 1054 − 1062. [24] 林雨萱, 哀建国, 宋新章, 等. 模拟氮沉降和磷添加对杉木林土壤呼吸的影响[J]. 浙江农林大学学报, 2021, 38(3): 494 − 501. LIN Yuxuan, AI Jianguo, SONG Xinzhang, et al. Effects of simulated nitrogen deposition and phosphorus addition on soil respiration in Chinese fir forest [J]. Journal of Zhejiang A&F University, 2021, 38(3): 494 − 501. [25] 杨振亚, 周本智, 陈庆标, 等. 干旱对杉木幼苗根系构型及非结构性碳水化合物的影响[J]. 生态学报, 2018, 38(18): 6729 − 6740. YANG Zhenya, ZHOU Benzhi, CHEN Qingbiao, et al. Effects of drought on root architecture and non-structural carbohydrate of Cunninghamia lanceolata [J]. Acta Ecologica Sinica, 2018, 38(18): 6729 − 6740. [26] 金涛涛, 张佛熠, 郑伟斌, 等. 南昌城乡不同绿地中小型土壤动物群落多样性及其影响因素[J]. 应用生态学报, 2023, 34(5): 1404 − 1414. JIN Taotao, ZHANG Foyi, ZHENG Weibin, et al. Diversity of medium and small-sized soil fauna community in different urban-rural green spaces and its influencing factors in Nanchang, China [J]. Chinese Journal of Applied Ecology, 2023, 34(5): 1404 − 1414. [27] 张瑞香. 不同林龄野生南方红豆杉碳氮磷生态化学计量与非结构性碳水化合物特征研究[D]. 郑州: 河南农业大学, 2024. ZHANG Ruixiang. Carbon, Nitrogen, and Phosphorus Stoichiometry and Non-structural Carbohydrate in Different Stand Ages in Wild Taxus wallichiana var. maire [D]. Zhengzhou: Henan Agricultural University, 2024. [28] 种培芳, 曾继娟, 单立山, 等. 干旱胁迫下荒漠草地植物红砂幼苗对外源ABA的生理响应[J]. 草地学报, 2016, 24(5): 1001 − 1008. CHONG Peifang, ZENG Jijuan, SHAN Lishan, et al. The physiological response of desert grassland plant Reamuria soongorica under drought stress to exogenous ABA [J]. Acta Agrestia Sinica, 2016, 24(5): 1001 − 1008. [29] 梁宽, 樊燕, 冯火炬, 等. 不同石灰岩生境淡竹非结构性碳水化合物浓度及分配特征[J]. 林业科学, 2019, 55(6): 22 − 27. LIANG Kuan, FAN Yan, FENG Huoju, et al. Concentration and distribution pattern of non-structural carbohydrate of Phyllostachys glauca in different limestone habitats [J]. Scientia Silvae Sinicae, 2019, 55(6): 22 − 27. [30] 欧阳园丽, 张参参, 林小凡, 等. 中国亚热带不同菌根树种的根叶形态学性状特征与生长差异: 以江西新岗山为例[J]. 生物多样性, 2021, 29(6): 746 − 758. OUYANG Yuanli, ZHANG Cancan, LIN Xiaofan, et al. Growth differences and characteristics of root and leaf morphological traits for different mycorrhizal tree species in the subtropical China: a case study of Xin’gangshan, Jiangxi Province [J]. Biodiversity Science, 2021, 29(6): 746 − 758. [31] JIN Yanqiang, LI Jing, LIU Chenggang, et al. Carbohydrate dynamics of three dominant species in a Chinese savanna under precipitation exclusion [J]. Tree Physiology, 2018, 38(9): 1371 − 1383. [32] 王凯, 沈潮, 曹鹏, 等. 沙地樟子松幼苗干旱致死过程中非结构性碳水化合物的变化[J]. 应用生态学报, 2018, 29(11): 3513 − 3520. WANG Kai, SHEN Chao, CAO Peng, et al. Changes of non-structural carbohydrates of Pinus sylvestris var. mongolica seedlings in the process of drought-induced mortality [J]. Chinese Journal of Applied Ecology, 2018, 29(11): 3513 − 3520. [33] SILVA E N, FERREIRA-SILVA S L, VIÉGAS R A, et al. The role of organic and inorganic solutes in the osmotic adjustment of drought-stressed Jatropha curcas plants [J]. Environmental and Experimental Botany, 2010, 69(3): 279 − 285. [34] 王凯, 宋琪, 张日升, 等. 科尔沁沙地防护林主要树种的非结构性碳水化合物分布特征[J]. 林业科学, 2020, 56(12): 39 − 48. WANG Kai, SONG Qi, ZHANG Risheng, et al. Distribution characteristics of non-structural carbohydrate in main tree species of shelterbelt forests in Horqin sandy land [J]. Scientia Silvae Sinicae, 2020, 56(12): 39 − 48. [35] 王睿照, 毛沂新, 云丽丽, 等. 氮添加对蒙古栎叶片碳氮磷化学计量与非结构性碳水化合物的影响[J]. 生态学杂志, 2022, 41(7): 1369 − 1377. WANG Ruizhao, MAO Yixin, YUN Lili, et al. Effects of nitrogen addition on leaf carbon, nitrogen and phosphorus stoichiometry and nonstructural carbohydrates in Mongolian oak (Quercus mongolica) [J]. Chinese Journal of Ecology, 2022, 41(7): 1369 − 1377. [36] 苏炜, 陈平, 吴婷, 等. 氮添加与干季延长对降香黄檀幼苗非结构性碳水化合物, 养分与生物量的影响[J]. 植物生态学报, 2023, 47(8): 1094 − 1104. SU Wei, CHEN Ping, WU Ting, et al. Effects of nitrogen addition and extended dry season on non-structural carbohydrates, nutrients and biomass of Dalbergia odorifera seedlings [J]. Chinese Journal of Plant Ecology, 2023, 47(8): 1094 − 1104. [37] ABRAHAM B. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production [J]. Plant,Cell &Environment, 2017, 40(1): 4 − 10. [38] CHAPIN III F S, SCHULZE E D, MOONEY H A. The ecology and economics of storage in plants [J]. Annual Review of Ecology and Systematics, 1990, 21: 423 − 447. [39] PIPER I F. Drought induces opposite changes in the concentration of non-structural carbohydrates of two evergreen Nothofagus species of differential drought resistance [J]. Annals of Forest Science, 2011, 68(2): 415 − 424. [40] LU Yanwei, DUAN Baoli, ZHANG Xiaolu, et al. Differences in growth and physiological traits of Populus cathayana populations as affected by enhanced UV-B radiation and exogenous ABA [J]. Environmental and Experimental Botany, 2009, 66(1): 100 − 109. [41] 王云霞, 刘莹, 付雨辰, 等. 干旱胁迫对连翘幼苗非结构性碳分配和水力特性的影响[J]. 生态学报, 2024, 44(11): 4698 − 4707. WANG Yunxia, LIU Ying, FU Yuchen, et al. Effects of drought on non-structural carbon allocation and hydraulic characteristics of Forsythia suspense seedlings [J]. Acta Ecologica Sinica, 2024, 44(11): 4698 − 4707. [42] 杨玉, 张芸, 魏绪英, 等. 应用13C示踪技术研究红花石蒜非结构性碳水化合物分配及转运模式[J]. 浙江农林大学学报, 2024, 41(2): 252 − 261. YANG Yu, ZHANG Yun, WEI Xuying, et al. Distribution and transport patterns of NSC in Lycoris radiata based on 13C tracing [J]. Journal of Zhejiang A&F University, 2024, 41(2): 252 − 261. [43] LIU Hongyan, SHANGGUAN Huailiang, ZHOU Mei, et al. Differentiated responses of nonstructural carbohydrate allocation to climatic dryness and drought events in the Inner Asian arid timberline [J]. Agricultural and Forest Meteorology, 2019, 271: 355 − 361. [44] 刘元玺, 王丽娜, 吴俊文, 等. 云南松幼苗非结构性碳水化合物对干旱胁迫的响应及其激素调控[J]. 西北农林科技大学学报(自然科学版), 2024, 52(1): 60 − 70. LIU Yuanxi, WANG Li’na, WU Junwen, et al. Response of non-structural carbohydrates of Pinus yunnanensis seedlings to drought stress and the hormonal regulation mechanism [J]. Journal of Northwest A&F University (Natural Science Edition), 2024, 52(1): 60 − 70. [45] 王凯, 芦珊, 刘畅, 等. 土壤含水量对樟子松幼苗非结构性碳水化合物及生长的影响[J]. 生态学杂志, 2023, 42(3): 617 − 625. WANG Kai, LU Shan, LIU Chang, et al. Effects of soil water content on non-structural carhohydrates and growth of Pinus sylvestris var. mongolica seedlings [J]. Chinese Journal of Ecology, 42(3): 617 − 625. [46] CHENG Xiaoli, AN Shuqing, LI Bo, et al. Summer rain pulse size and rainwater uptake by three dominant desert plants in a desertified grassland ecosystem in northwestern China [J]. Plant Ecology, 2006, 184(1): 1 − 12. [47] MCDOWELL N G. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality [J]. Plant Physiology, 2011, 155(3): 1051 − 1059. [48] 郭旭曼, 王佳敏, 杜浩瀚, 等. 桢楠幼苗适应喀斯特岩溶裂隙生境及降雨时间格局变化的方式[J]. 生态学报, 2023, 43(1): 379 − 387. GUO Xuman, WANG Jiamin, DU Haohan, et al. Phoebe zhennan S. Lee seedlings adjust the biomass allocation and root distribution to adapt to the karst fissure habitat and rainfall temporal pattern [J]. Acta Ecologica Sinica, 2023, 43(1): 379 − 387. [49] ANNA S, FRIDA P, GÜNTER H. Physiological mechanisms of drought-induced tree mortality are far from being resolved [J]. New Phytologist, 2010, 186(2): 274 − 281. [50] HARTMANN H, TRUMBORE S. Understanding the roles of nonstructural carbohydrates in forest trees-from what we can measure to what we want to know [J]. New Phytologist, 2016, 211(2): 386 − 403. [51] 段桂芳, 单立山, 李毅, 等. 降水格局变化对红砂幼苗生长的影响[J]. 生态学报, 2016, 36(20): 6457 − 6464. DUAN Guifang, SHAN Lishan, LI Yi, et al. Effects of changing precipitation patterns on seedling growth of Reaumuria soongorica [J]. Acta Ecologica Sinica, 2016, 36(20): 6457 − 6464. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240253
 点击查看大图
点击查看大图
计量
- 文章访问数: 1630
- HTML全文浏览量: 507
- PDF下载量: 55
- 被引次数: 0
