






-
杉木Cunninghamia lanceolata是中国南方最重要的速生造林树种,生长速度快,成材时间较短,其造林面积和蓄积量均居中国用材林树种的首位,人工林面积达8.95×106 hm2,占中国人工林面积的30%以上[1],木材产量约占全国商品材的25%[2]。长期以来杉木人工林经营主要依靠营造大面积、高密度的速生丰产林来满足市场对木材的需求[3]。这种经营模式下的杉木人工林往往只能输出中小径材,杉木大径材数量少,林分生产力和经营效益不高。大力发展杉木大径材定向培育技术已成为杉木人工林经营的重要方向[4]。20世纪90年代末,中国才开始开展对杉木大径材的培育技术研究,研究主要集中在良种壮苗选择、立地选择、林分密度调控、施肥等经营措施上[5],但缺乏杉木大径材培育的材种结构形成机制、不同培育措施的组装配套等关键技术的系统研究。间伐和施肥是大径材培育中用到较多的抚育措施,林分密度越大,小径阶林木株数越多,大中径阶林木株数越少[6],合理密度能有效改变林分的材种结构[7],带来最直观的林地生产效益和经济效益。晏姝等[8]对南岭山区现存杉木大径材林分的调查表明:林分密度和土壤肥力是培育杉木大径材的主导因子。但目前有关杉木人工林间伐和施肥管理的研究对象大多是幼龄林和中龄林,对于间伐和精准配方施肥共同作用于杉木成熟林的研究较少。本研究分析了不同间伐保留密度及不同氮、磷肥施用量对杉木成熟林材种结构、生产力、大径材出材量和出材率的影响,以期为高效培育杉木大径材提供参考。
-
试验林位于福建省南平市顺昌县洋口林场,地形属武夷山支脉的低山丘陵地带,海拔为300~700 m。属中亚热带海洋性季风气候区,年平均气温为18.6 ℃,年平均降水量为1 300~2 100 mm,无霜期可达305 d,气候温暖,雨量充沛。土壤以山地红壤为主,是适合杉木大径材培育的中心产区。样地内土壤理化性质见表1。
表 1 试验地土壤理化性质
Table 1. Physical properties of soil in the experimental site
土层/cm 最大持
水量/mm毛管持
水量/mm最小持
水量/mm非毛管孔
隙度/%毛管孔
隙度/%总孔隙
度/%土壤密度/
(g·cm−3)氮/
(g·kg−1)磷/
(g·kg−1)0~10 71.53±0.00 66.33±0.00 64.93±0.00 5.20±0.00 66.33±0.00 71.53±0.00 2.66±0.15 24.11±5.01 1.60±0.38 10~20 72.13±0.00 63.53±0.00 61.47±0.00 8.60±0.00 63.53±0.00 72.13±0.00 2.68±0.10 26.04±4.05 1.01±0.30 20~40 135.80±0.01 123.00±0.01 119.60±0.01 6.40±0.00 61.50±0.00 67.90±0.01 2.87±0.14 26.50±5.37 0.70±0.16 -
样地选自27年生立地指数均大于22的杉木成熟人工林,处理前林分密度为800~850株·hm−2。采用正交试验,设置集中连片的9块样地,设间伐保留密度、氮肥施用量、磷肥施用量共3个因素。间伐保留密度设置为300、600、825 (不间伐)株·hm−2,氮肥(尿素)施用量设置为0、100、200 g·株−1,磷肥(钙镁磷肥)施用量设置为0、250、500 g·株−1。间伐和施肥处理于2016年的11月杉木生长停滞期进行,施肥采用沟施方法,在每株树上坡开一条弧形沟,沟在树冠正投影下,沟长约100 cm,宽约20 cm,深约30 cm,氮肥和磷肥一次性施入后立即覆土。各样地具体处理措施如表2。
表 2 各样地间伐施肥处理情况
Table 2. Various thinning and fertilization treatments
样地号 间伐保留密度/
(株·hm−2)氮肥施用量/
(g·株−1)磷肥施用量/
(g·株−1)样地号 间伐保留密度/
(株·hm−2)氮肥施用量/
(g·株−1)磷肥施用量/
(g·株−1)1 825 0 0 6 600 200 0 2 600 0 500 7 300 0 250 3 825 100 500 8 825 200 250 4 300 100 0 9 300 200 500 5 600 100 250 -
2017—2020年每年的4月对各样地的杉木进行标记号牌管理和每木检尺,采用Haglof Vertex Ⅳ超声波树木测高仪测量树高,以围径尺测定胸径,以5 m标杆测量南北冠幅和东西冠幅,并以2017年的数据作为间伐和施肥处理前的生长情况。
-
为方便统计和计算,对杉木胸径进行径阶划分,以2 cm为1个径阶,以16为起测径阶,其立木胸径为[15 cm, 17 cm),18径阶为[17 cm, 19 cm),20径阶为[19 cm, 21 cm),以此类推。对各径阶株数进行统计,并分别计算各径阶杉木的平均胸径(Di),计算公式为
$D_i = \sqrt {\dfrac{1}{N_i}\displaystyle \sum {d_i^{\text{2}}}}$ 。其中,Ni表示第i径阶总株数,di表示第i径阶胸径值。 -
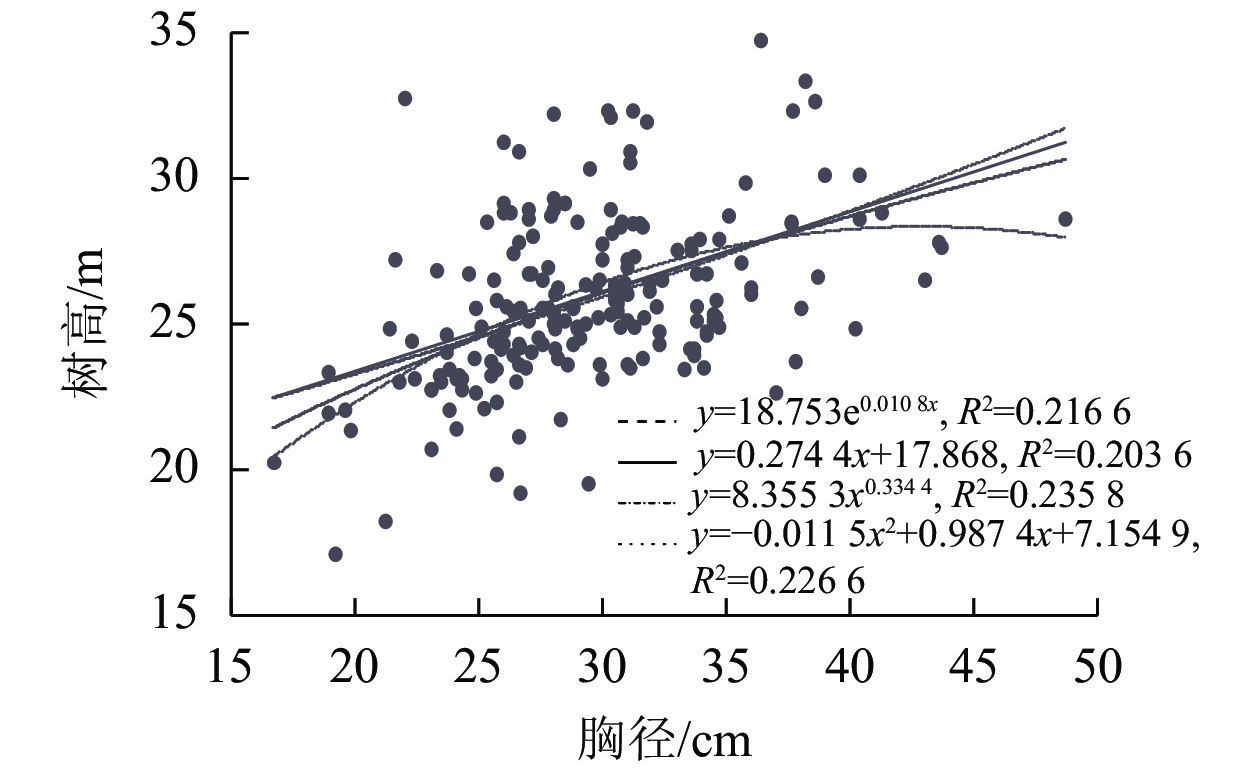
为提高树高测量值的精准度,通过建立试验样地内所有林木的胸径和树高生长模型,利用实测的平均胸径代入生长模型,算出拟合树高进行后续分析(图1)。胸径和树高生长模型的拟合优度分析表明:各拟合模型的精准度从大到小排序为幂函数(R2=0.235 8)、二次函数(R2=0.226 6)、指数函数(R2=0.216 6)、一次线性函数(R2=0.2036),所以幂函数y=8.3553x0.334 4作为树高回归计算的最优函数模型。
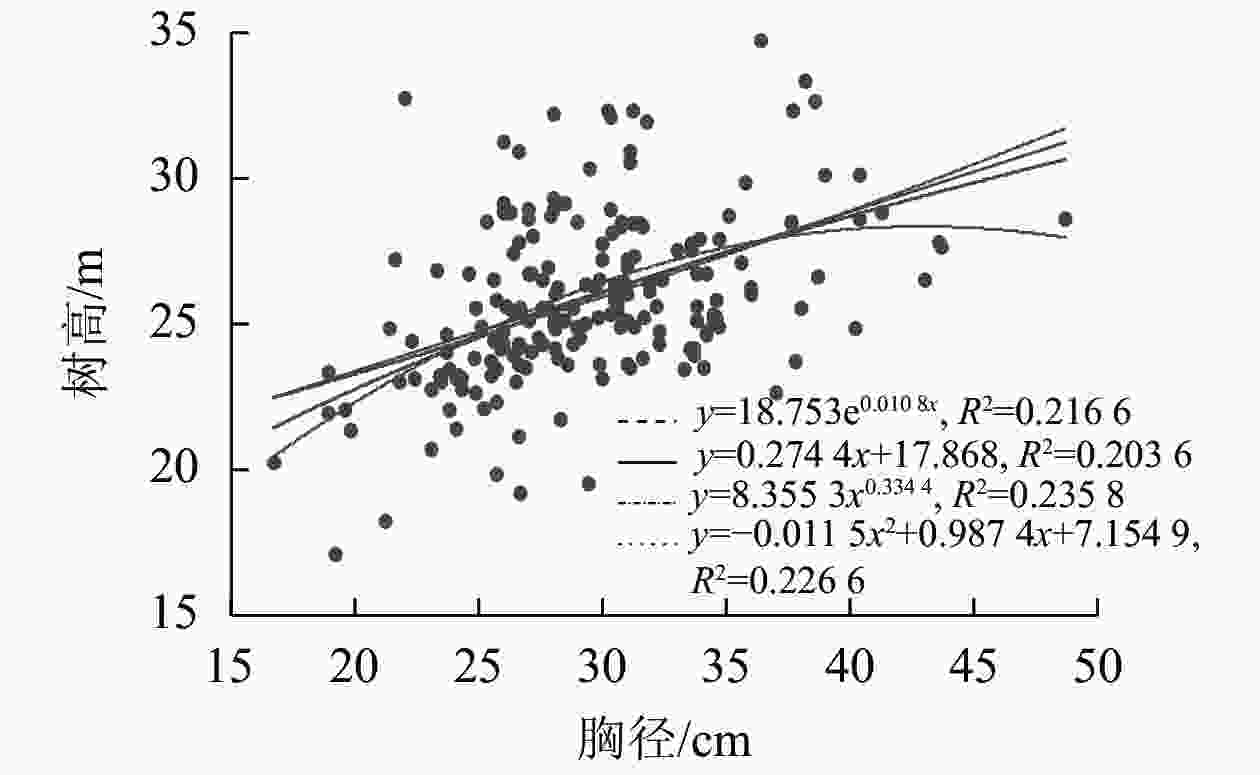
图 1 树高和胸径回归拟合结果
Figure 1. Regression fitting results of tree height and DBH
-
vi=0.000 058 777 042Di1.969 983 1Hi0.896 461 57。其中,Di为第i径阶平均胸径,Hi为第i径阶拟合树高。
-
分别计算样地内各径阶平均单株材积,乘以所对应的径阶株数,即得相对应径阶材积。对样地内各径阶材积累加求和,得到样地林分的总材积,并换算林分的蓄积量(m3·hm−2)。
-
林木材种按其直径大小的区分标准,进阶≥26为大径材,18~24为中径材,10~16为小径材。以径阶大小为区分标准计算各径阶立木出材量,并按照表3所示比例对林分立木各径阶中各材种所占株数进行归并。
表 3 各径级林木材种株数比例
Table 3. Proportion of tree species in different diameter classes
材种 不同径阶株数占比/% 16 18 20 22 24 ≥26 小径材 100 40 0 0 0 0 中径材 0 60 100 100 65 0 大径材 0 0 0 0 35 100 -
杉木规格材的原条用材出材量计算公式为V规(原条)=3.60243758×10−5×D1.94752076×H1.00793769。其中,D为径阶平均胸径(cm);H为径阶平均树高(m)。V规(原条)=V规(小径材)+V规(中径材)+V规(大径材),按比例求得小径材、中径材、大径材的出材量。出材率为出材量与林分蓄积量的比值。
-
使用SPSS软件统计数据和分析差异显著性,使用Excel进行树高胸径拟合,使用Origin 2018制图。
-
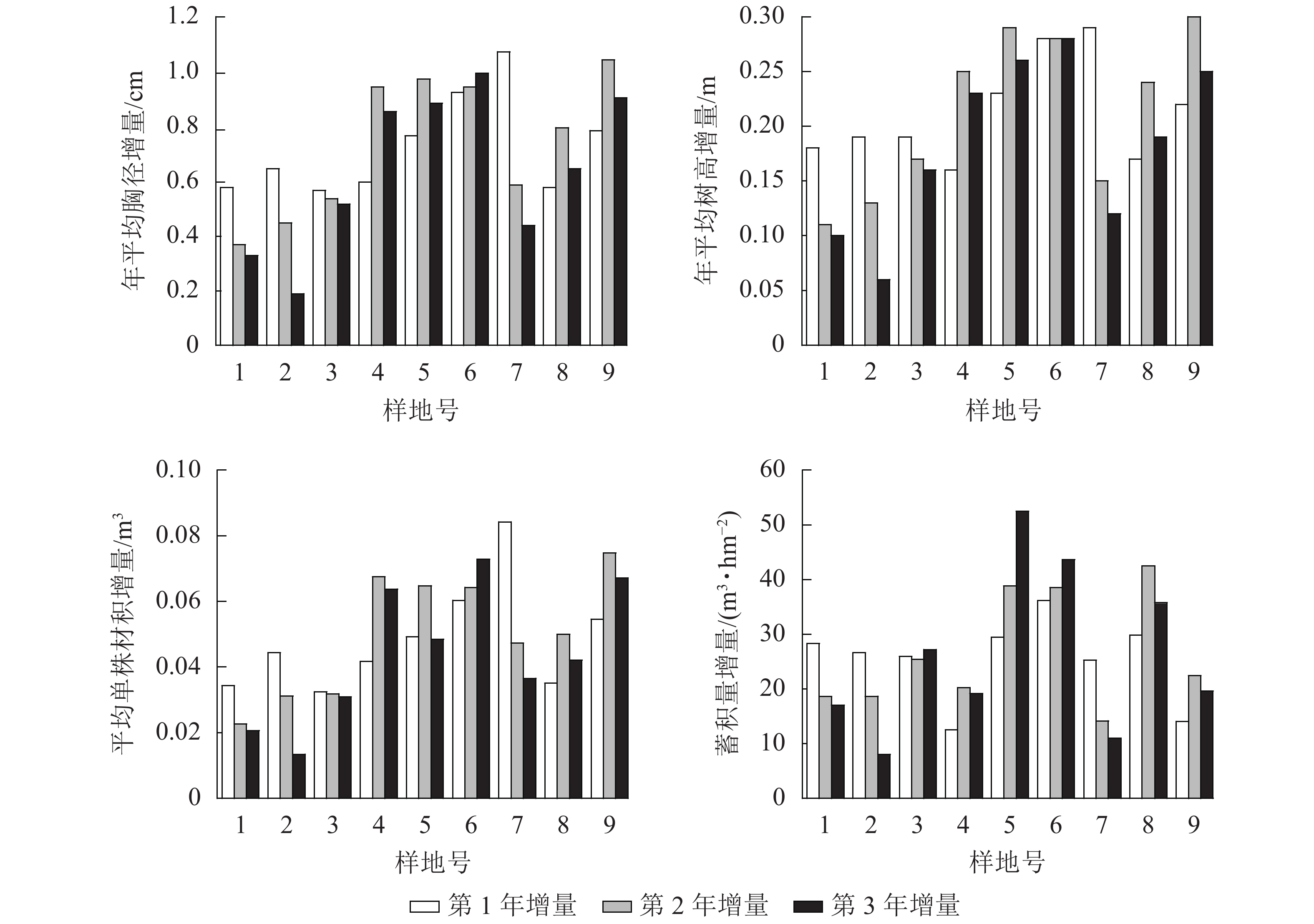
不同间伐和施肥处理下,林分的平均胸径、年平均树高、平均单株材积和蓄积量在3 a的生长中均有提升。处理前林木平均胸径为25.80~33.14 cm,处理后平均胸径为27.43~35.25 cm,平均胸径的增量为1.28~2.88 cm。处理前年平均树高为24.77~26.94 m,处理后年平均树高为25.29~27.50 m,年平均树高的增量为0.38~0.84 m。处理前平均单株材积为0.6351~1.123 4 m3,处理后平均单株材积为0.7302~1.2913 m3,平均单株材积的增量为0.0775~0.1972 m3。处理前蓄积量为270.3812~608.5037 m3·hm−2,处理后蓄积量为326.3937~716.550 0 m3·hm−2,蓄积量的增量为50.3838~120.7630 m3·hm−2。
不同处理下平均胸径、年平均树高、平均单株材积和蓄积量每年增量并不一致(图2),说明间伐抚育和施肥会对成熟林的杉木生长产生影响。就其增量变化趋势来看,并无一定规律,这可能与每个样地的施肥配比有关,对于增量连年增长或先增后减的样地,其氮肥配比较多,较高的速效氮肥施用量能够有效促进杉木的短期速生。氮肥施用量较少而磷肥施用量较多样地,其施肥效应在往后的生长过程中逐步显现,随着施肥效应的逐步减弱,林分密度将会成为制约杉木生长的主要因素。
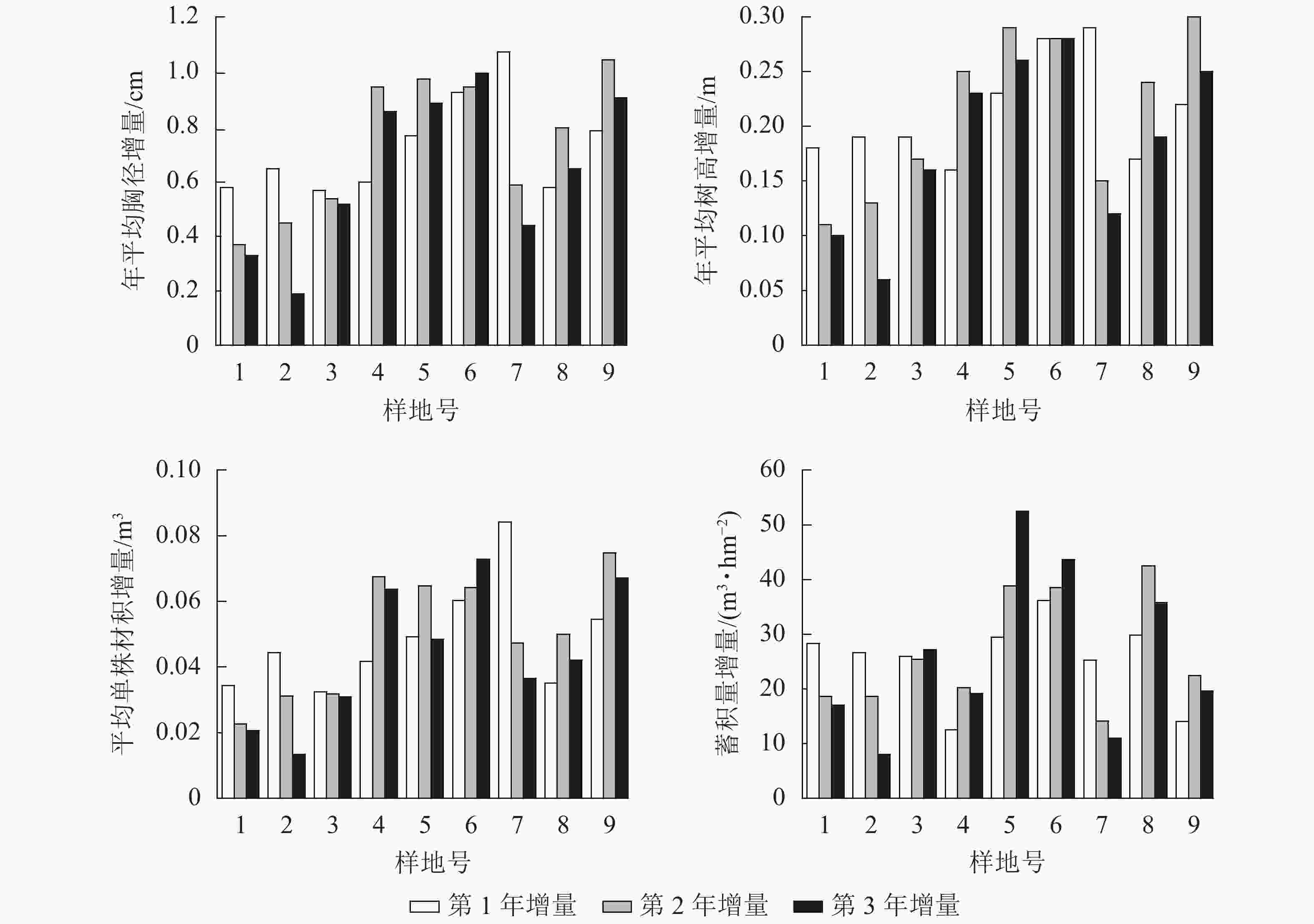
图 2 平均胸径、年平均树高、平均单株材积和蓄积量增量
Figure 2. Increment of average DBH, annual average tree height, average individual volume and stand volume in successive years
由图3显示:样地6平均胸径、年平均树高和平均单株材积的总增量最多,蓄积量总增量仅次于样地5,其处理方式为间伐保留密度600 株·hm−2、氮肥施用量200 g·株−1、磷肥施用量0 g·株−1。样地5的蓄积量总增量最多,其处理方式为间伐保留密度600 株·hm−2、氮肥施用量100 g·株−1、磷肥施用量250 g·株−1。相同的间伐保留密度下,高的氮肥施用配比能短期内增加杉木个体的生长,而氮肥磷肥配合施用能够短期内增加蓄积量的累积。总增量最少的4个样地分别为样地1、2、3、8,其中样地1、3、8的间伐保留密度均为825 株·hm−2,即不进行间伐。结果表明:虽然有施肥因素影响,但保留密度过大仍会限制杉木成熟林胸径增加。此外,林地3 a间的年平均树高总增量基本与平均胸径总增量一致,符合树高-胸径生长模型。平均单株材积总增量最少是样地1,其处理措施为不间伐,不施肥,也反映了合理的间伐保留密度和施肥处理能够有效促进杉木成熟林生长。样地2、4、7、9等的蓄积量总增量最少,其中样地4、7、9的间伐保留密度均为300 株·hm−2,表明间伐强度过大情况下会在3 a内明显影响林分蓄积量的增长。
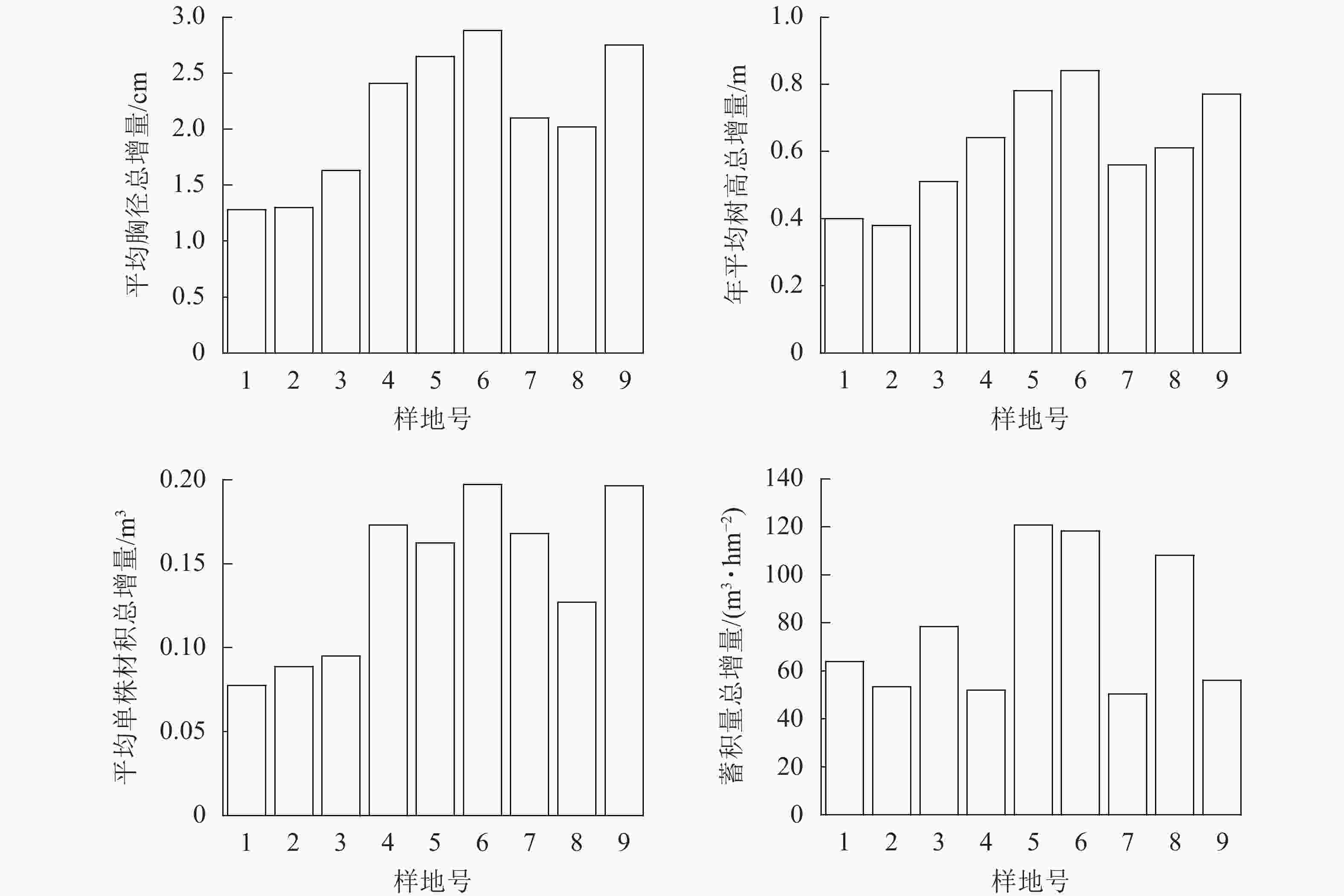
图 3 平均胸径、年平均树高、平均单株材积和蓄积量总增量
Figure 3. Total increment of average DBH, annual average tree height, average individual volume and stand volume
-
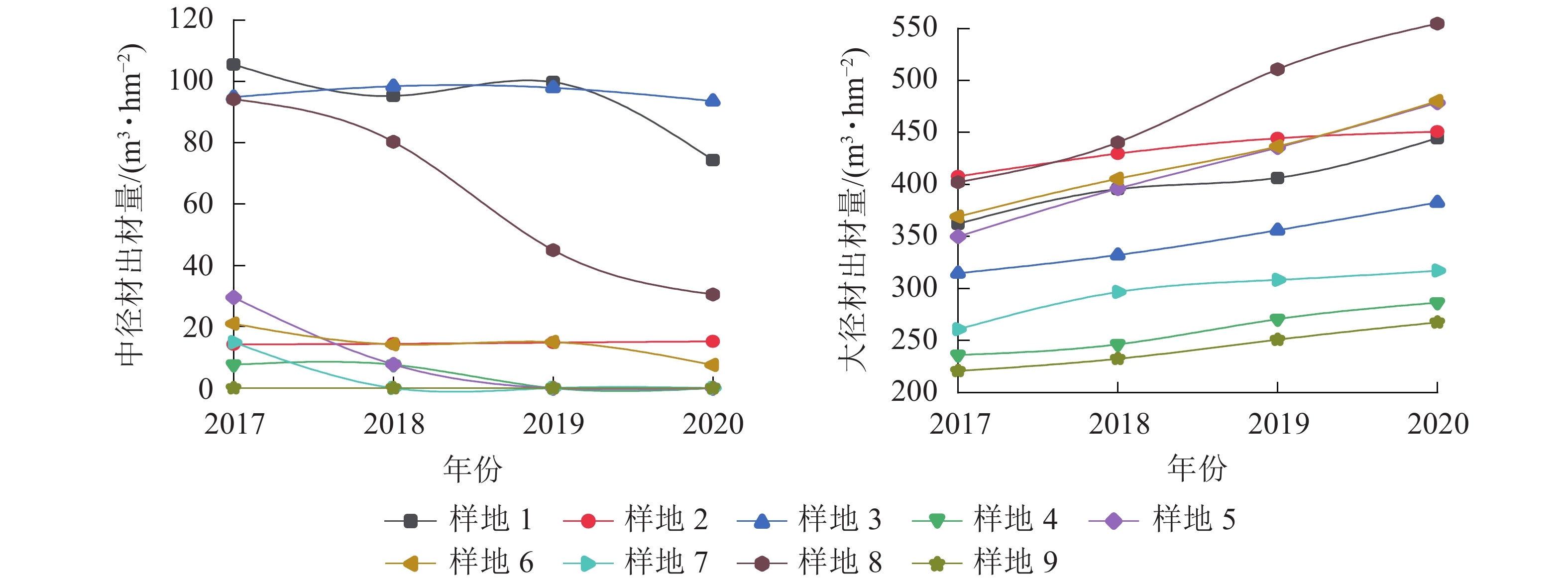
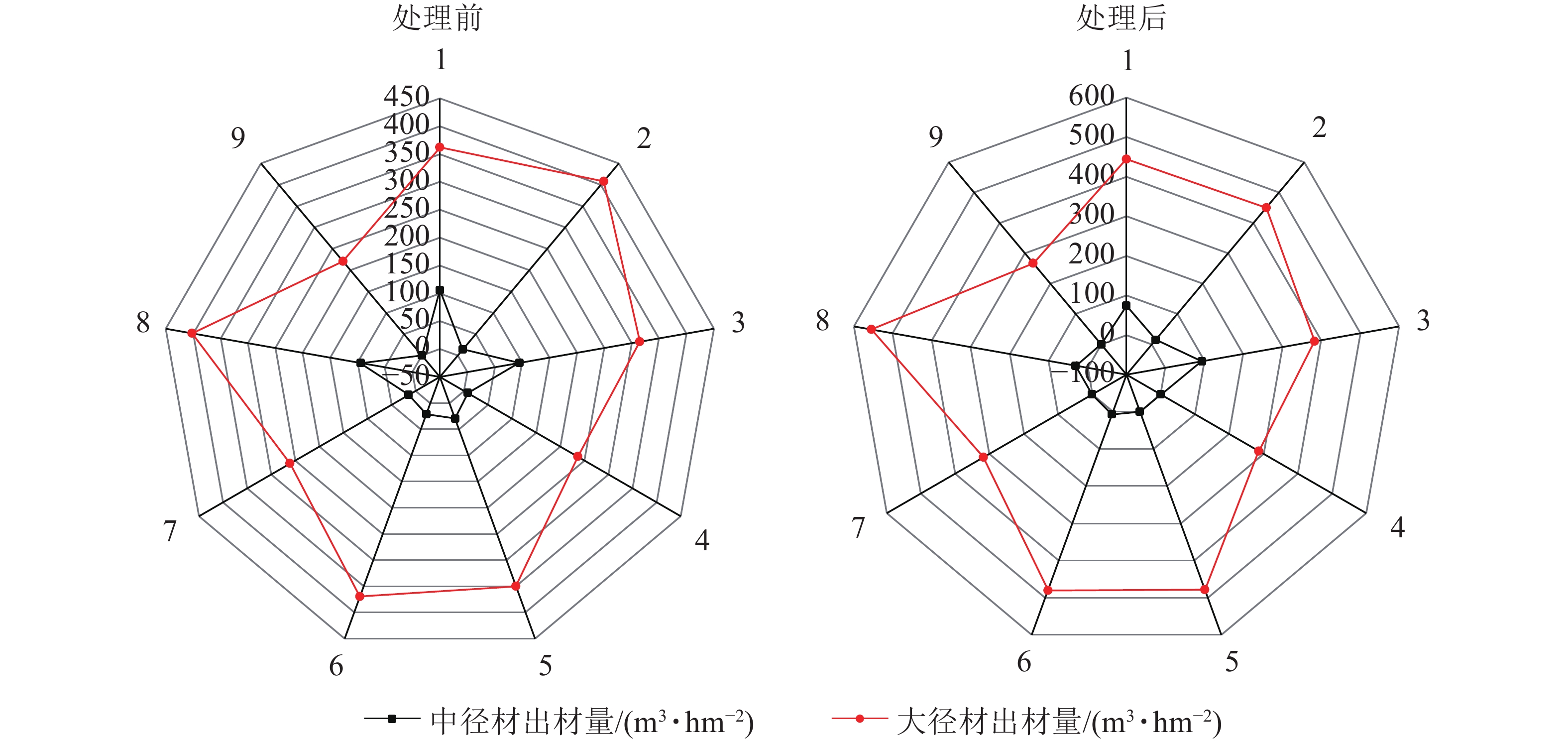
由图4可见:在间伐和施肥处理前后,除样地3有少量小径材,其余均只有中大径材,3 a间中径材出材量整体呈减少趋势,大径材出材量整体呈增长趋势。由图5~6可见:无论在处理前后,大径材出材量和出材率始终大于中径材出材量。大径材出材量在间伐和施肥处理后3 a的增量为42.9265~152.5223 m3·hm−2。样地8,即保留原林分密度825 株·hm−2、氮肥施用量为200 g·株−1、磷肥施用量为250 g·株−1处理下,大径材出材量的增量最大,比处理前增加了152.522 3 m3·hm−2。大径材出材率相比3 a前增加了0.10%~11.32%,其中增加最大的也是样地8,比处理前的大径材出材率增加了11.32%(图6)。
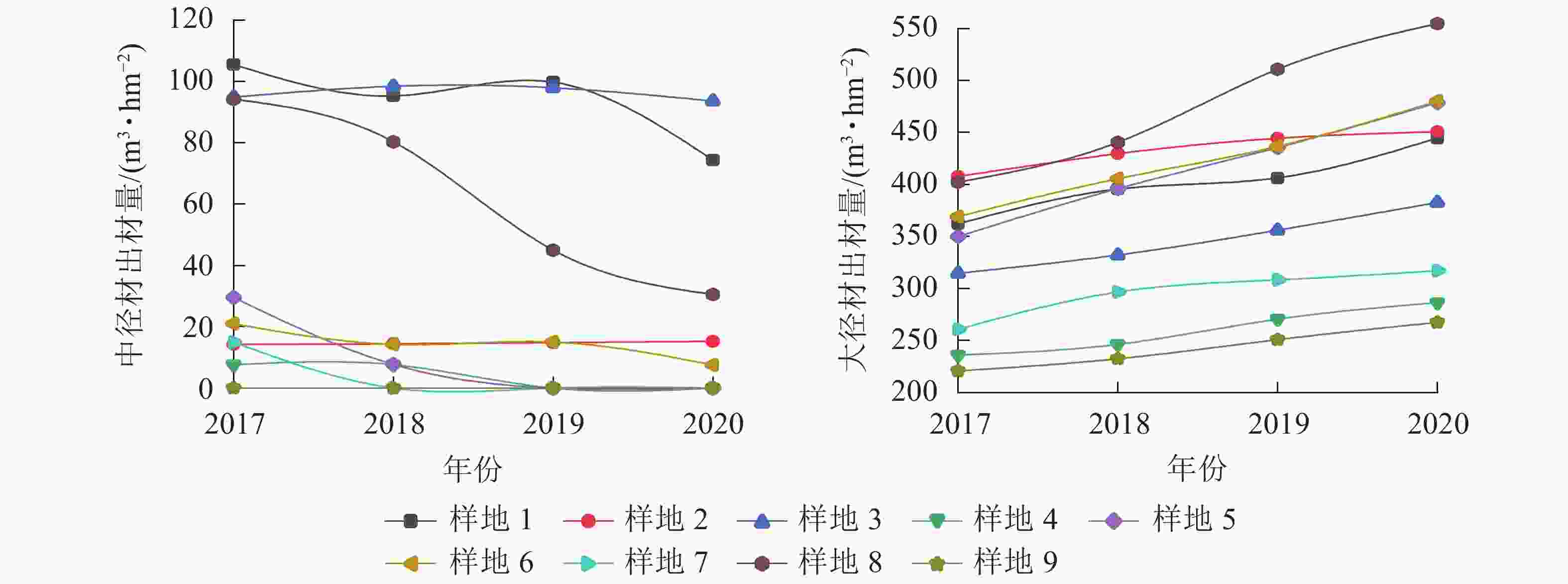
图 4 3 a间中大径材出材量变化情况
Figure 4. Change of output of medium and large diameter timber in the past three years
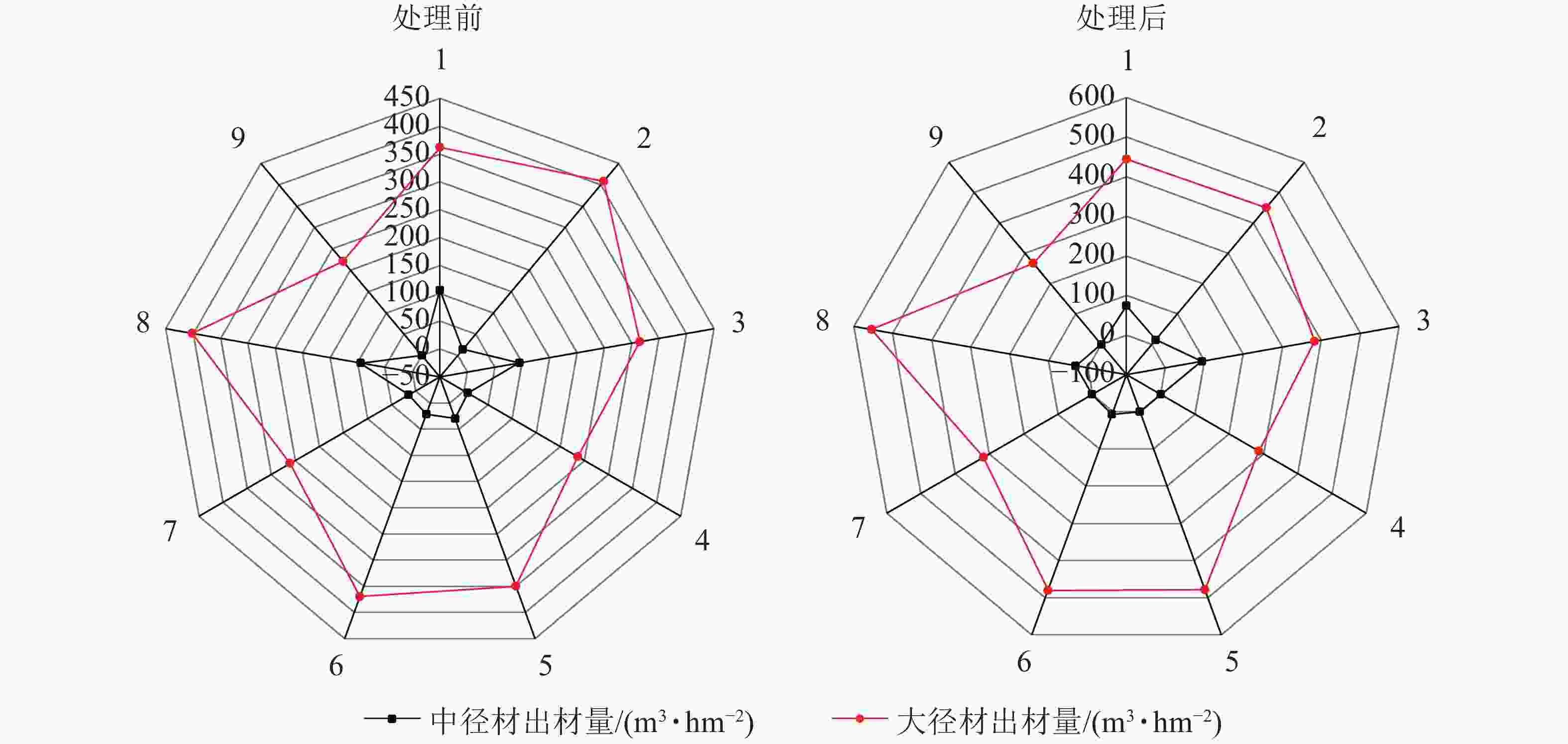
图 5 处理前后中大径材出材量雷达图
Figure 5. Radar chart of output of medium and large diameter wood before and after treatment
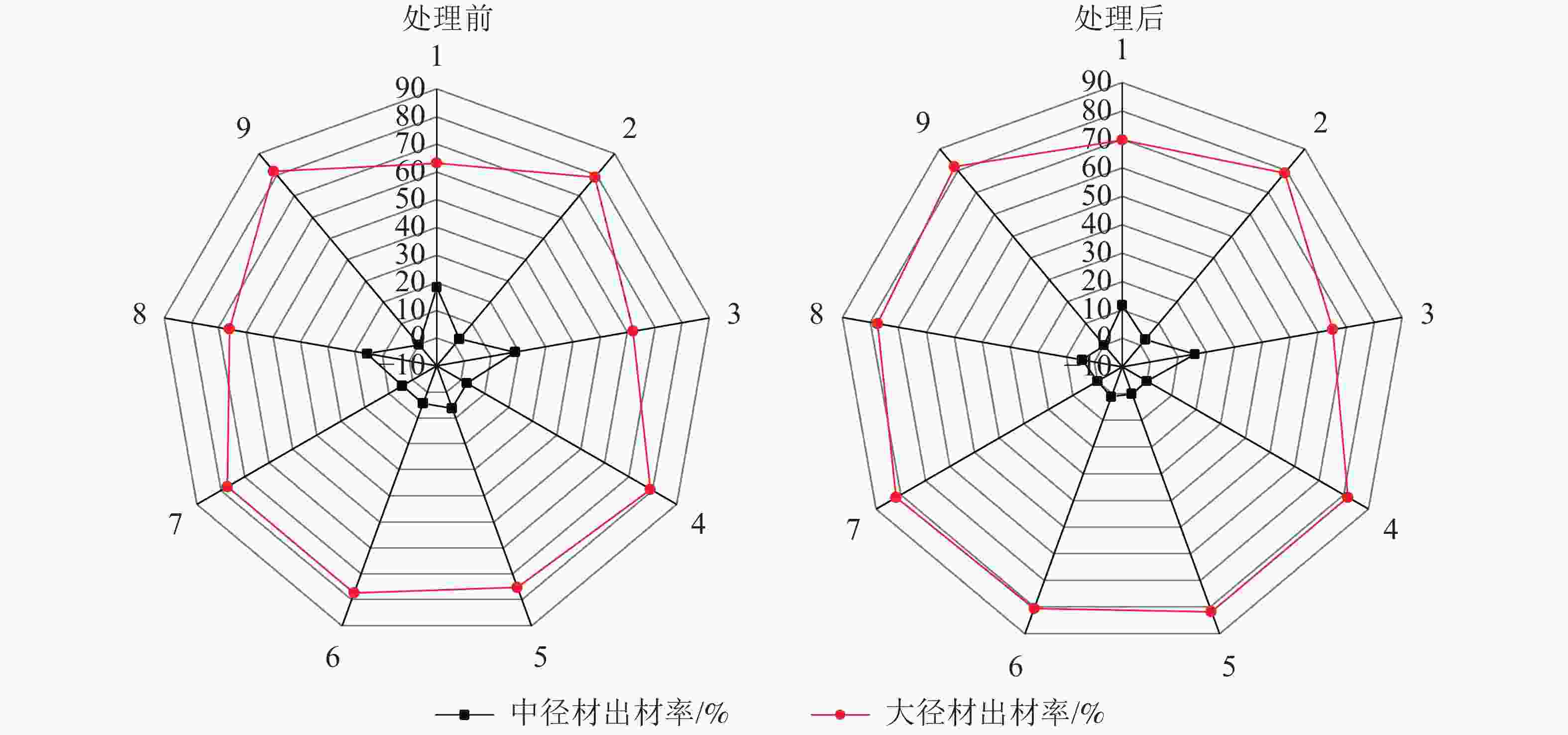
图 6 处理前后中大径材出材率雷达图
Figure 6. Radar chart of output rateof medium and large diameter wood before and after treatment
对于大径材出材率增量的分析(表4)表明:间伐保留密度和磷肥施用量对其具有显著影响(P<0.05),而氮肥施用量对其影响较小,3个因素的主次关系依次为磷肥施用量、间伐保留密度、氮肥施用量,最佳组合为间伐保留密度825株·hm−2(即不间伐)、氮肥施用量200 g·株−1、磷肥施用量250 g·株−1,这与大径材出材量的分析结果一致。因此,立地指数为22,间伐保留密度825株·hm−2的条件,能够使林分维持较高的大径材出材量和出材率。过低的间伐保留密度不再增加大径材,而合适的施肥量会有效提高林分大径材比例。
表 4 各影响因素对大径材出材量(率)的显著性情况
Table 4. Significance of the factors affecting the yield (rate) of large diameter materials
因变量 自变量 P 大径材出材量增量 间伐保留密度 0.019 氮肥量 0.029 磷肥量 0.015 大径材出材率增量 间伐保留密度 0.041 氮肥量 0.439 磷肥量 0.027 说明:P<0.05表示差异显著 -
由于林木之间对养分空间的竞争作用,使间伐成为杉木人工林抚育中必不可少的环节。徐金良等[11]研究发现:间伐会显著增加杉木胸径的增长,高强度的间伐对其胸径增长的影响更加显著。这是由于间伐使林分密度减小,一方面林木之间间隙增大,林木有了足够的生长空间,同时对阳光、水分和养分的竞争减小[12]。另一方面,林内光照增强,促进了林下植被的生长,增加了生物多样性,使人工林生态系统更加稳定,杉木林免受病虫害侵扰[1]。不同间伐强度对杉木中龄林也具有相同作用[13],中度和强度间伐林分的平均胸径显著高于弱度间伐,且平均胸径随着间伐强度的增大而增大。而这种规律对杉木成熟林同样适用,本研究表明:间伐保留的密度过大会限制杉木成熟林林分平均胸径的增加。其他树种也同样受间伐强度影响,对68年生的红松Pinus koraiensis人工林不同强度的抚育间伐研究结果[14]表明:随间伐强度的减弱,林分平均胸径达到较高水平所需年份越来越长。油松Pinus tabuliformis林间伐后2 a的平均胸径随间伐强度增大而升高[15];北美短叶松Pinus banksiana的间伐强度越大,平均胸径越大[16]。对巨尾桉Eucalyptus grandis×E. urophylla的研究[17]表明:施用过量氮肥会抑制桉树生长。本研究也表明:氮肥施用量能促进杉木个体的生长,但基于成本、生态环境保护和施肥效益考虑,杉木大径材培育的氮施用量不宜过高。
林木的平均单株材积与胸径树高息息相关,在杉木成熟林间伐和施肥试验中,与未间伐相比,不同强度的间伐和施肥均促进了平均单株材积的增长。但间伐强度过大,不利于林分蓄积量的增长,反而会使林分蓄积量减少。郑鸣鸣等[13]对杉木中龄林的研究发现:增大间伐强度,间伐3 a后,平均单株材积增长量增大,而林分蓄积量则减小。涂育合等[18]针对杉木成熟林间伐的研究得出:间伐保留密度最小的样地,其蓄积量也最少。李杰茹等[19]对不同抚育间伐强度下秦岭南坡锐齿栎Quercus aliena var. acuteserrata天然次生林研究表明:单株材积增长随间伐强度的增大而增大,但低强度间伐下,其蓄积量最大。SULLIVAN等[20]研究了不同间伐强度对美国黑松Pinus contorta的影响得出:低强度间伐林分蓄积量增长最大,未间伐林分次之,中度和重度间伐则小于未间伐样地。说明间伐强度过大,并不利于林分蓄积量的增长,与本研究结果一致。但尤文忠等[21]认为:强度间伐有利于蒙古栎Quercus mongolica蓄积量的积累,在中龄林阶段应加大间伐强度。而龚固堂等[22]对柏木Cupressus funebris的研究表明:不同间伐强度对蓄积量的影响并不明显。关于间伐能否提高林分总蓄积历来有较大争议,这可能是由于间伐强度、立地条件、树种等各方面的差异,导致结果不同。相同间伐强度下,氮、磷肥共同施用能够增加林分蓄积量。湛红辉[23]研究表明:施肥对马尾松Pinus massoniana幼林及中龄林阶段的蓄积量增加都有影响,但磷肥在其中起关键作用。彭桂永[24]研究表明:杉木中龄林间伐和施肥6 a后,间伐和施肥均对蓄积量的增加起到了极显著的作用。
本研究认为:间伐和施肥导致立地指数为22的杉木成熟林中径材出材量和出材率不断减少,大径材出材量和出材率不断增加。可见立地指数高的林地更适合培育大径材,这可能是由于立地指数高的林地提供的养分更多,林木生长速度更快,林分间的竞争就越大,在成熟时期样地上保留的株数就越少,大径材的出材率越大[25]。郭光智等[26]在南亚热带杉木人工林材种结构长期立地与密度效应的研究中指出:立地指数为18的林分中,中径材比例随年龄增长表现出先增加后减少的趋势,与本研究结果一致,其大径材比例在成熟年龄之后也不断增加。惠刚盈等[27]研究表明:在材种的形成过程中,立地条件是其主要限制因子,在立地指数低的林地中,无论何种间伐保留密度,都难以形成大径材。本研究正交试验的方差分析表明:间伐保留密度、氮肥量、磷肥量对杉木成熟林大径材出材量的增加均具有显著的影响;间伐保留密度和磷肥量对大径材具有显著影响,而氮肥量这个因素对其的影响较小。
本研究得到的最佳间伐密度和施肥配比为间伐保留密度825株·hm−2、氮肥施用量200 g·株−1、磷肥施用量250 g·株−1。在高立地指数(22)杉木成熟林大径材培育过程中,保留适宜的间伐密度,并增施氮肥和适量磷肥,可有效促进大径材出材量和出材率,过低的间伐保留密度在短期内不能有效促进林地大径材出材量和出材率的增加。该结论可能与林地较高的立地指数有关,对于立地指数为18~22的杉木大径材林地,这一结论的一致性还有待继续研究。
Effects of thinning and fertilization on growth and timber structure of mature Chinese fir forest
-
摘要:
目的 分析不同间伐保留密度及不同氮、磷肥施用量对杉木Cunninghamia lanceolata成熟林生长和材种结构的影响,为高效培育大径材杉木提供理论依据。 方法 选择27年生立地指数大于22的杉木人工林,采用正交试验设置9块标准地,间伐保留密度设为300、600和825株·hm−2,氮肥施用量设为0、100、200 g·株−1,磷肥施用量设为0、250、500 g·株−1。 结果 间伐保留密度过大会限制杉木成熟林胸径增加,间伐保留密度过小会降低林分蓄积量,间伐和施肥均能有效促进单株材积增长,较高氮肥施用量能够促进杉木胸径、树高和平均单株材积的生长,氮、磷肥混合施用能够更有效地增加林分蓄积量的积累。间伐保留密度及氮、磷肥施用量3个因素均显著影响杉木成熟林大径材出材量 (P<0.05),间伐保留密度和磷肥施用量对杉木成熟林大径材出材率具有显著影响 (P<0.05),而氮肥施用量的影响较小。在立地指数为22条件下,能够使杉木成熟林大径材出材量和出材率最高的处理为间伐保留密度825株·hm−2、氮肥施用量200 g·株−1、磷肥施用量250 g·株−1。 结论 高立地指数下,杉木成熟林大径材培育过程中,保留适宜的间伐密度,并增施氮肥和磷肥,能有效促进杉木大径材出材率和出材量。图6表4参27 Abstract:Objective This study aims to investigate the effects of different thinning retention density and different proportion of N and P fertilizer application on the growth and timber structure of mature Chinese fir forest. Method The 27 year-old Chinese fir plantation with a site index greater than 22 was selected, and 9 standard plots were set up by orthogonal experimental design. The thinning retention densities were 300, 600 and 825 plants·hm−2. The application rates of N fertilizer were 0, 100 and 200 g·plant−1, and the application rates of P fertilizer were 0, 250 and 500 g·plant−1. Result Excessive thinning retention density limited the diameter at breast height (DBH) increase of mature Chinese fir. Too small thinning retention density reduced the stand stock. Thinning and fertilization could effectively promote the growth of individual tree volume. Higher N application rate could promote the growth of DBH, tree height and average individual tree volume of Chinese fir, and the mixed application of N and P could increase the accumulation of stand volume more effectively. The thinning retention density, the application rate of N and P fertilizer all significantly affected the yield of large-diameter timber of mature Chinese fir forest (P<0.05). The thinning retention density and the application rate of P fertilizer had a significant impact on the yield of large-diameter timber of mature Chinese fir forest (P<0.05), while the application rate of N fertilizer had little impact. Under the site index of 22, the combination that produced the maximum yield and outturn rate of large-diameter timber of mature Chinese fir forest was thinning retention density of 825 plants·hm−2, N fertilizer application rate of 200 g·plant−1 and P fertilizer application rate of 250 g·plant−1. Conclusion Under the condition of high site index, the outturn and yield of large-diameter timber of Chinese fir could be effectively promoted by keeping appropriate thinning density and increasing N and P fertilizer application in the cultivation process of mature Chinese fir forest. [Ch, 6 fig. 4 tab. 27 ref.] -
Key words:
- Chinese fir /
- thinning /
- fertilization /
- large diameter timber /
- timber structure
-
图 2 平均胸径、年平均树高、平均单株材积和蓄积量增量
Figure 2 Increment of average DBH, annual average tree height, average individual volume and stand volume in successive years
图 3 平均胸径、年平均树高、平均单株材积和蓄积量总增量
Figure 3 Total increment of average DBH, annual average tree height, average individual volume and stand volume
图 4 3 a间中大径材出材量变化情况
Figure 4 Change of output of medium and large diameter timber in the past three years
图 5 处理前后中大径材出材量雷达图
Figure 5 Radar chart of output of medium and large diameter wood before and after treatment
图 6 处理前后中大径材出材率雷达图
Figure 6 Radar chart of output rateof medium and large diameter wood before and after treatment
表 1 试验地土壤理化性质
Table 1. Physical properties of soil in the experimental site
土层/cm 最大持
水量/mm毛管持
水量/mm最小持
水量/mm非毛管孔
隙度/%毛管孔
隙度/%总孔隙
度/%土壤密度/
(g·cm−3)氮/
(g·kg−1)磷/
(g·kg−1)0~10 71.53±0.00 66.33±0.00 64.93±0.00 5.20±0.00 66.33±0.00 71.53±0.00 2.66±0.15 24.11±5.01 1.60±0.38 10~20 72.13±0.00 63.53±0.00 61.47±0.00 8.60±0.00 63.53±0.00 72.13±0.00 2.68±0.10 26.04±4.05 1.01±0.30 20~40 135.80±0.01 123.00±0.01 119.60±0.01 6.40±0.00 61.50±0.00 67.90±0.01 2.87±0.14 26.50±5.37 0.70±0.16  下载: 导出CSV
下载: 导出CSV
表 2 各样地间伐施肥处理情况
Table 2. Various thinning and fertilization treatments
样地号 间伐保留密度/
(株·hm−2)氮肥施用量/
(g·株−1)磷肥施用量/
(g·株−1)样地号 间伐保留密度/
(株·hm−2)氮肥施用量/
(g·株−1)磷肥施用量/
(g·株−1)1 825 0 0 6 600 200 0 2 600 0 500 7 300 0 250 3 825 100 500 8 825 200 250 4 300 100 0 9 300 200 500 5 600 100 250
下载: 导出CSV
表 3 各径级林木材种株数比例
Table 3. Proportion of tree species in different diameter classes
材种 不同径阶株数占比/% 16 18 20 22 24 ≥26 小径材 100 40 0 0 0 0 中径材 0 60 100 100 65 0 大径材 0 0 0 0 35 100
下载: 导出CSV
表 4 各影响因素对大径材出材量(率)的显著性情况
Table 4. Significance of the factors affecting the yield (rate) of large diameter materials
因变量 自变量 P 大径材出材量增量 间伐保留密度 0.019 氮肥量 0.029 磷肥量 0.015 大径材出材率增量 间伐保留密度 0.041 氮肥量 0.439 磷肥量 0.027 说明:P<0.05表示差异显著
下载: 导出CSV
-
[1] 徐雪蕾, 孙玉军, 周华, 等. 间伐强度对杉木人工林林下植被和土壤性质的影响[J]. 林业科学, 2019, 55(3): 1 − 12. XU Xuelei, SUN Yujun, ZHOU Hua, et al. Effects of thinning intensity on understory growth and soil properties in Chinese fir plantation [J]. Sci Silv Sin, 2019, 55(3): 1 − 12. [2] 国家林业局. 第八次全国森林资源清查结果[EB/OL]. 2016-01-15 [2021-03-15]. http: //www. forestry. gov. cn/. State Forestry Administration. The 8 th National Forest Resources Inventory[EB/OL]. 2016-01-15 [2021-03-15]. http://www. forestry. gov. cn/. [3] 赵隽宇, 张泽尧, 覃祚玉, 等. 长期连作下广西杉木人工林主产区土壤肥力演变特征与评价[J]. 中南林业科技大学学报, 2020, 40(9): 124 − 131. ZHAO Junyu, ZHANG Zeyao, QIN Zuoyu, et al. Evolution characteristics and evaluation of soil fertility in main producing areas of Cunninghamia lanceolata plantation in Guangxi under long-term cropping [J]. J Cent South Univ For Technol, 2020, 40(9): 124 − 131. [4] WU Chuping, JIANG Bo, YUAN Weigao, et al. On the management of large-diameter trees in China’s forests[J/OL]. Forests, 2020, 11(1): 111[2021-02-12]. doi: 10.3390/f11010111. [5] 蔡年辉, 李根前. 人工林大径材定向培育研究现状[J]. 安徽农业科学, 2009, 37(31): 15518 − 15519. CAI Nianhui, LI Genqian. Research status of directional cultivation of large diameter timber in plantation [J]. J Anhui Agric Sci, 2009, 37(31): 15518 − 15519. [6] RUSSELL M B, BURKHART H E, AMATEIS R L, et al. Regional locale and its influence on the prediction of Loblolly pine diameter distributions [J]. Southern J Appl For, 2012, 36(4): 198 − 203. [7] SUN Honggang, ZHANG Jianguo, DUAN Aiguo, et al. Effect of pre-commercial thinning on diameter class distribution and total merchantable volume growth in Chinese fir stands in southern China [J]. Afr J Agric Res, 2012, 7(21): 3158 − 3165. [8] 晏姝, 王润辉, 邓厚银, 等. 南岭山区杉木大径材成材影响因子研究[J]. 华南农业大学学报, 2021, 42(2): 80 − 89. YAN Shu, WANG Runhui, DENG Houyin, et al. Study on impact factors of large-diameter wood formation of Cunninghamia lanceolata in Nanling Mountains [J]. J South China Agric Univ, 2021, 42(2): 80 − 89. [9] 邓伦秀. 杉木人工林林分密度效应及材种结构规律研究[D]. 北京: 中国林业科学研究院, 2010. DENG Lunxiu. Study on Stand Density Effect and Timber Species Structure of Cunninghamia lanceolata Plantations[D]. Beijing: Chinese Academy of Forestry, 2010. [10] 相聪伟. 杉木人工林生长及材种结构规律研究[D]. 北京: 中国林业科学研究院, 2013. XIANG Congwei. Study on Regulation of Stand Growth and Timber Assortment Structure in Cunninghamia lanceolata Plantation[D]. Beijing: Chinese Academy of Forestry, 2013. [11] 徐金良, 毛玉明, 郑成忠, 等. 抚育间伐对杉木人工林生长及出材量的影响[J]. 林业科学研究, 2014, 27(1): 99 − 107. XU Jinliang, MAO Yuming, ZHENG Chengzhong, et al. Effect of thinning on growth and timber outturn in Cunninghamia lanceolata plantation [J]. For Res, 2014, 27(1): 99 − 107. [12] CRECENTE-CAMPO F, POMMERENING A, RODRIGUEZ-SOALLEIRO R. Impacts of thinning on structure, growth and risk of crown fire in a Pinus sylvestris L. plantation in northern Spain [J]. For Ecol Manage, 2009, 257(9): 1945 − 1954. [13] 郑鸣鸣, 任正标, 王友良, 等. 间伐强度对杉木中龄林生长和结构的影响[J]. 森林与环境学报, 2020, 40(4): 369 − 376. ZHENG Mingming, REN Zhengbiao, WANG Youliang, et al. Effect of thinning intensity on the growth and structure of a middle-aged Chinese fir forest [J]. J For Environ, 2020, 40(4): 369 − 376. [14] 孙白冰, 陈晓波, 张启昌, 等. 我国红松大径级用材林近自然培育探讨[J]. 世界林业研究, 2021, 34(1): 84 − 89. SUN Baibing, CHEN Xiaobo, ZHANG Qichang, et al. Discussions on close-to-nature silviculture of Pinus koraiensis stand for large-diameter timber in China [J]. World For Res, 2021, 34(1): 84 − 89. [15] 马履一, 李春义, 王希群, 等. 不同强度间伐对北京山区油松生长及其林下植物多样性的影响[J]. 林业科学, 2007, 43(5): 1 − 9. MA Lüyi, LI Chunyi, WANG Xiqun, et al. Effects of thinning on the growth and the diversity of undergrowth of Pinus tabulaeformis plantation in Beijing mountainous areas [J]. Sci Silv Sin, 2007, 43(5): 1 − 9. [16] 童雀菊, 张述垠. 抚育间伐对北美短叶松的生长及材质的影响(英文)[J]. 南京林业大学学报(自然科学版), 2005, 29(6): 73 − 76. TONG Queju, ZHANG Shuyin. Effects of precommercialthinning on tree growth and stem quality in Jack pine stands [J]. J Nanjing For Univ Nat Sci Ed, 2005, 29(6): 73 − 76. [17] 胡厚臻. 配方施肥下巨尾桉的生长生理及土壤肥力特征研究[D]. 南宁: 广西大学, 2016. HU Houzhen. Study on the Growth Physiology and the Soil Fertility Character of Eucalyptus grandis × E. urophylla under the Formulated Fertilizer Application [D]. Nanning: Guangxi University, 2016. [18] 涂育合, 叶功富, 林武星, 等. 杉木大径材定向培育的适宜经营密度[J]. 浙江林学院学报, 2005, 22(5): 530 − 534. TU Yuhe, YE Gongfu, LIN Wuxing, et al. Suitable management density for directional cultivation of large diameter timber of Cunninghamia lanceolata [J]. J Zhejiang For Coll, 2005, 22(5): 530 − 534. [19] 李杰茹, 党坤良, 王先初, 等. 抚育间伐对秦岭南坡锐齿栎天然次生林生长的影响[J]. 西北林学院学报, 2021, 36(1): 62 − 68. LI Jieru, DANG Kunliang, WANG Xianchu, et al. Effects of thinning on the growth of Quercus aliena var. acuteserrata natural secondary forest on the southern slope of Qinling Mountains [J]. J Northwest For Univ, 2021, 36(1): 62 − 68. [20] SULLIVAN T P, SULLIVAN D S. Acceleration of old-growth structural attributes in lodgepole pine forest: tree growth and stand structure 25 years after thinning [J]. For Ecol Manage, 2016, 365: 96 − 106. [21] 尤文忠, 赵刚, 张慧东, 等. 抚育间伐对蒙古栎次生林生长的影响[J]. 生态学报, 2015, 35(1): 56 − 64. YOU Wenzhong, ZHAO Gang, ZHANG Huidong, et al. Effects of thinning on growth of Mongolian oak (Quercus mongolica) secondary forests [J]. Acta Ecol Sin, 2015, 35(1): 56 − 64. [22] 龚固堂, 牛牧, 慕长龙, 等. 间伐强度对柏木人工林生长及林下植物的影响[J]. 林业科学, 2015, 51(4): 8 − 15. GONG Gutang, NIU Mu, MU Changlong, et al. Impacts of different thinning intensities on growth of Cupressus funebris plantation and understory plants [J]. Sci Silv Sin, 2015, 51(4): 8 − 15. [23] 谌红辉. 广西马尾松人工林定向培育关键技术研究[D]. 南京: 南京林业大学, 2010. CHEN Honghui. A Study on Key Techniques for Directive Cultivation of Masson Pine Plantations in Guangxi Province[D]. Nanjing: Nanjing Forestry University, 2010. [24] 彭桂永. 杉木中龄林间伐后不同保留密度和施肥试验研究[J]. 安徽农学通报, 2016, 22(12): 94 − 95, 117. PENG Guiyong. Experimental study on different retention density and fertilization of middle-aged Chinese fir forest after thinning [J]. Anhui Agric Sci Bull, 2016, 22(12): 94 − 95, 117. [25] 相聪伟, 张建国, 段爱国, 等. 杉木人工林材种结构的立地及密度效应研究[J]. 林业科学研究, 2015, 28(5): 654 − 659. XING Congwei, ZHANG Jianguo, DUAN Aiguo, et al. Effects of site quality and planting density on wood assortment rate in Chinese fir plantation [J]. For Res, 2015, 28(5): 654 − 659. [26] 郭光智, 段爱国, 张建国, 等. 南亚热带杉木人工林材种结构长期立地与密度效应[J]. 林业科学研究, 2020, 33(1): 35 − 43. GUO Guangzhi, DUAN Aiguo, ZHANG Jianguo, et al. Long-term effects of site and density on timber assortment structure of Chinese fir plantations in south subtropical area, China [J]. For Res, 2020, 33(1): 35 − 43. [27] 惠刚盈, 胡艳波, 罗云伍, 等. 杉木中大径材成材机理的研究[J]. 林业科学研究, 2000, 13(2): 177 − 181. HUI Gangying, HU Yanbo, LUO Yunwu, et al. Study on the mechanism of producing high quality wood of Chinese fir [J]. For Res, 2000, 13(2): 177 − 181. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210226
 点击查看大图
点击查看大图
计量
- 文章访问数: 2324
- HTML全文浏览量: 454
- PDF下载量: 59
- 被引次数: 0

